Abstract
Sourdough is one of the oldest starters traditionally used for making baked goods, offering several advantages to the sensory, rheology, and shelf life of final products. The present study investigated, for the first time, the microbiota of spontaneously fermented Maiorca dough samples collected from bakeries located in Sicily (Italy). Four sourdough samples (M1, M2, M3, and M4), were produced using Triticum vulgare Host. var. albidum Koern (Maiorca grain) were subjected to LAB and yeasts isolation and identification at the species level. The in-depth characterization of the lactobacilli population revealed that Lactiplantibacillus plantarum and Levilactobacillus brevis unquestionably dominated the Maiorca sourdough ecosystem. Concerning the yeasts community, high species diversity was found. Saccharomyces cerevisiae and Wickerhamomyces anomalus were the most frequently isolated species. In addition, Torulaspora delbrueckii, Pichia kluyveri, Candida boidinii, and Candida diddensiae were also detected. Investigations on both pro-technological and functional traits of the isolated strains could lead to the selection of starters for the production of baked goods.
1. Introduction
In the last decade, the new trend towards the production and consumption of natural, traditional, and healthy foods helped to drive changes in the formulation of cereal-based products [1]. In fact, both consumers and the industry experienced increasing interest in the formulation of bakery foods using traditional sourdough [2].
Sourdough has been used since ancient times as a biotechnological strategy for the fermentation of cereals. Based on its microbiota, sourdough harbors a very complex microbial consortium [3], mainly resulting from the type of flour and ingredients used [4,5,6,7,8]. In addition, the applied technological parameters consistently affect the sourdough microbiota, which in turn influence the sensory, nutritional, texture, and shelf-life features of baked goods [9]. Notably, flour represents the main source of autochthonous lactic acid bacteria (LAB) in spontaneous sourdough fermentation, whose role is essential to establish a balanced microbial consortium between LAB and yeasts [10,11]. Usually, this microbial consortium consists of yeasts and obligate or facultative heterofermentative lactobacilli [10,12,13,14,15]. Among lactobacilli, strains ascribed to Levilactobacillus brevis, Lactiplantibacillus plantarum, and Fructilactobacillus sanfranciscensis species are frequently isolated from Italian sourdoughs [16,17,18,19]. In addition, other LAB, such as enterococci, lactococci, pediococci, streptococci, as well as members of the Leuconostoc and Weissella genera have been detected [4]. Among yeasts, Saccharomyces cerevisiae, Kazachstania exigua, Kazachstania humilis, Torulaspora delbrueckii, Wickerhamomyces anomalus, and Pichia kudriavzevii commonly dominate the fungal community [20,21,22,23]. During the fermentation process, the yeasts are primarily responsible for leavening whereas the LAB carry out the acidification of the dough and both contribute to the flavor of the resulting bread [21,24]. Although functional and nutritional features of sourdough have been traditionally attributed to the LAB metabolism [25,26], several studies highlighted a link with the activities of the yeast population. Several yeasts, isolated from traditional sourdough samples, exhibited functional properties, such as the ability to produce vitamins [27,28], to exert phytase [29] and antioxidant [30,31] activities as well as probiotic properties [4,23,32,33,34].
One of the new trends in bakery productions is the rediscovery of landraces/historical, which is in line with the increased interest in locally grown crops with particular nutritional properties [35]. Compared to modern varieties, wheat landraces have a better nutritional profile, higher content of phenols, vitamins, minerals, and nutraceutical compounds [36]. Sicily, a region of Southern Italy, claims several durum wheat landraces. Among these, Triticum aestivum L. subsp. aestivum, commonly named Maiorca or Majorca, is one of the durum wheat landraces used in the production of bread, pasta, and cakes until the 1900s. Recently, Maiorca flour was re-discovered and re-employed by local bakers for the distinctive sensory properties generated in the final products [23,36]. As reported by Visioli and co-workers [36], the bread wheat Maiorca showed the highest amount of both high molecular weight and low molecular weight glutenins, compared to other Sicilian durum wheat landraces, such as Margherito, Perciasacchi, Russello, and Timilia. The aforementioned glutenin subunits are important parameters affecting the gluten strength, and therefore, the ability of the proteins to form a tenacious network, promoting extrusion properties. In addition, bread wheat Maiorca exhibited low total gluten proteins [36].
Although the microbial population of many types of sourdough was in-depth investigated, no information is now available about the microbiological profile of the Maiorca sourdough. Therefore, the aim of the present study was to characterize the microbiota of spontaneously fermented Maiorca doughs and to collect detailed information about the composition of both LAB and yeast populations.
2. Materials and Methods
2.1. Sourdough Sampling
Sourdough samples (M1, M2, M3, and M4), produced using Triticum vulgare Host. var. albidum Koern (Maiorca grain), were collected from four artisanal bakeries located in the Sicily region. The characteristics of the sourdough samples are detailed in Table S1. All sourdoughs were propagated by one daily back slopping using the sourdough coming from a previous fermentation without the addition of baker’s yeast or any other starter cultures. Mature sourdough M1, M2, M3, and M4 samples were aseptically collected and transferred to the microbiology laboratory under refrigerated conditions. Three independent samples were taken from each selected bakery from each of three different bread-making runs.
2.2. Microbiological Analysis
Each sourdough sample (25 g) was mixed with 225 mL of sterile saline water (0.9% NaCl w/v) in a stomacher bag and homogenized for 4 min using a Stomacher (Interscience). Ten-fold serial dilutions were made and plated on the following agar media and conditions: Plate Count Agar (PCA) (Biolife Italiana srl, Milan, Italy), incubated for 48–72 h at 30 ± 2 °C, for total mesophilic bacteria counts; Slanetz Bartley Agar (SLA) (Biolife Italiana srl), incubated for 24–48 h at 37 °C, for enterococci enumeration; Violet Red Bile Glucose Agar (VRBGA) (Biolife Italiana srl), anaerobically incubated at 37 °C for 24–48 h for Enterobacteria; Brilliance E. coli (BEC) (Oxoid, Milan, Italy), incubated at 37 °C for 18–24 h for Escherichia coli; Mannitol Salt Agar (MSA) (Biolife Italiana srl), incubated at 32 °C for 48 h, for staphylococci; modified MRS (mMRS), according to Corsetti and co-workers [13], anaerobically incubated at 30 ± 2 °C for 48 h, for lactic acid bacteria; Sabouraud Dextrose Agar (SDA) (Liofilchem, Roseto degli Abruzzi, Italy) and Wallerstein Laboratory Nutrient Agar (WLN) (Liofilchem), supplemented with chloramphenicol (100 mg/L), incubated at 30 ± 2 °C for 48–72 h, for yeasts count.
2.3. Isolation, DNA Extraction and Identification of Lactic Acid Bacteria and Yeasts
From each mMRS agar plate, 20% of the total number of colonies were randomly selected. After purification, by streaking three times onto mMRS agar plates, each colony was microscopically examined, checked for catalase activity and Gram reaction. Isolates were stored at −80 °C in liquid culture using 20% (v/v) of glycerol until the use. Total genomic DNA was extracted from pure overnight cultures according to the protocol reported by Pino et al. [37]. DNA concentration was assessed by using the Fluorometer Qubit (Invitrogen, Carlsbad, CA, USA). PCR amplification of the 16S rRNA gene was performed using the primer pair 7-f (5′-AGA GTT TGA TC/TA/C TGG CTCAG-3′) and 1510-r (5′-ACG G (C/T) TACC TTG TTA CGA CTT-3′) according to Pino and co-workers [38]. The restriction endonucleases AluI and MspI (Thermo Fisher Scientific, Waltham, MA, USA) were used for the digestion of amplification products as previously reported [39]. PCR products, of at least one strain representative of each restriction pattern, were purified using the QIAquick PCR Purification Kit (Qiagen, Milan Italy) and sequenced on both strands using external primers through a DNA Sanger sequencing process performed by Eurofins Genomics (Vimodrone, Italy). LAB taxonomic identification was assessed by sequence analysis of the 16S ribosomal RNA gene using the the basic local alignment search tool (BLASTn) software in the NCBI-curated bacterial and Archaea RefSeq database. Sequences with a percentage identity ≥98% were considered to belong to the same species. The 16S rRNA gene sequences were deposited in GenBank under the accession numbers from OL314786 to OL314809.
Presumptive yeasts (20% of the total number of colonies) were randomly selected from both SDA and WLN agar media. Isolates were purified by streaking three times onto the same isolation medium, microscopically examined and stocked at −80 °C in liquid culture using 20% (w/v) of glycerol. DNA concentration was determined using a Nanodrop Nd 1000 spectrophotometer (260/280 nm) (Nano-drop Technologies, Wilmington, DE, USA). PCR amplification of 5.8S rRNA gene and the upstream and downstream internal transcribed spacer regions ITS1 and ITS2 (globally referred to as ITS regions) was carried out with the primers ITS1 (5′-TCCGTAGGTGAACCTGCGG-3′) and ITS4 (5′-TCCTCCGCTTATTGATATGC-3′) [40]. A 40 μL reaction mixture contained 1X Dream Taq Green Buffer (Mg2+ plus) (Thermo Scientific, Waltham, MA, USA), 200 µM of each dNTPs (Thermo Scientific, Waltham, MA, USA), 0.3 µM of each primer, 1 U of Dream Taq DNA polymerase (Thermo Scientific, Waltham, MA, USA) and 50 ng of gDNA template. Amplification reactions were performed in a T100 thermal cycler (BioRad, Hercules, CA, USA) with the following cycling conditions: initial denaturation at 95 °C for 5 min; 35 cycles of denaturation at 95 °C for 1 min, annealing at 55 °C for 2 min, and polymerization at 72 °C for 2 min; the polymerization was finalized by one more cycle at 72 °C for 10 min. ITS amplicons were subjected to RFLP analysis using the endonucleases HaeIII and HinfI (Thermo Fisher Scientific, Waltham, MA, USA) as reported by Esteve-Zarzoso et al. [41] and according to the manufacturer’s instructions. The restriction fragments were checked by electrophoresis in 2.0% (w/v) agarose gel. PCR and RFLP fragment lengths were compared with those contained in the database at www.yeast-id.org (CSIC and University of Valencia, Spain) with rank parameters at +/−10. At least one strain representative of each restriction pattern was submitted to PCR amplification of D1/D2 region of the 26S rRNA gene using primers NL1 (5′-GCATATCAATAAGCGGAGGAAAAG-3′) and NL4 (5′-GGTCCGTGTTTCAAGACGG-3′) [42]. The thermal conditions consisted of initial denaturation at 95 °C for 5 min followed by 36 cycles of denaturation at 95 °C for 1 min, annealing at 52 °C for 45 s and extension at 72 °C for 2 min; final extension at 72 °C for 10 min. All PCR products were purified with DNA Clean & Concentrator™-5 Kit (Zymo Research, Orange, CA, USA) and sequenced on both strands using NL1 and NL4 primers by a Sanger method process performed by external service (Bio-Fab Research srl, Rome, Italy). Sequences were edited manually, and consensus sequences were generated using the program SeqMan (DNASTAR, Madison, WI, USA). Sequences were compared against NCBI-RefSeq database for 26S rRNA gene sequences using BLASTn [43]. The sequences of 26S rRNA D1/D2 regions were deposited in GenBank under the accession numbers from MN058031 to MN058041 and from MZ221124 to MZ221135.
