
Polyphasic Characterization and Genomic Insights into an Aerobic Denitrifying Bacterium, Shewanella zhuhaiensis sp. nov., Isolated from a Tidal Flat Sediment

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolation
2.2. Morphological, Physiological, and Biochemical Characteristics
2.3. The 16S rRNA Gene Sequencing and Phylogenetic Analysis
2.4. Whole-Genome Sequencing, Phylogenomic, and Comparative Analyses
2.5. Accession Numbers
3. Results and Discussion
3.1. Physiological Characterization
3.2. Chemotaxonomic Analysis
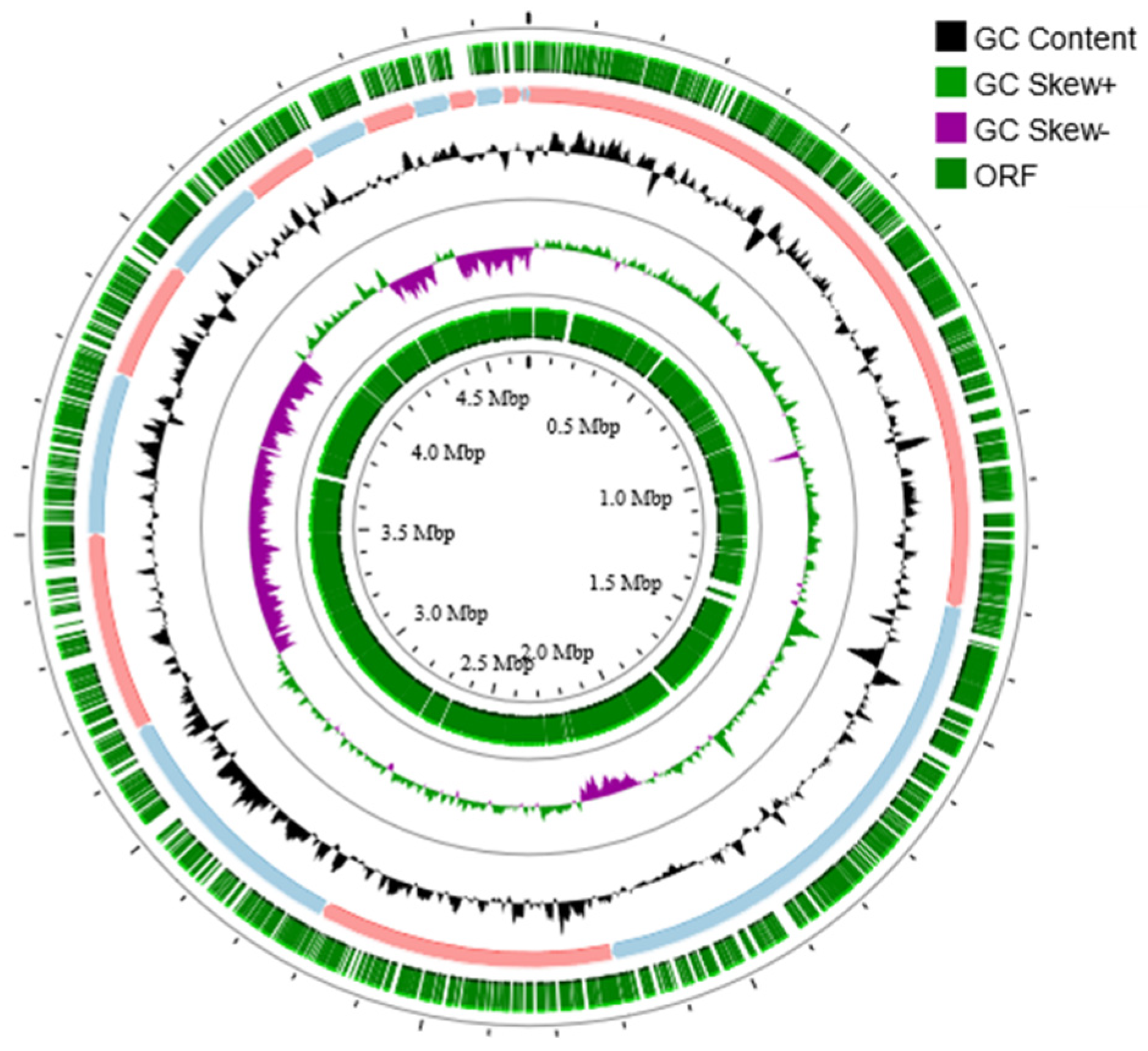
3.3. Genome Structural Features
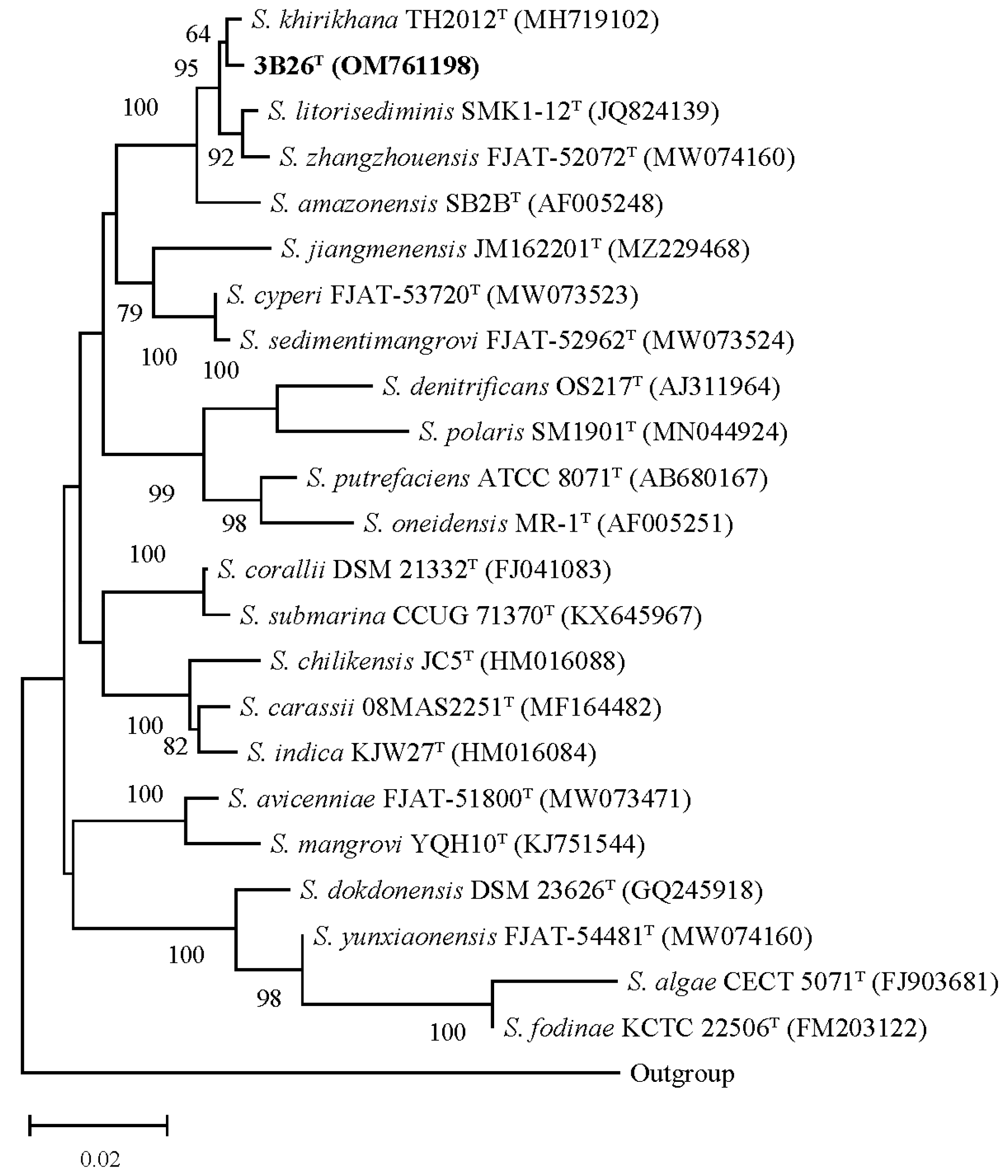
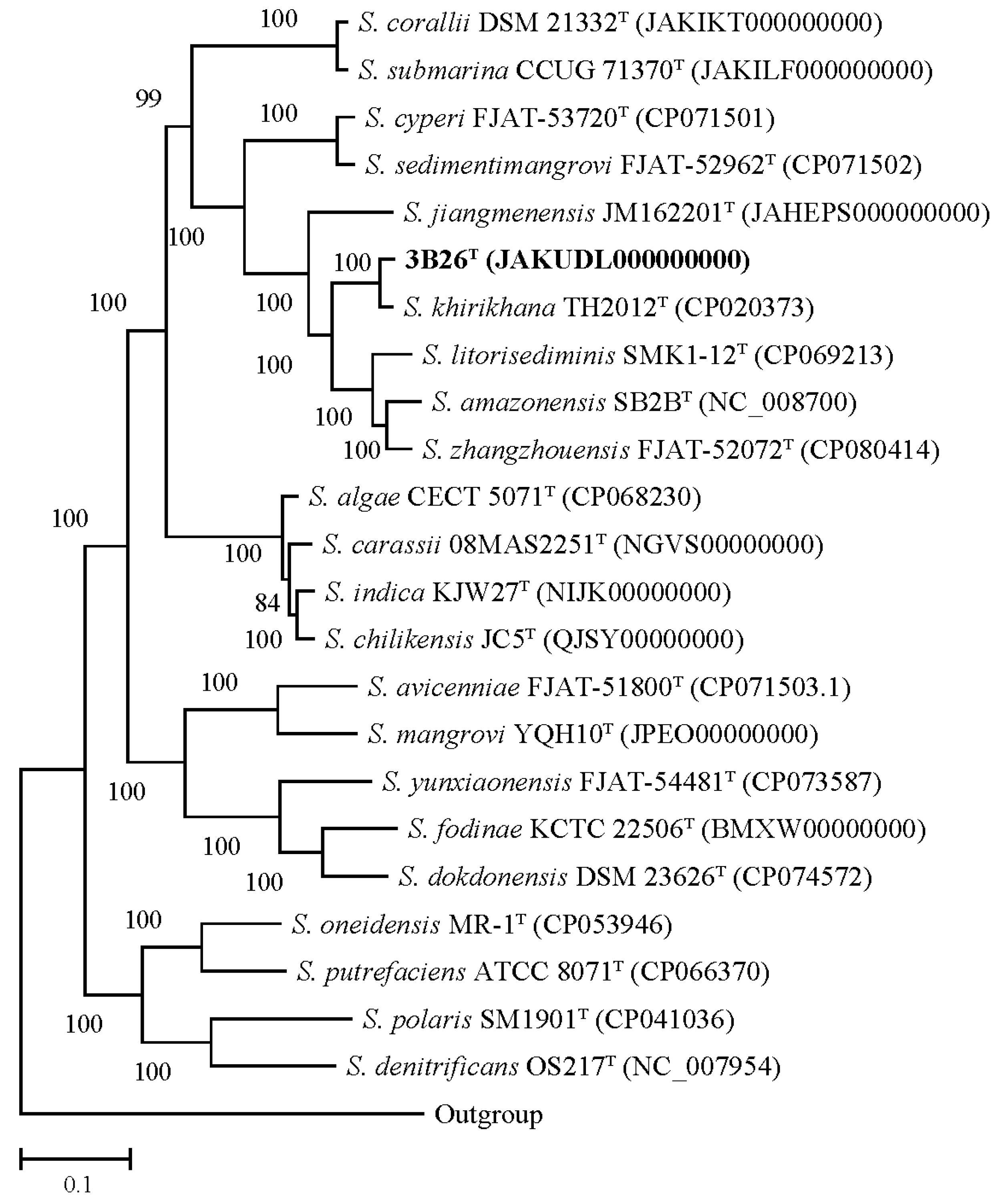
3.4. Phylogenetic and Taxonogenomic Analyses
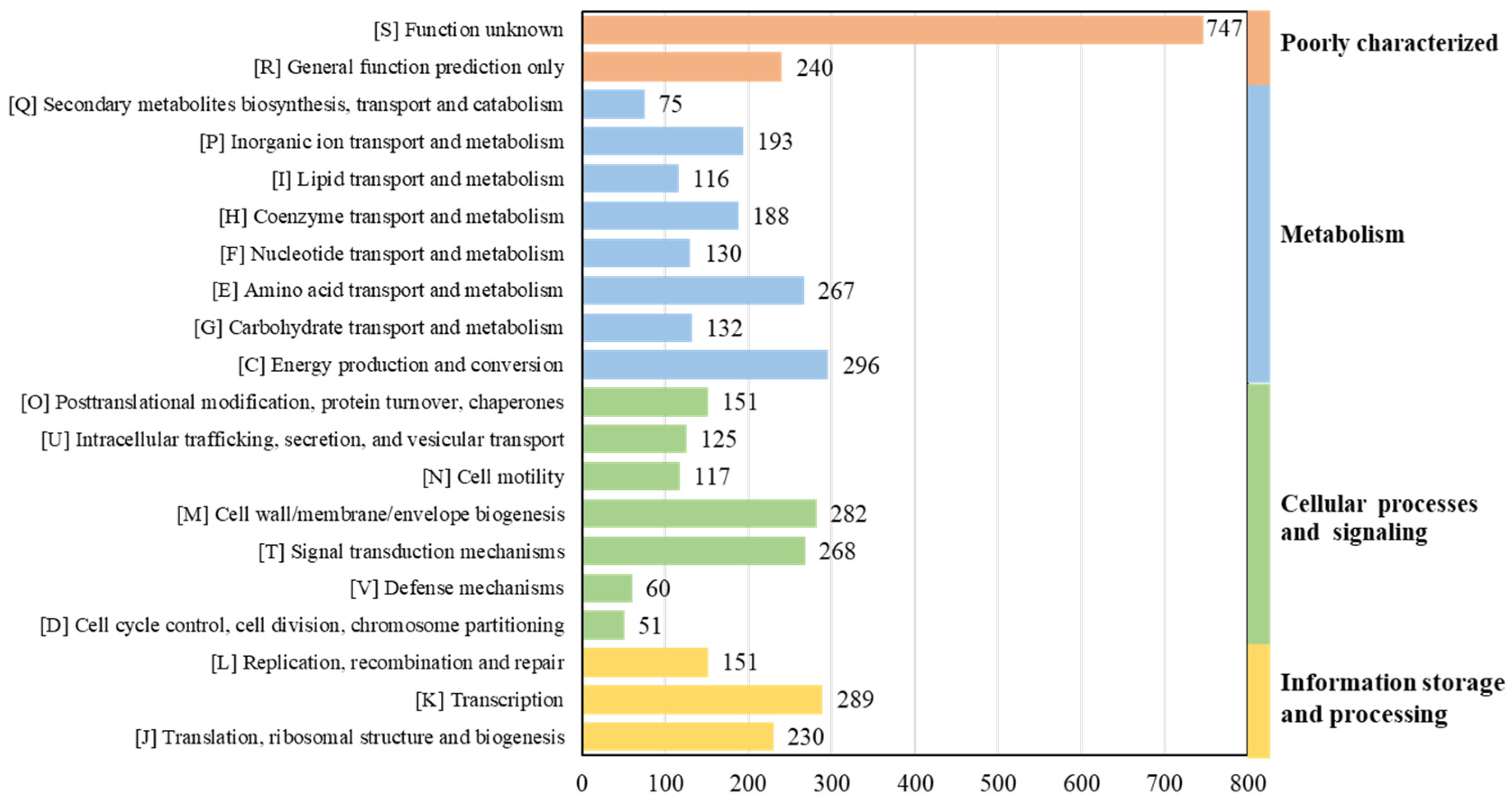
3.5. Genomic Functional Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethical Statement
Abbreviations
References
- Ivanova, E.P.; Flavier, S.; Christen, R. Phylogenetic relationships among marine Alteromonas-like proteobacteria: Emended description of the family Alteromonadaceae and proposal of Pseudoalteromonadaceae fam. nov., Colwelliaceae fam. nov., Shewanellaceae fam. nov., Moritellaceae fam. nov., Ferrimonadaceae fam. nov., Idiomarinaceae fam. nov. and Psychromonadaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2004, 54, 1773–1788. [Google Scholar]
- MacDonell, M.T.; Colwell, R.R. Phylogeny of the Vibrionaceae, and recommendation for two new genera, Listonella and Shewanella. Syst. Appl. Microbiol. 1985, 6, 171–182. [Google Scholar] [CrossRef]
- Martín-Rodríguez, A.J.; Meier-Kolthoff, J.P. Whole genome-based taxonomy of Shewanella and Parashewanella. Int. J. Syst. Evol. Microbiol. 2022, 72, 005438. [Google Scholar] [CrossRef]
