Betanodavirus and VER Disease: A 30-year Research Review


Abstract
1. Introduction
2. The Virus
2.1. Viral Structure
2.2. Viral Replication
2.3. Taxonomic Classification
2.4. Geographical Distribution and Host Range
2.5. Viral Thermotolerance
3. The Disease: Viral Encephalopathy and Retinopathy (VER) or Viral Nervous Necrosis (VNN)
3.1. Routes of Infection and Spread through the Fish Body
3.2. Viral Transmission
3.3. Host Response
3.4. Disease Outbreaks
4. Epidemiology
4.1. VER Outbreaks and NNV Detections in Farmed Fish
4.2. NNV in Wild Fish
4.3. NNV in Invertebrates and other Marine Animals
5. Diagnostics
5.1. Cell Cultures
5.2. Diagnostic Procedures
5.3. Control of the Disease
6. Prevention
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Glazebrook, J.S.; Campbell, R.S.F. Diseases of barramundi (Lates calcarifer) in Australia: A review. In Management of Wild and Cultured Sea Bass/Barramundi (Lates calcarifer), Proceedings of an international workshopm Darwin, N.T. Australia; Coppland, J.W., Grey, D.L., Eds.; Australian Centre for International Agruculture Research (ACIAR): Camberra, Australia, 1987; pp. 204–206. [Google Scholar]
- Callinan, R.B. Diseases of Australian native fishes. In Fish Diseases; Bryden, D.I., Ed.; Post Graduate Committee in Veterinary Science, University of Sydney: Sydney, Australia, 1988; pp. 459–472. [Google Scholar]
- Bellance, R.; Gallet de Saint-Aurin, D. L’encephalite virale de loup de mer. Caraibes Med. 1998, 2, 105–114. [Google Scholar]
- Breuil, G.; Bonami, J.R.; Pepin, J.F.; Pichot, Y. Viral infection (picorna-like virus) associated with mass mortalities in hatchery-reared sea-bass (Dicentrarchus labrax) larvae and juveniles. Aquaculture 1991, 97, 109–116. [Google Scholar] [CrossRef]
- Glazebrook, J.S.; Heasman, M.P.; Beer, S.W. Picorna-like viral particles associated with mass mortalities in larval barramundi, Lates calcarifer Bloch. J. Fish Dis. 1990, 13, 245–249. [Google Scholar] [CrossRef]
- Mori, K.-I.; Nakai, T.; Muroga, K.; Arimoto, M.; Mushiake, K.; Furusawa, I. Properties of a new virus belonging to nodaviridae found in larval striped jack (Pseudocaranx dentex) with nervous necrosis. Virology 1992, 187, 368–371. [Google Scholar] [CrossRef]
- Comps, M.; Pépin, J.F.; Bonami, J.R. Purification and characterization of two fish encephalitis viruses (FEV) infecting Lates calcarifer and Dicentrarchus labrax. Aquaculture 1994, 123, 1–10. [Google Scholar] [CrossRef]
- Ball, L.; Hendry, D.; Johnson, J.; Ruechert, R.; Scotti, P. Family Nodaviridae. In Virus Taxonomy. Seventh Report of the International Committee on Taxonomy of Viruses; Van Regenmortel, M.H., Fauquet, C.M., Bishop, D.H.L., Cartens, E.B., Estes, M.K., Lemon, S.M., Maniloff, J., Mayo, M.A., McGeoch, D.J., Pringle, C.R., et al., Eds.; Academic Press: San Diego, CA, USA, 2000; pp. 747–755. [Google Scholar]
- Schneemann, A.; Ball, L.A.; Delsert, C.; Johnson, J.E.; Nishizawa, T. Nodaviridae. In Virus Taxonomy, Eighth Report of the International Committee on Taxonomy of Viruses; Fauquet, C.M., Mayo, M.A., Maniloff, J., Desselberger, U., Ball, L.A., Eds.; Elsevier/Academic Press: London, UK, 2005. [Google Scholar]
- Frerichs, G.N.; Rodger, H.D.; Peric, Z. Cell culture isolation of piscine neuropathy nodavirus from juvenile sea bass, Dicentrarchus labrax. J. Gen. Virol. 1996, 77, 2067–2071. [Google Scholar] [CrossRef]
- Delsert, C.; Morin, N.; Comps, M. A fish encephalitis virus that differs from other nodaviruses by its capsid protein processing. Arch. Virol. 1997, 142, 2359–2371. [Google Scholar] [CrossRef]
- Nagai, T.; Nishizawa, T. Sequence of the non-structural protein gene encoded by RNA1 of striped jack nervous necrosis virus. J. Gen. Virol. 1999, 80, 3019–3022. [Google Scholar] [CrossRef]
- Iwamoto, T.; Mise, K.; Takeda, A.; Okinaka, Y.; Mori, K.I.; Arimoto, M.; Okuno, T.; Nakai, T. Characterization of striped jack nervous necrosis virus subgenomic RNA3 and biological activities of its encoded protein B2. J. Gen. Virol. 2005, 86, 2807–2816. [Google Scholar] [CrossRef]
- Sommerset, I.; Nerland, A. Complete sequence of RNA1 and subgenomic RNA3 of Atlantic halibut nodavirus (AHNV). Dis. Aquat. Organ. 2004, 58, 117–125. [Google Scholar] [CrossRef]
- Tan, C.; Huang, B.; Chang, S.F.; Ngoh, G.H.; Munday, B.; Chen, S.C.; Kwang, J. Determination of the complete nucleotide sequences of RNA1 and RNA2 from greasy grouper (Epinephelus tauvina) nervous necrosis virus, Singapore strain. J. Gen. Virol. 2001, 82, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, T.; Mori, K.; Furuhashi, M.; Nakai, T.; Furusawa, I.; Muroga, K. Comparison of the coat protein genes of five fish nodaviruses, the causative agents of viral nervous necrosis in marine fish. J. Gen. Virol. 1995, 76, 1563–1569. [Google Scholar] [CrossRef] [PubMed]
- Hegde, A.; Teh, H.C.; Lam, T.J.; Sin, Y.M. Nodavirus infection in freshwater ornamental fish, guppy, Poicelia reticulata—Comparative characterization and pathogenicity studies. Arch. Virol. 2003, 148, 575–586. [Google Scholar] [CrossRef] [PubMed]
- Venter, P.A.; Schneemann, A. Recent insights into the biology and biomedical applications of Flock House virus. Cell. Mol. Life Sci. 2008, 65, 2675–2687. [Google Scholar] [CrossRef] [PubMed]
- Krondiris, J.V.; Sideris, D.C. Intramolecular disulfide bonding is essential for betanodavirus coat protein conformation. J. Gen. Virol. 2002, 83, 2211–2214. [Google Scholar] [CrossRef]
- Wang, C.-H.; Hsu, C.-H.; Wu, Y.-M.; Luo, Y.-C.; Tu, M.-H.; Chang, W.; Cheng, R.H.; Lin, C.-S. Roles of cysteines Cys115 and Cys201 in the assembly and thermostability of grouper betanodavirus particles. Virus Genes 2010, 41, 73–80. [Google Scholar] [CrossRef][Green Version]
- Chen, N.C.; Yoshimura, M.; Guan, H.H.; Wang, T.Y.; Misumi, Y.; Lin, C.C.; Chuankhayan, P.; Nakagawa, A.; Chan, S.I.; Tsukihara, T.; et al. Crystal Structures of a Piscine Betanodavirus: Mechanisms of Capsid Assembly and Viral Infection. PLoS Pathog. 2015, 11, 1–25. [Google Scholar] [CrossRef]
- Wu, Y.M.; Hsu, C.H.; Wang, C.H.; Liu, W.; Chang, W.H.; Lin, C.S. Role of the DxxDxD motif in the assembly and stability of betanodavirus particles. Arch. Virol. 2008, 153, 1633–1642. [Google Scholar] [CrossRef]
- Guo, Y.X.; Dallmann, K.; Kwang, J. Identification of nucleolus localization signal of betanodavirus GGNNV protein α. Virology 2003, 306, 225–235. [Google Scholar] [CrossRef]
- Guo, Y.X.; Wei, T.; Dallmann, K.; Kwang, J. Induction of caspase-dependent apoptosis by betanodaviruses GGNNV and demonstration of protein α as an apoptosis inducer. Virology 2003, 308, 74–82. [Google Scholar] [CrossRef]
- Wu, H.-C.C.; Chiu, C.-S.S.; Wu, J.-L.L.; Gong, H.-Y.Y.; Chen, M.-C.C.; Lu, M.-W.W.; Hong, J.-R.R. Zebrafish anti-apoptotic protein zfBcl-xL can block betanodavirus protein α-induced mitochondria-mediated secondary necrosis cell death. Fish Shellfish Immunol. 2008, 24, 436–449. [Google Scholar] [CrossRef] [PubMed]
