1. Introduction
Haliotis discus hannai, a pivotal subspecies within the genus Haliotis (family Haliotidae) [
1], predominantly inhabits rocky substrates across tropical to temperate marine zones [
2]. Within these habitats, specific water temperature, transparency, and light intensity constitute critical ecological determinants, and wild populations establish stable ecological niches in algae- and kelp-rich environments [
3], exhibiting trophic specialization through macroalgal consumption. Despite these specific habitat requirements in the wild, H. discus hannai is recognized as the cornerstone species in China’s abalone aquaculture industry. H. discus hannai is the most extensively farmed abalone species in northern China, especially in the Shandong, Liaoning, and Fujian provinces. It constitutes > 70% of China’s farmed abalone output due to its rapid growth, superior flesh quality, and adaptability to controlled environments [
4]. However, industrial-scale farming faces dual challenges: escalating biosecurity risks from pathogenic infections, and systemic nutritional deficits.
Vibriosis and pseudomonad-induced pathologies (such as pustule disease and abdominal rupture) have emerged as primary biosecurity threats [
5,
6,
7,
8], with elevated water temperatures during summer exacerbating pathogen proliferation, immunosuppression, and mass mortality events. Concurrently, high-density farming amplifies pathogen transmission via hydrodynamic vectors. Traditional algal-based feeds fail to meet protein–lipid requirements, while artificial formulations, despite storage stability advantages, suffer from suboptimal cellulose digestibility and imbalanced amino acid profiles, collectively impairing growth performance and elevating production costs [
9]. These intertwined constraints necessitate innovative technological interventions.
Probiotics, as bioactive microbial preparations, demonstrate multifaceted therapeutic potential through host–microbiome crosstalk modulation. Their tripartite mechanisms encompass the following: gut microbiota regulation via the competitive exclusion of pathogens (Vibrio harveyi) and short-chain fatty acid (SCFA)-mediated enteric homeostasis [
10]; nutrient metabolism enhancement through the enzymatic hydrolysis of recalcitrant feed components (cereal proteins/polysaccharides), achieving 8–34% weight gain improvement [
11,
12,
13]; and immunomodulation via microbial-associated molecular patterns (MAMPs) activating pattern recognition receptors (PRRs), thereby triggering TLR4/NRF2/KEAP1 signaling cascades to suppress pro-inflammatory cytokines (TNF-α, IL-16) [
14]. Notably, Shewanella-supplemented diets reduce
V. harveyi mortality from 77% to 27% [
15], while
Bacillus strains enhance hemocyte vitality and apoptosis resistance [
16]. Furthermore, probiotics exhibit an environmental bioremediation capacity, degrading 99% of nitrites and ammonia nitrogen in aquaculture systems [
17].
Lactobacillus plantarum (LP) was selected as the focal probiotic agent in this study due to its (1) well-documented safety (GRAS/QPS status) and stability in feed matrices, (2) robust enzymatic repertoire (proteases, carbohydrases) that enables the efficient degradation of complex plant-derived antinutrients, (3) potent SCFA (particularly lactate) production capacity, which acidifies the gut lumen to inhibit Vibrio spp. colonization, and (4) proven immunostimulatory effects in aquatic species via the upregulation of antioxidant pathways and antimicrobial peptide expression [
17]. Its resilience to industrial processing (lyophilization, pelleting) further enhances its practical applicability in aquaculture feed production.
The synergistic integration of probiotic engineering with feed biotechnology presents transformative potential. LP-fermented feeds degrade antinutritional factors, enrich γ-aminobutyric acid (GABA) and α-linolenic acid (ALA) profiles [
18], and activate the tryptophan metabolism. Probiotic formulations achieve mortality reduction [
19] and reshape gut microbiomes by elevating Firmicutes abundance, fostering nutrient assimilation and immune priming [
20]. The primary goal of this investigation is to rigorously evaluate the efficacy of a novel LP-fermented feed in (1) augmenting growth performance and feed utilization efficiency, (2) enhancing resistance against Vibrio harveyi challenges, and (3) modulating key immune-physiological and metabolic responses (e.g., microbiome structure, antioxidant status). This paradigm not only truncates traditional 3–4 year cultivation cycles, but enables precision health management through microbiome–metabolome axis elucidation (Halomonas–hydroxyethyl sulfonate metabolic coupling).
