

Potential of LP as a Biocontrol Agent for Vibriosis in Abalone Farming

Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Reagents and Instruments
2.3. Isolation of LP
2.4. Strain Purification and Cultivation
2.5. Morphological Characterization
2.6. Physiological and Biochemical Analysis
2.7. 16S rDNA Sequencing and Phylogenetic Analysis
2.8. Probiotic Property Evaluation
2.8.1. Antibacterial Activity Assay
2.8.2. Minimum Inhibitory Concentration (MIC)
2.8.3. Hemolytic Activity
2.8.4. Biofilm Formation Assay
2.8.5. Antibiotic Susceptibility
2.8.6. Bile Salt Tolerance
2.8.7. Acid and Alkali Resistance
2.8.8. Heat Tolerance
2.9. Adhesion Assays
2.9.1. Auto-Aggregation
2.9.2. Hydrophobicity
2.10. Pathogenicity Challenge in Abalone
2.10.1. Abalone Rearing
2.10.2. Bacterial Preparation
2.10.3. Challenge Experiment
2.11. Statistical Analysis
3. Results
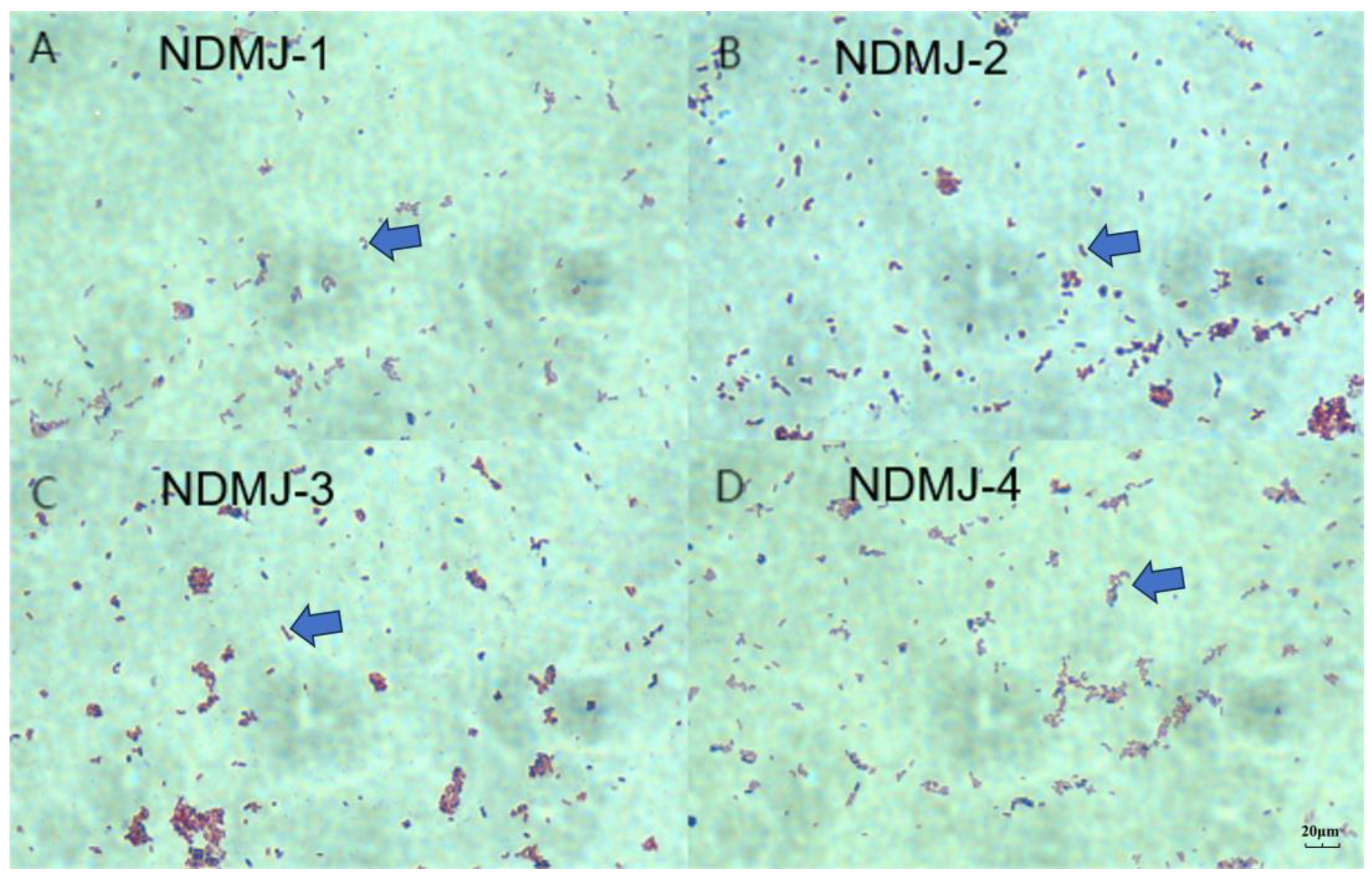
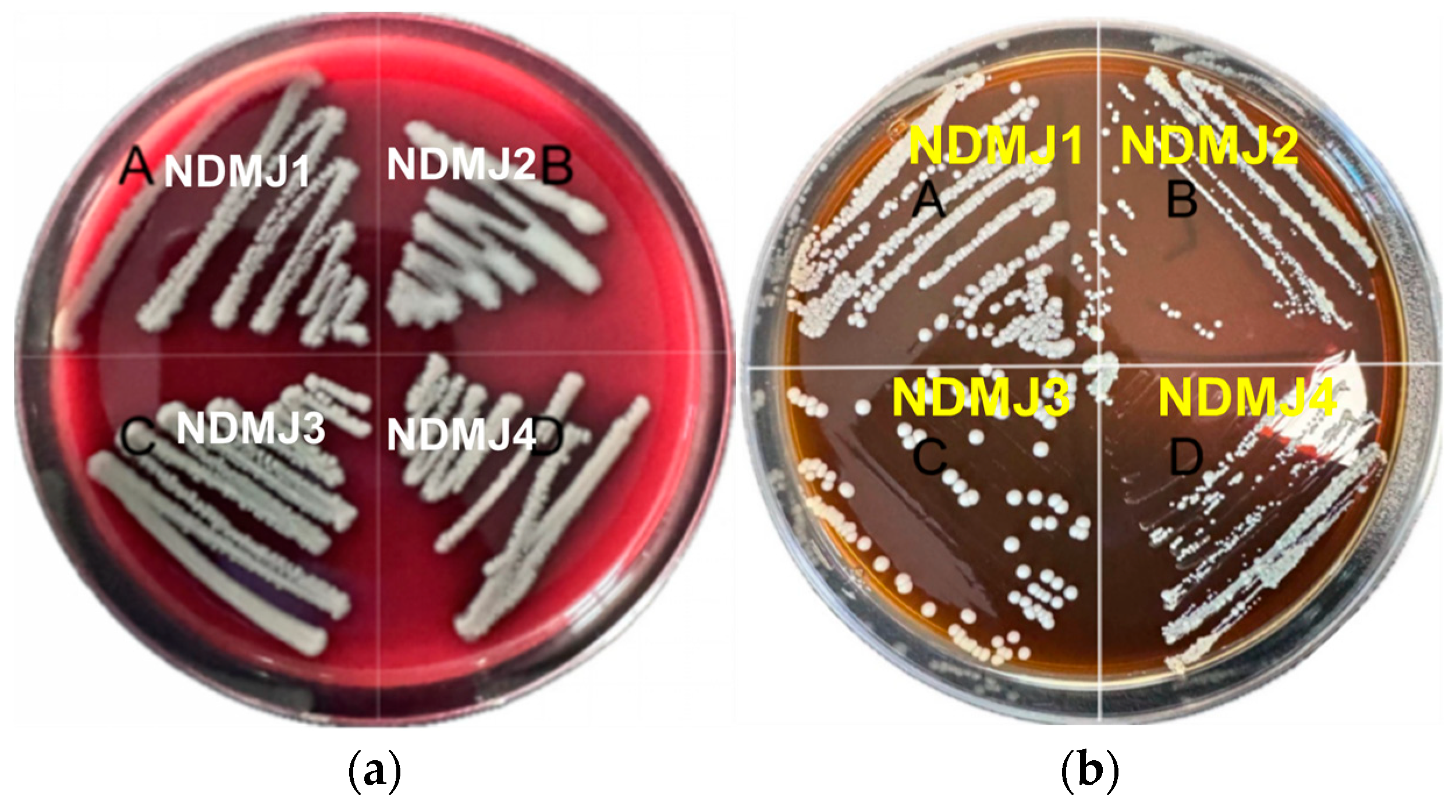
3.1. Isolation and Identification of NDMJ
3.2. Identification of NDMJ Strains
3.3. In Vitro Antibacterial Assay
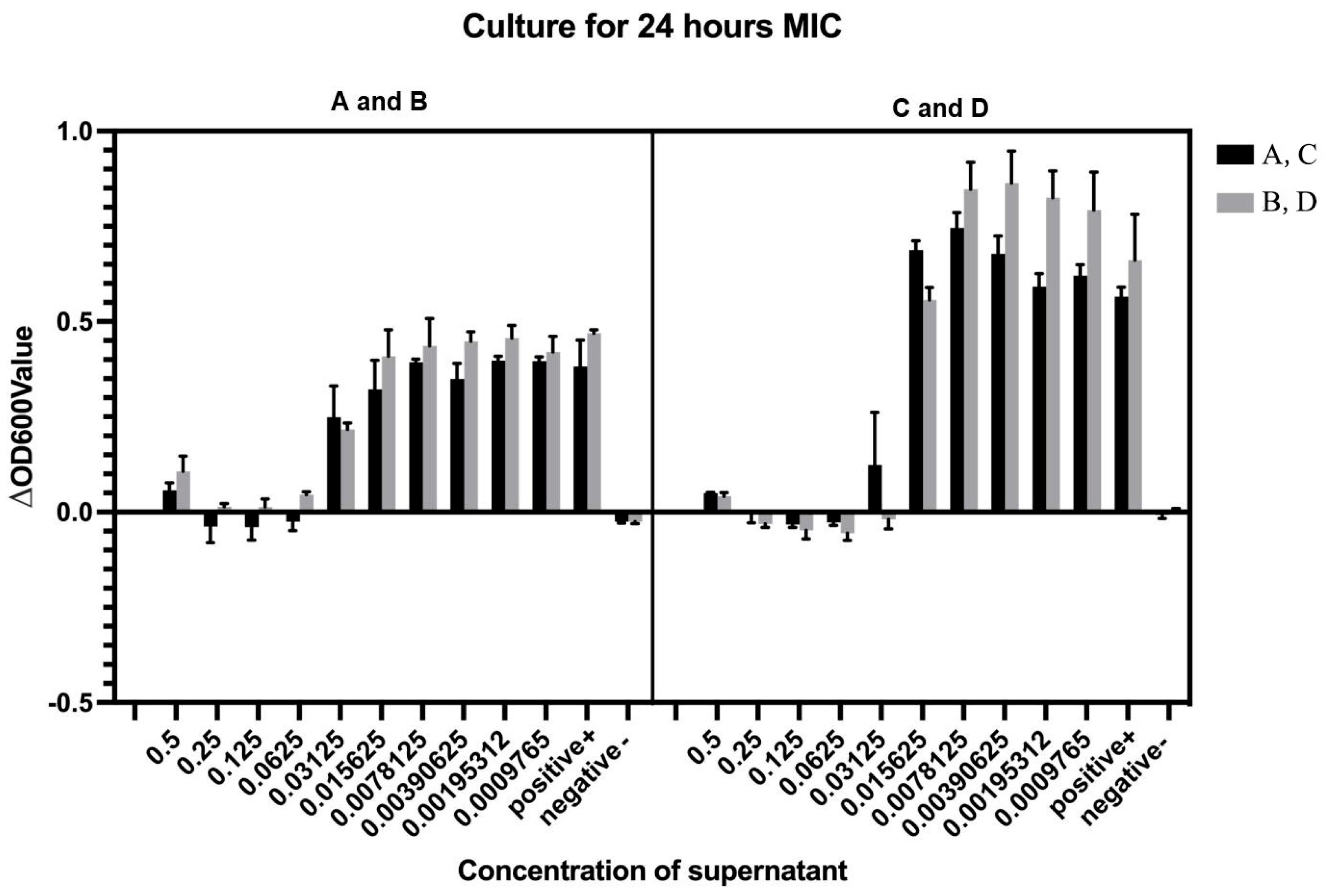
3.4. Determination of Minimum Inhibitory Concentration (MIC) Against Vibrio Parahaemolyticus V-25
3.5. Hemolytic Activity and Biofilm Formation Assessment of NDMJ Isolates
3.6. Antimicrobial Susceptibility Profile of NDMJ Isolates
3.7. Stress Tolerance of NDMJ Isolates
3.7.1. Bile Salt Tolerance
3.7.2. Acid Tolerance
3.7.3. Thermal Tolerance
3.8. Adhesion-Related Properties of NDMJ Isolates
3.8.1. Auto-Aggregation Capacity
3.8.2. Surface Hydrophobicity
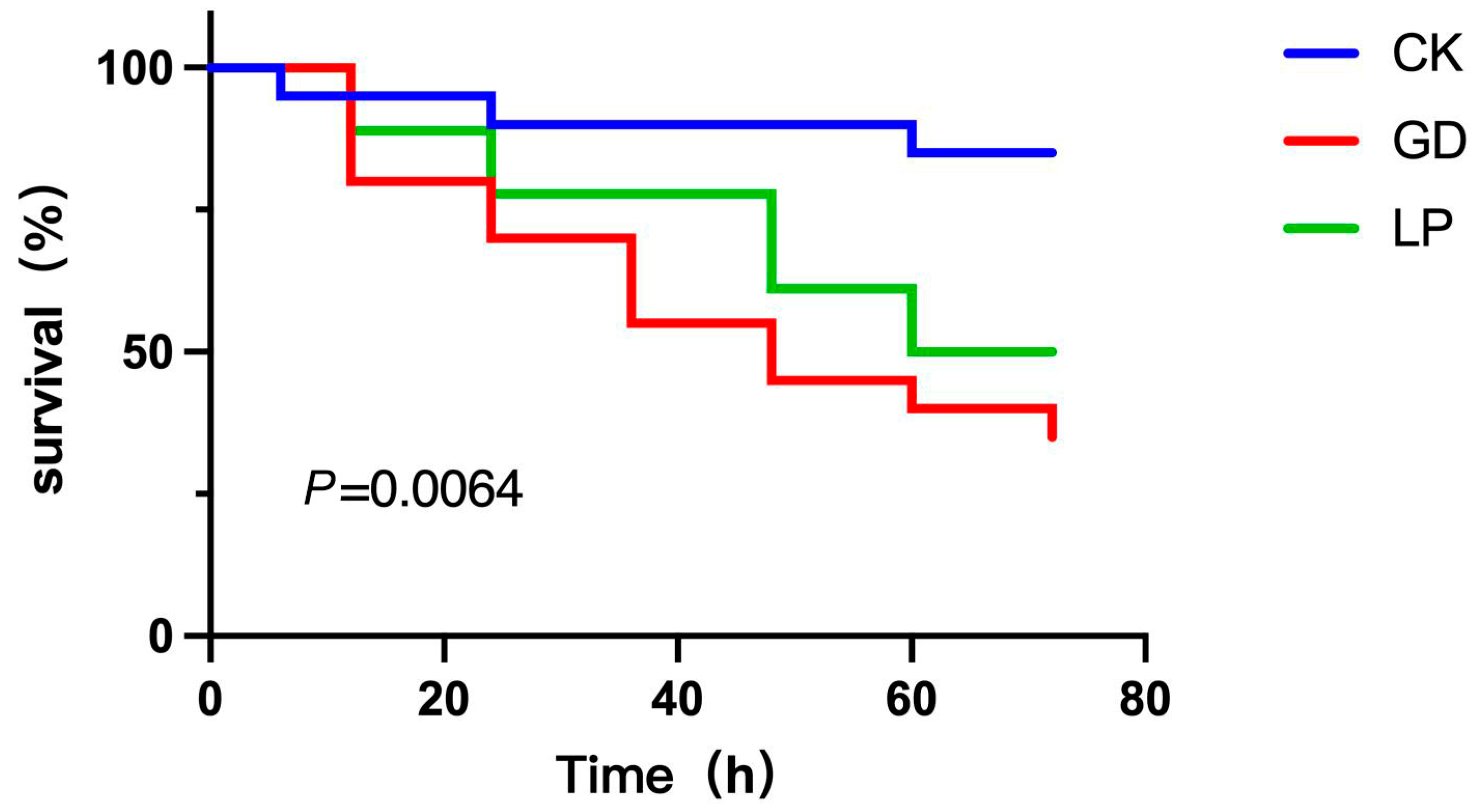
3.9. Vibrio Parahaemolyticus Challenge Assay
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Iehata, S.; Inagaki, T.; Okunishi, S.; Nakano, M.; Tanaka, R.; Maeda, H. Colonization and Probiotic Effects of Lactic Acid Bacteria in the Gut of the Abalone Haliotis Gigantea. Fish. Sci. 2009, 75, 1285–1293. [Google Scholar] [CrossRef]