- Parte, A.C.; Sardà Carbasse, J.; Meier-Kolthoff, J.P.; Reimer, L.C.; Göker, M. List of Prokaryotic names with Standing in Nomenclature (LPSN) moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, O.N.; Méjean, V.; Iobbi-Nivol, C. The Shewanella genus: Ubiquitous organisms sustaining and preserving aquatic ecosystems. FEMS Microbiol. Rev. 2020, 44, 155–170. [Google Scholar] [CrossRef]
- Yu, K.; Huang, Z.; Xiao, Y.; Wang, D. Shewanella infection in humans: Epidemiology, clinical features and pathogenicity. Virulence 2022, 13, 1515–1532. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Pei, T.; Du, J.; Chao, M.; Deng, M.R.; Zhu, H. Roseibium litorale sp. nov., isolated from a tidal flat sediment and proposal for the reclassification of Labrenzia polysiphoniae as Roseibium polysiphoniae comb. nov. Int. J. Syst. Evol. Microbiol. 2021, 71, 004634. [Google Scholar] [CrossRef] [PubMed]
- Venkateswaran, K.; Dollhopf, M.E.; Aller, R.; Stackebrandt, E.; Nealson, K.H. Shewanella amazonensis sp. nov., a novel metal-reducing facultative anaerobe from Amazonian shelf muds. Int. J. Syst. Bacteriol. 1998, 48, 965–972. [Google Scholar] [CrossRef]
- Lee, M.H.; Yoon, J.H. Shewanella litorisediminis sp. nov., a gammaproteobacterium isolated from a tidal flat sediment. Antonie Leeuwenhoek 2012, 102, 591–599. [Google Scholar] [CrossRef]
- Song, Y.; Li, A.; Xu, S.; Zhang, M.; Yao, Q.; Ma, L.; Zhu, H. Shewanella jiangmenensis sp. nov., isolated from aquaculture water. Arch. Microbiol. 2022, 204, 198. [Google Scholar] [CrossRef]
- Prachumwat, A.; Wechprasit, P.; Srisala, J.; Kriangsaksri, R.; Flegel, T.W.; Thitamadee, S.; Sritunyalucksana, K. Shewanella khirikhana sp. nov.-a shrimp pathogen isolated from a cultivation pond exhibiting early mortality syndrome. Microb. Biotechnol. 2020, 13, 781–795. [Google Scholar] [CrossRef]
- Liu, G.H.; Yang, S.; Narsing Rao, M.P.; Han, S.; Xie, C.J.; Alwathnani, H.A.; Herzberg, M.; Rensing, C.; Zhou, S.G. Isolation and genomics of ten novel Shewanella species from mangrove wetland. Int. J. Syst. Evol. Microbiol. 2023, 73, 005929. [Google Scholar] [CrossRef] [PubMed]