3. Results
3.1. Analysis of Feed Quality Before and After Fermentation
3.1.1. Sensory Evaluation of Fermented Feed
As shown in
Table 1 and
Figure 1, the color of LP-fermented feed exhibited minimal variation compared to the control group, both presenting a brownish-yellow hue. Post-fermentation, the aroma of soybean meal transitioned from its original bean cake scent to a distinctive sour and alcoholic fragrance. The addition of fermentation broth increased feed moisture content, resulting in a softer texture with minor agglomeration, although no significant pH alteration was observed (pH 6.5 vs. 6.7).
3.1.2. Impact of Probiotic Fermentation on the Nutritional Profile of Soybean Meal
As indicated in
Table 2, LP fermentation notably enhanced specific fatty acid contents. The LP group demonstrated a two-fold increase in cis-9-palmitoleic acid (106.85 ± 21.37 mg/kg vs. 48.32 ± 9.18 mg/kg in CK) and a 14% elevation in α-linolenic acid (ALA) content (292.6 ± 55.59 mg/kg vs. 256.6 ± 33.36 mg/kg). However, docosahexaenoic acid (DHA) levels decreased by 11.8% (66.47 ± 13.29 mg/kg vs. 75.33 ± 10.55 mg/kg), warranting further investigation into fermentation-induced metabolic pathway alterations. A marginal increase was observed in cis-11,14,17-eicosatrienoic acid (2.47 ± 0.3 mg/kg vs. 1.97 ± 0.28 mg/kg).
Concerning saturated fatty acids (
Table 3), caprylic acid (C8:0) content decreased by 20.53% in the LP group, while lauric acid (C12:0), tridecanoic acid (C13:0), and myristic acid (C14:0) exhibited dramatic increases of 357.14%, 163.64%, and 51.42%, respectively.
3.1.3. Amino Acid Profile Modifications Induced by Probiotic Fermentation
Fermentation significantly elevated amino acid levels in the LP group compared to CK(
Table 4). The most pronounced increases were observed in L-histidine (+38.69%), L-asparagine (+36.10%), γ-aminobutyric acid (+32.88%), and L-Arginine (+29.51%).
3.2. Production Performance
Production performance refers to the comprehensive evaluation of abalone growth, survival rate, and feed utilization efficiency during aquaculture, serving as a critical indicator for assessing rearing conditions, feed quality, and management efficacy. As detailed in
Table 5, the growth performance of abalone varied significantly across feed treatment groups. The weight gain rate in the LP group (421%) was significantly higher than that in the CK (201%) and HD (263%) groups.
The feed conversion ratio (FCR), a key metric for evaluating feed utilization efficiency, was calculated as 2633.12 for the LP group, lower than both the CK (1586.37) and HD (1205.36) groups. In terms of survival rate, the LP group achieved 61.66%, outperforming the CK group (45.33%), but remaining lower than the HD group (69%). Notably, while the total weight gain rate of the LP group (263%) substantially exceeded that of the CK group (201%), it was markedly lower than the HD group’s 421%.
3.3. Determination of Abalone Quality
The experiment compared changes in amino acid content in abalone muscle among the HD group (kelp-based feed), LP group (LP-fermented feed), and CK group (basal feed) (
Table 6). The results reveal that the LP group exhibited higher concentrations of most amino acids in abalone muscle compared to both the HD and CK groups. Specifically, histidine, phenylalanine, and cystine levels in the LP group were 0.21 g/kg, 0.41 g/kg, and 0.11 g/kg, respectively. These values represented increases of 0.10 g/kg, 0.09 g/kg, and 0.10 g/kg compared to the CK group (0.11 g/kg, 0.32 g/kg, and 0.09 g/kg), and increments of 0.03 g/kg, 0.04 g/kg, and 0.10 g/kg relative to the HD group (0.18 g/kg, 0.37 g/kg, and 0.01 g/kg). These findings indicate that fermented feed enhances the umami-related components in abalone muscle.