- Okinaka, Y.; Nakai, T. Comparisons among the complete genomes of four betanodavirus genotypes. Dis. Aquat. Organ. 2008, 80, 113–121. [Google Scholar] [CrossRef]
- Chen, L.J.; Su, Y.C.; Hong, J.R. Betanodavirus non-structural protein B1: A novel anti-necrotic death factor that modulates cell death in early replication cycle in fish cells. Virology 2009, 385, 444–454. [Google Scholar] [CrossRef]
- Su, Y.-C.; Reshi, L.; Chen, L.-J.; Li, W.-H.; Chiu, H.-W.; Hong, J.-R. Nuclear targeting of the betanodavirus B1 protein via two arginine-rich domains induces G1/S cell cycle arrest mediated by upregulation of p53/p21. Sci. Rep. 2018, 8, 3079. [Google Scholar] [CrossRef]
- Fenner, B.J.; Thiagarajan, R.; Chua, H.K.; Kwang, J. Betanodavirus B2 is an RNA interference antagonist that facilitates intracellular viral RNA accumulation. J. Virol. 2006, 80, 85–94. [Google Scholar] [CrossRef]
- Chen, S.P.; Wu, J.L.; Su, Y.C.; Hong, J.R. Anti-Bcl-2 family members, zfBcl-xL and zfMcl-1a, prevent cytochrome c release from cells undergoing betanodavirus-induced secondary necrotic cell death. Apoptosis 2007, 12, 1043–1060. [Google Scholar] [CrossRef]
- Su, Y.C.; Wu, J.L.; Hong, J.R. Betanodavirus non-structural protein B2: A novel necrotic death factor that induces mitochondria-mediated cell death in fish cells. Virology 2009, 385, 143–154. [Google Scholar] [CrossRef]
- Su, Y.C.; Hong, J.R. Betanodavirus B2 causes ATP depletion-induced cell death via mitochondrial targeting and complex II inhibition in vitro and in vivo. J. Biol. Chem. 2010, 285, 39801–39810. [Google Scholar] [CrossRef]
- Guo, Y.X.; Chan, S.-W.; Kwang, J. Membrane association of greasy grouper nervous necrosis virus protein A and characterization of its mitochondrial localization targeting signal. J. Virol. 2004, 78, 6498–6508. [Google Scholar] [CrossRef]
- Mézeth, K.B.; Nylund, S.; Henriksen, H.; Patel, S.; Nerland, A.H.; Szilvay, A.M. RNA-dependent RNA polymerase from Atlantic Halibut Nodavirus contains two signals for localization to the mitochondria. Virus Res. 2007, 130, 43–52. [Google Scholar] [CrossRef]
- Panzarin, V.; Cappellozza, E.; Mancin, M.; Milani, A.; Toffan, A.; Terregino, C.; Cattoli, G. In vitro study of the replication capacity of the RGNNV and the SJNNV betanodavirus genotypes and their natural reassortants in response to temperature. Vet. Res. 2014, 45, 1–11. [Google Scholar] [CrossRef]
- Souto, S.; Salgado, L.V.; Olveira, J.G.; Bandín, I. Amino acidic substitutions in the polymerase N-terminal region of a reassortant betanodavirus strain causing poor adaptation to temperature increase. Vet. Res. 2019, 50, 1–11. [Google Scholar] [CrossRef]
- Rosskopf, J.J.; Upton, J.H.; Rodarte, L.; Romero, T.A.; Leung, M.-Y.; Taufer, M.; Johnson, K.L. A 3’ terminal stem–loop structure in Nodamura virus RNA2 forms an essential cis-acting signal for RNA replication. Virus Res. 2010, 150, 12–21. [Google Scholar] [CrossRef]
- Taufer, M.; Leung, M.-Y.; Solorio, T.; Licon, A.; Mireles, D.; Araiza, R.; Johnson, K.L. RNAVLab: A virtual laboratory for studying RNA secondary structures based on grid computing technology. Parallel Comput. 2008, 34, 661–680. [Google Scholar] [CrossRef]
- Souto, S.; Olveira, J.G.; Dopazo, C.P.; Borrego, J.J.; Bandín, I. Modification of betanodavirus virulence by substitutions in the 3’ terminal region of RNA2. J. Gen. Virol. 2018, 99, 1210–1220. [Google Scholar] [CrossRef]
- Kim, J.-O.O.; Kim, W.-S.S.; Oh, M.-J.J. Investigation of nervous necrosis virus (NNV) replication in vitro using RNA in situ hybridization. Virus Res. 2019, 260, 78–85. [Google Scholar] [CrossRef]
- Fenner, B.J.; Goh, W.; Kwang, J. Sequestration and Protection of Double-Stranded RNA by the Betanodavirus B2 Protein. J. Virol. 2006, 80, 6822–6833. [Google Scholar] [CrossRef]
- Nishizawa, T.; Furuhashi, M.; Nagai, T.; Nakai, T.; Muroga, K. Genomic classification of fish nodaviruses by molecular phylogenetic analysis of the coat protein gene. Appl. Environ. Microbiol. 1997, 63, 1633–1636. [Google Scholar] [CrossRef]
- Johansen, R.; Sommerset, I.; Torud, B.; Korsnes, K.; Hjortaas, M.J.; Nilsen, F.; Nerland, A.H.; Dannevig, B.H. Characterization of nodavirus and viral encephalopathy and retinopathy in farmed turbot, Scophthalmus maximus (L.). J. Fish Dis. 2004, 27, 591–601. [Google Scholar] [CrossRef]
- Gagné, N.; Johnson, S.C.; Cook-Versloot, M.; MacKinnon, A.M.; Olivier, G. Molecular detection and characterization of nodavirus in several marine fish species from the northeastern Atlantic. Dis. Aquat. Organ. 2004, 62, 181–189. [Google Scholar] [CrossRef]
- Kim, Y.C.; Kwon, W.J.; Min, J.G.; Kim, K., II; Jeong, H.D. Complete genome sequence and pathogenic analysis of a new betanodavirus isolated from shellfish. J. Fish Dis. 2019, 42, 1–13. [Google Scholar] [CrossRef]
- Moody, N.J.G.; Horwood, P.F.; Reynolds, A.; Mahony, T.J.; Anderson, I.G.; Oakey, H.J. Phylogenetic analysis of betanodavirus isolates from Australian finfish. Dis. Aquat. Organ. 2009, 87, 151–160. [Google Scholar] [CrossRef]
- Nylund, A.; Karlsbakk, E.; Nylund, S.; Isaksen, T.E.; Karlsen, M.; Korsnes, K.; Handeland, S.; Martinsen, R.; Mork Pedersen, T.; Ottem, K.F. New betanodaviruses detected in wild and farmed cod (Gadus morhua) in Norway. Arch. Virol. 2008, 153, 541–547. [Google Scholar] [CrossRef]
- Binesh, C.P.; Greeshma, C. Genomic classification of betanodavirus by molecular phylogenetic analysis of the coat protein gene. Arch. Virol. 2013, 158, 1589–1594. [Google Scholar] [CrossRef]
- Keawcharoen, J.; Techangamsuwan, S.; Ponpornpisit, A.; Lombardini, E.D.; Patchimasiri, T.; Pirarat, N. Genetic characterization of a betanodavirus isolated from a clinical disease outbreak in farm-raised tilapia Oreochromis niloticus (L.) in Thailand. J. Fish Dis. 2015, 38, 49–54. [Google Scholar] [CrossRef]
- Korsnes, K.; Karlsbakk, E.; Skår, C.K.; Sælemyr, L.; Nylund, A.; Kvamme, B.O.; Mortensen, S. High nervous necrosis virus (NNV) diversity in wild wrasse (Labridae) in Norway and Sweden. Dis. Aquat. Organ. 2017, 126, 43–50. [Google Scholar] [CrossRef]
- NaveenKumar, S.; Shekar, M.; Karunasagar, I.; Karunasagar, I. Genetic analysis of RNA1 and RNA2 of Macrobrachium rosenbergii nodavirus (MrNV) isolated from India. Virus Res. 2013, 173, 377–385. [Google Scholar] [CrossRef]
- Ho, K.L.; Gabrielsen, M.; Beh, P.L.; Kueh, C.L.; Thong, Q.X.; Streetley, J.; Tan, W.S.; Bhella, D. Structure of the Macrobrachium rosenbergii nodavirus: A new genus within the Nodaviridae? PLoS Biol. 2018, 16, e3000038. [Google Scholar] [CrossRef]
- Thiéry, R.; Cozien, J.; de Boisséson, C.; Kerbart-Boscher, S.; Névarez, L. Genomic classification of new betanodavirus isolates by phylogenetic analysis of the coat protein gene suggests a low host-fish species specificity. J. Gen. Virol. 2004, 85, 3079–3087. [Google Scholar] [CrossRef]