- Minnikin, D.E.; O’Donnell, A.G.; Goodfellow, M.; Alderson, G.; Athalye, M.; Schaal, A.; Parlett, J.H. An integrated procedure for the extraction of bacterial isoprenoid quinones and polar lipids. J. Microbiol. Methods 1984, 2, 233–241. [Google Scholar] [CrossRef]
- Collins, M.D.; Pirouz, T.; Goodfellow, M.; Minnikin, D.E. Distribution of menaquinones in actinomycetes and corynebacteria. J. Gen. Microbiol. 1977, 100, 221–230. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.-H.; Ha, S.-M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef]
- Rzhetsky, A.; Nei, M. Theoretical foundation of the minimum-evolution method of phylogenetic inference. Mol. Biol. Evol. 1993, 10, 1073–1095. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Auch, A.F.; von Jan, M.; Klenk, H.-P.; Göker, M. Digital DNA-DNA hybridization for microbial species delineation by means of genome-to-genome sequence comparison. Stand. Genom. Sci. 2010, 2, 117–134. [Google Scholar] [CrossRef] [PubMed]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat. Commun. 2018, 9, 5114. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res. 2014, 42, D206–D214. [Google Scholar] [CrossRef]
- Cantalapiedra, C.P.; Hernández-Plaza, A.; Letunic, I.; Bork, P.; Huerta-Cepas, J. eggNOG-mapper v2: Functional annotation, orthology assignments, and domain prediction at the metagenomic scale. Mol. Biol. Evol. 2021, 38, 5825–5829. [Google Scholar] [CrossRef]
- Na, S.-I.; Kim, Y.O.; Yoon, S.-H.; Ha, S.-M.; Baek, I.; Chun, J. UBCG: Up-to-date bacterial core gene set and pipeline for phylogenomic tree reconstruction. J. Microbiol. 2018, 56, 280–285. [Google Scholar] [CrossRef]