- Zhang, Z.; Lv, J.; Pan, L.; Zhang, Y. Roles and Applications of Probiotic Lactobacillus Strains. Appl. Microbiol. Biotechnol. 2018, 102, 8135–8143. [Google Scholar] [CrossRef] [PubMed]
- Hadi, J.; Gutierrez, N.; Alfaro, A.; Roberts, R. Use of Probiotic Bacteria to Improve Growth and Survivability of Farmed New Zealand Abalone (Haliotis iris). N. Z. J. Mar. Freshw. Res. 2014, 48, 405–415. [Google Scholar] [CrossRef]
- Wang, H.Z. Isolation of Vibrio Parahaemolyticus from Abalone (Haliotis diversicolor supertexta L.) Postlarvae Associated with Mass Mortalities. Aquaculture 2006, 257, 161–166. [Google Scholar] [CrossRef]
- Qing-Yan, Z.; Shi-Chao, Z.; Xin-Ling, L.I.; Sheng-Nan, H. Classification and Identification of Lactic Acid Bacteria and Application in Food Industry. J. Food Saf. Qual. 2019, 10, 5260–5265. [Google Scholar]
- Nath, D.; Sethi, P.; Kumar, R.; Singh, D. Lactobacillus As a Reservoir of Antibiotic Resistance Genes. In Futuristic Trends in Agriculture Engineering & Food Sciences Volume 3 Book 21; IIP Series: Karnataka, India, 2024; Volume 3, pp. 107–117. ISBN 978-93-5747-435-1. [Google Scholar] [CrossRef]
- Floris, R.; Sanna, G.; Mura, L.; Fiori, M.; Culurgioni, J.; Diciotti, R.; Rizzo, C.; Lo Giudice, A.; Laganà, P.; Fois, N. Isolation and Identification of Bacteria with Surface and Antibacterial Activity from the Gut of Mediterranean Grey Mullets. Microorganisms 2021, 9, 2555. [Google Scholar] [CrossRef]
- Sharifuzzaman, S.M.; Rahman, H.; Austin, D.A.; Austin, B. Properties of Probiotics Kocuria SM1 and Rhodococcus SM2 Isolated from Fish Guts. Probiotics Antimicrob. Proteins 2018, 10, 534–542. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, J.; Lv, M.; Shao, Z.; Hungwe, M.; Wang, J.; Bai, X.; Xie, J.; Wang, Y.; Geng, W. Metabolism Characteristics of Lactic Acid Bacteria and the Expanding Applications in Food Industry. Front. Bioeng. Biotechnol. 2021, 9, 612285. [Google Scholar] [CrossRef]