2.4. Genotyping of Lactic Acid Bacteria and Yeasts
All the LAB and yeast isolates were genotyped with a repetitive element palindromic PCR (rep-PCR) fingerprinting analysis with (GTG)5 primer (5′-GTG GTG GTG GTG GTG-3′) [44], according to the protocols reported in Solieri et al. [45] and Dakal et al. [46], respectively. Briefly, both PCR amplification reactions of LAB and yeasts isolates were carried out in a 20 µL final volume containing 1 x DreamTaq Green Buffer, 3 mM MgCl2, 200 µM of each dNTP, 5 U of DreamTaq DNA polymerase, 50 ng of template DNA and either 0.6 µM or 1 µM of (GTG)5 primer for yeast and LAB, respectively. In addition, the PCR mixture for LAB genotyping contained 100 µg/mL of BSA (bovine serum albumin) (Thermo Fisher Scientific, Waltham, MA, USA). All rep-PCR reactions were carried out in the same T100™ thermal cycler, using the following cycling parameters: for LAB, initial denaturation at 94 °C for 5 min; 35 cycles of denaturation at 94 °C for 30 s, annealing at 40 °C for 1 min, and extension at 72 °C for 4 min; final extension at 72 °C for 7 min; for yeasts, initial denaturation at 94 °C for 5 min; 40 cycles of denaturation at 94 °C for 15 s, annealing at 55 °C for 45 min, and extension at 72 °C for 1.30 min; final extension at 72 °C for 4 min. The products were run on 1.8% (w/v) agarose gel containing SYBR™ Safe DNA Gel Stain (Invitrogen, Waltman, MA, USA) for 6 h at the constant voltage of 70 V in 0.5X TBE buffer under refrigerated conditions. GeneRuler 100 bp Plus DNA Ladder (Thermo Fisher Scientific, Waltham, MA, USA) was used as molecular-weight standard. The obtained digitalized images were analyzed using the BioNumerics software v3.0 (Applied Maths, Sint-Martens-Latem, Belgium). Pearson’s correlation similarity coefficient was used to convert computed band patterns into a similarity matrix. Optimization and curve smoothening parameters were optimized with the specific script present in BioNumerics. The unweighted pair group method analysis using the arithmetic means (UPGMA) was used for the tree construction. LAB and yeast isolates of ≥80.2 and 92% similarity values were treated as a single strain, respectively. Reproducibility cut-off values were established in at least two PCR amplification reactions using the same DNA preparation from three randomly selected LAB or yeast strains.
2.5. Statistical and Phylogenetic Analysis
All experiments were performed in triplicate and data were reported as average values, provided with standard deviation. Data were subjected to one-way ANOVA and pair-comparisons were achieved by Tukey’s procedure at p < 0.05, using the Minitab 19 Statistical software.
Regarding LAB and yeasts phylogenetics, the related sequences were aligned with Muscle program [47] in MEGA X software [48] and the resulting alignment was subjected to a DNA substitution model analysis to select the best-fitting model. Phylogenetic relationships were computed using the Kimura 2-parameter (K2P) model and the Neighbor Joining (NJ) method, whereas a gamma distribution (+G) was used to model rate variation among sites. Bootstrap support values were obtained from 1000 random re-samplings. Interactive Tree of Life (ITOL) was used to visualize all trees [49].
3. Results
3.1. Microbiological Analysis
Results of the main microbial groups detected in M1, M2, M3, and M4 sourdough samples are reported in Table 1. Overall, high variability was observed among samples except for Enterobacteriaceae, Staphylococcus spp., and Escherichia coli were never detected. In detail, the highest mesophilic aerobic bacteria population was detected in the M1 sample whereas M3 and M4 samples exhibited the highest enterococci count. Lactic acid bacteria were detected in all samples with an average value of 8.59 log CFU/g. The M2 sample exhibited the highest LABs cell density (9.14 log CFU/g) whereas the lowest was observed in the M1 sample (7.88 log CFU/g). The yeast population showed a cell density of about 8 log CFU/g in all analyzed samples except for the M2 sample which was characterized by a lower count (6.73 log CFU/g) (Table 1).