3.4. Effects of LP-Fermented Feed on Intestinal Morphology of Abalone
A comparative analysis of intestinal tissue structure (
Figure 2) revealed significant morphological differences in abalone intestines across feed treatment groups. In the HD group, the intestinal architecture remained relatively intact, characterized by well-developed villi and a continuous mucosal layer. The abundant villous structure effectively expanded the nutrient absorption surface area, thereby enhancing dietary nutrient utilization and promoting growth. These findings suggest that kelp-based feed may exert protective effects on intestinal integrity, supporting barrier function and nutrient assimilation. In contrast, the CK group exhibited disorganized intestinal folds with shortened, sparse villi and localized structural damage, indicating the limited benefits of basal feed on intestinal health. Such compromised morphology likely reduced nutrient absorption efficiency, ultimately impairing growth performance.
The LP group demonstrated markedly improved intestinal integrity compared to CK, with significantly elongated and denser villi and no observable structural defects. Although slightly inferior to the HD group, the LP group’s intestinal morphology surpassed that of CK, highlighting that LP NDHJ-4 fermentation enhanced digestive capacity and intestinal health in H. discus hannai through structural optimization.
3.5. Effects of LP-Fermented Feed on Intestinal Barrier Function, Cytokines, and Antioxidant Factors in Abalone
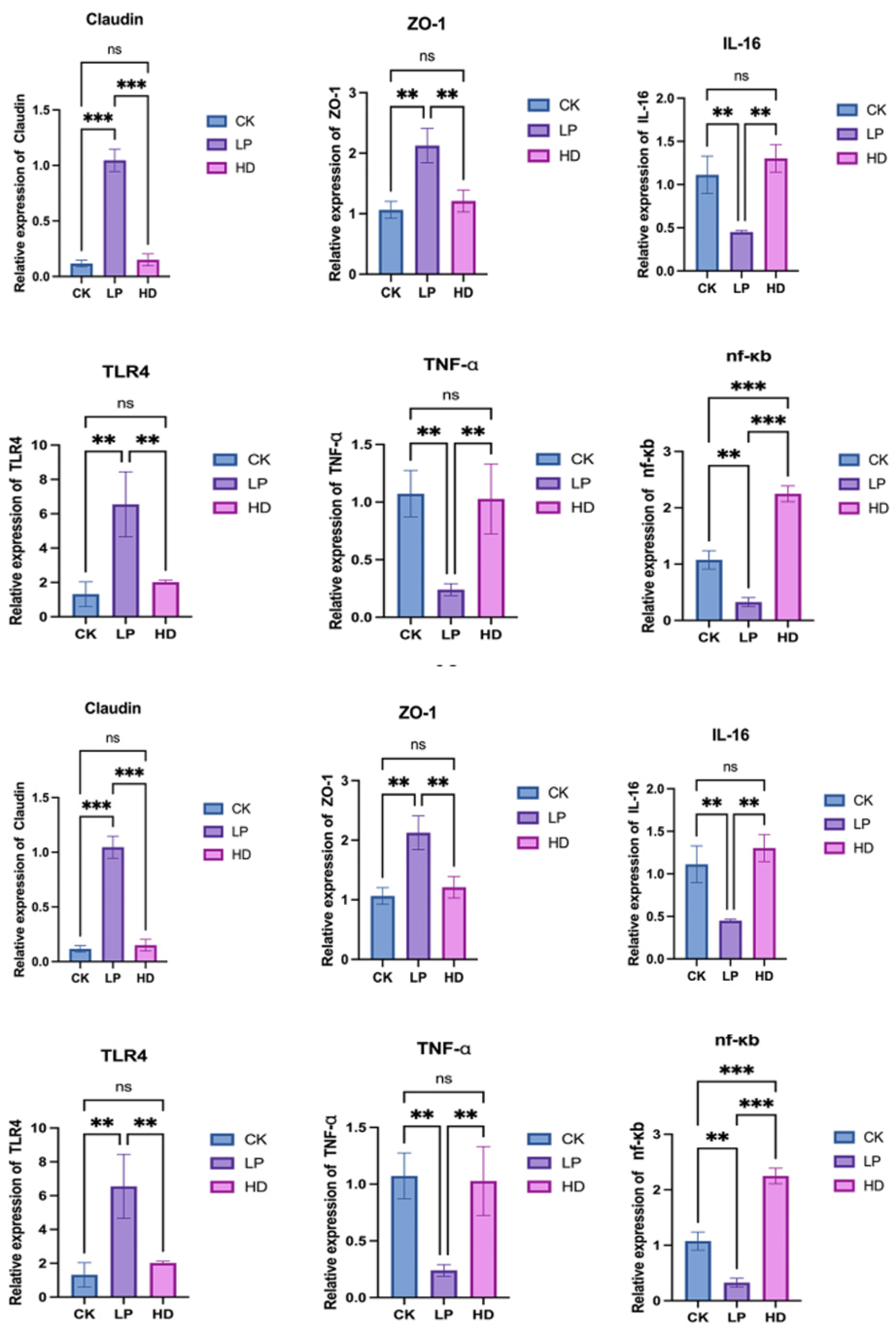
A comparative analysis of intestinal tight junction protein mRNA expression across feed treatment groups (
Figure 3) revealed distinct molecular responses. The LP group exhibited significantly elevated Claudin mRNA expression compared to both CK and HD groups (
p < 0.001), with no significant difference observed between CK and HD (ns). Similarly, ZO-1 mRNA expression was markedly higher in the LP group than in CK and HD (
p < 0.01), indicating enhanced tight junction assembly and intestinal barrier integrity in the fermented feed group.