- Cutrín, J.M.; Dopazo, C.P.; Thiéry, R.; Leao, P.; Olveira, J.G.; Barja, J.L.; Bandín, I. Emergence of pathogenic betanodaviruses belonging to the SJNNV genogroup in farmed fish species from the Iberian Peninsula. J. Fish Dis. 2007, 30, 225–232. [Google Scholar] [CrossRef]
- Olveira, J.G.; Souto, S.; Dopazo, C.P.; Thiéry, R.; Barja, J.L.; Bandín, I. Comparative analysis of both genomic segments of betanodaviruses isolated from epizootic outbreaks in farmed fish species provides evidence for genetic reassortment. J. Gen. Virol. 2009, 90, 2940–2951. [Google Scholar] [CrossRef]
- Panzarin, V.; Fusaro, A.; Monne, I.; Cappellozza, E.; Patarnello, P.; Bovo, G.; Capua, I.; Holmes, E.C.; Cattoli, G. Molecular epidemiology and evolutionary dynamics of betanodavirus in southern Europe. Infect. Genet. Evol. 2012, 12, 63–70. [Google Scholar] [CrossRef]
- Toffan, A.; Pascoli, F.; Pretto, T.; Panzarin, V.; Abbadi, M.; Buratin, A.; Quartesan, R.; Gijon, D.; Padros, F. Viral nervous necrosis in gilthead sea bream (Sparus aurata) caused by reassortant betanodavirus RGNNV/SJNNV: An emerging threat for Mediterranean aquaculture. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Bitchava, K.; Chassalevris, T.; Lampou, E.; Athanassopoulou, F.; Economou, V.; Dovas, C.I. Occurrence and molecular characterization of betanodaviruses in fish and invertebrates of the Greek territorial waters. J. Fish Dis. 2019, 42, 1773–1783. [Google Scholar] [CrossRef]
- Mori, K.; Mangyoku, T.; Iwamoto, T.; Arimoto, M.; Tanaka, S.; Nakai, T. Serological relationships among genotypic variants of betanodavirus. Dis. Aquat. Organ. 2003, 57, 19–26. [Google Scholar] [CrossRef]
- Panzarin, V.; Toffan, A.; Abbadi, M.; Buratin, A.; Mancin, M.; Braaen, S.; Olsen, C.M.; Bargelloni, L.; Rimstad, E.; Cattoli, G. Molecular basis for antigenic diversity of genus Betanodavirus. PLoS ONE 2016, 11, 1–18. [Google Scholar] [CrossRef]
- Barker, D.E.; MacKinnon, A.M.; Boston, L.; Burt, M.D.B.; Cone, D.K.; Speare, D.J.; Griffiths, S.; Cook, M.; Ritchie, R.; Olivier, G. First report of piscine nodavirus infecting wild winter flounder Pleuronectes americanus in Passamaquoddy Bay, New Brunswick, Canada. Dis. Aquat. Organ. 2002, 49, 99–105. [Google Scholar] [CrossRef]
- Ciulli, S.; Galletti, E.; Grodzki, M.; Alessi, A.; Battilani, M.; Prosperi, S. Isolation and genetic characterization of Betanodavirus from wild marine fish from the Adriatic Sea. Vet. Res. Commun. 2007, 31, 221–224. [Google Scholar] [CrossRef]
- Gomez, D.K.; Mori, K.; Okinaka, Y.; Nakai, T.; Park, S.C. Trash fish can be a source of betanodaviruses for cultured marine fish. Aquaculture 2010, 302, 158–163. [Google Scholar] [CrossRef]
- Haddad-Boubaker, S.; Boughdir, W.; Sghaier, S.; Souissi, J.B.; Megdich, A.; Dhaouadi, R.; Amara, A.; Panzarin, V.; Fakhfakh, E. Outbreak of viral nervous necrosis in endangered fish species Epinephelus costae and E. Marginatus in northern Tunisian coasts. Fish Pathol. 2014, 49, 53–56. [Google Scholar] [CrossRef]
- Haddad-Boubaker, S.; Bigarré, L.; Bouzgarou, N.; Megdich, A.; Baud, M.; Cabon, J.; Chéhida, N.B. Molecular epidemiology of betanodaviruses isolated from sea bass and sea bream cultured along the Tunisian coasts. Virus Genes 2013, 46, 412–422. [Google Scholar] [CrossRef]
- Moreno, P.; Olveira, J.G.; Labella, A.; Cutrín, J.M.; Baro, J.C.; Borrego, J.J.; Dopazo, C.P. Surveillance of viruses in wild fish populations in areas around the Gulf of Cadiz (South Atlantic Iberian Peninsula). Appl. Environ. Microbiol. 2014, 80, 6560–6571. [Google Scholar] [CrossRef]
- Cherif, N.; Fatma, A. Nodaviruses in wild fish population collected around aquaculture cage sites from coastal areas of tunisia. Fish. Aquac. J. 2017, 8, 2–7. [Google Scholar] [CrossRef]
- Gomez, D.K.; Okinaka, Y.; Nakai, T.; Sato, J.; Mushiake, K.; Isshiki, T. PCR-based detection of betanodaviruses from cultured and wild marine fish with no clinical signs. J. Fish Dis. 2004, 27, 603–608. [Google Scholar] [CrossRef]
- Sakamoto, T.; Okinaka, Y.; Mori, K.-I.; Sugaya, T.; Nishioka, T.; Oka, M.; Yamashita, H.; Nakai, T. Phylogenetic analysis of betanodavirus RNA2 identified from wild marine fish in oceanic regions. Fish Pathol. 2008, 43, 19–27. [Google Scholar] [CrossRef][Green Version]
- Ma, H.; Wen, W.; Su, Y.; Feng, J.; Xu, L.; Peng, C.; Guo, Z. Epidemiological characterization of VNNV in hatchery-reared and wild marine fish on Hainan Island, China, and experimental infection of golden pompano (Trachinotus ovatus) juveniles. Arch. Virol. 2015, 160, 2979–2989. [Google Scholar] [CrossRef]
- Liu, X.D.L.; Huang, J.N.; Weng, S.P.; Hu, X.Q.; Chen, W.J.; Qin, Z.D.; Dong, X.X.; Liu, X.D.L.; Zhou, Y.; Asim, M.; et al. Infections of nervous necrosis virus in wild and cage-reared marine fish from South China Sea with unexpected wide host ranges. J. Fish Dis. 2015, 38, 533–540. [Google Scholar] [CrossRef]
- Nishioka, T.; Sugaya, T.; Kawato, Y.; Mori, K.; Nakai, T. Pathogenicity of striped jack nervous necrosis virus (SJNNV) Isolated from asymptomatic wild Japanese jack mackerel Trachurus japonicus. Fish Pathol. 2016, 51, 176–183. [Google Scholar] [CrossRef][Green Version]
- Curtis, P.A.; Drawbridge, M.; Iwamoto, T.; Nakai, T.; Hedrick, R.P.; Gendron, A.P. Nodavirus infection of juvenile white seabass, Atractoscion nobilis, cultured in southern California: First record of viral nervous necrosis (VNN) in North America. J. Fish Dis. 2001, 24, 263–271. [Google Scholar] [CrossRef]
- Munday, B.L.; Kwang, J.; Moody, N. Review article Betanodavirus infections of teleost fish: A review. J. Fish Dis. 2002, 25, 127–142. [Google Scholar] [CrossRef]
- Yoshikoshi, K.; Inoue, K. Viral nervous necrosis in hatchery-reared larvae and juveniles of Japanese parrotfish, Oplegnathus fasciatus (Temminck & Schlegel). J. Fish Dis. 1990, 13, 69–77. [Google Scholar]
- Muroga, K. Viral and bacterial diseases in larval and juvenile marine fish and shellfish: A review. Fish Pathol. 1995, 30, 71–85. [Google Scholar] [CrossRef]
- Skliris, G.P.; Krondiris, J.V.; Sideris, D.C.; Shinn, A.P.; Starkey, W.G.; Richards, R.H. Phylogenetic and antigenic characterization of new fish nodavirus isolates from Europe and Asia. Virus Res. 2001, 75, 59–67. [Google Scholar] [CrossRef]
- Johnson, S.C.; Sperker, S.A.; Leggiadro, C.T.; Groman, D.B.; Griffiths, S.G.; Ritchie, R.J.; Cook, M.D.; Cusack, R.R. Identification and characterization of a piscine neuropathy and nodavirus from juvenile Atlantic cod from the Atlantic coast of North America. J. Aquat. Anim. Health 2002, 14, 124–133. [Google Scholar] [CrossRef]
- Mao, M.-G.; Wen, S.-H.; Perálvarez-Marín, A.; Li, H.; Jiang, J.-L.; Jiang, Z.-Q.; Li, X.; Sun, H.; Lü, H.-Q. Evidence for and characterization of nervous necrosis virus infection in Pacific cod (Gadus macrocephalus). Arch. Virol. 2015, 160, 2237–2248. [Google Scholar] [CrossRef]
- Sethi, S.N.; Vinod, K.; Rudhramurthy, N.; Kokane, M.R.; Pattnaik, P. Detection of betanodavirus in wild caught fry milk fish, Chanos chanos, (Lacepeds 1803). Indian J. Geo Mar. Sci. 2018, 47, 1620–1624. [Google Scholar]