- Chun, J.; Oren, A.; Ventosa, A.; Christensen, H.; Arahal, D.R.; da Costa, M.S.; Rooney, A.P.; Yi, H.; Xu, X.-W.; De Meyer, S.; et al. Proposed minimal standards for the use of genome data for the taxonomy of prokaryotes. Int. J. Syst. Evol. Microbiol. 2018, 68, 461–466. [Google Scholar] [CrossRef]
- Wayne, L.G.; Brenner, D.J.; Colwell, R.R.; Grimont, P.A.D.; Kandler, O.; Krichevsky, M.I.; Moore, L.H.; Moore, W.E.C.; Murray, R.G.E.; Stackebrandt, E.; et al. Report of the Ad Hoc committee on reconciliation of approaches to bacterial systematics. Int. J. Syst. Bacteriol. 1987, 37, 463–464. [Google Scholar] [CrossRef]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Zhou, J.; Zhou, J.; Hu, M.; Zhang, Q.; Kong, N.; Ren, H.; Liang, L.; Yue, J. Genome-based classification of Burkholderia cepacia complex provides new insight into its taxonomic status. Biol. Direct 2020, 15, 6. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Li, K.; Zhang, Y.; Wang, Y.; Fu, L.; Xiao, Y.; Tang, X.; Gao, J. New insights into the threshold values of multi-locus sequence analysis, average nucleotide identity and digital DNA-DNA hybridization in delineating Streptomyces species. Front. Microbiol. 2022, 13, 910277. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lai, Q.; Du, J.; Shao, Z. Genetic diversity and population structure of the Bacillus cereus group bacteria from diverse marine environments. Sci. Rep. 2017, 7, 689. [Google Scholar] [CrossRef]
- Moi, I.M.; Leow, A.T.C.; Ali, M.S.M.; Rahman, R.; Salleh, A.B.; Sabri, S. Polyunsaturated fatty acids in marine bacteria and strategies to enhance their production. Appl. Microbiol. Biotechnol. 2018, 102, 5811–5826. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, F. Insights on nitrate respiration by Shewanella. Front. Mar. Sci. 2015, 1, 80. [Google Scholar] [CrossRef]