- Seddik, H.A.; Bendali, F.; Gancel, F.; Fliss, I.; Spano, G.; Drider, D. LP and Its Probiotic and Food Potentialities. Probiotics Antimicrob. Proteins 2017, 9, 111–122. [Google Scholar] [CrossRef]
- Dave, R.I.; Shah, N.P. Ingredient Supplementation Effects on Viability of Probiotic Bacteria in Yogurt. J. Dairy Sci. 1998, 81, 2804–2816. [Google Scholar] [CrossRef]
- Vinderola, C.G.; Reinheimer, J.A. Lactic Acid Starter and Probiotic Bacteria: A Comparative “In Vitro” Study of Probiotic Characteristics and Biological Barrier Resistance. Food Res. Int. 2003, 36, 895–904. [Google Scholar] [CrossRef]
- Wu, L.; Wang, W.; Wu, Z.; Pan, D.; Zeng, X.; Guo, Y.; Lian, L. Effect of Acid and Alkali Stress on Extracellular Metabolite Profile of LP ATCC 14917. J. Basic Microbiol. 2020, 60, 722–729. [Google Scholar] [CrossRef] [PubMed]
- Ayyash, M.; Abdalla, A.; Alkalbani, N.; Baig, M.; Turner, M.; Liu, S.; Shah, N. Invited Review: Characterization of New Probiotics from Dairy and Nondairy Products-Insights into Acid Tolerance, Bile Metabolism and Tolerance, and Adhesion Capability. J. Dairy Sci. 2021, 104, 8363–8379. [Google Scholar] [CrossRef] [PubMed]
- Verónica, U.; Josep, C. Interactions between Bacteria and Bile Salts in the Gastrointestinal and Hepatobiliary Tracts. Front. Med. 2017, 4, 163. [Google Scholar]
- Llamas-Arriba, M.G.; Puertas, A.I.; Prieto, A.; López, P.; Cobos, M.; Miranda, J.I.; Marieta, C.; Ruas-Madiedo, P.; Dueñas, M.T. Characterization of Dextrans Produced by Lactobacillus Mali CUPV271 and Leuconostoc Carnosum CUPV411. Food Hydrocoll. 2019, 89, 613–622. [Google Scholar] [CrossRef]