Inflammatory and immune-related cytokine profiling demonstrated the regulatory effects of LP treatment. While TLR4 mRNA expression showed no difference between CK and HD, it was significantly upregulated in LP (p < 0.01). Conversely, TNF-α mRNA expression in LP decreased sharply compared to CK and HD (p < 0.01), with no intergroup difference between CK and HD. Notably, NF-κB mRNA levels were lowest in LP, while HD displayed the highest expression (p < 0.001 vs. both groups), and CK levels exceeded LP (p < 0.01). Additionally, IL-16 mRNA expression in LP was reduced by 42% compared to CK and HD (p < 0.01), further suggesting that fermented feed mitigates inflammatory signaling.
An antioxidant pathway analysis revealed that Keap1 mRNA expression peaked in CK, but was suppressed in LP and HD (p < 0.001). Although Nrf-2 mRNA levels showed no difference between CK and LP, HD exhibited significantly higher Nrf-2 expression than CK (p < 0.05), highlighting divergent antioxidant activation mechanisms.
3.6. Effects of Fermented Feed on the Intestinal Microbiota of Abalone
3.6.1. OTU Distribution and Diversity Analysis
The Venn diagram (
Figure 4) illustrates the similarity of operational taxonomic unit (OTU) composition among three abalone intestinal microbiota groups. A total of 46,574 OTUs were identified across all groups, with 3063 OTUs shared among all three groups. Specifically, the CK group contained 13,159 OTUs (4879 unique), the HD group exhibited the highest richness with 17,963 OTUs (10,191 unique), and the LP group comprised 16,318 OTUs (9795 unique). This hierarchical distribution indicates that the HD group demonstrated the highest OTU richness, followed by the LP and CK groups.
Alpha diversity indices were analyzed to evaluate microbial community characteristics (
Table 7). The HD group showed significantly higher Observed_species, Shannon, and Simpson indices compared to the CK and LP groups (
p < 0.05), while no significant differences were observed between the CK and LP groups. Notably, the CK group exhibited the highest Chao1 and ACE indices, reflecting distinct patterns in species richness estimation. These results suggest that LP and HD feed modulates intestinal microbiota by enhancing microbial diversity (Shannon: 4.12 in CK vs. 4.27 in LP vs. 4.79 in HD) while maintaining comparable species richness to basal feed.
As illustrated in the PCoA plot (Unweighted Unifrac distance), each point represents an individual abalone sample, with the color corresponding to experimental groups (HD, LP, CK). The analysis revealed distinct clustering patterns: samples within the same group exhibited tight aggregation, while intergroup points were spatially separated without overlap. This clear segregation (PERMANOVA, * p < 0.001) indicates that dietary interventions significantly restructured the intestinal microbial community composition of abalone.
3.6.2. Effects of Fermented Feed on Gut Microbiota Composition in Abalone
Following species annotation processing, a comparative analysis of microbial communities was conducted based on relative abundance at both the phylum and genus levels. A total of 37 bacterial phyla and 1156 bacterial genera were identified across all intestinal samples. Based on annotation results, the top 10 phyla accounting for over 99% of total annotated species in abalone gut microbiota from different marine areas were selected for analysis.