- Ucko, M.; Colorni, A.; Diamant, A. Nodavirus infections in Israeli mariculture. J. Fish Dis. 2004, 27, 459–469. [Google Scholar] [CrossRef]
- Zorriehzahra, M.E.J.; Ghasemi, M.; Ghiasi, M.; Karsidani, S.H.; Bovo, G.; Nazari, A.; Adel, M.; Arizza, V.; Dhama, K. Isolation and confirmation of viral nervous necrosis (VNN) disease in golden grey mullet (Liza aurata) and leaping mullet (Liza saliens) in the Iranian waters of the Caspian Sea. Vet. Microbiol. 2016, 190, 27–37. [Google Scholar] [CrossRef]
- Sharma, S.R.K.; Pradeep, M.A.; Dube, P.N.; Kumar, T.V.A.; Kumar, R.; Swaminathan, T.R. Betanodavirus-associated mortality in Asian seabass (Lates calcarifer, Bloch) cultured in indoor tanks and sea cages. Aquac. Int. 2019, 27, 279–286. [Google Scholar] [CrossRef]
- Chi, S.C.; Shieh, J.R.; Lin, S.J. Genetic and antigenic analysis of betanodaviruses isolated from aquatic organisms in Taiwan. Dis. Aquat. Organ. 2003, 55, 221–228. [Google Scholar] [CrossRef]
- Chi, S.; Lee, K.; Hwang, S. Investigation of Host Range of Fish Nodavirus in Taiwan. In Proceedings of the Tenth International Conference on Diseases of Fish and Shellfish, Dublin, Ireland, 9–14 September 2001; European Association of Fish Pathologist: Dublin, Ireland, 2001; p. P-O-049. [Google Scholar]
- Ransangan, J.; Manin, B.O. Mass mortality of hatchery-produced larvae of Asian seabass, Lates calcarifer (Bloch), associated with viral nervous necrosis in Sabah, Malaysia. Vet. Microbiol. 2010, 145, 153–157. [Google Scholar] [CrossRef]
- Jung, S.J.; Miyazaki, T.; Miyata, M.; Oishi, T. Histopathological studies on viral nervous necrosis in a new host, Japanese sea bass Lateolabrax japonicus. Bullettin Fac. Bioresour. 1996, 16, 9–16. [Google Scholar]
- David, R.; Tréguier, C.; Montagnani, C.; Belliard, C.; Levy, P.; Nédélec, G.; Joufoques, V.; Remoissenet, G.; Gueguen, Y.; Cochennec-Laureau, N. Molecular detection of betanodavirus from the farmed fish, Platax orbicularis (Forsskal) (Ephippidae), in French Polynesia. J. Fish Dis. 2010, 33, 451–454. [Google Scholar] [CrossRef]
- Ransangan, J.; Manin, B.O.; Abdullah, A.; Roli, Z.; Sharudin, E.F. Betanodavirus infection in golden pompano, Trachinotus blochii, fingerlings cultured in deep-sea cage culture facility in Langkawi, Malaysia. Aquaculture 2011, 315, 327–334. [Google Scholar] [CrossRef]
- Li, P.; Yu, Q.; Li, F.; Qin, X.; Dong, D.; Chen, B.; Qin, Q. First identification of the nervous necrosis virus isolated from cultured golden pompano (Trachinotus ovatus) in Guangxi, China. J. Fish Dis. 2018, 41, 1177–1180. [Google Scholar] [CrossRef]
- Abdullah, A.; Ramli, R.; Ridzuan, M.S.M.; Murni, M.; Hashim, S.; Sudirwan, F.; Abdullah, S.Z.; Mansor, N.N.; Amira, S.; Saad, M.Z.; et al. The presence of Vibrionaceae, Betanodavirus and Iridovirus in marine cage-cultured fish: Role of fish size, water physicochemical parameters and relationships among the pathogens. Aquac. Rep. 2017, 7, 57–65. [Google Scholar] [CrossRef]
- Toffolo, V.; Negrisolo, E.; Maltese, C.; Bovo, G.; Belvedere, P.; Colombo, L.; Valle, L.D. Phylogeny of betanodaviruses and molecular evolution of their RNA polymerase and coat proteins. Mol. Phylogenet. Evol. 2007, 43, 298–308. [Google Scholar] [CrossRef]
- Bovo, G.; Nishizawa, T.; Maltese, C.; Borghesan, F.; Mutinelli, F.; Montesi, F.; De Mas, S. Viral encephalopathy and retinopathy of farmed marine fish species in Italy. Virus Res. 1999, 63, 143–146. [Google Scholar] [CrossRef]
- Le Breton, A.; Grisez, L.; Sweetman, J.; Ollevier, F. Viral nervous necrosis (VNN) associated with mass mortalities in cage-reared sea bass, Dicentrarchus labrax (L.). J. Fish Dis. 1997, 20, 145–151. [Google Scholar] [CrossRef]
- Athanassopoulou, F.; Billinis, C.; Psychas, V.; Karipoglou, K. Viral encephalopathy and retinopathy of Dicentrarchus labrax (L.) farmed in fresh water in Greece. J. Fish Dis. 2003, 26, 361–365. [Google Scholar] [CrossRef]
- Vendramin, N.; Patarnello, P.; Toffan, A.; Panzarin, V.; Cappellozza, E.; Tedesco, P.; Terlizzi, A.; Terregino, C.; Cattoli, G. Viral Encephalopathy and Retinopathy in groupers (Epinephelus spp.) in southern Italy: A threat for wild endangered species? BMC Vet. Res. 2013, 9, 20. [Google Scholar] [CrossRef]
- Binesh, C.P.; Renuka, K.; Malaichami, N.; Greeshma, C. First report of viral nervous necrosis-induced mass mortality in hatchery-reared larvae of clownfish, Amphiprion sebae Bleeker. J. Fish Dis. 2013, 36, 1017–1020. [Google Scholar] [CrossRef]
- Oh, M.J.; Jung, S.J.; Kim, S.R.; Rajendran, K.V.; Kim, Y.J.; Choi, T.J.; Kim, H.R.; Kim, J. Do A fish nodavirus associated with mass mortality in hatchery-reared red drum, Sciaenops ocellatus. Aquaculture 2002, 211, 1–7. [Google Scholar] [CrossRef]
- Comps, M.; Trindade, M.; Delsert, C. Investigation of fish encephalitis viruses (FEV) expression in marine fishes using DIG-labelled probes. Aquaculture 1996, 143, 113–121. [Google Scholar] [CrossRef]
- Pavoletti, E.; Prearo, M.; Ghittino, M.; Ghittino, C. Casi di encefaloretinopatia in ombrina (Umbrina cirrosa) con descrizione della sintomatologia clinica e del quadro anatomoistopatologico. Boll. Soc. Patol. Ittica 1998, 23, 24–33. [Google Scholar]
- Sugaya, T.; Mori, K.; Nishioka, T.; Masuma, S.; Oka, M.; Mushiake, K.; Okinaka, Y.; Nakai, T. Genetic heterogeneity of betanodaviruses in juvenile production trials of Pacific bluefin tuna, Thunnus orientalis (Temminck & Schlegel). J. Fish Dis. 2009, 32, 815–824. [Google Scholar]
- Chi, S.C.; Lo, C.F.; Kou, G.H.; Chang, P.S.; Peng, S.E.; Chen, S.N. Mass mortalities associated with viral nervous necrosis (VNN) disease in two species of hatchery-reared grouper, Epinephelus fuscogutatus and Epinephelus akaara (Temminck & Schlegel). J. Fish Dis. 1997, 20, 185–193. [Google Scholar]
- Mori, K.; Nakai, T.; Nagahara, M.; Muroga, K.; Mekuchi, T.; Kanno, T. A viral disease in hatchery-reared larvae and juveniles of redspotted grouper. Fish Pathol. 1991, 26, 209–210. [Google Scholar] [CrossRef]
- Lai, Y.-S.S.; Murali, S.; Chiu, H.-C.C.; Ju, H.-Y.Y.; Lin, Y.-S.S.; Chen, S.-C.C.; Guo, I.-C.C.; Fang, K.; Chang, C.-Y.Y. Propagation of yellow grouper nervous necrosis virus (YGNNV) in a new nodavirus-susceptible cell line from yellow grouper, Epinephelus awoara (Temminck & Schlegel), brain tissue. J. Fish Dis. 2001, 24, 299–309. [Google Scholar]
- Maeno, Y.; de la Pena, L.D.; Cruz-Lacierda, E.R. Nodavirus infection in hatchery-reared orange-spotted grouper Epinephelus coioides: First record of viral nervous necrosis in the Philippines. Fish Pathol. 2002, 37, 87–89. [Google Scholar] [CrossRef][Green Version]
- Kara, H.M.; Chaoui, L.; Derbal, F.; Zaidi, R.; de Boisséson, C.; Baud, M.; Bigarré, L. Betanodavirus-associated mortalities of adult wild groupers Epinephelus marginatus (Lowe) and Epinephelus costae (Steindachner) in Algeria. J. Fish Dis. 2014, 37, 273–278. [Google Scholar] [CrossRef]