Table 1.
Microbiological counts expressed as log10 CFU/g ± standard deviation of the main microbial groups detected in M1, M2, M3, and M4 sourdough samples collected from four different Sicilian bakeries.
3.2. LAB identification and Species Distribution
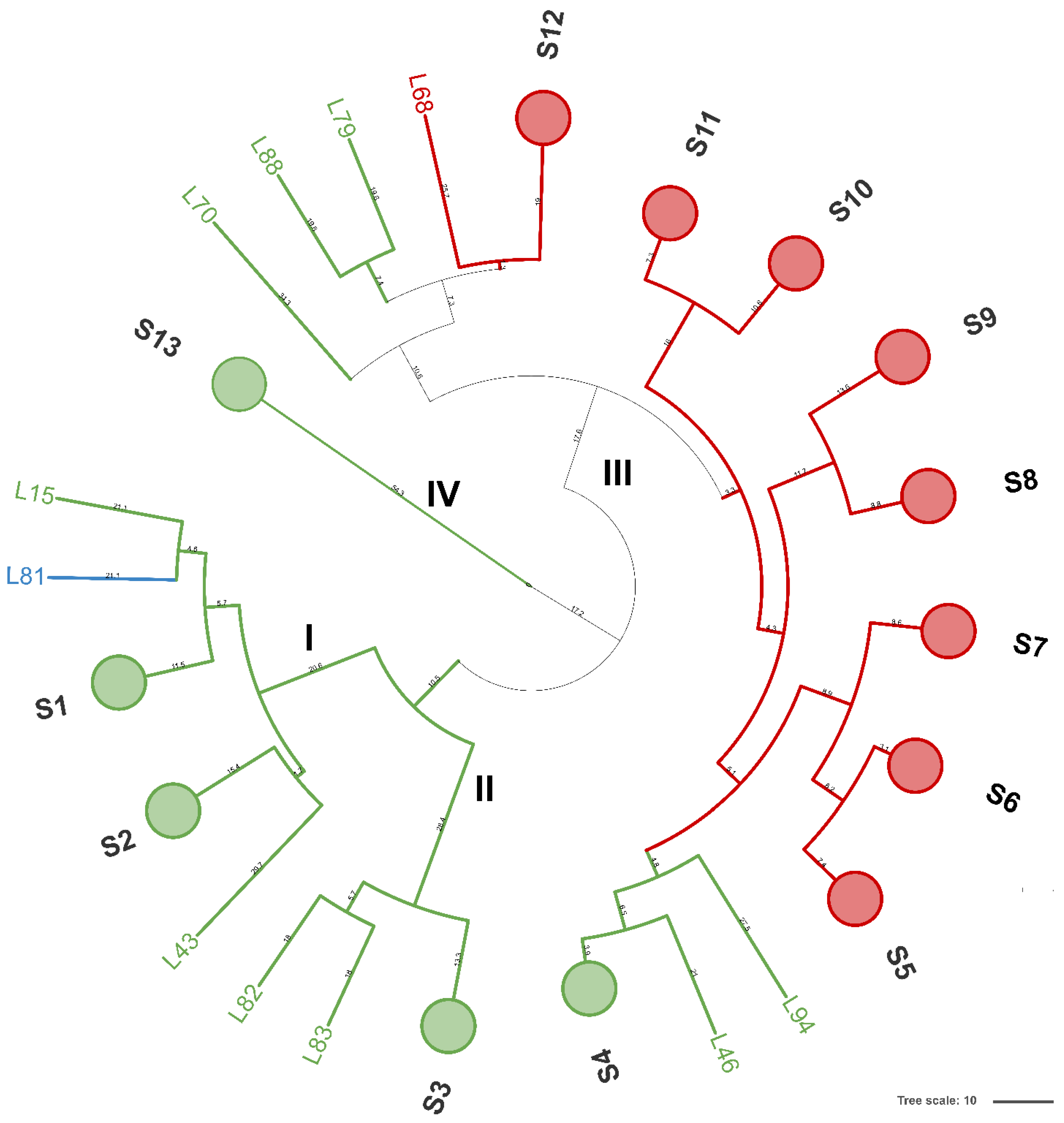
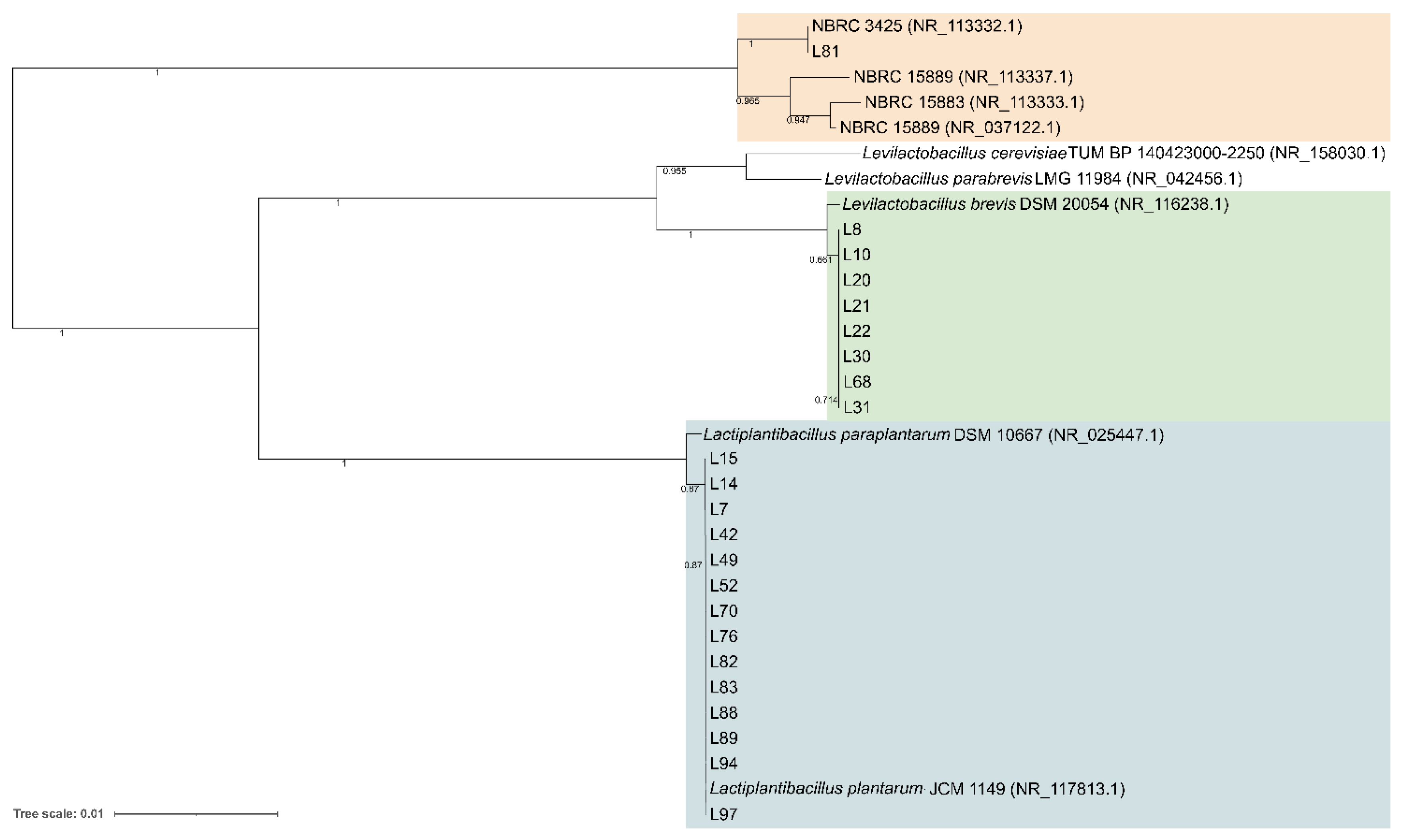
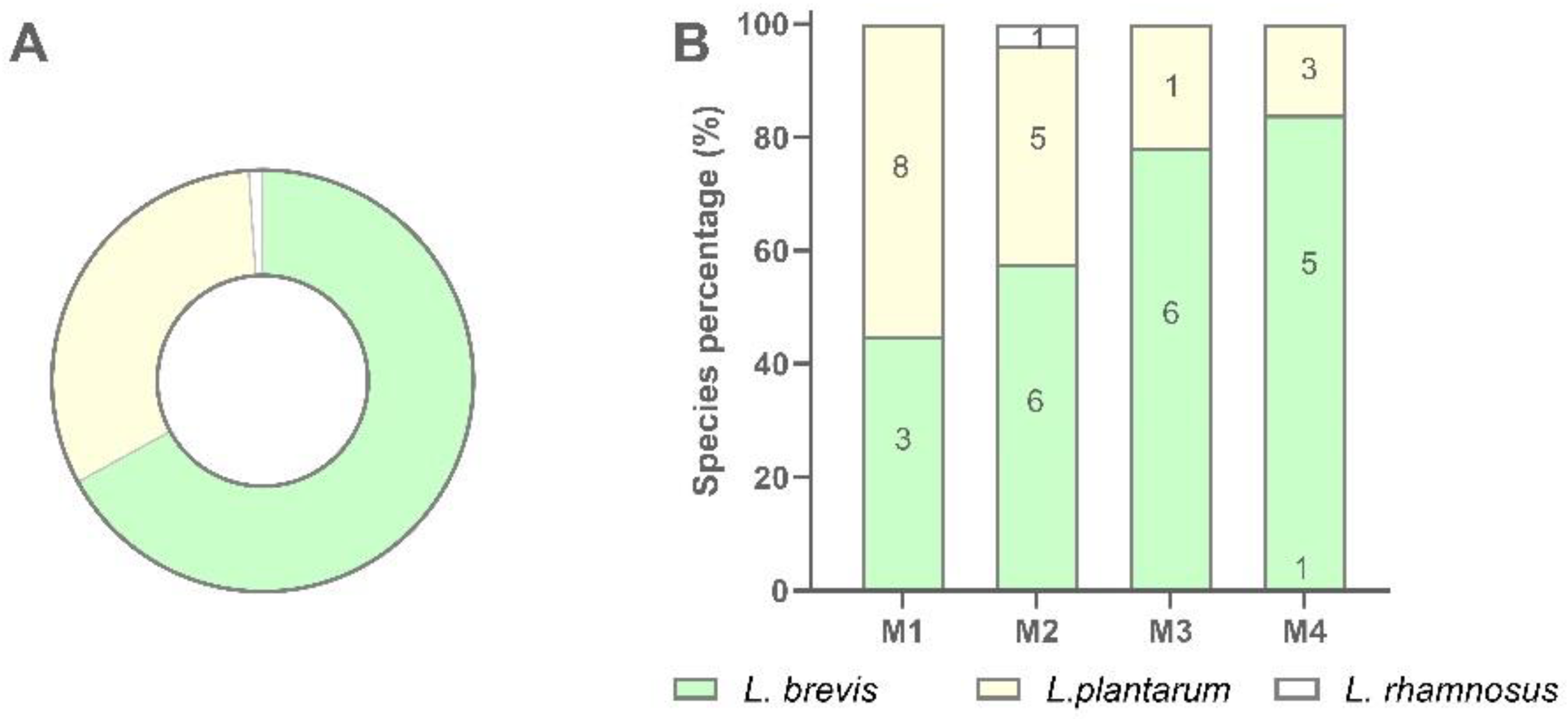
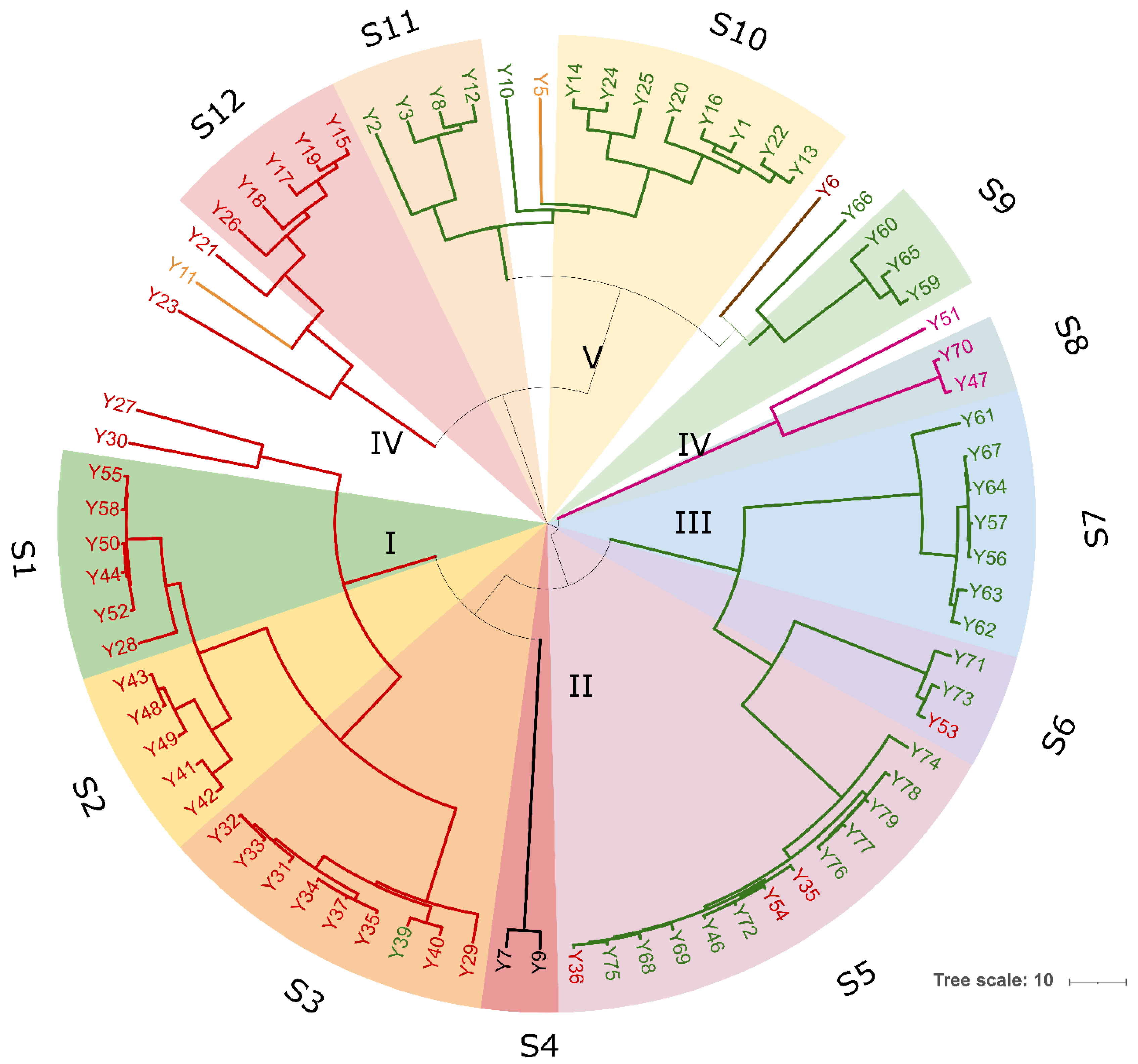
A total of 94 LAB isolates were preliminarily characterized by 16S rRNA gene ARDRA analysis. Simultaneous double digestion with the restriction enzymes AluI and MspI allowed for the discrimination of isolates into three different restriction patterns, namely A, B, and C which were congruent with those expected for the species Levilactobacillus brevis, Lactiplantibacillus plantarum, and those of the Lacticaseibacillus casei group, respectively (supplementary Table S2). rep-PCR fingerprinting with (GTG)5 primer was performed to genotype the library of LAB isolates. (GTG)5 fingerprints consisted of band numbers from 5 to 19 and band sizes ranging from 280 to 2630 bp. Clustering analysis of (GTG)5-fingerprints showed that four major clusters, named from I to IV, were found at the similarity threshold of 50% (Figure 1). Cluster I mainly grouped 13 isolates with 16S rRNA gene ARDRA pattern B and one isolate with pattern C, whereas cluster II consisted of five isolates most of which showed 16S rRNA gene ARDRA profile A. Cluster III was an intermixed group of 71 isolates with patterns A and B, whilst cluster IV grouped five isolates showing the restriction profile B. Using 80.2% as reproducibility threshold we grouped 83 isolates into 13 subclusters and identified 11 singletons. This similarity cut-off was chosen from duplicate experiments to estimate the experimental reproducibility of (GTG)5 typing method (data not shown). At least one strain representative of each subcluster and/or strains for which incongruence was scored between (GTG)5 clustering and 16S rRNA gene ARDRA profile was submitted to the sequencing of the 16S rRNA gene and BLASTn analysis (Table S3). As reported in Figure 2, an NJ-based tree was built to identify the phylogenetic relationships among isolates. Strains with 16S rRNA gene ARDRA profile B formed a monophyletic and statistically supported clade with L. plantarum JCM1149, indicating their affiliation to this species. Strains with 16S rRNA gene ARDRA profile A clustered with the type strain of L. brevis DSM 20054T, while strain L81 clustered with L. rhamnosus NBR3425 with 100% bootstrapping (Figure 2). Congruently with the species attribution, eight subclusters, namely S1, S2, S3, S4, and S13, grouped 21 L. plantarum strains, while eight L. plantarum isolates and L. rhamnosus L81 were singletons. Sixty-four L. brevis isolates grouped into eight subclusters within the major cluster III, namely from S5 to S12, while L68 and L79 were the only two L. brevis singletons. Strain L14 belonged to L. plantarum but incongruently clustered into S5 together with L. brevis isolates (Table S3). This evidence supported that (GTG)5 rep-PCR should be integrated with other techniques, such as 16S rRNA gene ARDRA to properly select strains for sequencing and to avoid any misidentifications. Figure 3 shows that L. brevis and L. plantarum were scored in all the samples but with different percentages. In particular, L. plantarum was the dominant species in sample M1, while L. brevis dominated M2, M3, and M4 samples. L. rhamnosus was scored only in sample M2. The number of genotypes was high for every species in all the samples suggesting a high diversity in LAB communities populating these sourdoughs.