- Endo, R.; Hotta, S.; Wakinaka, T.; Mogi, Y.; Watanabe, J. Identification of an Operon and Its Regulator Required for Autoaggregation in Tetragenococcus halophilus. Appl. Environ. Microbiol. 2023, 89, e0145823. [Google Scholar] [CrossRef]
- Bhakdi, S.; Mackman, N.; Menestrina, G.; Gray, L.; Hugo, F.; Seeger, W.; Holland, I.B. The Hemolysin of Escherichia coli. Eur. J. Epidemiol. 1988, 4, 135–143. [Google Scholar] [CrossRef]
- Chalmeau, J.; Monina, N.; Shin, J.; Vieu, C.; Noireaux, V. α-Hemolysin Pore Formation into a Supported Phospholipid Bilayer Using Cell-Free Expression. Biochim. Biophys. Acta 2011, 1808, 271–278. [Google Scholar] [CrossRef]
- Palela, M.; Giol, E.D.; Amzuta, A.; Ologu, O.G.; Stan, R.C. Fever Temperatures Impair Hemolysis Caused by Strains of Escherichia coli and Staphylococcus aureus. Heliyon 2022, 8, e08958. [Google Scholar] [CrossRef]
- Elliott, S.J.; Srinivas, S.; Albert, M.J.; Alam, K.; Robins-Browne, R.M.; Gunzburg, S.T.; Mee, B.J.; Chang, B.J. Characterization of the Roles of Hemolysin and Other Toxins in Enteropathy Caused by Alpha-Hemolytic Escherichia coli Linked to Human Diarrhea. Infect. Immun. 1998, 66, 2040–2051. [Google Scholar] [CrossRef]
- Joseph, A.; Philip, R. Immunocompetence of Penaeus Monodon under Acute Salinity Stress and Pathogenicity of Vibrio Harveyi with Respect to Ambient Salinity. Fish Shellfish Immunol. 2020, 106, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Feng, Z.; Wang, J. Vibrio Vulnificus Hemolysin: Biological Activity, Regulation of VvhA Expression, and Role in Pathogenesis. Front. Immunol. 2020, 11, 599439. [Google Scholar] [CrossRef]
- Greene, C.; Vadlamudi, G.; Newton, D.; Foxman, B.; Xi, C. The Influence of Biofilm Formation and Multidrug Resistance on Environmental Survival of Clinical and Environmental Isolates of Acinetobacter Baumannii. Am. J. Infect. Control 2016, 44, e65–e71. [Google Scholar] [CrossRef] [PubMed]
