1. Introduction
With the rapid development of aquaculture and marine fishing industries, the demand for fishmeal as a high-quality protein source has continued to rise, and its insufficient supply has caused prices to soar [
1]. Plant protein materials, due to their wide availability and low cost, are commonly used as alternatives to fishmeal in aquafeeds. However, anti-nutritional factors and amino acid imbalances in plant protein materials can affect the growth and health of aquatic animals to varying degrees [
2,
3]. When fishmeal is largely replaced by plant protein, functional feed additives (such as organic acids, nucleotides, and probiotics) are typically added to mitigate the negative effects of the alternative protein sources, thereby enhancing fish growth performance, boosting immunity and disease resistance, and improving gut health [
4,
5].
Sodium butyrate (SB), the sodium salt of the short-chain volatile fatty acid butyric acid, is widely used in poultry and aquaculture feed due to its high stability, rapid absorption, and environmental friendliness [
6,
7]. However, different forms of SB show significant differences in bioavailability and efficacy [
8]. Raw SB powder, with its strong odor and short shelf life, is rapidly absorbed in the stomach, making it difficult to exert sustained effects in the intestine [
9]. Encapsulated SB (ESB), encapsulated with lipids or polymers, forms protective and controlled-release layers. It is released gradually in the stomach and intestinal regions [
10] and is targeted to the cecum and colon [
11], enhancing intestinal digestion and absorption and reducing inflammation [
12,
13].
SB is an important energy source for intestinal epithelial cells, promoting cell growth and metabolism [
14], and can reduce intestinal pH to suppress harmful bacteria and maintain gut homeostasis [
15]. Studies have demonstrated that dietary SB can enhance the growth performance and digestive enzyme activities of juvenile thin-lipped mullet (
Liza ramada) [
16]. Moreover, dietary SB has been proven to enhance intestinal absorption and immunity in various fish species, including Nile tilapia (
Oreochromis niloticus) [
17], grass carp (
Ctenopharyngodon idella) [
18], turbot (
Scophthalmus maximusa) [
19], and rice field eel (
Monopterus albus) [
20]. Interestingly, Jesus et al. [
21] observed that the addition of encapsulated SB at 0.5% in either buffer or oil improved the yield and biomass gain compared with the unprotected forms during sexual reversion of Nile tilapia. Additionally, Zhou et al. [
22] found that different SB forms improved
C. idella health by up-regulating
myd88 and
tlr22 gene expressions, enriching beneficial gut bacteria, and reducing lipid accumulation, despite showing no significant effects on growth. These findings highlight the need for further investigation into the optimal dietary supplementation levels of different forms of SB (raw powder or encapsulated), as well as their impacts on gut antioxidant status, microbiota composition, and immune function.
The largemouth bass (
Micropterus salmoides), a carnivorous fish, preys on small fish and insects. It is highly regarded in the market for its firm flesh, delicious taste, few bones, and high nutritional value [
23]. With its rapid growth and adaptability to intensive farming, it has a robust market demand. Largemouth bass has a high demand for dietary protein and is more sensitive to adverse intestinal effects caused by plant-based feed, making it a key species for enhancing feed efficiency and health management and an ideal model for studying carnivorous fish intestinal physiology [
24]. Research showed that dietary supplementation with SB in feed could alleviate the negative influence of high-fat diets on fish growth, intestinal microbiota homeostasis, and liver health in largemouth bass [
23,
24]. However, the appropriate inclusion level of encapsulated SB in largemouth bass feed, along with comparisons to raw powder SB at equivalent doses, remains an area for further investigation. This study investigates how different doses of encapsulated SB and raw powder SB affect the growth, serum biochemistry, gut structure, antioxidant capacity, immunity, and gut microbiota of largemouth bass, aiming to guide the optimal use of encapsulated SB in feed.