- Simpson, P.J.L.; Richardson, D.J.; Codd, R. The periplasmic nitrate reductase in Shewanella: The resolution, distribution and functional implications of two NAP Isoforms, NapEDABC and NapDAGHB. Microbiology 2010, 156, 302–312. [Google Scholar] [CrossRef]
- Wu, G.; Li, N.; Mao, Y.; Zhou, G.; Gao, H. Endogenous generation of hydrogen sulfide and its regulation in Shewanella oneidensis. Front. Microbiol. 2015, 6, 374. [Google Scholar] [CrossRef]
- Baek, J.H.; Lee, S.Y. Transcriptome analysis of phosphate starvation response in Escherichia coli. J. Microbiol. Biotechnol. 2007, 17, 244–252. [Google Scholar]
- Yang, Y.L.; Goldrick, D.; Hong, J.S. Identification of the products and nucleotide sequences of two regulatory genes involved in the exogenous induction of phosphoglycerate transport in Salmonella typhimurium. J. Bacteriol. 1988, 170, 4299–4303. [Google Scholar] [CrossRef] [PubMed]
- Coon, S.L.; Kotob, S.; Jarvis, B.B.; Wang, S.; Fuqua, W.C.; Weiner, R.M. Homogentisic acid is the product of MelA, which mediates melanogenesis in the marine bacterium Shewanella colwelliana D. Appl. Environ. Microbiol. 1994, 60, 3006–3010. [Google Scholar] [CrossRef] [PubMed]
- Kuttan, S.P.; Abdulaziz, A.; Chekidhenkuzhiyil, J.; Raj, D.; Mohan, M.; Athiyanathil, S. Characterization of pyomelanin secreted by Shewanella sp. and their application in metal recovery. Environ. Sci. Pollut. Res. Int. 2023, 30, 6705–6715. [Google Scholar] [CrossRef] [PubMed]
- Verma, H.; Bajaj, A.; Kumar, R.; Kaur, J.; Anand, S.; Nayyar, N.; Puri, A.; Singh, Y.; Khurana, J.P.; Lal, R. Genome organization of Sphingobium indicum B90A: An archetypal hexachlorocyclohexane (HCH) degrading genotype. Genome Biol. Evol. 2017, 9, 2191–2197. [Google Scholar] [CrossRef]
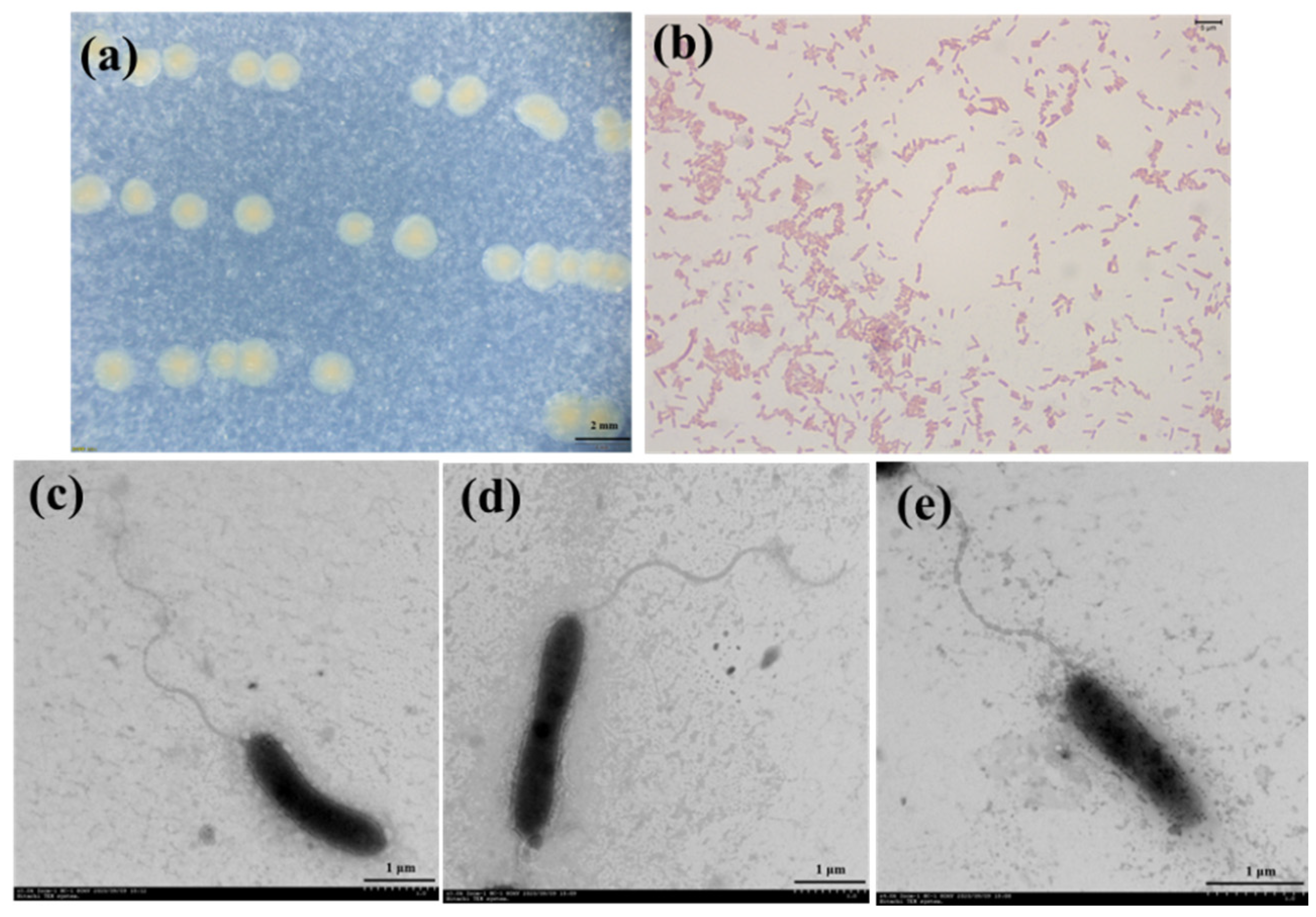

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| Colony color | Light-yellow | Yellowish | Orange-red | Beige to pinkish | Orange yellow | nd |