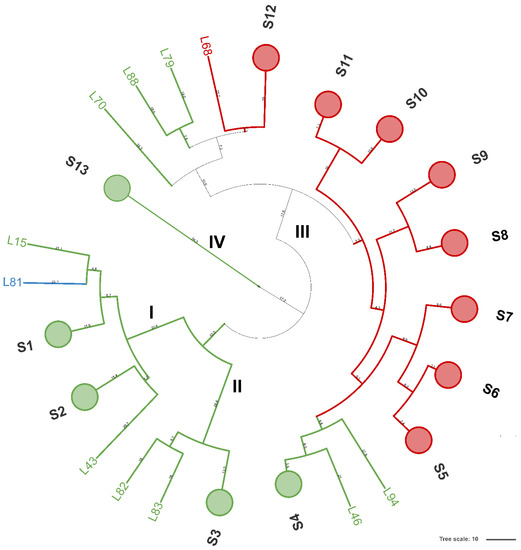
Figure 1.
Dendrogram analysis of (GTG)5-based rep-PCR patterns of LAB isolates. Similarities were calculated as Pearson r product-moment correlation coefficient and dendrogram was built using the Unweighted Pair Group Method with Arithmetic Mean (UPGMA). Numbers near the branch represent branch length. Circles are subclusters of biotypes obtained with a reproducibility cut-off of 80.2% and are labeled from S1 to S13. Major clusters are numbered with Roman letters from I to IV. Cluster and/or singletons belonging to L. brevis, L. plantarum, and L. rhamnosus are reported in red, green, and light blue, respectively. The tree was visualized using Itol.
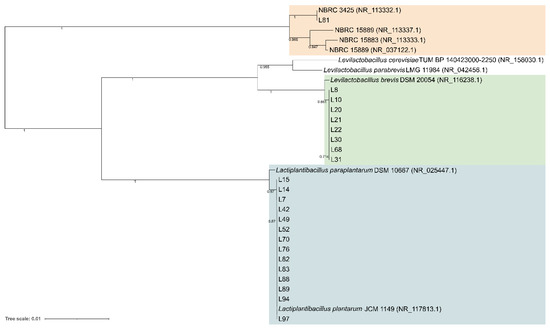
Figure 2.
Neighbor-joining phylogeny of LAB strains based on 16S rRNA gene partial sequences. Bootstrap supports (1000 replicates) >0.4 are shown next to the branches. The evolutionary distances were computed using the Kimura 2-parameter and the rate variation among sites was modeled with a gamma distribution (shape parameter = 1). The tree is drawn to scale, with branch lengths measured in the number of base substitutions per site. The analysis involved 32 nucleotide sequences. All ambiguous positions were removed for each sequence pair. There were a total of 1436 positions in the final dataset.
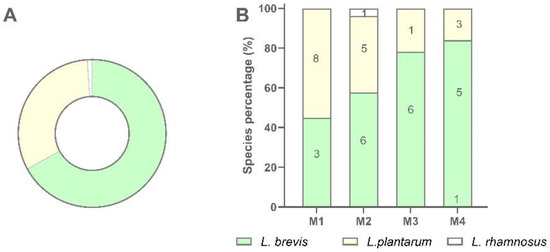
Figure 3.
Recovery percentage and species distribution of LAB isolates in sourdough samples. (A) Percentage of recovery of each LAB species; (B) Distribution of each species in M1, M2, M3, and M4 sourdough samples.
3.3. Yeast Identification and Species Distribution
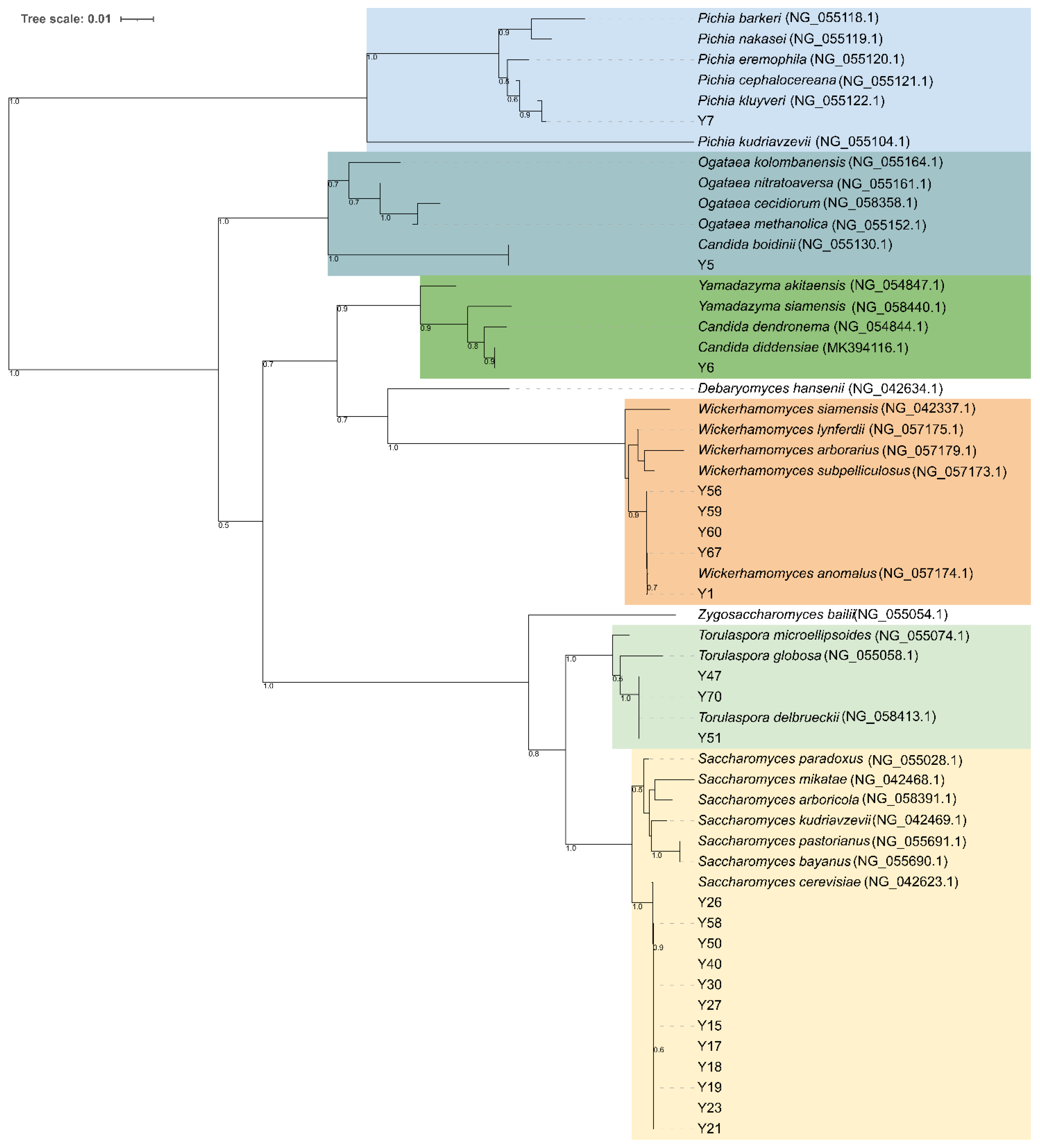
A total of 77 yeast isolates were successfully submitted to PCR amplification of the ITS region. Thirty-seven isolates showed an ITS amplicon of 660 bp, 32 isolates one of 880 bp, while five amplicons of 800, 700, 630, and 450 bp were found for 3, 1, 2, and 2 isolates, respectively. Combination of the ITS amplicons size and the restriction profiles obtained with endonucleases HaeIII and HinfI allowed the identification of seven different restriction patterns (referred to as A to F) corresponding to Wickerhamomyces anomalus (100%), Saccharomyces cerevisiae (100%), Torulaspora delbrueckii (100%), Candida boidinii (73%), Candida ethanolica (83%), and Candida maritima/Candida zeylanoides/Candida diddensiae (100%) in yeast-id database, respectively. Digestion of 620 bp amplicons with HaeIII resulted in undigested fragments in all 37 isolates, while digestion with HinfI produced two fragments of 310 and 300 bp (Table S4). Using HaeIII and HinfI all the 32 isolates having an 880 bp long fragment resulted in the same restriction patterns (325, 230, 170, and 125 bp with HaeIII and 375, 365, and 110 bp for HinfI, respectively) as the reference strains S. cerevisiae CBS1171T. Conversely, two isolates showing 630 bp long ITS amplicon, exhibited pattern F consisting in 420, 130 and 80 bp long fragments with HaeIII and two fragments of approximatively 310 bp with HinfI, respectively. This pattern did not allow for discrimination between C. maritima, C. zeylenoides, and C. diddensiae. Isolates producing ITS amplicons of 800, 700, and 450 bp length exhibited three distinct restriction patterns (Table S4). The BLASTn searches of 26S D1/D2 region sequences against the GenBank database confirmed species attribution for all the restriction patterns, except for pattern E, which corresponded to Pichia kluyveri instead of C. ethanolica (Table S5). Phylogenetic analysis showed that all the S. cerevisiae strains formed a monophyletic group with S. cerevisiae NRRL Y-12632 confirming species attribution based on PCR-RFLP analysis of ITS regions (Figure 4). Strains Y56, Y59, Y60, Y67, and Y1 clustered to W. anomalus with a high bootstrapping, forming a taxonomically congruent group within the Debaryomycetaceae clade. Strains Y47, Y70, and Y51 (pattern C) were phylogenetically identical to T. delbrueckii, while isolates Y6 representative for pattern F grouped to C. diddensiae (90% bootstrapping) (Table S4). Isolates Y5 and Y7 showing patterns D and E respectively, were close to C. boidinii and P. kluyveri species (Figure 4).