Figure 4 illustrates the phylum-level effects of fermented feed on abalone gut microbiota. The dominant phyla included Proteobacteria, Tenericutes, Bacteroidetes, Firmicutes, Actinobacteria, Planctomycetes, and Verrucomicrobia. Significant intergroup differences were observed in Acidobacteria, Spirochaetes, and Chloroflexi (
p < 0.05).
Notably, Proteobacteria showed no significant variation among the experimental groups. The LP group exhibited the significant upregulation of Tenericutes (p = 0.017) and downregulation of Bacteroidetes (p = 0.023) compared to controls. Although Firmicutes abundance showed no statistical difference from the CK group, it was significantly lower than the HD group (p = 0.035).
At the genus level (
Figure 4C), the dominant genera comprised
Curvibacter,
Mycoplasma,
Asinibacterium,
Mycoplasmopsis,
Ochrobactrum,
Pandoraea,
Hydrotalea, and
Pelagibacterium. The LP group demonstrated 3.08-fold and 14.42-fold increases in
Mycoplasma and
Ochrobactrum abundance, respectively, versus CK group, concomitant with 2.35-fold reduction in
Pelagibacterium.
Curvibacter,
Asinibacterium,
Pandoraea, and
Hydrotalea showed no significant differences from the CK group, but were notably reduced compared to the HD group.
Linear discriminant analysis (LDA > 4) revealed 31 differentially abundant species across groups (
Figure 4(A3)). The CK, HD, and LP groups contained seven, fourteen, and ten discriminant species, respectively, contributing to microbial structure variation. Notably,
Lactobacillus plantarum-fermented feed enhanced differential microbiota diversity in H. discus hannai compared to the CK group. Although the LP group showed slightly lower diversity than the seaweed-fed HD group, its increased discriminant species count relative to the CK group indicates probiotic-fermented feed’s modulatory effect on intestinal microbial diversity.
3.7. Metabolite Identification and Quantification
A total of 2478 compounds were identified through LC-MS-based metabolomic profiling combined with an in-house database. The annotated metabolites were classified into 14 major categories (
Figure 5): lipids and lipid-like molecules (877), organoheterocyclic compounds (391), organic oxides (167), organic acids and derivatives (460), organohalides (3), organosulfur compounds (11), organonitrogen compounds (52), 1,3-dipolar organic compounds (1), homogeneous non-metallic compounds (1), hydrocarbons and derivatives (7), alkaloids and derivatives (55), lignans/neolignans (8), nucleosides/nucleotides/analogs (52), benzenoids (243), phenylpropanoids/polyketides (120), and others (30).
3.7.1. OPLS-DA Modeling
Untargeted LC-MS/MS metabolomics was performed on ten intestinal samples per group. Orthogonal Partial Least Squares Discriminant Analysis (OPLS-DA) with 1000 permutation tests (to mitigate overfitting risks) revealed distinct clustering patterns among the three groups (
Figure 4B). The clear spatial separation (R2Y > 0.98 for all pairwise comparisons) and absence of intergroup overlap confirmed significant metabolic profile differences. Model validity was further supported by high predictive capacity: CK vs. HD (R2Y = 0.99, Q2 = 0.693), CK vs. LP (R2Y = 0.993, Q2 = 0.951), and HD vs. LP (R2Y = 0.985, Q2 = 0.922). All Q2 values exceeded the 0.5 threshold, confirming robust model reliability (
Figure 4C).
3.7.2. Screening of Differential Metabolites
Differential metabolites were identified using Variable Importance in Projection (VIP) scores from the first principal component of Orthogonal Partial Least Squares-Discriminant Analysis (OPLS-DA) models combined with independent
t-test
p-values. Among the 3254 detected metabolites, 959 showed significant intergroup variations (VIP > 1.0,
p < 0.05), including 664 upregulated and 405 downregulated compounds in LP compared to CK (
Table 8). The key metabolic pathways impacted by fermented feed included lipid biosynthesis (23.1%), amino acid metabolism (18.7%), and secondary metabolite production (14.5%).