| Cellular width (μm) | 0.4–0.6 | 0.5–0.6 | 0.6–1.0 | 0.4–0.7 | 0.3–0.7 | 0.4–0.6 |
| Cellular length (μm) | 2.0–2.8 | 1.7–2.4 | 1.7–3.2 | 2.0–3.0 | 0.9–5.0 | 1.4–2.3 |
| Motility | + | + | + | + | - | + |
| Growth of | ||||||
| Temperature range (°C) | 10–40 | 25–37 | 20–40 | 4–45 | 10–40 | 10–40 |
| Optimal temperature (°C) | 30 | 30 | 30 | 37 | 30–37 | 30 |
| pH range | 4.0–9.0 | 6.5–9.5 | 6.0–9.0 | 6.0–9.0 | 5.0–8.0 | 4.0–10.0 |
| Optimal pH | 7.0 | 7.0–7.5 | 7.0 | 7.0–8.0 | 7.0–8.0 | 7.0–8.0 |
| NaCl range (w/v, %) | 0–5.0 | 0.5–5.5 | 0–6.0 | 0–3.0 | 0–6.0 | 0–6.0 |
| Optimal NaCl (w/v, %) | 1.0 | 1.5–2.0 | 2.0 | 1.0 | 2.0 | 0–1.0 |
| Enzymatic activity | ||||||
| α-Glucosidase | - | nd | - | - | + | - |
| Casein hydrolysis | - | + | nd | + | + | + |
| Reduction of nitrate to nitrite | + | + | - | + | + | + |
| Denitrification | - | - | - | + | + | - |
| D-glucose fermentation | - | - | - | + | + | + |
| Utilization of | ||||||
| D-Glucose | + | - | - | + | + | + |
| D-Mannose | + | nd | - | + | + | + |
| Potassium gluconate | - | nd | nd | - | + | + |
| Fatty Acids | 1 | 2 | 3 | 4 | 5 |
|---|---|---|---|---|---|
| C12:0 | 4.7 | 4.0 | 3.6 | 4.8 | 5.9 |
| C13:0 | 2.7 | 3.6 | 2.5 | 2.0 | 3.1 |
| C14:0 | 2.0 | 1.8 | 1.5 | 1.7 | 2.4 |
| C15:0 | 4.1 | nd | 4.7 | 3.5 | 5.0 |
| C16:0 | 11.0 | 7.3 | 10.2 | 12.0 | 11.5 |
| C17:0 | 3.4 | 2.6 | 4.5 | 4.2 | 3.6 |
| C18:0 | 0.8 | nd | 1.3 | 1.8 | 0.6 |
| C11:0 3OH | 0.9 | 1.8 | 1.3 | 1.4 | 1.5 |
| C12:0 3OH | 2.1 | 2.3 | 1.9 | 3.1 | 2.8 |
| C16:0 3OH | 0.6 | nd | 0.6 | 0.7 | tr |
| C15:1 ω6c | tr | 1.2 | 0.9 | tr | 0.7 |
| C17:1 ω6c | 0.9 | 1.9 | 1.4 | 1.0 | 1.5 |
| C17:1 ω8c | 8.5 | 15.1 | 11.1 | 7.5 | 8.5 |
| C18:1 ω9c | 2.0 | 1.5 | 1.4 | 1.5 | 1.5 |
| iso-C13:0 | 3.7 | 4.6 | 3.5 | 3.4 | 4.2 |
| iso-C13:0 3OH | 5.3 | 4.7 | 4.0 | 3.0 | 3.2 |
| iso-C14:0 | 0.6 | 1.0 | 0.5 | tr | 0.7 |