2. Materials and Methods
2.1. Diet Formulation
Largemouth bass (
M. salmoides) with an initial weight of 29.75 ± 0.25 g were used in this study. Sodium butyrate (SB) as the feed additive was obtained from Hunan Perfly Biotech Co., Ltd. (Hunan, China). Five experimental diets were formulated: a control diet (SB0, basal diet containing 25% fish meal by weight with no SB) and four treatment diets supplemented with 1000 mg/kg (ESB1), 1500 mg/kg (ESB2), and 2000 mg/kg (ESB3) of encapsulated SB, or 2000 mg/kg of raw powder SB (RSB). The addition concentration of SB in the feed referenced effective levels reported in published studies on grass carp (
C. idella) [
22], yellow drum (
Nibea albiflora) [
13], and Nile tilapia [
21], with a gradient design covering 1000–2000 mg/kg to identify optimal concentrations for largemouth bass. The main ingredients and nutritional compositions of the five diets are listed in
Table 1.
The feed ingredients were finely ground through a 60-mesh sieve, uniformly mixed according to the formulated ratios, and extruded into strip-shaped pellets using an aquafeed extruder (Jiangsu Zhengchang Grain and Feed Machinery Co., Ltd., SZLH200, Liyang, China) at a temperature of 50 °C. The extruded feed was dried to a moisture content of less than 10% and then formed into approximately 3 mm diameter particles using a feed pellet mill (Jiangsu Zhengchang Grain and Feed Machinery Co., Ltd., 1208 Pellet Mill, Liyang, China) for low-temperature storage until use.
2.2. Experimental Animals and Management
After two weeks of acclimation, 600 largemouth bass with an initial weight of 29.75 ± 0.25 g were evenly distributed into five groups (n = 120 fish per group). Each group was allocated three net cages (2 m × 2 m × 1.6 m; total 15 cages) with 40 fish per cage. The cages were suspended in an aerated pond. The 49-day feeding trial involved daily feeding at 3% of body weight, distributed twice daily (07:00 and 17:00) until apparent satiation (1–1.5 h per feeding). Weekly, four fish per group were randomly weighed to adjust feeding rations. Water quality parameters were maintained within the following ranges: dissolved oxygen at 5–7 mg/L, water temperature at 16–32 °C, pH at 7.0–8.5, and pond water transparency at 30–40 cm.
2.3. Sample Collection and Ethics Statement
At the end of the 49-day rearing period, the largemouth bass were subjected to a 24 h fasting period. Subsequently, 20 fish were randomly sampled from each net cage (60 fish per group) for experimentation. The fish were anesthetized with an MS-222 solution (50 mg/L, Changsha Shanghe Biotechnology Co., Ltd., Changsha, China) for 5 min. All experiments complied with the Guidelines for the Care and Use of Laboratory Animals in China and adhered to local animal welfare regulations and institutional guidelines.
Surface moisture was gently removed from the 60 anesthetized fish per group using sterile gauze before body weight measurement. Blood was collected from the caudal vein of 30 randomly selected fish per group. The blood samples were left to stand at 4 °C for 24 h and then centrifuged at 3000× g for 15 min to collect the serum, which was stored at −80 °C for subsequent serum biochemical analysis. The intestines of these 30 fish were quickly dissected on ice, and tissue samples from the proximal (3 cm posterior to the stomach), middle, and distal (3 cm anterior to the anus) segments of the intestine were collected. Tissue samples from six fish were fixed in 4% paraformaldehyde at 4 °C for histological section preparation, while samples from the remaining 24 fish per experimental group (8 fish from each of the 3 tanks) were snap-frozen in liquid nitrogen for 12 h and then stored at −80 °C for total RNA extraction and antioxidant enzyme activity assays.
Finally, from the remaining 30 anesthetized fish in each group, 18 fish per experimental group (6 fish from each of the 3 tanks) were selected for gut content collection. The entire intestinal contents of these fish per experimental group (4 fish from each of the 3 tanks) were collected aseptically and stored in sterile vials. The samples were snap-frozen in liquid nitrogen for 12 h and then stored at −80 °C for subsequent gut microbiota sequencing. The remaining 12 fish per experimental group (4 fish from each of the 3 tanks) were stored at −20 °C for whole-body composition analysis (moisture, crude fat, crude protein, and crude ash).