3.7.3. Classification of Differential Metabolites
In the CK vs. HD comparison, 494 differential metabolites were identified, including 167 lipids and lipid-like molecules, 96 organoheterocyclic compounds, 35 organic oxides, 93 organic acids/derivatives, 1 organohalide, 2 organosulfur compounds, 11 organonitrogen compounds, 2 hydrocarbons, 14 alkaloids/derivatives, 1 lignan/neolignan, 12 nucleosides/nucleotides/analogs, 32 benzenoids, 24 phenylpropanoids/polyketides, and 4 others, with 286 upregulated and 208 downregulated metabolites.
The CK vs. LP comparison revealed 959 differential metabolites: 346 lipids/lipid-like molecules, 185 organoheterocyclic compounds, 68 organic oxides, 153 organic acids/derivatives, 3 organohalides, 4 organosulfur compounds, 14 organonitrogen compounds, 2 hydrocarbons, 18 alkaloids/derivatives, 4 lignans/neolignans, 18 nucleosides/nucleotides/analogs, 89 benzenoids, 46 phenylpropanoids/polyketides, and 9 others, showing 664 upregulated and 405 downregulated species.
For HD vs. LP, 963 differential metabolites were detected: 355 lipids/lipid-like molecules, 173 organoheterocyclic compounds, 65 organic oxides, 155 organic acids/derivatives, 2 organohalides, 5 organosulfur compounds, 13 organonitrogen compounds, 4 hydrocarbons, 15 alkaloids/derivatives, 2 lignans/neolignans, 1 homogeneous non-metallic compound, 17 nucleosides/nucleotides/analogs, 99 benzenoids, 50 phenylpropanoids/polyketides, and 7 others, with 460 upregulated and 503 downregulated metabolites.
Cross-group analysis identified lipids/lipid-like molecules, organoheterocyclic compounds, organic acids/derivatives, and benzenoids as the predominant differential metabolite categories. A core set of 149 shared differential metabolites was observed across all comparisons (
Figure 5).
3.7.4. KEGG Pathway Enrichment Analysis
KEGG pathway analysis (FDR-adjusted
p < 0.05) revealed the significant enrichment of differential metabolites in ovarian steroidogenesis (
p = 0.018), tryptophan metabolism (
p = 0.026), and pantothenate/CoA biosynthesis (
p = 0.042) (
Figure 4). These pathways are hypothesized to regulate growth performance and immune responses in H. discus hannai.
3.7.5. Shared Differential Metabolites Across Groups
Fourteen high-confidence differential metabolites (VIP > 1.0,