- Lin, C.S.; Lu, M.W.; Tang, L.; Liu, W.; Chao, C.B.; Lin, C.J.; Krishna, N.K.; Johnson, J.E.; Schneemann, A. Characterization of virus-like particles assembled in a recombinant baculovirus system expressing the capsid protein of a fish nodavirus. Virology 2001, 290, 50–58. [Google Scholar] [CrossRef]
- Agnihotri, K.; Pease, B.; Chong, R. Molecular analysis of RNA1 and RNA2 sequences from a betanodavirus isolated from giant grouper (Epinephelus lanceolatus) in Australia. Virol. Rep. 2016, 6, 25–31. [Google Scholar] [CrossRef]
- Valencia, J.; Grau, A.; Pretto, T.; Pons, J.; Jurado-Rivera, J.; Castro, J.; Toffan, A.; Catanese, G. Viral encephalopathy and retinopathy (VER) disease in Epinephelus marginatus from the Balearic Islands marine protected areas. Dis. Aquat. Organ. 2019, 135, 49–58. [Google Scholar] [CrossRef]
- Nakai, T.; Dung, N.H.; Nishizawa, T.; Muroga, K.; Arimoto, M.; Ootsuki, K. Occurrence of viral nervous necrosis in kelp grouper and tiger puffer. Fish Pathol. 1994, 29, 211–212. [Google Scholar] [CrossRef][Green Version]
- Fukuda, Y.; Nguyen, H.D.; Furuhashi, M.; Nakai, T. Mass Mortality of Cultured Sevenband Grouper, Epinephelus septemfasciatus, Associated with Viral Nervous Necrosis. Fish Pathol. 1996, 31, 165–170. [Google Scholar] [CrossRef]
- Sohn, S.-G.; Park, M.-A.; Oh, M.-J.; Chun, S.-K. A Fish Nodavirus Isolated from Cultured Sevenband Groupe, Epinephelus septemfasciatus. J. Fish Pathol. 1998, 11, 97–104. [Google Scholar]
- Chua, F.; Loo, J.; Wee, J. Mass mortality in juvenile greasy grouper, Epinephelus tauvina, associated with vacuolating encephalopathy and retinopathy. In Dis Asian Aquacult; Fish Health Section, Asian Fisheries Society: Manila, Philippines, 1995; Volume II, pp. 235–241. [Google Scholar]
- Hegde, A.; Chen, C.L.; Qin, Q.W.; Lam, T.J.; Sin, Y.M. Characterization, pathogenicity and neutralization studies of a nervous necrosis virus isolated from grouper, Epinephelus tauvina, in Singapore. Aquaculture 2002, 213, 55–72. [Google Scholar] [CrossRef]
- Khumaidi, A.; Fadjar, M.; Iranawati, F.; Kilawati, Y.; Yanuhar, U. Mass Mortality Associated with Viral Nervous Necrosis of Hybrid Grouper (Epinephelus sp.) Cultured in City of Grouper. In Proceedings of the International Conference on Biology and Applied Science (ICOBAS), Malang, Indonesia, 13–14 March 2019; AIP Publishing LLC: Melville, NY, USA, 2019; Volume 2120, p. 070007. [Google Scholar]
- Knibb, W.; Luu, G.; Premachandra, H.K.A.; Lu, M.-W.; Nguyen, N.H. Regional genetic diversity for NNV grouper viruses across the Indo-Asian region—Implications for selecting virus resistance in farmed groupers. Sci. Rep. 2017, 7, 10658. [Google Scholar] [CrossRef]
- Ransangan, J.; Manin, B.O. Genome analysis of Betanodavirus from cultured marine fish species in Malaysia. Vet. Microbiol. 2012, 156, 16–44. [Google Scholar] [CrossRef]
- Zafran; Koesharyani, I.; Johnny, F.; Yuasa, K.; Harada, T.; Hatai, K. Viral nervous necrosis in humpback grouper Cromileptes altivelis larvae and juveniles in Indonesia. Fish Pathol. 2000, 35, 95–96. [Google Scholar] [CrossRef][Green Version]
- Pirarat, N.; Ponpornpisit, A.; Traithong, T.; Nakai, T.; Katagiri, T.; Maita, M.; Endo, M. Nodavirus associated with pathological changes in adult spotted coralgroupers (Plectropomus maculatus) in Thailand with viral nervous necrosis. Res. Vet. Sci. 2009, 87, 97–101. [Google Scholar]
- NaveenKumar, S.; Hassan, M.A.; Mahmoud, M.A.; Al-Ansari, A.; Al-Shwared, W.K. Betanodavirus infection in reared marine fishes along the Arabian Gulf. Aquac. Int. 2017, 25, 1543–1554. [Google Scholar] [CrossRef]
- Kim, S.R.; Jung, S.J.; Kim, Y.J.; Kim, J.D.; Jung, T.S.; Choi, T.J.; Yoshimizu, M.; Oh, M.J. Phylogenic Comparison of Viral Nervous Necrosis (VNN) Viruses Occurring Seed Production Period. Korean J. Fish. Aquat. Sci. 2002, 35, 237–241. [Google Scholar]
- Grotmol, S.; Totland, G.K.; Kvellestad, A.; Fjell, K.; Olsen, A.B. Mass mortality of larval and juvenile hatchery-reared halibut (Hippoglossus hippoglossus L.) associated with the presence of virus-like particles in vacuolated lesions in the central nervous system and retina. Bull. Eur. Assoc. Fish Pathol. 1995, 15, 176–180. [Google Scholar]
- Grotmol, S.; Nerland, A.H.; Biering, E.; Totland, G.K.; Nishizawa, T. Characterisation of the capsid protein gene from a nodavirus strain affecting the Atlantic halibut Hippoglossus hippoglossus and design of an optimal reverse-transcriptase polymerase chain reaction (RT-PCR) detection assay. Dis. Aquat. Organ. 2000, 39, 79–88. [Google Scholar] [CrossRef]
- Starkey, W.G.; Ireland, J.H.; Muir, K.F.; Shinn, A.; Richards, R.H.; Ferguson, H.W. Isolation of nodavirus from Scottish farmed halibut, Hippoglossus hippoglossus (L). J. Fish Dis. 2000, 23, 418–422. [Google Scholar] [CrossRef]
- Pirarat, N.; Katagiri, T.; Maita, M.; Nakai, T.; Endo, M. Viral encephalopathy and retinopathy in hatchery-reared juvenile thread-sail filefish (Stephanolepis cirrhifer). Aquaculture 2009, 288, 349–352. [Google Scholar] [CrossRef]
- Gomez, D.K.; Lim, D.J.; Baeck, G.W.; Youn, H.J.; Shin, N.S.; Youn, H.Y.; Hwang, C.Y.; Park, J.H.; Park, S.C. Detection of betanodaviruses in apparently healthy aquarium fishes and invertebrates. J. Vet. Sci. 2006, 7, 369–374. [Google Scholar] [CrossRef]
- Kim, Y.C.; Kwon, W.J.; Min, J.G.; Jeong, H.D. Isolation and initial characterization of new betanodaviruses in shellfish. Transbound. Emerg. Dis. 2018, 65, 1557–1567. [Google Scholar] [CrossRef]
- Panzarin, V.; Patarnello, P.; Mori, A.; Rampazzo, E.; Cappellozza, E.; Bovo, G.; Cattoli, G. Development and validation of a real-time TaqMan PCR assay for the detection of betanodavirus in clinical specimens. Arch. Virol. 2010, 155, 1193–1203. [Google Scholar] [CrossRef] [PubMed]
- Castric, J.; Thiéry, R.; Jeffroy, J.; De Kinkelin, P.; Raymond, J.C. Sea bream Sparus aurata, an asymptomatic contagious fish host for nodavirus. Dis. Aquat. Organ. 2001, 47, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Berzak, R.; Scheinin, A.; Davidovich, N.; Regev, Y.; Diga, R.; Tchernov, D.; Morick, D. Prevalence of nervous necrosis virus (NNV) and Streptococcus species in wild marine fish and crustaceans from the Levantine Basin, Mediterranean Sea. Dis. Aquat. Organ. 2019, 133, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.C.; Kwon, W.J.; Kim, M.S.; Kim, K.I.; Min, J.G.; Jeong, H.D. High prevalence of betanodavirus barfin flounder nervous necrosis virus as well as red-spotted grouper nervous necrosis virus genotype in shellfish. J. Fish Dis. 2018, 41, 233–246. [Google Scholar] [CrossRef]
- Olveira, J.G.; Souto, S.; Dopazo, C.P.; Bandín, I. Isolation of betanodavirus from farmed turbot Psetta maxima showing no signs of viral encephalopathy and retinopathy. Aquaculture 2013, 406–407, 125–130. [Google Scholar] [CrossRef]