2.4. Survival and Growth Performance
The survival and growth parameters measured in this study included initial body weight (IBW), final body weight (FBW), weight gain rate (WGR), specific growth rate (SGR), Feed coefficient (FC), feed intake (FI), viscera somatic index (VSI), and hepatosomatic index (HSI).
2.5. Proximate Composition Analysis
The crude protein content was determined using the Kjeldahl method (N × 6.25). Crude fat content was measured by Soxhlet extraction. Moisture content was determined by drying samples to constant weight in an oven at 105 °C. Ash content was analyzed by incinerating samples to constant weight in a muffle furnace at 550 °C. All procedures followed the methods described by Su et al. [
25].
2.6. Histological Observation
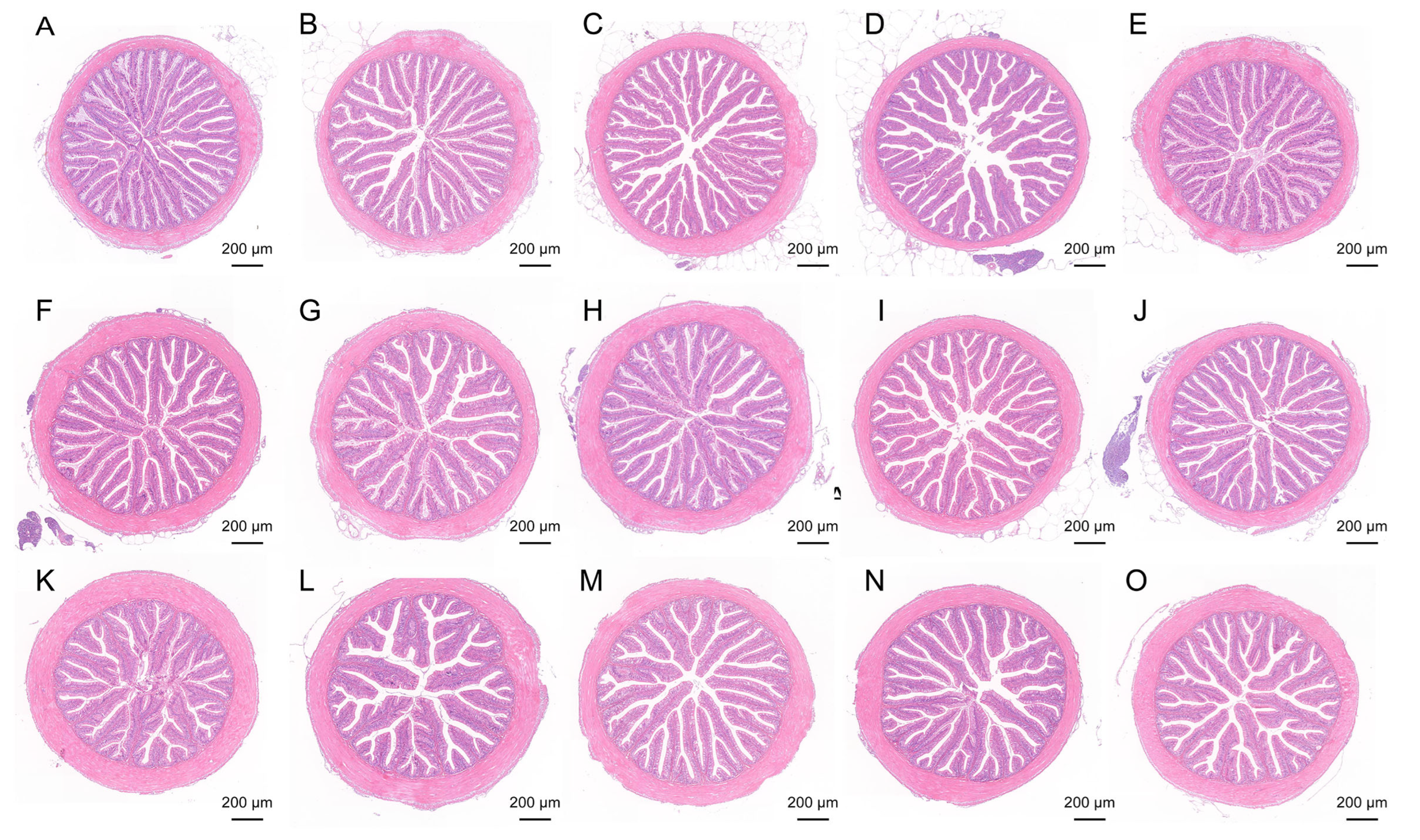
Proximal, middle, and distal intestinal samples from each group (n = 3) were dehydrated through a graded ethanol series and embedded in paraffin. Transverse sections approximately 1 cm in length were cut into 5 μm thick slices and stained with hematoxylin and eosin (H&E) following the method of Su et al. [
25]. H&E staining was performed and scanned by Wuhan Sevicebio Technology Co., Ltd. (Wuhan, China). Sections were scanned using a NanoZoomer S60 (Hamamatsu Photonics, Hamamatsu, Japan), and measurements of villus length, villus width, and muscle layer thickness were taken using Image-Pro Plus 6.0 software.