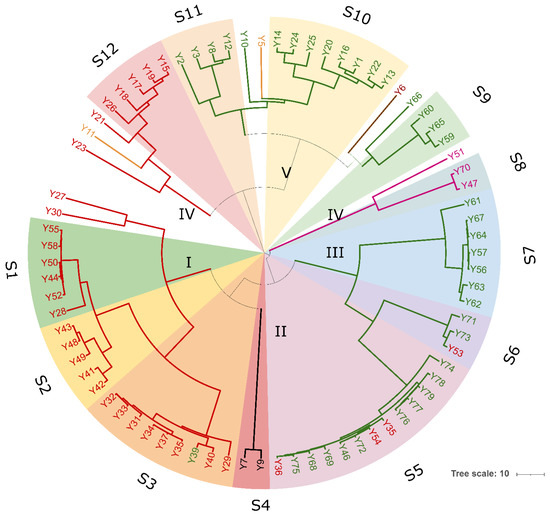
Figure 4.
Clustering analysis of (GTG)5-based rep-PCR patterns of yeast isolates. Similarities were calculated as Pearson r product-moment correlation coefficient and dendrogram was built using the Unweighted Pair Group Method with Arithmetic Mean (UPGMA) method. Major clusters are identified at 50% of similarity and numbered with Roman letters from I to IV. Biotypes and/or singletons obtained with a reproducibility cut-off of 92% are labeled from S1 to S12 and drawn with the following colour code: S. cerevisiae, red; W. anomalus, green; P. kluyveri, black; T. delbrueckii, purple; C. diddensiae, brown; and C. boidinii, orange. The tree was visualized using Itol.
Yeast isolates were genotyped with (GTG)5-based rep-PCR. The different band patterns generated by this method ranged from a minimum of six to a maximum of 12 bands, and sizes of amplified fragments between 274 and 2630 bp. The UPGMA dendrogram obtained from the cluster analyses of (GTG)5 fingerprints with the Pearson’s similarity coefficient grouped 77 yeast isolates into six major clusters (named from I to VI) at the cut-out value of 60% (Figure 5). Except for some strains, UPGMA cluster analysis provided a differentiation of the isolates according to the species attribution based on ITS PCR-RFLP analysis and 26S D1/D2 domains sequence-based phylogeny. Cluster I and II were homogeneous in species composition and grouped 23 S. cerevisiae and two P. kluyveri isolates, respectively (Figure 5). Cluster III was heterogeneous in species composition as it included 19 W. anomalus and four S. cerevisiae isolates, while cluster IV grouped only three isolates one belonging to S. cerevisiae and two to T. delbrueckii. Most of W. anomalus isolates grouped within cluster V, except for one C. diddensiae and one C. boidinii strains. Cluster VI was formed by seven S. cerevisiae strains and one C. diddensiae isolate (Figure 5). When a similarity value of 92% was applied as a reproducibility cut-off, 12 genotypes and 11 singletons strains were scored, suggesting a high level of intra-strain diversity among yeast isolates. We scored 10 biotypes for S. cerevisiae and W. anomalus, respectively; two for T. delbrueckii, two for C. diddensiae and one for P. kluyveri.
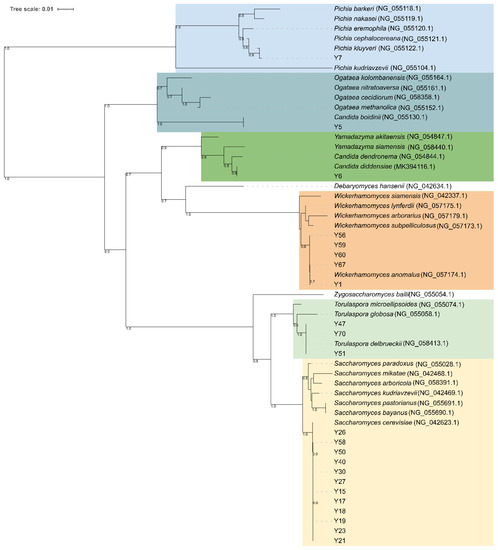
Figure 5.
Neighbor-joining phylogeny of yeast strains based on 26S rRNA gene partial sequences. Bootstrap supports (1000 replicates) > 0.5 are shown next to the branches. The evolutionary distances were computed using the Kimura 2-parameter and the rate variation among sites was modeled with a gamma distribution (shape parameter = 1). The tree is drawn to scale, with branch lengths measured in the number of base substitutions per site. The analysis involved 55 nucleotide sequences. All ambiguous positions were removed for each sequence pair. There was a total of 581 positions in the final dataset.
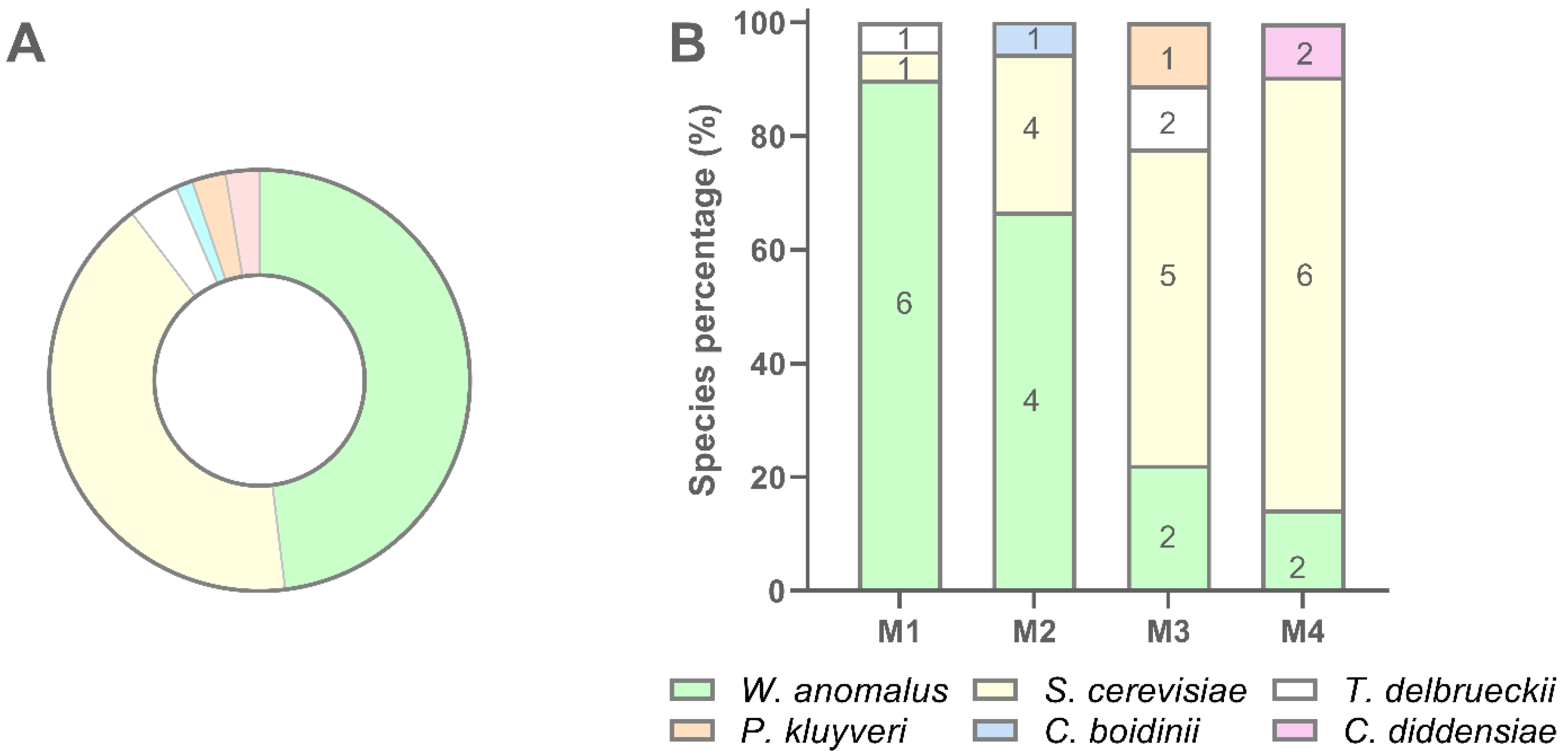
Globally W. anomalus accounted for 48.1% of isolates, followed by S. cerevisiae (41.6%), T. delbrueckii (3.9%), C. diddensiae (2.6%), P. kluyveri (2.6%), and C. boidinii (1.3%) (Figure 6, panel A). W. anomalus dominated sample M1 while S. cerevisiae was the dominant species in sample M4. Samples M2 and M3 showed W. anomalus and S. cerevisiae at intermediated distribution compared with M1 and M4 (Figure 6, panel B). In particular, M3 exhibited the most complex species composition as four species, namely W. anomalus, S. cerevisiae, T. delbrueckii, and P. kluyveri were recovered in this sourdough. In every sample, a high number of biotypes was found to dominate the yeast population, even if the number of biotypes per sample decreased with the decreasing of the number of isolates (Figure 6, panel B). In several cases, the same biotype was scored into different samples. C. boidinii and C. diddensiae were scored in only one sample, M2 and M4 respectively, and seemed to be occasional contaminant microorganisms for the low number of isolates recovered (Figure 6).