- Athanassopoulou, F.; Billinis, C.; Prapas, T. Important disease conditions of newly cultured species in intensive freshwater farms in Greece: First incidence of nodavirus infection in Acipenser sp. Dis. Aquat. Organ. 2004, 60, 247–252. [Google Scholar] [CrossRef]
- Jithendran, K.P.; Shekhar, M.S.; Kannappan, S.; Azad, I.S. Nodavirus infection in freshwater ornamental fishes in India-diagnostic histopathology and nested RT-PCR. Asian Fish. Sci. 2011, 24, 12–19. [Google Scholar]
- Vendramin, N.; Padrós, F.; Pretto, T.; Cappellozza, E.; Panzarin, V.; Bovo, G.; Toffan, A.; Terregino, C. Viral encephalopathy and retinopathy outbreak in restocking facilities of the endangered freshwater species, Salaria fluviatilis (Asso). J. Fish Dis. 2012, 35, 867–871. [Google Scholar] [CrossRef]
- Bovo, G.; Gustinelli, A.; Quaglio, F.; Gobbo, F.; Panzarin, V.; Fusaro, A.; Mutinelli, F.; Caffara, M.; Fioravanti, M. Viral encephalopathy and retinopathy outbreak in freshwater fish farmed in Italy. Dis. Aquat. Organ. 2011, 96, 45–54. [Google Scholar] [CrossRef]
- Bigarré, L.; Cabon, J.; Baud, M.; Heimann, M.; Body, A.; Lieffrig, F.; Castric, J. Outbreak of betanodavirus infection in tilapia, Oreochromis niloticus (L.), in fresh water. J. Fish Dis. 2009, 32, 667–673. [Google Scholar] [CrossRef]
- Bandín, I.; Souto, S.; Cutrín, J.M.; López-Vázquez, C.; Olveira, J.G.; Esteve, C.; Alcaide, E.; Dopazo, C.P. Presence of viruses in wild eels Anguilla anguilla L, from the Albufera Lake (Spain). J. Fish Dis. 2014, 37, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Giacopello, C.; Foti, M.; Bottari, T.; Fisichella, V.; Barbera, G. Detection of viral encephalopathy and retinopathy virus (VERV) in wild marine fish species of the South Tyrrhenian Sea (Central Mediterranean). J. Fish Dis. 2013, 36, 819–821. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Jimena, B.; Cherif, N.; Garcia-Rosado, E.; Infante, C.; Cano, I.; Castro, D.; Hammami, S.; Borrego, J.J.; Alonso, M.C. A combined RT-PCR and dot-blot hybridization method reveals the coexistence of SJNNV and RGNNV betanodavirus genotypes in wild meagre (Argyrosomus regius). J. Appl. Microbiol. 2010, 109, 1361–1369. [Google Scholar] [CrossRef] [PubMed]
- Dalla Valle, L.; Zanella, L.; Patarnello, P.; Paolucci, L.; Belvedere, P.; Colombo, L. Development of a sensitive diagnostic assay for fish nervous necrosis virus based on RT-PCR plus nested PCR. J. Fish Dis. 2000, 23, 321–327. [Google Scholar] [CrossRef]
- Nakai, T.; Mori, K.; Sugaya, T.; Nishioka, T.; Mushiake, K.; Yamashita, H. Current knowledge on viral nervous necrosis (VNN) and its causative betanodaviruses. Isr. J. Aquac. 2010, 61, 198–207. [Google Scholar]
- García-Rosado, E.; Cano, I.; Martín-Antonio, B.; Labella, A.; Manchado, M.; Alonso, M.C.; Castro, D.; Borrego, J.J. Co-occurrence of viral and bacterial pathogens in disease outbreaks affecting newly cultured sparid fish. Int. Microbiol. 2007, 10, 193–199. [Google Scholar]
- Maltese, C.; Bovo, G. MONOGRAFIE Viral encephalopathy and retinopathy Encefalopatia e retinopatia virale. Ittiopatologia 2007, 4, 93–146. [Google Scholar]
- Toffan, A.; Panzarin, V.; Toson, M.; Cecchettin, K.; Pascoli, F. Water temperature affects pathogenicity of different betanodavirus genotypes in experimentally challenged Dicentrarchus labrax. Dis. Aquat. Organ. 2016, 119, 231–238. [Google Scholar] [CrossRef]
- Grotmol, S.; Bergh, Ø.; Totland, G.K. Transmission of viral encephalopathy and retinopathy (VER) to yolk-sac larvae of the Atlantic halibut Hippoglossus hippoglossus: Occurrence of nodavirus in various organs and a possible route of infection. Dis. Aquat. Organ. 1999, 36, 95–106. [Google Scholar] [CrossRef]
- Vendramin, N.; Toffan, A.; Mancin, M.; Cappellozza, E.; Panzarin, V.; Bovo, G.; Cattoli, G.; Capua, I.; Terregino, C. Comparative pathogenicity study of ten different betanodavirus strains in experimentally infected European sea bass, Dicentrarchus labrax (L.). J. Fish Dis. 2014, 37, 371–383. [Google Scholar] [CrossRef]
- Souto, S.; Olveira, J.G.; Bandín, I. Influence of temperature on Betanodavirus infection in Senegalese sole (Solea senegalensis). Vet. Microbiol. 2015, 179, 162–167. [Google Scholar] [CrossRef]
- Tanaka, S.; Aoki, H.; Nakai, T. Pathogenicity of the nodavirus detected from diseased sevenband grouper Epinephelus septemfasciatus. Fish Pathol. 1998, 33, 31–36. [Google Scholar] [CrossRef]
- Totland, G.K.; Grotmol, S.; Morita, Y.; Nishioka, T.; Nakai, T. Pathogenicity of nodavirus strains from striped jack Pseudocaranx dentex and Atlantic halibut Hippoglossus hippoglossus, studied by waterborne challenge of yolk-sac larvae of both teleost species. Dis. Aquat. Organ. 1999, 38, 169–175. [Google Scholar] [CrossRef]
- Hata, N.; Okinaka, Y.; Iwamoto, T.; Kawato, Y.; Mori, K.I.; Nakai, T. Identification of RNA regions that determine temperature sensitivities in betanodaviruses. Arch. Virol. 2010, 155, 1597–1606. [Google Scholar] [CrossRef]
- de Silva, S.S.; Soto, D. Climate change and aquaculture: Potential impacts, adaptation and migraton. In Climate Change Implications for Fisheries and Aquaculture: Overview of Current Scientific Knowledge; Cochrane, K., de Young, C., Soto, D., Bahri, T., Eds.; FAO: Rome, Italy, 2009; pp. 151–212. [Google Scholar]
- Slenning, B.D. Global Climate Change and Implications for Disease Emergence. Vet. Pathol. 2010, 47, 28–33. [Google Scholar] [CrossRef]
- Bett, B.; Kiunga, P.; Gachohi, J.; Sindato, C.; Mbotha, D.; Robinson, T.; Lindahl, J.; Grace, D. Effects of climate change on the occurrence and distribution of livestock diseases. Prev. Vet. Med. 2017, 137, 119–129. [Google Scholar] [CrossRef]
- Brand, S.P.C.; Keeling, M.J. The impact of temperature changes on vector-borne disease transmission: Culicoides midges and bluetongue virus. J. R. Soc. Interface 2017, 14. [Google Scholar] [CrossRef]
- Niedbalski, W.; Fitzner, A. Impact of climate change on the occurrence and distribution of bluetongue in Europe. Med. Weter. 2018, 74, 634–639. [Google Scholar] [CrossRef]
- Nguyen, H.D.; Nakai, T.; Muroga, K. Progression of striped jack nervous necrosis virus (SJNNV) infection in naturally and experimentally infected striped jack Pseudocaranx dentex larvae. Dis. Aquat. Organ. 1996, 24, 99–105. [Google Scholar] [CrossRef]
- Mladineo, I. The immunohistochemical study of nodavirus changes in larval, juvenile and adult sea bass tissue. J. Appl. Ichthyol. 2003, 19, 366–370. [Google Scholar] [CrossRef]
- Souto, S.; Olveira, J.G.; Alonso, M.C.; Dopazo, C.P.; Bandín, I. Betanodavirus infection in bath-challenged Solea senegalensis juveniles: A comparative analysis of RGNNV, SJNNV and reassortant strains. J. Fish Dis. 2018, 41, 1571–1578. [Google Scholar] [CrossRef]
- Péducasse, S.; Castric, J.; Thiéry, R.; Jeffroy, J.; Le Ven, A.; Baudin Laurencin, F. Comparative study of viral encephalopathy and retinopathy in juvenile sea bass Dicentrarchus labrax infected in different ways. Dis. Aquat. Organ. 1999, 36, 11–20. [Google Scholar] [CrossRef][Green Version]