2.7. Serum Biochemical Assays
Blood samples collected from the caudal vein were left to clot at 4 °C, then centrifuged at 3000×
g for 10 min to obtain serum, which was stored at −80 °C. Serum samples (n = 3) were analyzed for alanine aminotransferase (ALT), aspartate aminotransferase (AST), albumin (ALB), glucose (GLU), and total protein (TP) using a fully automatic biochemical analyzer (Mindray, BS280, Shenzhen, China) [
26].
2.8. Intestinal Antioxidant Enzyme Activity Assays
Antioxidant enzyme activities in the proximal, middle, and distal intestines of largemouth bass were measured using commercial kits from Nanjing Jiancheng Bioengineering Institute (Nanjing, China). Superoxide dismutase (SOD) and catalase (CAT) activities and the total protein content (TP) were determined following the manufacturer’s protocols [
25,
26].
2.9. Real-Time Quantitative PCR Analysis
Total RNA was extracted from the proximal, middle, and distal intestine of each group of largemouth bass using the RNAex Pro Reagent (Hunan Akrie Biotechnology Co., Ltd., Changsha, China) and quantified using a NanoDrop 2000 (Thermo Scientific, Waltham, MA, USA). cDNA was synthesized from total RNA using the gDNA Clean Reaction Mix Ver.2 and 5X Evo M-MLV RT Reaction Mix II (Yeasen Biotechnology (Shanghai) Co., Ltd., Shanghai, China) and stored at −20 °C. Primers for real-time quantitative PCR (RT-qPCR) were designed using Primer Premier 5.0 and synthesized by Sangon Biotech (Shanghai) Co., Ltd. (Shanghai, China) (
Table 2). RT-qPCR was performed using the CFX 96TM Real-Time System (Bio-Rad, Hercules, CA, USA). The total reaction volume was 20 μL, including 10 μL of SYBR Green, 0.4 μL of upstream and downstream specific primers (10 mM each), 1 μL of template cDNA, and 8.2 μL of sterile deionized water. The reaction protocol included an initial denaturation at 95 °C for 30 s, followed by 40 cycles of 95 °C for 5 s, annealing at 60 °C for 30 s, and a final melting curve analysis from 65 °C to 95 °C with a 5 s hold every 0.5 °C. β-actin was used as the reference gene, and relative gene expression was measured using the 2
−△△Ct method [
26,
27].
2.10. Gut Microbiota Analysis
Total intestinal genomic DNA was extracted using the E.Z.N.A.