| iso-C15:0 | 16.5 | 18.9 | 15.7 | 16.6 | 8.9 |
| iso-C15:0 3OH | tr | tr | tr | tr | 0.5 |
| iso-C16:0 | 1.3 | 0.6 | 0.9 | 1.0 | 0.6 |
| iso-C17:0 | 2.2 | 0.8 | 1.8 | 2.4 | 1.1 |
| Summed feature 1 # | 0.9 | 1.0 | 0.7 | 0.8 | 1.7 |
| Summed feature 2 # | 0.8 | 0.7 | tr | 0.6 | 2.4 |
| Summed feature 3 # | 14.1 | 17.3 | 15.4 | 14.5 | 18.2 |
| Summed feature 8 # | 4.2 | 4.6 | 3.5 | 5.1 | 4.5 |
| Pathways | K number | Gene | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|---|---|
| Nitrogen metabolism | ||||||||
| Denitrification | K02567 | napA | + | + | + | + | + | + |
| K02568 | napB | + | + | + | + | + | + | |
| K00368 | nirK | + | + | + | + | + | − | |
| K04561 | norB | + | + | + | + | + | + | |
| K02305 | norC | + | + | + | + | + | + | |
| Dissimilatory nitrate reduction | K02567 | napA | + | + | + | + | + | + |
| K02568 | napB | + | + | + | + | + | + | |
| K00362 | nirB | + | − | − | − | − | − | |
| K00363 | nirD | + | − | − | − | − | − | |
| K03385 | nrfA | + | + | + | + | + | + | |
| K15876 | nrfH | + | + | + | + | + | + | |
| Assimilatory nitrate reduction | K00372 | nasA | + | − | − | − | − | − |
| K00360 | nasB | + | − | − | − | − | − | |
| Sulfur metabolism | ||||||||
| Assimilatory sulfate reduction | K00956 | cysN | + | + | + | + | + | + |
| K00957 | cysD | + | + | + | + | + | + | |
| K00955 | cysC | + | + | + | + | + | + | |
| K00390 | cysH | + | + | + | + | + | + | |
| K00380 | cysJ | + | + | + | + | + | + | |
| K00381 | cysI | + | + | + | + | + | + | |
| Phosphorus metabolism | ||||||||
| Two-component system | K02039 | phoU | + | + | + | + | + | + |
| K07636 | phoR | + | + | + | + | + | + | |
| K07657 | phoB | + | + | + | + | + | + | |
| K08478 | pgtC | + | + | + | + | + | + | |
| K08475 | pgtB | + | + | + | + | + | + | |
| K08476 | pgtA | + | + | + | + | + | + | |
| Transporters | K02040 | pstS | + | + | + | + | + | + |