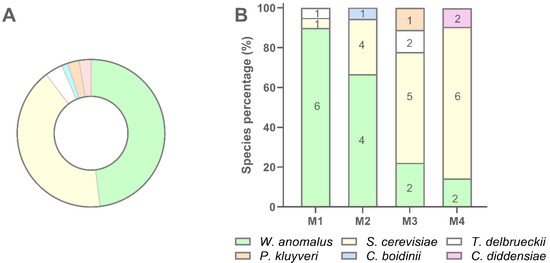
Figure 6.
Recovery percentage and species distribution of yeast isolates in sourdough samples. (A) Percentage of recovery of each yeast species; (B) Distribution of each species in M1, M2, M3, and M4 sourdough samples.
4. Discussion
Sourdough is one of the oldest starters traditionally used for making baked goods. Its use, alternatively to baker’s yeast and chemical leavening, offers several advantages since sourdough has been claimed as able to improve both sensory and rheology of the final product as well as shelf life. Recent data revealed that sourdough fermentation could increase mineral bioavailability, reduce the glycaemic index, improve protein digestibility, and decrease the anti-nutritional factors content [50]. Recently, the increasing attention to organic and natural products promoted the rediscovery of landrace/historical species of wheat. The renewed interest in ancient grains is correlated to the recognition of ancient wheat as a healthy cereal recommended for the treatment of several diseases, such as hypercholesterolemia, colitis, allergies, and insulin resistance [13,23,36]. Among these, Triticum aestivum L. subsp. aestivum, commonly named Maiorca or Majorca, is one of the Sicilian durum wheat landraces recently re-discovered by both consumers and local bakers.
Up to now, the microbial population of several traditional sourdoughs has been characterized although, based on our knowledge, no study has been conducted on Maiorca sourdough produced in Sicily. According to that, the present study aimed to characterize the native microbiota of spontaneously fermented Maiorca dough samples, obtained from four different bakeries located in Sicily.
Microbiological data revealed a quite similar cell density for both LAB and yeast populations suggesting the absence of competitiveness between the two communities. Although several studies reported higher LAB counts compared to yeasts one [10,51,52], the ability of both LAB and yeasts to co-dominate the sourdough ecosystems, creating a stable association, is renowned [24]. In the present study, the in-depth characterization of the lactobacilli population revealed that Lactiplantibacillus plantarum and Levilactobacillus brevis unquestionably dominated the Maiorca sourdough ecosystem. As recently revised by Arora and co-workers [53] the aforementioned species were commonly identified in more than 15 sourdoughs worldwide, meaning that they are the most common representatives of the LAB community. In particular, the dominance of L. plantarum was observed in Sardinian [54], Apulian [55], Molisan [56], and Sicilian traditional sourdoughs [16,57]. Similarly, Alfonzo and co-workers [11], by studying the microbial composition of flour samples, used in Sicily to make bread, reported the dominance of the L. plantarum species. In addition, Ventimiglia and co-workers [11] highlighted the codominance of L. plantarum with obligate heterofermentative species. According to that, the high occurrence of L. plantarum could be due to the bread-making technologies applied in Italian sourdough rather than the type of flour, as previously reported [51,58]. The robustness of L. plantarum strains, recovered in the sourdoughs analyzed in this study, could be due to several factors, such as rapid acidification, competition for nutrients, adaptability to environmental stresses, as well as ability to ferment hexose and pentose, synthesize diacetyl and hydrogen peroxide, and produce bacteriocins [59]. Similarly, to artisanal sourdough samples, obtained from bakeries located in Northern Italy [60,61], as well as French sourdoughs [62], the co-dominance of L. plantarum and L. brevis species was found in samples analyzed in the present study. This could be attributed to low incubation temperatures and continuous back-slopping creating a favorable environment for the development of the aforementioned species, as reported by De Vuyst and co-workers [4]. In discordance to previously reported data, which highlighted that Fructilactobacillus sanfranciscensis and Limosilactobacillus fermentum were frequently isolated from Italian and European sourdoughs [16,53,57,63], the aforementioned species were not detected in the analyzed Maiorca sourdough samples. According to that, Gobbetti and co-workers [64] reported on the strict association between L. sanfranciscensis and L. plantarum in Italian wheat sourdoughs and Minervini and co-workers [65] demonstrated the robustness of the L. plantarum under daily back-slopping propagation using wheat flour.
Concerning yeasts community, the rep-PCR with (GTG)5 primer method has been demonstrated to be powerful for discriminating biotypes in the yeast sourdough population [66] However, Ramírez-Castrillón et al. [67] proved that fingerprinting analysis carried out with only one microsatellite primers can group isolates belonging to different species in the same cluster leading to misidentification. We also found incongruences between species attribution carried out with conventional DNA barcoding approaches, such as ITS PCR-RFLP and 26S D1/D2 rDNA sequencing, and clustering results obtained by DNA fingerprinting with the (GTG)5 primed rep-PCR. These results supported that a combination of different molecular approaches is required for proper species attribution and intra-specific polymorphism detection of sourdough yeasts isolates. Compared to extensive sourdoughs studies [10,68,69,70], high species diversity was found albeit only four bakeries were sampled. Saccharomyces cerevisiae and Wickerhamomyces anomalus were the most frequently isolated species. It is well known that both S. cerevisiae and W. anomalus species are characterized by high adaptability to stressful conditions in terms of temperature, pH and osmolarity [24,69,70,71,72,73]. In particular, strains ascribed to the S. cerevisiae species are frequently isolated in sourdoughs from central and southern Italy [10,21,23]. S. cerevisiae is the yeast most frequently present in fermented sourdough, as it is able to produce a significant amount of ethanol in sourdoughs by consuming both maltose and glucose [74]. It was interesting to point out that L. brevis was recovered with a high percentage of isolation in samples dominated by S. cerevisiae. This result is in discordance to previously reported data, demonstrating the greatest adaptability of S. cerevisiae when it is associated with homofermentative compared to heterofermentative LAB [60,75]. In addition, the presence of both Candida boidinii and L. brevis, revealed in the M2 sample, could be associated with the absence of competition for maltose. As previously reported by Solieri and co-workers [76], Candida spp. ferments glucose and trehalose but not maltose. The high frequency of isolates ascribed to the W. anomalus species corroborates its well-known ability to thrive in a wide range of microbial ecosystems thanks to its competitiveness under stressful environmental conditions [71]. Congruently to the absence of L. sanfranciscensis, we also did not observe the maltose negative K. humilis in the yeast community. The stable association between maltose positive and maltose negative K. humilis due to the lack of antagonism for maltose has been stated by many authors [15,77]. The absence of both L. sanfranciscensis and K. humilis, in the analyzed Maiorca sourdough samples, could be probably related to external conditions, such as the environmental temperature, occurring during the sourdough fermentation [77,78].
Torulaspora delbrueckii and P. kudriavzevii have been recovered among the six yeast species most frequently associated with sourdoughs [24]. Congruently these species were scored in the analyzed Maiorca sourdoughs even if at low frequency. T. delbrueckii is recognized as a superior choice due to its importance in the production of frozen dough products since it exhibits a very good baking ability and a high capacity to resist osmotic and freeze–thaw stresses [79]. According to that, T. delbrueckii has already been used in the bakery industry in Japan, for the production of sweetbreads and pastries. P. kudriavzevii was found as a non-dominant yeast species in one out of four Danish sourdoughs in association with K. exigua, while P. kudriavzevii was found as a pure yeast culture in two Lithuanian sourdoughs. Only a few studies have, as yet, reported on the isolation of P. kudriavzevii from artisan Belgian [69] and Chinese [80] sourdoughs.
5. Conclusions
The culture-dependent approach, applied in the present study, allowed for in-depth investigation, for the first time, of the LAB and yeasts population of Sicilian Maiorca sourdough samples. LAB and yeasts co-dominated the sourdough ecosystem of the analyzed samples, creating a stable association. Among LAB, Lactiplantibacillus plantarum and Levilactobacillus brevis dominated the Maiorca sourdough ecosystem, whereas high species diversity was found among yeasts. Further studies will be conducted in order to evaluate both the pro-technological and functional properties of the isolated strains to be used as starters for the production of baked goods.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/microorganisms10020283/s1, Table S1. Characteristics of the sourdough samples M1, M2, M3, and M4 collected from the four selected bakeries located in Sicily. Table S2. Overview of the 16S rRNA gene ARDRA profiles and clustering data based on (GTG)5 rep-PCR. Table S3. 16S rRNA gene sequencing results and BLAST search on NCBI RefSeq database (16S ribosomal RNA nucleotide sequence records for Bacteria and Archaea). Table S4. Overview of the ITS restriction patterns of sourdough yeast isolates. Table S5. 26S rRNA D1/D2 region sequencing results and BLAST search on NCBI RefSeq database (28S ribosomal RNA Nucleotide sequence records).
Author Contributions
Conceptualization, C.L.R., C.C. and L.S. (Lisa Solieri); methodology, C.L.R., C.C. and L.S. (Lisa Solieri); formal analysis, A.P., L.S. (Laura Sola), N.R.; data curation, A.P., N.R., L.S. (Laura Sola); writing—original draft preparation, A.P., C.L.R. and L.S. (Lisa Solieri); writing—review and editing, A.P., C.L.R. and L.S. (Lisa Solieri). All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data presented in this study are included within the article.
Acknowledgments
The authors thank the bakeries for providing the sourdough samples.
Conflicts of Interest
The authors declare no conflict of interest.