- Grotmol, S.; Totland, G.K.; Thorud, K.; Hjeltnes, B.K. Vacuolating encephalopathy and retinopathy associated with a nodavirus-like agent: A probable cause of mass mortality of cultured larval and juvenile Atlantic halibut Hippoglossus hippoglossus. Dis. Aquat. Org. 1997, 29, 85–97. [Google Scholar] [CrossRef][Green Version]
- Tanaka, S.; Takagi, M.; Miyazaki, T. Histopathological studies on viral nervous necrosis of sevenband grouper, Epinephelus septemfasciatus Thunberg, at the grow-out stage. J. Fish Dis. 2004, 27, 385–399. [Google Scholar] [CrossRef]
- Costa, J.Z.; Thompson, K.D. Understanding the interaction between Betanodavirus and its host for the development of prophylactic measures for viral encephalopathy and retinopathy. Fish Shellfish Immunol. 2016, 53, 35–49. [Google Scholar] [CrossRef]
- Skliris, G.P.; Richards, R.H. Induction of nodavirus disease in seabass, Dicentrarchus labrax, using different infection models. Virus Res. 1999, 63, 85–93. [Google Scholar] [CrossRef]
- Gjessing, M.C.; Kvellestad, A.; Ottesen, K.; Falk, K. Nodavirus provokes subclinical encephalitis and retinochoroiditis in adult farmed Atlantic cod, Gadus morhua L. J. Fish Dis. 2009, 32, 421–431. [Google Scholar] [CrossRef]
- Tanaka, S.; Kuriyama, I.; Nakai, T.; Miyazaki, T. Susceptibility of cultured juveniles of several marine fish to the sevenband grouper nervous necrosis virus. J. Fish Dis. 2003, 26, 109–115. [Google Scholar] [CrossRef]
- Korsnes, K.; Devold, M.; Nerland, A.H.; Nylund, A. Viral encephalopathy and retinopathy (VER) in Atlantic salmon Salmo salar after intraperitoneal challenge with a nodavirus from Atlantic halibut Hippoglossus hippoglossus. Dis. Aquat. Organ. 2005, 68, 7–15. [Google Scholar] [CrossRef]
- Bitchava, K.; Xylouri, E.; Fragkiadaki, E.; Athanassopoulou, F.; Papanastassopoulou, M.; Sabatakou, O. First incidence of clinical signs of nodavirus infection in sea bream, Sparus auratus L. Isr. J. Aquac. Bamidgeh 2007, 59, 3–9. [Google Scholar]
- Huang, R.; Zhu, G.; Zhang, J.; Lai, Y.; Xu, Y.; He, J.; Xie, J. Betanodavirus-like particles enter host cells via clathrin-mediated endocytosis in a cholesterol-, pH- and cytoskeleton-dependent manner. Vet. Res. 2017, 48, 8. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Hsu, C.-H.; Hong, Y.-R.; Wu, S.-C.; Wang, C.-H.; Wu, Y.-M.; Chao, C.-B.; Lin, C.-S. Early endocytosis pathways in SSN-1 cells infected by dragon grouper nervous necrosis virus. J. Gen. Virol. 2005, 86, 2553–2561. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, T.; Nakai, T.; Mori, K.; Arimoto, M.; Furusawa, I.; Nakai, T. High permissivity of the fish cell line SSN-1 for piscine nodaviruses. Dis. Aquat. Organ. 2000, 43, 81–89. [Google Scholar] [CrossRef]
- Chang, J.-S.; Chi, S.-C. GHSC70 is involved in the cellular entry of nervous necrosis virus. J. Virol. 2015, 89, 61–70. [Google Scholar] [CrossRef]
- Krishnan, R.; Kim, J.-O.; Kim, J.-O.; Qadiri, S.S.N.; Kim, S.-J.; Oh, M.-J. Immunoglobulin-like cell adhesion molecules, nectins—Characterization, functional prediction and expression profiling from seven-band grouper, Hyporthodus septemfasciatus. Aquaculture 2019, 506, 387–393. [Google Scholar] [CrossRef]
- Krishnan, R.; Qadiri, S.S.N.; Oh, M.-J. Functional characterization of seven-band grouper immunoglobulin like cell adhesion molecule, Nectin4 as a cellular receptor for nervous necrosis virus. Fish Shellfish Immunol. 2019, 93, 720–725. [Google Scholar] [CrossRef]
- Iwamoto, T.; Okinaka, Y.; Mise, K.; Mori, K.-I.; Arimoto, M.; Okuno, T.; Nakai, T. Identification of host-specificity determinants in betanodaviruses by using reassortants between striped jack nervous necrosis virus and sevenband grouper nervous necrosis virus. J. Virol. 2004, 78, 1256–1262. [Google Scholar] [CrossRef]
- Ito, Y.; Okinaka, Y.; Mori, K.I.; Sugaya, T.; Nishioka, T.; Oka, M.; Nakai, T. Variable region of betanodavirus RNA2 is sufficient to determine host specificity. Dis. Aquat. Organ. 2008, 79, 199–205. [Google Scholar] [CrossRef]
- Souto, S.; Olveira, J.G.; Vázquez-Salgado, L.; Dopazo, C.P.; Bandín, I. Betanodavirus infection in primary neuron cultures from sole. Vet. Res. 2018, 49, 86. [Google Scholar] [CrossRef]
- Ikenaga, T.; Tatecho, Y.; Nakai, T.; Uematsu, K. Betanodavirus as a novel transneuronal tracer for fish. Neurosci. Lett. 2002, 331, 55–59. [Google Scholar] [CrossRef]
- Olveira, J.G.; Soares, F.; Engrola, S.; Dopazo, C.P.; Bandín, I. Antemortem versus postmortem methods for detection of betanodavirus in Senegalese sole (Solea senegalensis). J. Vet. Diagn. Investig. 2008, 20, 215–219. [Google Scholar] [CrossRef]
- Korsnes, K.; Karlsbakk, E.; Devold, M.; Nerland, A.H.; Nylund, A. Tissue tropism of nervous necrosis virus (NNV) in Atlantic cod, Gadus morhua L., after intraperitoneal challenge with a virus isolate from diseased Atlantic halibut, Hippoglossus hippoglossus (L.). J. Fish Dis. 2009, 32, 655–665. [Google Scholar] [CrossRef]
- Kim, J.-O.; Kim, S.-J.; Kim, J.-O.; Kim, W.-S.; Oh, M.-J. Distribution of nervous necrosis virus (NNV) in infected sevenband grouper, Hyporthodus septemfasciatus by intramuscular injection or immersion challenge. Aquaculture 2018, 489, 1–8. [Google Scholar] [CrossRef]
- Nguyen, H.D.; Mushiake, K.; Nakai, T.; Muroga, K. Tissue distribution of striped jack nervous necrosis virus (SJNNV) in adult striped jack. Dis. Aquat. Organ. 1997, 28, 87–91. [Google Scholar] [CrossRef]
- Azad, I.S.; Jithendran, K.P.; Shekhar, M.S.; Thirunavukkarasu, A.R.; De La Peña, L.D. Immunolocalisation of nervous necrosis virus indicates vertical transmission in hatchery produced Asian sea bass (Lates calcarifer Bloch)—A case study. Aquaculture 2006, 255, 39–47. [Google Scholar] [CrossRef][Green Version]
- Grove, S.; Johansen, R.; Reitan, L.J.; Press, C.M.; Dannevig, B.H. Quantitative investigation of antigen and immune response in nervous and lymphoid tissues of Atlantic halibut (Hippoglossus hippoglossus) challenged with nodavirus. Fish Shellfish Immunol. 2006, 21, 525–539. [Google Scholar] [CrossRef]
- Lopez-Jimena, B.; Alonso, M.C.; Thompson, K.D.; Adams, A.; Infante, C.; Castro, D.; Borrego, J.J.; Garcia-Rosado, E. Tissue distribution of Red Spotted Grouper Nervous Necrosis Virus (RGNNV) genome in experimentally infected juvenile European seabass (Dicentrarchus labrax). Vet. Microbiol. 2011, 154, 86–95. [Google Scholar] [CrossRef]
- Mazelet, L.; Dietrich, J.; Rolland, J.L. New RT-qPCR assay for viral nervous necrosis virus detection in sea bass, Dicentrarchus labrax (L.): Application and limits for hatcheries sanitary control. Fish Shellfish Immunol. 2011, 30, 27–32. [Google Scholar] [CrossRef]
- Valero, Y.; Arizcun, M.; Esteban, M.Á.; Bandín, I.; Olveira, J.G.; Patel, S.; Cuesta, A.; Chaves-Pozo, E. Nodavirus colonizes and replicates in the testis of gilthead seabream and european sea bass modulating its immune and reproductive functions. PLoS ONE 2015, 10, 1–24. [Google Scholar] [CrossRef]