- Rather, M.A.; Gupta, K.; Mandal, M. Microbial Biofilm: Formation, Architecture, Antibiotic Resistance, and Control Strategies. Braz. J. Microbiol. 2021, 52, 1701–1718. [Google Scholar] [CrossRef]
- Sauer, K.; Stoodley, P.; Goeres, D.M.; Hall-Stoodley, L.; Burmølle, M.; Stewart, P.S.; Bjarnsholt, T. The Biofilm Life Cycle: Expanding the Conceptual Model of Biofilm Formation. Nat. Rev. Microbiol. 2022, 20, 608–620. [Google Scholar] [CrossRef] [PubMed]
- Swanson, K.S.; Allenspach, K.; Amos, G.; Auchtung, T.A.; Bassett, S.A.; Bjørnvad, C.R.; Everaert, N.; Martín-Orúe, S.M.; Ricke, S.C.; Ryan, E.P.; et al. Use of Biotics in Animals: Impact on Nutrition, Health, and Food Production. J. Anim. Sci. 2025, 103, skaf061. [Google Scholar] [CrossRef]
- Devadas, S.; Zakaria, Z.; Shariff, M.; Bhassu, S.; Karim, M.; Natrah, I. Methodologies and Standards for Monitoring Antimicrobial Use and Antimicrobial Resistance in Shrimp Aquaculture. Aquaculture 2024, 579, 740216. [Google Scholar] [CrossRef]
- Li, Z.; Li, C.; Lin, F.; Yan, L.; Wu, H.; Zhou, H.; Guo, Q.; Lin, B.; Xie, B.; Xu, Y.; et al. Duck Compound Probiotics Fermented Diet Alters the Growth Performance by Shaping the Gut Morphology, Microbiota and Metabolism. Poult. Sci. 2024, 103, 103647. [Google Scholar] [CrossRef]
- Li, Z.; Guo, Q.; Lin, F.; Li, C.; Yan, L.; Zhou, H.; Huang, Y.; Lin, B.; Xie, B.; Lin, Z.; et al. Lactobacillus Plantarum Supernatant Inhibits Growth of Riemerella Anatipestifer and Mediates Intestinal Antimicrobial Defense in Muscovy Ducks. Poult. Sci. 2024, 103, 103216. [Google Scholar] [CrossRef]
- Wu, S.; Luo, Y.; Wei, F.; Li, Y.; Fan, J.; Chen, Y.; Zhang, W.; Li, X.; Xu, Y.; Chen, Z.; et al. Lactic Acid Bacteria Target NF-ΚB Signaling to Alleviate Gastric Inflammation. Food Funct. 2025, 16, 3101–3119. [Google Scholar] [CrossRef]