References
- De Angelis, M.; Minervini, F.; Siragusa, S.; Rizzello, C.G.; Gobbetti, M. Wholemeal wheat flours drive the microbiome and functional features of wheat sourdoughs. Int. J. Food Microbiol. 2019, 302, 35–46. [Google Scholar] [CrossRef]
- Pétel, C.; Onno, B.; Prost, C. Sourdough volatile compounds and their contribution to bread: A review. Trends Food Sci. Technol. 2017, 59, 105–123. [Google Scholar] [CrossRef]
- Minervini, F.; De Angelis, M.; Di Cagno, R.; Gobbetti, M. Ecological parameters influencing microbial diversity and stability of traditional sourdough. Int. J. Food Microbiol. 2014, 171, 136–146. [Google Scholar] [CrossRef]
- De Vuyst, L.; Vrancken, G.; Ravyts, F.; Rimaux, T.; Weckx, S. Biodiversity, ecological determinants, and metabolic exploitation of sourdough microbiota. Food Microbiol. 2009, 26, 666–675. [Google Scholar] [CrossRef]
- Gänzle, M.; Ripari, V. Composition and function of sourdough microbiota: From ecological theory to bread quality. Int. J. Food Microbiol. 2016, 239, 19–25. [Google Scholar] [CrossRef]
- Minervini, F.; Celano, G.; Lattanzi, A.; De Angelis, M.; Gobbetti, M. Added ingredients affect the microbiota and biochemical characteristics of durum wheat type-I sourdough. Food Microbiol. 2016, 60, 112–123. [Google Scholar] [CrossRef]
- Minervini, F.; Dinardo, F.R.; Celano, G.; De Angelis, M.; Gobbetti, M. Lactic Acid bacterium population dynamics in artisan sourdoughs over one year of daily propagations is mainly driven by flour microbiota and nutrients. Front. Microbiol. 2018, 9, 1984. [Google Scholar] [CrossRef]
- Van Kerrebroeck, S.; Maes, D.; De Vuyst, L. Sourdoughs as a function of their species diversity and process conditions, a meta-analysis. Trends Food Sci. Technol. 2017, 68, 152–159. [Google Scholar] [CrossRef]
- Vancanneyt, M.; Neysens, P.; De Wachter, M.; Engelbeen, K.; Snauwaert, C.; Cleenwerck, I.; Swings, J. Lactobacillus acidifarinae sp. nov. and Lactobacillus zymae sp. nov., from wheat sourdoughs. Int. J. Syst. Evol. 2005, 55, 615–620. [Google Scholar] [CrossRef]
- Minervini, F.; Di Cagno, R.; Lattanzi, A.; De Angelis, M.; Antonielli, L.; Cardinali, G.; Cappelle, S.; Gobbetti, M. Lactic acid bacterium and yeast microbiotas of 19 sourdoughs used for traditional/typical Italian breads: Interactions between ingredients and microbial species diversity. Appl. Environ. Microbiol. 2012, 78, 1251–1264. [Google Scholar] [CrossRef] [Green Version]
- Alfonzo, A.; Ventimiglia, G.; Corona, O.; Di Gerlando, R.; Gaglio, R.; Francesca, N.; Settanni, L. Diversity and technological potential of lactic acid bacteria of wheat flours. Food Microbiol. 2013, 36, 343–354. [Google Scholar] [CrossRef] [Green Version]
- Vogel, R.F.; Müller, M.R.A.; Stolz, P.; Ehrmann, M.A. Ecology in sourdough produced by traditional and modern technologies. Adv. Food Sci. 1996, 18, 152–159. [Google Scholar]
- Corsetti, A.; Lavermicocca, P.; Morea, M.; Baruzzi, F.; Tosti, N.; Gobbetti, M. Phenotypic and molecular identification and clustering of lactic acid bacteria and yeasts from wheat (species Triticum durum and Triticum aestivum) sourdoughs of Southern Italy. Int. J. Food Microbiol. 2001, 64, 95–104. [Google Scholar] [CrossRef]
- Palomba, S.; Blaiotta, G.; Ventorino, V.; Saccone, A.; Pepe, O. Microbial characterization of sourdough for sweet baked products in the Campania region (Southern Italy) by a polyphasic approach. Ann. Microbiol. 2011, 61, 307–314. [Google Scholar] [CrossRef]
- Venturi, M.; Guerrini, S.; Vincenzini, M. Stable and non-competitive association of Saccharomyces cerevisiae, Candida milleri and Lactobacillus sanfranciscensis during manufacture of two traditional sourdough baked goods. Food Microbiol. 2012, 1, 107–115. [Google Scholar] [CrossRef]
- Randazzo, C.L.; Heilig, H.; Restuccia, C.; Giudici, P.; Caggia, C. Bacterial population in traditional sourdough evaluated by molecular methods. J. Appl. Microbiol. 2005, 99, 251–258. [Google Scholar] [CrossRef]
- Siragusa, S.; Di Cagno, R.; Ercolini, D.; Minervini, F.; Gobbetti, M.; De Angelis, M. Taxonomic structure and monitoring of the dominant population of lactic acid bacteria during wheat flour sourdough type I propagation using Lactobacillus sanfranciscensis starters. Appl. Environ. Microbiol. 2009, 75, 1099–1109. [Google Scholar] [CrossRef] [Green Version]
- Picozzi, C.; Bonacina, G.; Vigentini, I.; Foschino, R. Genetic diversity in Italian Lactobacillus sanfranciscensis strains assessed by multilocus sequence typing and pulsed field gel electrophoresis analyses. Microbiology 2010, 156, 2035–2045. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Pulvirenti, A.; Solieri, L.; Gullo, M.; De Vero, L.; Giudici, P. Occurrence and dominance of yeast species in sourdough. Lett. Appl. Microbiol. 2004, 38, 113–117. [Google Scholar] [CrossRef] [Green Version]
- Valmorri, S.; Tofalo, R.; Settanni, L.; Corsetti, A.; Suzzi, G. Yeast microbiota associated with spontaneous sourdough fermentations in the production of traditional wheat sourdough breads of the Abruzzo region (Italy). Antonie Van Leeuwenhoek 2010, 97, 119–129. [Google Scholar] [CrossRef]
- Di Cagno, R.; Pontonio, E.; Buchinb, S.; De Angelis, M.; Lattanzia, A.; Valerio, F.; Gobbetti, M.; Calasso, M. Diversity of the lactic acid bacterium and yeast microbiota in the switch from firm- to liquid-sourdough fermentation. Appl. Environ. Microbiol. 2014, 80, 3161–3172. [Google Scholar] [CrossRef] [Green Version]
- Palla, M.; Agnoluccia, M.; Calzonea, A.; Giovannettia, M.; Di Cagno, R.; Gobbetti, M.; Rizzello, C.G.; Pontonio, E. Exploitation of autochthonous Tuscan sourdough yeasts as potential starters. Int. J. Food Microbiol. 2019, 302, 59–68. [Google Scholar] [CrossRef]
- De Vuyst, L.; Harth, H.; Van Kerrebroeck, S.; Leroy, F. Yeast diversity of sourdoughs and associated metabolic properties and functionalities. Int. J. Food Microbiol. 2016, 239, 26–34. [Google Scholar] [CrossRef]
- Gobbetti, M.; Rizzello, C.G.; Di Cagno, R.; De Angelis, M. How the sourdough may affect the functional features of leavened baked goods. Food Microbiol. 2014, 37, 30–40. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Montemurro, M.; Gobbetti, M. Characterization of the bread made with durum wheat semolina rendered gluten free by sourdough biotechnology in comparison with commercial gluten-free products. J. Food Sci. 2016, 81, 2263–2272. [Google Scholar] [CrossRef]
- Kariluoto, S.; Edelmann, M.; Nyström, L.; Sontag-Strohm, T.; Salovaara, H.; Kivelä, R.; Herranenb, M.; Korholab, M.; Piironen, V. In situ enrichment of folate by microorganisms in beta-glucan rich oat and barley matrices. Int. J. Food Microbiol. 2014, 176, 38–48. [Google Scholar] [CrossRef]
- Liukkonen, K.H.; Katina, K.; Wilhelmsson, A.; Myllymaki, O.; Lampi, A.M.; Kariluoto, S.; Poutanen, K. Process-induced changes on bioactive compounds in whole grain rye. Proc. Nutr. Soc. 2003, 62, 117–122. [Google Scholar] [CrossRef]
- Türk, M.; Sandberg, A.S.; Carlsson, N.G.; Andlid, T. Inositol hexaphosphate hydrolysis by baker’s yeast. Capacity, kinetics, and degradation products. J. Agric. Food Chem. 2000, 48, 100–104. [Google Scholar]
- Moore, J.; Cheng, Z.; Hao, J.; Guo, G.; Liu, J.G.; Lin, C.; Yu, L. Effects of solid-state yeast treatment on the antioxidant properties and protein and fiber compositions of common hard wheat bran. J. Agric. Food Chem. 2007, 55, 10173–10182. [Google Scholar] [CrossRef]
- Wang, T.; He, F.; Chen, G. Improving bioaccessibility and bioavailability of phenolic compounds in cereal grains through processing technologies: A concise review. J. Funct. Foods 2014, 7, 101–111. [Google Scholar] [CrossRef]
- Moslehi-Jenabian, S.; Pedersen, L.L.; Jespersen, L. Beneficial effects of probiotic and food borne yeasts on human health. Nutrients 2010, 2, 449–473. [Google Scholar] [CrossRef]
- Katina, K.; Poutanen, K. Nutritional aspects of cereal fermentation with lactic acid bacteria and yeast. In Handbook on Sourdough Biotechnology; Gobbetti, M., Gänzle, M., Eds.; Springer: New York, NY, USA, 2014; Volume 1, pp. 229–244. [Google Scholar]
- Perricone, M.; Bevilacqua, A.; Corbo, M.R.; Sinigaglia, M. Technological characterization and probiotic traits of yeasts isolated from Altamura sourdough to select promising microorganisms as functional starter cultures for cereal-based products. Food Microbiol. 2014, 38, 26–35. [Google Scholar] [CrossRef]
- Boukid, F.; Folloni, S.; Sforza, S.; Vattidini, E.; Prandi, B. Current trends in ancient grains-based foodstuffs: Insights into nutritional aspects and technological applications. Compr. Rev. Food Sci. Food Saf. 2018, 17, 123–136. [Google Scholar] [CrossRef] [Green Version]
- Visioli, G.; Giannelli, G.; Agrimonti, C.; Spina, A.; Pasini, G. Traceability of Sicilian Durum Wheat Landraces and Historical Varieties by High Molecular Weight Glutenins. Agronomy 2021, 11, 143. [Google Scholar] [CrossRef]
- Pino, A.; Liotta, L.; Randazzo, C.L.; Todaro, A.; Mazzaglia, A.; De Nardo, F.; Chiofalo, V.; Caggia, C. Polyphasic approach to study physico-chemical, microbiological and sensorial characteristics of artisanal Nicastrese goat’s cheese. Food Microbiol. 2018, 70, 143–154. [Google Scholar] [CrossRef]