p < 0.05) were identified between the CK and experimental groups (HD/LP) after filtering unnamed compounds (
Table 9). Three metabolites were downregulated in the LP/HD groups: cis-11,14,17-eicosatrienoic acid, thermozymocidin, and nabilone. Eleven metabolites exhibited upregulated trends, including the key intermediates of lipid remodeling (lysophosphatidylcholine 18:1) and amino acid derivatives (N-acetylglutamine).
3.8. Correlation Analysis of Differential Microbial Genera, Metabolites, and Production Performance in H. discus hannai
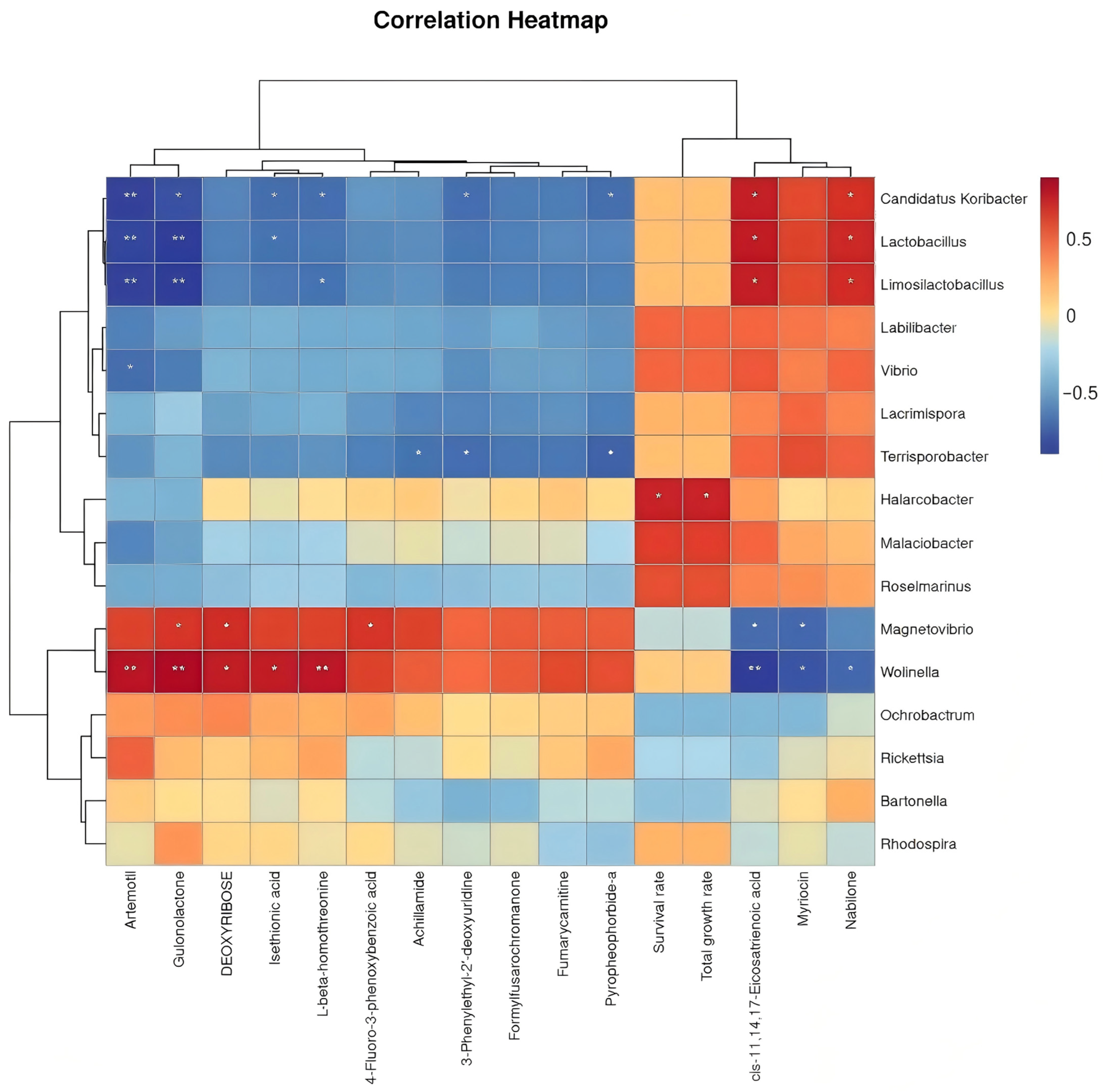
To elucidate the potential mechanisms by which LP-fermented feed modulates abalone production performance through intestinal microbiota–metabolite crosstalk, a Spearman correlation analysis was conducted between the differential bacterial genera, metabolites, growth indices (total weight gain rate), and survival rate. The association heatmap (
Figure 6) reveals distinct interaction patterns, where the red and blue regions denote positive and negative correlations, respectively, reflecting the potential microbial regulation of metabolic pathways and host physiology.
4. Discussion
This study multidimensionally evaluates the effects of soybean meal fermented with Lactobacillus plantarum (LP) on feed quality, growth performance, intestinal health, and metabolic regulation in abalone (H. discus hannai). The results demonstrate that LP fermentation significantly enhanced key fatty acids in soybean meal, including a two-fold increase in cis-9-palmitoleic acid and a 14% elevation in α-linolenic acid (ALA), alongside notable improvements in amino acids such as L-histidine and L-asparagine.
Critically, these nutritional enhancements translated to significant biological outcomes. Feeding with LP-fermented feed markedly improved abalone growth rate (263% vs. 201% in the control group, CK) and survival rate (61.66% vs. 45.33% in CK). Beyond growth metrics, mechanistic investigations revealed profound effects on intestinal physiology: LP fermentation enhanced intestinal morphology (elongated villi and structural integrity), upregulated tight junction proteins (Claudin and ZO-1), suppressed inflammatory cytokines (TNF-α, IL-16), and activated antioxidant pathways (Keap1 downregulation), collectively optimizing intestinal health.