- Dubief, B.; Nunes, F.; Basuyaux, O.; Richard, O.; Paillard, C. Differential Resistance to Bacterial Infection of Two Populations of the European Abalone Haliotis Tuberculata against the Bacterium Vibrio Harveyi. Fish Shellfish Immunol. 2016, 53, 90–91. [Google Scholar] [CrossRef]
- Guarin, M.; Ny, A.; De Croze, N.; Maes, J.; Léonard, M.; Annaert, P.; de Witte, P.A.M. Pharmacokinetics in Zebrafish Embryos (ZFE) Following Immersion and Intrayolk Administration: A Fluorescence-Based Analysis. Pharmaceuticals 2021, 14, 576. [Google Scholar] [CrossRef] [PubMed]
- Watson, T.R.; Bruce, T.J.; Ma, J.; Cain, K.D. Comparison of Injection and Immersion Challenges of Renibacterium Salmoninarum Strains in Rainbow Trout. J. Aquat. Anim. Health 2023, 35, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Phudinsai, P.; Wangkahart, E. Effect of the MONTANIDETM IMS 1312 Adjuvant on the Innate and Adaptive Immune Responses of Nile Tilapia (Oreochromis niloticus) against Streptococcusagalactiae through Immersion Vaccination. Fish Shellfish Immunol. 2024, 155, 110012. [Google Scholar] [CrossRef]
- Yao, T.; Lu, J.; Bai, C.; Xie, Z.; Ye, L. The Enhanced Immune Protection in Small Abalone Haliotis Diversicolor Against a Secondary Infection with Vibrio Harveyi. Front. Immunol. 2021, 12, 685896. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | NDMJ-1 | NDMJ-2 | NDMJ-3 | NDMJ-4 |
|---|---|---|---|---|
| Esculin | + | + | + | + |
| Maltose | + | + | + | + |
| Salicin fermentation tube | + | + | + | + |
| Sorbitol fermentation tube | − | + | + | + |
| Cellobiose | + | + | + | + |
| Lactose | + | + | + | + |
| Raffinose | + | + | + | + |
| Sucrose | + | + | + | + |
| Inulin | + | + | + | + |
| Antibiotic Class | Drug Content (μg/Tablet) | Probiotics No. | |||
|---|---|---|---|---|---|
| NDMJ-1 | NDMJ-2 | NDMJ-3 | NDMJ-4 | ||
| β-Lactams | Erythromycin (15) | R | S | S | S |
| Minocycline (30) | R | S | S | R | |
| Doxycycline (30) | S | R | R | R | |