® Soil DNA Kit (Omega, Irving, TX, USA). The V3+V4 region of 16S rRNA was amplified with primers 338F (5′-ACTCCTACGGGAGGCAGCA-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′). Sequencing was performed on the Illumina MiSeq platform (Shanghai Majorbio Biopharm Technology Co., Ltd., Shanghai, China). Raw data were merged using FLASH (v1.2.7) and filtered for quality with QIIME2 (v2020.2), followed by chimera removal with UCHIME (v8.1) to obtain high-quality tags. Effective sequences were analyzed using USEARCH 10.0 for OTU clustering at 97% similarity. Taxonomic annotation of OTUs was performed with the RDP Classifier 2.2 against the Silva 16S rRNA gene database (v132,
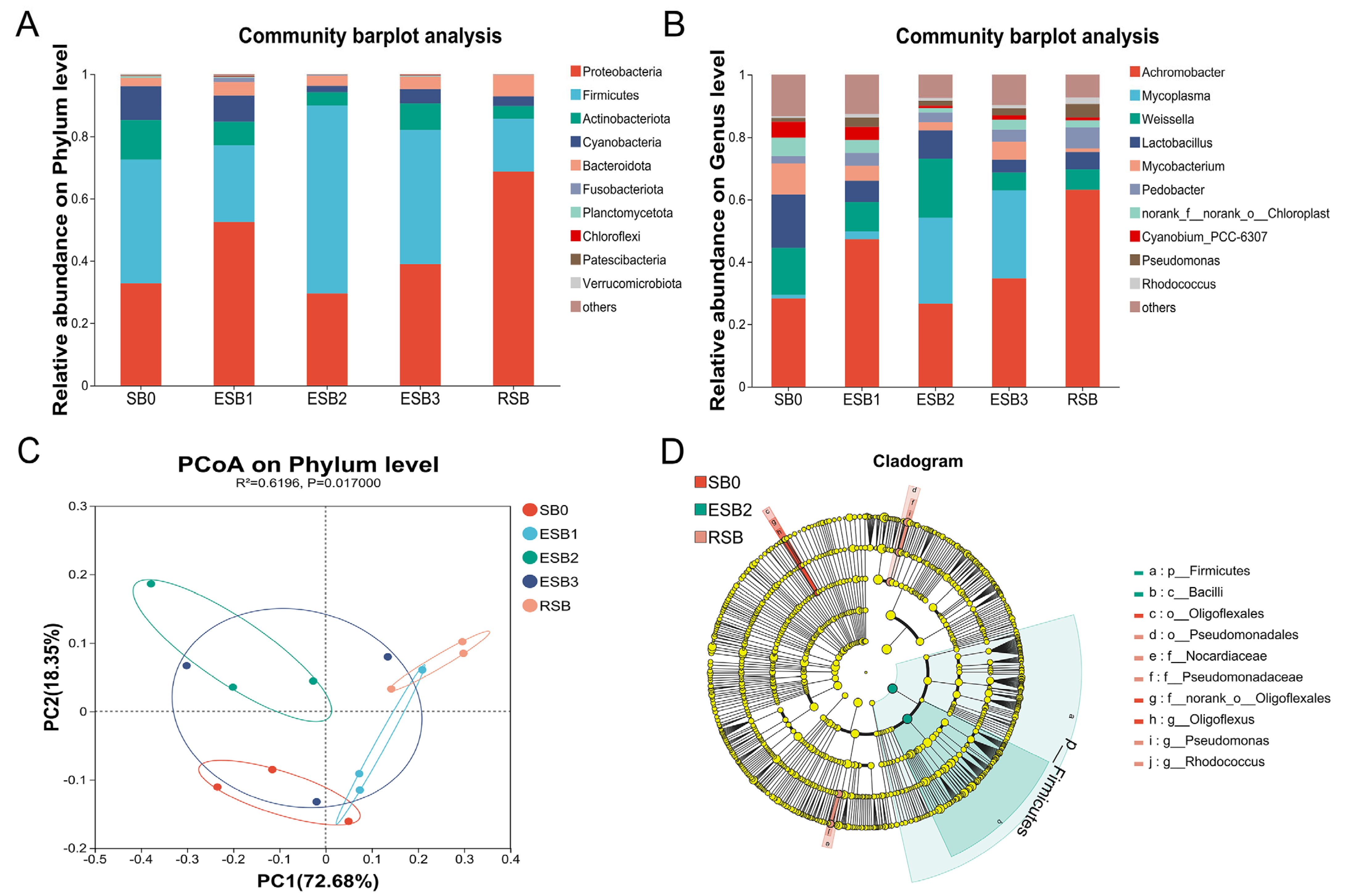
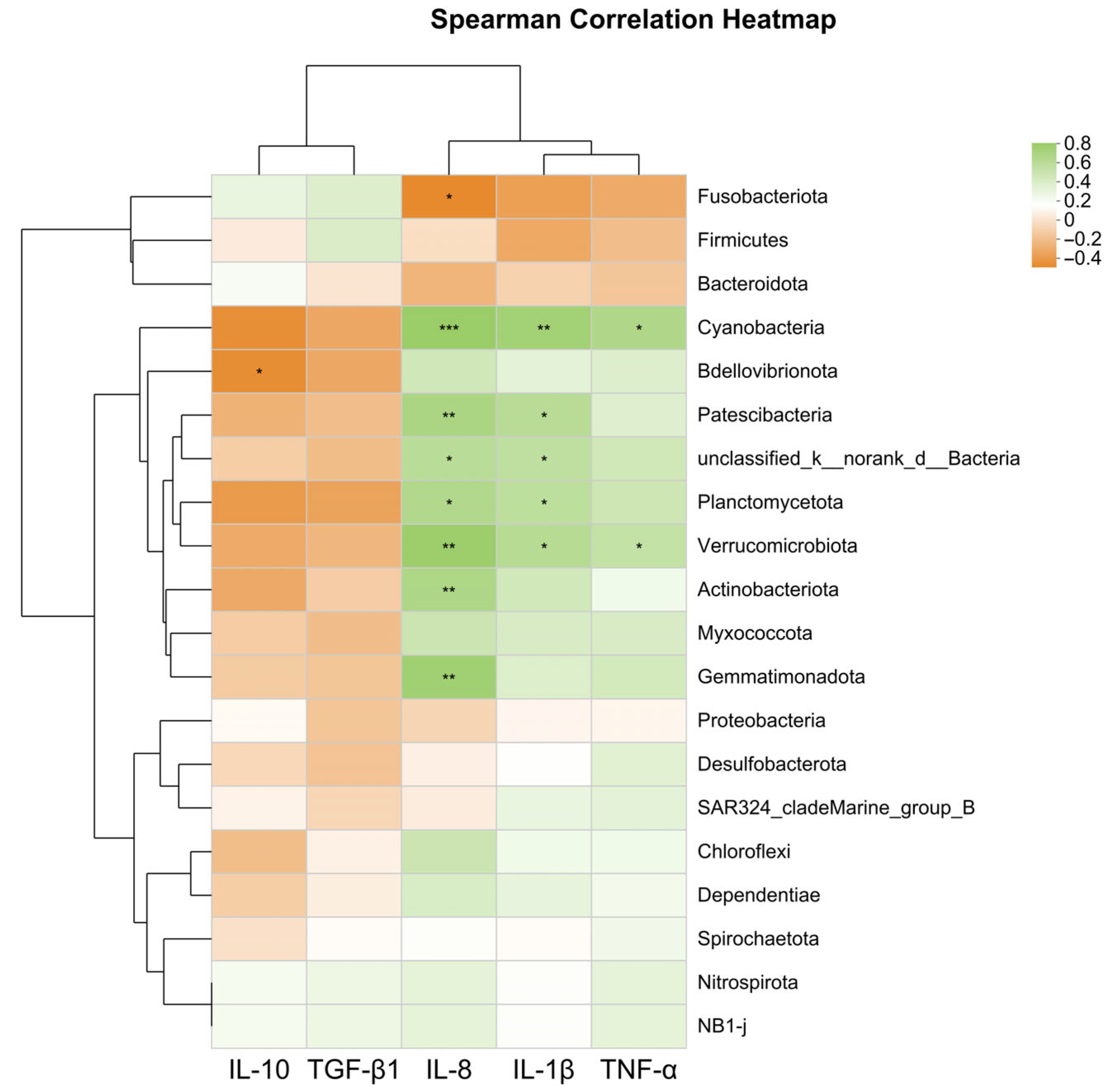
http://www.arb-silva.de accessed on 11 April 2025). Microbial community composition was profiled at various taxonomic levels (phylum, class, order, family, genus, species), and diversity indices (Shannon, Simpson, Chao, Ace) were calculated. β-diversity was assessed via principal coordinate analysis (PCoA, Vegan v2.4.3), and differentially abundant biomarkers were identified using linear discriminant analysis effect size (LEfSe) [
28].