Further elucidating the drivers of improved health and growth, LP-fed abalone exhibited increased gut microbiota diversity, elevated abundances of Mycoplasma and Ochrobactrum, and differential metabolites enriched in tryptophan metabolism and the pantothenate/CoA biosynthesis pathways, revealing a potential microbiota–metabolism axis driving growth promotion.
Positioning these findings within existing knowledge, this study aligns with prior research on probiotic-fermented feeds in aquaculture, but also unveils novel insights. Regarding nutritional enhancement, earlier studies consistently reported that microbial fermentation degrades antinutritional factors in soybean meal and liberates free amino acids. For instance, N. Muhamad Nor [
21] observed significant increases in lysine and methionine content after lactic acid bacteria fermentation, consistent with the marked elevation of L-histidine and L-asparagine here. However, our work uniquely highlights the paradoxical enrichment of ω-3 fatty acids (ALA) alongside DHA reduction—a phenomenon seldom reported. This discrepancy may stem from LP’s metabolic specificity: earlier studies suggested that lactic acid bacteria preferentially degrade long-chain fatty acids via β-oxidation [
22], potentially accelerating DHA catabolism, while ALA accumulation could arise from bacterial biosynthesis or selective substrate utilization [
23]. These nuanced findings underscore the need for multi-omics approaches (e.g., lipidomics and transcriptomics) to dissect fatty acid metabolic networks.
Transitioning to growth performance, while LP-fed abalone exhibited higher weight gain than CK, it underperformed compared to the seaweed-based HD group—a result partially congruent with [
24], who demonstrated that fermented soybean meal could replace 30% of fishmeal without compromising fish growth. The likely explanation for HD’s superiority resides in its polysaccharide-rich matrix and trace element content, unmatched by fermented feeds. Notably, a key advantage of the LP diet emerged in efficiency: the significantly lower feed conversion ratio (FCR) implies enhanced nutrient utilization, a mechanism plausibly linked to the observed intestinal remodeling and microbiota modulation. For example, villus elongation directly expands nutrient absorption surfaces [
25], while upregulated tight junction proteins fortify barrier integrity, mitigating leaky gut risks [
26]. These observations not only corroborate the “gut health–nutrient utilization” axis proposed in probiotic studies, but extend it by integrating structural, immunological, and microbial dimensions—a holistic perspective that was previously absent.
The gut microbiota findings further reveal both consonance and divergence from the literature. The marked enrichment of Tenericutes and Mycoplasma in LP-fed abalone contrasts with Ifra Ghori et al. (2022) [
27], who reported the probiotic-driven suppression of conditional pathogens in fish. Significantly, the 14-fold surge in Ochrobactrum—a genus renowned for xenobiotic degradation [
28]—suggests its role in detoxifying feed-derived contaminants, indirectly bolstering host health. Nevertheless, comparative analysis highlights a challenge: the higher OTU diversity in the HD group underscores the ecological superiority of natural diets in maintaining microbial stability, urging the future exploration of hybrid feeding strategies.
Metabolomic insights add another layer of understanding, delineating the multidimensional regulatory effects of LP fermentation. Differential metabolites enriched in the tryptophan metabolism (kynurenine pathway) align with suppressed TLR4/NF-κB signaling, implying that tryptophan derivatives mitigate intestinal inflammation—a mechanism known in mammals [
29], now extended to mollusks. Crucially, the upregulation of artemotil and isethionic acid, correlated with Wolinella abundance and growth metrics, provides direct evidence for a “microbiota–metabolite–host” axis. This multi-omics integration transcends conventional single-parameter analyses, offering unprecedented resolution into probiotic mechanisms—an advancement addressing critical gaps in earlier aquaculture research.
Finally, this study identifies some key limitations that point to future directions: the metabolic mechanism driving DHA reduction remains unclear, potentially addressable by stable isotope tracing; the absence of long-term assessments on reproductive performance necessitates extended trials; and the lack of functional gut microbiota profiling (metagenomics) limits causal insights into microbiota–metabolite interactions, highlighting the need to integrate metagenomics with metabolic flux studies. And the potential for enhanced resistance against specific pathogens like Vibrio harveyi requires validation through dedicated challenge trials. Additionally, exploring yeast co-fermentation strategies may further optimize fatty acid composition and feed’s nutritional value.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}