| Tetracycline (30) | R | R | R | R | |
| Neomycin (30) | R | R | R | R | |
| Kanamycin (30) | R | R | R | R | |
| Gentamycin (10) | S | S | R | R | |
| Amikacin (30) | R | S | S | S | |
| Cefoperazone (75) | R | S | S | S | |
| Ceftriaxone (30) | R | R | S | S | |
| Ceftazidime (30) | S | S | R | S | |
| Cefuroxime (30) | S | S | S | S | |
| Aminoglycosides | Cefuroxime (30) | R | S | S | S |
| Cefazolin (30) | S | S | R | S | |
| Cephalexin (30) | R | R | R | R | |
| Piperacillin (100) | S | S | S | S | |
| Macrolides | Carbenicillin (100) | S | S | S | S |
| Tetracyclines | Ampicillin (10) | S | S | S | S |
| Oxacillin (1) | R | R | R | R | |
| Penicillin (10) | R | R | S | S | |
| Treat 4 h | Live Bacteria Concentration (CFU/mL) at Different Bile Salt Contents | |||
|---|---|---|---|---|
| Initial Concentration (0.0%) | 0.1% | 0.3% | 0.5% | |
| NDMJ-1 | 4.36 × 104 ± 3.27 × 102 | 3.29 × 104 ± 1.00 × 102 | 1.97 × 104 ± 1.53 × 102 | 1.26 × 104 ± 1.00 × 102 |
| NDMJ-2 | 4.80 × 104 ± 1.53 × 102 | 5.30 × 104 ± 1.00 × 102 | 1.27 × 103 ± 2.50 × 101 | 8.40 × 103 ± 5.00 × 101 |
| NDMJ-3 | 4.80 × 104 ± 1.53 × 102 | 4.21 × 104 ± 1.00 × 102 | 8.70 × 103 ± 5.00 × 101 | 7.00 × 103 ± 5.00 × 101 |
| NDMJ-4 | 5.11 × 104 ± 3.61 × 102 | 4.42 × 104 ± 2.50 × 102 | 2.78 × 104 ± 2.00 × 102 | 1.35 × 104 ± 1.00 × 102 |
| Processing 4 h | Live Bacteria Concentration at Different pH (CFU/mL) | |||||
|---|---|---|---|---|---|---|
| (pH = 6.5) | pH = 1.0 | pH = 3.0 | pH = 5.0 | pH = 7.0 | pH = 9.0 | |
| NDMJ-1 | 7.07 × 105 ± 1.55 × 104 | 3.51 × 103 ± 9.00 × 101 | 8.50 × 104 ± 9.50 × 102 | 5.20 × 105 ± 1.00 × 102 | 8.03 × 105 ± 6.24 × 103 | 4.20 × 104 ± 1.00 × 103 |
| NDMJ-2 | 4.90 × 106 ± 1.00 × 105 | 4.10 × 104 ± 1.00 × 102 | 5.51 × 105 ± 8.54 × 103 | 1.05 × 106 ± 5.00 × 102 | 7.10 × 106 ± 1.00 × 103 | 7.70 × 105 ± 1.00 × 102 |