2.11. Data Analysis
Data are expressed as the mean ± standard error (mean ± S.E.) (n = 3). Statistical analyses were performed using IBM SPSS Statistics 26.0, and graphs were generated with GraphPad Prism 9.0. Prior to analysis, data that deviated from the population mean were identified and excluded through a one-sample t-test (α = 0.05). One-way ANOVA followed by Duncan’s multiple range test was used for intergroup comparisons. Paired-samples t-tests were used for intragroup comparisons of gut locations. Significance was set at p < 0.05. Significant differences among different gut locations within the same SB group are indicated by different capital letters, while significant differences among different SB groups within the same gut location are indicated by different lowercase letters.
5. Conclusions
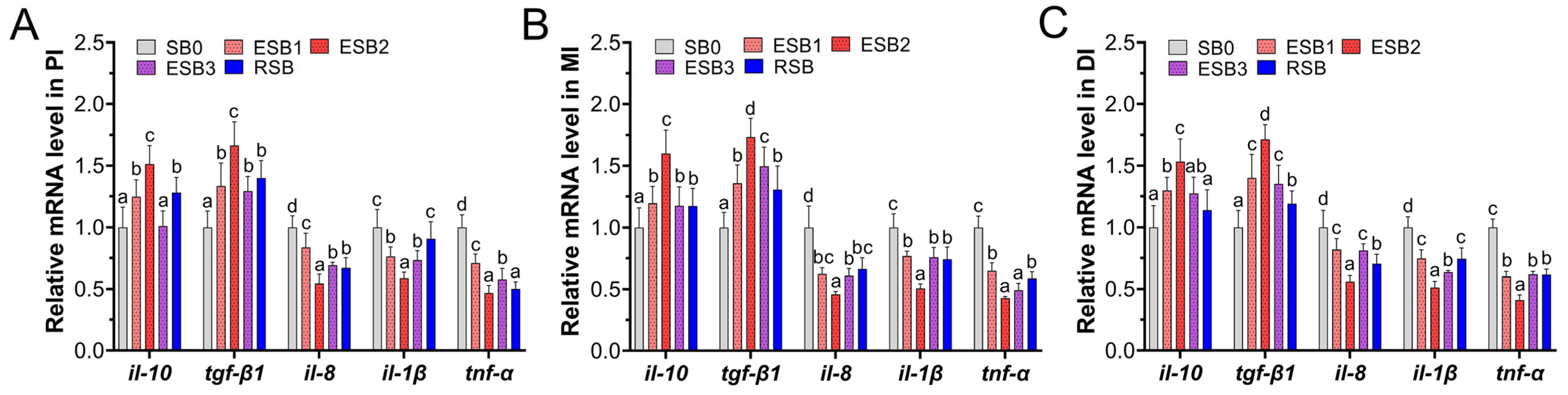
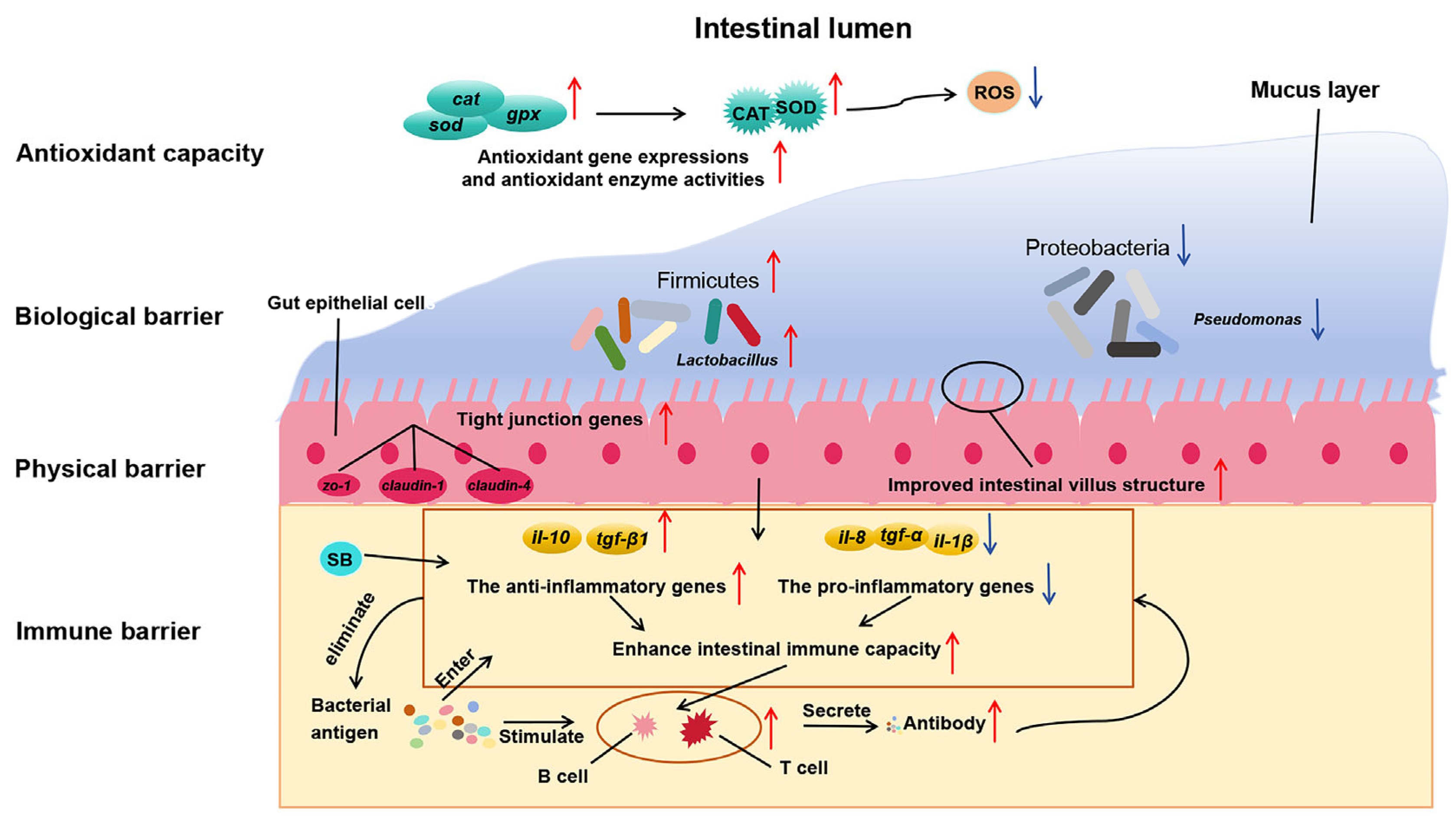
In this study, both encapsulated SB and raw powder SB up-regulated the expression of tight junction genes (zo-1, claudin-1, and claudin-4), thereby enhancing the intestinal physical barrier function and improving nutrient absorption efficiency. They also promoted the gene expression of anti-inflammatory factors (tgf-β1 and il-10) and inhibited the gene expression of pro-inflammatory factors (tnf-α, il-1β, and il-8), thereby strengthening the intestinal immune barrier. Moreover, adding 1000 and 1500 mg/kg encapsulated SB enhanced the expression of antioxidant-related genes (sod, cat, and gpx) and enzyme activities (SOD and CAT), alleviating oxidative stress damage and systematically improving the fish’s antioxidant capacity. In this study, adding 1500 mg/kg encapsulated SB improved nutrient absorption efficiency, thereby achieving efficient growth conversion, enhancing the growth performance of largemouth bass, and significantly increasing the abundance of beneficial bacteria (Firmicutes) while reducing the proportion of potential pathogenic bacteria (Proteobacteria), thus maintaining intestinal microbial balance and enhancing the intestinal biological barrier. In summary, adding 1500 mg/kg encapsulated SB (ESB2 group) was the most effective, significantly improving the growth performance, intestinal health, and immune function of largemouth bass. This study provides a theoretical basis for developing efficient and eco-friendly feed additives and reduces the farming cost of largemouth bass.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}