- Pino, A.; Liotta, L.; Caggia, C.; Chiofalo, V.; De Nardo, F.; Zumbo, A.; Todaro, A.; Randazzo, C.L. Effect of seasonality on physico-chemical and microbiological properties of nicastrese milk and artisanal cheese. FEMS Microbiol. Lett. 2021, 368, fnab055. [Google Scholar] [CrossRef]
- Pino, A.; Russo, N.; Van Hoorde, K.; De Angelis, M.; Sferrazzo, G.; Randazzo, C.L.; Caggia, C. Piacentinu Ennese PDO Cheese as reservoir of promising probiotic bacteria. Microorganisms 2019, 7, 254. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Esteve-Zarzoso, B.; Belloch, C.; Uruburu, F.; Querol, A. Identification of yeasts by RFLP analysis of the 5.8S rRNA gene and the two ribosomal internal transcribed spacers. Int. J. Syst. Bacteriol. 1999, 49, 329–337. [Google Scholar] [CrossRef] [Green Version]
- Kurtzman, C.; Robnett, C.J. Identification and phylogeny of ascomycetous yeasts from analysis of nuclear large subunit (26S) ribosomal DNA partial sequences. Antonie Van Leeuwenhoek 1998, 73, 331–371. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Versalovic, J.; Schneider, M.; de Bruijn, F.J.; Lupski, J. R Genomic fingerprinting of bacteria using repetitive sequence-based PCR (rep-PCR). Methods Mol. Cell. Biol. 1994, 5, 25–40. [Google Scholar]
- Solieri, L.; Bianchi, A.; Giudici, P. Inventory of non starter lactic acid bacteria from ripened Parmigiano Reggiano cheese as assessed by a culture dependent multiphasic approach. Syst. Appl. Microbiol. 2012, 35, 270–277. [Google Scholar] [CrossRef]
- Dakal, T.C.; Solieri, L.; Giudici, P. Evaluation of fingerprinting techniques to assess genotype variation among Zygosaccharomyces strains. Food Microbiol. 2018, 72, 135–145. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, 293–296. [Google Scholar] [CrossRef]
- Graça, C.; Lima, A.; Raymundo, A.; Sousa, I. Sourdough fermentation as a tool to improve the nutritional and health-promoting properties of its derived-products. Fermentation 2021, 7, 246. [Google Scholar] [CrossRef]
- Gobbetti, M.; Minervini, F.; Pontonio, E.; Di Cagno, R.; De Angelis, M. Drivers for the establishment and composition of the sourdough lactic acid bacteria biota. Int. J. Food Microbiol. 2016, 239, 3–18. [Google Scholar] [CrossRef]
- Zhang, G.; Tu, J.; Sadiq, F.A.; Zhang, W.; Wang, W. Prevalence, genetic diversity, and technological functions of the Lactobacillus sanfranciscensis in Sourdough: A Review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1209–1226. [Google Scholar] [CrossRef] [Green Version]
- Arora, K.; Ameur, H.; Polo, A.; Di Cagno, R.; Rizzello, C.G.; Gobbetti, M. Thirty years of knowledge on sourdough fermentation: A systematic review. Trends Food Sci. Technol. 2021, 108, 71–83. [Google Scholar] [CrossRef]
- Catzeddu, P.; Mura, E.; Parente, E.; Sanna, M.; Farris, G.A. Molecular characterization of lactic acid bacteria from sourdough breads produced in Sardinia (Italy) and multivariate statistical analyses of results. Syst. Appl. Microbiol. 2006, 29, 138–144. [Google Scholar] [CrossRef]
- Ricciardi, A.; Parente, E.; Piraino, P.; Paraggio, M.; Romano, P. Phenotypic characterization of lactic acid bacteria from sourdoughs for Altamura bread produced in Apulia (southern Italy). Int. J. Food Microbiol. 2005, 98, 63–72. [Google Scholar] [CrossRef]
- Reale, A.; Di Renzo, T.; Succi, M.; Tremonte, P.; Coppola, R.; Sorrentino, E. Identification of lactobacilli isolated in traditional ripe wheat sourdoughs by using molecular methods. World J. Microbiol. Biotechnol. 2011, 27, 237–244. [Google Scholar] [CrossRef]
- Ventimiglia, G.; Alfonzo, A.; Galluzzo, P.; Corona, O.; Francesca, N.; Caracappa, S.; Moschetti, G.; Settanni, L. Codominance of Lactobacillus plantarum and obligate heterofermentative lactic acid bacteria during sourdough fermentation. Food Microbiol. 2015, 51, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Reale, A.; Di Renzo, T.; Preziuso, M.; Panfili, G.; Cipriano, L.; Messia, M.C. Stabilization of sourdough starter by spray drying technique: New bread making perspective. LWT 2019, 99, 468–475. [Google Scholar] [CrossRef]
- Gobbetti, M.; De Angelis, M.; Corsetti, A.; Di Cagno, R. Biochemistry and physiology of sourdough lactic acid bacteria. Trends Food Sci. Technol. 2005, 16, 57–69. [Google Scholar] [CrossRef]
- Iacumin, L.; Cecchini, F.; Manzano, M.; Osualdini, M.; Boscolo, D.; Orlic, S.; Comi, G. Description of the microflora of sourdoughs by culture-dependent and cultureindependent methods. Food Microbiol. 2009, 26, 128–135. [Google Scholar] [CrossRef]
- Osimani, A.; Zannini, E.; Aquilanti, L.; Mannazzu, I.M.; Comitini, F.; Clementi, F. Lactic acid bacteria and yeasts from wheat sourdoughs of the Marche region. Ital. J. Food Sci. 2009, 3, 269–286. [Google Scholar]
- Michel, E.; Monfort, C.; Deffrasnes, M.; Guezenec, S.; Lhomme, E.; Barret, M.; Sicard, D.; Dousset, X.; Onno, B. Characterization of relative abundance of lactic acid bacteria species in French organic sourdough by cultural, qPCR and MiSeq high-throughput sequencing methods. Int. J. Food Microbiol. 2016, 239, 35–43. [Google Scholar] [CrossRef] [Green Version]
- DeVuyst, L.; Schrijvers, V.; Paramithiotis, S.; Hoste, B.; Vancanneyt, M.; Swings, J.; Kalatzopoulos, G.; Tsakalidou, E.; Messens, W. The biodiversity of lactic acid bacteria in Greek traditional sourdoughs is reflected in both composition and metabolite formation. Appl. Environ. Microbiol. 2002, 68, 6059–6069. [Google Scholar] [CrossRef] [Green Version]
- Gobbetti, M. The sourdough microflora: Interactions of lactic acid bacteria and yeasts. Trends Food Sci. Technol. 1998, 9, 267–274. [Google Scholar] [CrossRef]
- Minervini, F.; De Angelis, M.; Di Cagno, R.; Pinto, D.; Siragusa, S.; Rizzello, C.G.; Gobbetti, M. Robustness of Lactobacillus plantarum starters during daily propagation of wheat flour sourdough type I. Food Microbiol. 2010, 27, 897–908. [Google Scholar] [CrossRef]
- Palla, M.; Cristani, C.; Giovannetti, M.; Agolucci, M. Large Genetic Intraspecific Diversity of Autochthonous Lactic Acid Bacteria and Yeasts Isolated from PDO Tuscan Bread Sourdough. Appl. Sci. 2020, 10, 1043. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Castrillón, M.; Mendes, S.D.; Inostroza-Ponta, M.; Valente, P. (GTG)5 MSP-PCR fingerprinting as a technique for discrimination of wine associated yeasts? PLoS ONE 2014, 9, e105870. [Google Scholar]
- Lattanzi, A.; Minervini, F.; Di Cagno, R.; Diviccaro, A.; Antonielli, L.; Cardinali, G.; Cappelle, S.; De Angelis, M.; Gobbetti, M. The lactic acid bacteria and yeast microbiota of eighteen sourdoughs used for the manufacture of traditional Italian sweet leavened baked goods. Int. J. Food Microbiol. 2013, 15, 71–79. [Google Scholar] [CrossRef]
- Vrancken, G.; De Vuyst, L.; Van der Meulen, R.; Huys, G.; Vandamme, P.; Daniel, H.M. Yeast species composition differs between artisan bakery and spontaneous laboratory sourdoughs. FEMS Yeast Res. 2010, 10, 471–481. [Google Scholar] [CrossRef]
- Lhomme, E.; Urien, C.; Legrand, J.; Dousset, X.; Onno, B.; Sicard, D. Sourdough microbial community dynamics: An analysis during French organic bread-making processes. Food Microbiol. 2016, 53, 41–50. [Google Scholar] [CrossRef]
- Daniel, H.M.; Moons, M.C.; Huret, S.; Vrancken, G.; De Vuyst, L. Wickerhamomyces anomalus in the sourdough microbial ecosystem. Antonie Van Leeuwenhoek 2011, 99, 63–73. [Google Scholar] [CrossRef]
- Huys, G.; Daniel, H.M.; De Vuyst, L. Taxonomy and Biodiversity of Sourdough Yeasts and Lactic Acid Bacteria. In Handbook on Sourdough Biotechnology; Springer: Boston, MA, USA, 2011; pp. 105–154. [Google Scholar]
- Cappelli, A.; Ulissi, U.; Valzano, M.; Damiani, C.; Epis, S.; Gabrielli, M.G.; Conti, S.; Polonelli, L.; Bandi, C.; Favia, G.; et al. A Wickerhamomyces anomalus killer strain in the malaria vector Anopheles stephensi. PLoS ONE 2014, 9, e95988. [Google Scholar] [CrossRef]
- Lahue, C.; Madden, A.A.; Dunn, R.R.; Smukowski Heil, C. History and Domestication of Saccharomyces cerevisiae in Bread Baking. Front. Genet. 2020, 11, 1373. [Google Scholar] [CrossRef]
- Gobbetti, M.; Corsetti, A.; Rossi, J. Interaction between lactic acid bacteria and yeasts in sourdough using a rheofermentometer. World J. Microbiol. Biotechnol. 1995, 11, 625–630. [Google Scholar] [CrossRef]
- Solieri, L.; De Vero, L.; Pulvirenti, A. Studio fenotipico e molecolare di lieviti in paste acide tradizionali. Ind. Aliment. 2003, 42, 971–978. [Google Scholar]
- Carbonetto, B.; Nidelet, T.; Guezenec, S.; Perez, M.; Segond, D.; Sicard, D. Interactions between Kazachstania humilis yeast species and Lactic Acid Bacteria in Sourdough. Microorganisms 2020, 8, 240. [Google Scholar] [CrossRef] [Green Version]
- Gänzle, M.G.; Ehmann, M.; Hammes, W.P. Modeling of Growth of Lactobacillus sanfranciscensis and Candida milleri in Response to Process Parameters of Sourdough Fermentation. Appl. Environ. Microbiol. 1998, 7, 2616–2623. [Google Scholar] [CrossRef] [Green Version]
- Almeida, M.; Pais, C. Leavening ability and freeze tolerance of yeasts isolated from traditional corn and rye bread doughs. Appl. Environ. Microbiol. 1997, 62, 4401–4404. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Liu, W.; Sun, Z.; Bao, Q.; Wang, F.; Yu, J.; Zhang, H. Diversity of lactic acid bacteria and yeasts in traditional sourdoughs collected from western region in Inner Mongolia of China. Food Control 2011, 22, 767–774. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).