| NDMJ-3 | 6.33 × 105 ± 1.55 × 104 | 7.90 × 103 ± 9.00 × 101 | 4.62 × 104 ± 7.23 × 102 | 3.20 × 105 ± 1.00× 102 | 4.60 × 105 ± 1.00 × 102 | 1.15 × 105 ± 5.00 × 102 |
| NDMJ-4 | 3.10 × 106 ± 1.00 × 105 | 5.10 × 103 ± 1.00 × 102 | 5.61 × 104 ± 8.08 × 102 | 9.10 × 104 ± 1.00 × 102 | 2.05 × 106 ± 5.00 × 104 | 1.05 × 106 ± 5.00 × 103 |
| Treat 30 min | Live Bacteria Concentration at Different Temperatures (CFU/mL) | ||||
|---|---|---|---|---|---|
| 35 °C | 45 °C | 55 °C | 65 °C | 75 °C | |
| NDMJ-1 | 2.41 × 105 ± 3.61 × 103 | 2.50 × 105 ± 2.00 × 103 | 4.12 × 102 ± 7.64 × 100 | 0 ± 0 | 0 ± 0 |
| NDMJ-2 | 4.37 × 106 ± 3.61 × 102 | 4.41 × 106 ± 1.53 × 102 | 3.45 × 103 ± 5.00 × 101 | 5.00 × 101 ± 1.00 × 100 | 0 ± 0 |
| NDMJ-3 | 2.08 × 106 ± 1.53 × 102 | 1.92 × 106 ± 1.00 × 102 | 4.50 × 102 ± 1.00 × 101 | 0 ± 0 | 0 ± 0 |
| NDMJ-4 | 1.86 × 106 ± 1.00 × 102 | 1.62 × 106 ± 1.00 × 102 | 3.12 × 102 ± 7.64 × 100 | 2.00 × 101 ± 1.00 × 100 | 0 ± 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ke, L.; Huang, C.; Peng, S.; Zhao, M.; Lin, F.; Li, Z. Potential of LP as a Biocontrol Agent for Vibriosis in Abalone Farming. Microorganisms 2025, 13, 1554. https://doi.org/10.3390/microorganisms13071554
Ke L, Huang C, Peng S, Zhao M, Lin F, Li Z. Potential of LP as a Biocontrol Agent for Vibriosis in Abalone Farming. Microorganisms. 2025; 13(7):1554. https://doi.org/10.3390/microorganisms13071554
Chicago/Turabian StyleKe, Ling, Chenyu Huang, Song Peng, Mengshi Zhao, Fengqiang Lin, and Zhaolong Li. 2025. "Potential of LP as a Biocontrol Agent for Vibriosis in Abalone Farming" Microorganisms 13, no. 7: 1554. https://doi.org/10.3390/microorganisms13071554
APA StyleKe, L., Huang, C., Peng, S., Zhao, M., Lin, F., & Li, Z. (2025). Potential of LP as a Biocontrol Agent for Vibriosis in Abalone Farming. Microorganisms, 13(7), 1554. https://doi.org/10.3390/microorganisms13071554