| K02037 | pstC | + | + | + | + | + | + | |
| K02038 | pstA | + | + | + | + | + | + | |
| K02036 | pstB | + | + | + | + | + | + | |
| K03306 | pit | + | + | + | + | + | + | |
| K02044 | phnD | + | + | + | + | + | + | |
| K02042 | phnE | + | − | − | − | − | − | |
| K02041 | phnC | + | + | + | + | + | + | |
| Organic phosphoester hydrolysis | K01077 | phoA | + | + | + | + | + | + |
| K01113 | phoD | + | + | + | + | + | + | |
| K07093 | phoX | + | + | + | + | + | + | |
| K09474 | phoN | − | − | − | − | − | + | |
| Melanin biosynthesis | K00500 | phhA | + | + | + | + | + | + |
| K00832 | tyrB | + | + | + | + | + | + | |
| K00457 | hppD | + | + | + | + | + | + | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Pei, T.; Du, J.; Zhu, H. Polyphasic Characterization and Genomic Insights into an Aerobic Denitrifying Bacterium, Shewanella zhuhaiensis sp. nov., Isolated from a Tidal Flat Sediment. Microorganisms 2023, 11, 2870. https://doi.org/10.3390/microorganisms11122870
Liu Y, Pei T, Du J, Zhu H. Polyphasic Characterization and Genomic Insights into an Aerobic Denitrifying Bacterium, Shewanella zhuhaiensis sp. nov., Isolated from a Tidal Flat Sediment. Microorganisms. 2023; 11(12):2870. https://doi.org/10.3390/microorganisms11122870
Chicago/Turabian StyleLiu, Yang, Tao Pei, Juan Du, and Honghui Zhu. 2023. "Polyphasic Characterization and Genomic Insights into an Aerobic Denitrifying Bacterium, Shewanella zhuhaiensis sp. nov., Isolated from a Tidal Flat Sediment" Microorganisms 11, no. 12: 2870. https://doi.org/10.3390/microorganisms11122870
APA StyleLiu, Y., Pei, T., Du, J., & Zhu, H. (2023). Polyphasic Characterization and Genomic Insights into an Aerobic Denitrifying Bacterium, Shewanella zhuhaiensis sp. nov., Isolated from a Tidal Flat Sediment. Microorganisms, 11(12), 2870. https://doi.org/10.3390/microorganisms11122870