- Su, Y.; Xu, H.; Ma, H.; Feng, J.; Wen, W.; Guo, Z. Dynamic distribution and tissue tropism of nervous necrosis virus in juvenile pompano (Trachinotus ovatus) during early stages of infection. Aquaculture 2015, 440, 25–31. [Google Scholar] [CrossRef]
- Parameswaran, V.; Kumar, S.R.; Ahmed, V.P.I.; Hameed, A.S.S. A fish nodavirus associated with mass mortality in hatchery-reared Asian Sea bass, Lates calcarifer. Aquaculture 2008, 275, 366–369. [Google Scholar] [CrossRef]
- Souto, S.; Lopez-Jimena, B.; Alonso, M.C.; García-Rosado, E.; Bandín, I. Experimental susceptibility of European sea bass and Senegalese sole to different betanodavirus isolates. Vet. Microbiol. 2015, 177, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Hick, P.; Schipp, G.; Bosmans, J.; Humphrey, J.; Whittington, R. Recurrent outbreaks of viral nervous necrosis in intensively cultured barramundi (Lates calcarifer) due to horizontal transmission of betanodavirus and recommendations for disease control. Aquaculture 2011, 319, 41–52. [Google Scholar] [CrossRef]
- Pascoli, F.; Serra, M.; Toson, M.; Pretto, T.; Toffan, A. Betanodavirus ability to infect juvenile European sea bass, Dicentrarchus labrax, at different water salinity. J. Fish Dis. 2016, 39, 1061–1068. [Google Scholar] [CrossRef]
- Aranguren, R.; Tafalla, C.; Novoa, B.; Figueras, A. Experimental transmission of encephalopathy and retinopathy induced by nodavirus to sea bream, Sparus aurata L., using different infection models. J. Fish Dis. 2002, 25, 317–324. [Google Scholar] [CrossRef]
- Manin, B.O.; Ransangan, J. Experimental evidence of horizontal transmission of Betanodavirus in hatchery-produced Asian seabass, Lates calcarifer and brown-marbled grouper, Epinephelus fuscoguttatus fingerling. Aquaculture 2011, 321, 157–165. [Google Scholar] [CrossRef]
- Korsnes, K.; Karlsbakk, E.; Nylund, A.; Nerland, A.H. Horizontal transmission of nervous necrosis virus between turbot Scophthalmus maximus and Atlantic cod Gadus morhua using cohabitation challenge. Dis. Aquat. Organ. 2012, 99, 13–21. [Google Scholar] [CrossRef]
- Arimoto, M.; Mori, K.; Nakai, T.; Muroga, K.; Furusawa, I. Pathogenicity of the causative agent of viral nervous necrosis disease in striped jack, Pseudocaranx dentex (Bloch & Schneider). J. Fish Dis. 1993, 16, 461–469. [Google Scholar]
- Frerichs, G.N.; Tweedie, A.; Starkey, W.G.; Richards, R.H. Temperature, pH and electrolyte sensitivity, and heat, UV and disinfectant inactivation of sea bass (Dicentrarchus labrax) neuropathy nodavirus. Aquaculture 2000, 185, 13–24. [Google Scholar] [CrossRef]
- Skliris, G.P.; Richards, R.H. Assessment of the susceptibility of the brine shrimp Artemia salina and rotifer Brachionus plicatilis to experimental nodavirus infections. Aquaculture 1998, 169, 133–141. [Google Scholar] [CrossRef]
- Vazquez-Salgado, L.; Olveira, J.G.; Dopazo, C.P.; Bandín, I. Live food can play a role in nervous necrosis virus (NNV) transmission in marine fish hatcheries (manuscript in preparation).
- Gomez, D.K.; Baeck, G.W.; Kim, J.H.; Choresca, C.H.; Park, S.C. Molecular detection of betanodaviruses from apparently healthy wild marine invertebrates. J. Invertebr. Pathol. 2008, 97, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Volpe, E.; Grodzki, M.; Panzarin, V.; Guercio, A.; Purpari, G.; Serratore, P.; Ciulli, S. Detection and molecular characterization of betanodaviruses retrieved from bivalve molluscs. J. Fish Dis. 2018, 41, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Fichi, G.; Cardeti, G.; Perrucci, S.; Vanni, A.; Cersini, A.; Lenzi, C.; De Wolf, T.; Fronte, B.; Guarducci, M.; Susini, F. Skin lesion-associated pathogens from Octopus vulgaris: First detection of Photobacterium swingsii, Lactococcus garvieae and betanodavirus. Dis. Aquat. Organ. 2015, 115, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Arimoto, M.; Mushiake, K.; Mizuta, Y.; Nakai, T.; Muroge, K.; Furusawa, I. Detection of Striped Jack Nervous Necrosis Virus (SJNNV) by Enzyme-Linked Immunosorbent Assay (ELISA). Fish Pathol. 1992, 27, 191–195. [Google Scholar] [CrossRef]
- Yoshimizu, M.; Suzuki, K.; Nishizawa, T.; Winton, J.R.; Ezura, Y. Antibody Screening for the Identification of Nervous Necrosis Carriers in Flounder Broodstock. In Proceedings of the NRIA International Workshop on New Approaches to Viral Diseases of Aquatic Animals, Kyoto, Japan, 21–24 January 1997; pp. 124–130. [Google Scholar]
- Breuil, G.; Pépin, J.F.P.; Boscher, S.; Thiéry, R. Experimental vertical transmission of nodavirus from broodfish to eggs and larvae of the sea bass, Dicentrarchus labrax (L.). J. Fish Dis. 2002, 25, 697–702. [Google Scholar] [CrossRef]
- Mori, K.; Mushiake, K.; Arimoto, M. Control measures for viral nervous necrosis in striped jack. Fish Pathol. 1998, 33, 443–444. [Google Scholar] [CrossRef]
- Watanabe, K.; Nishizawa, T.; Yoshimizu, M. Selection of brood stock candidates of barfin flounder using an ELISA system with recombinant protein of barfin flounder nervous necrosis virus. Dis. Aquat. Organ. 2000, 41, 219–223. [Google Scholar] [CrossRef]
- Jaramillo, D.; Hick, P.; Whittington, R.J. Age dependency of nervous necrosis virus infection in barramundi Lates calcarifer (Bloch). J. Fish Dis. 2017, 40, 1089–1101. [Google Scholar] [CrossRef]
- Juniar, E.; Kurniasih, K.; Sumiarto, B. Risk factors of a viral nervous necrosis disease in grouper (Epinephelus spp.) cultured in Bintan district, Indonesia. Vet. World 2018, 11, 1558–1563. [Google Scholar] [CrossRef]
- Verrier, E.R.; Langevin, C.; Benmansour, A.; Boudinot, P. Early antiviral response and virus-induced genes in fish. Dev. Comp. Immunol. 2011, 35, 1204–1214. [Google Scholar] [CrossRef]
- Zou, J.; Secombes, C.J. Teleost fish interferons and their role in immunity. Dev. Comp. Immunol. 2011, 35, 1376–1387. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-M.M.; Wang, T.-Y.Y.; Chen, T.-Y.Y. Immunity to betanodavirus infections of marine fish. Dev. Comp. Immunol. 2014, 43, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Scapigliati, G.; Buonocore, F.; Randelli, E.; Casani, D.; Meloni, S.; Zarletti, G.; Tiberi, M.; Pietretti, D.; Boschi, I.; Manchado, M.; et al. Cellular and molecular immune responses of the sea bass (Dicentrarchus labrax) experimentally infected with betanodavirus. Fish Shellfish Immunol. 2010, 28, 303–311. [Google Scholar] [CrossRef]
- Øvergård, A.C.; Nerland, A.H.; Fiksdal, I.U.; Patel, S. Atlantic halibut experimentally infected with nodavirus shows increased levels of T-cell marker and IFNγ transcripts. Dev. Comp. Immunol. 2012, 37, 139–150. [Google Scholar] [CrossRef]
- Chen, C.W.; Wu, M.S.; Huang, Y.J.; Cheng, C.A.; Chang, C.Y. Recognition of linear B-cell epitope of Betanodavirus coat protein by RG-M18 neutralizing mAB inhibits giant grouper nervous necrosis virus (GGNNV) infection. PLoS ONE 2015, 10, 10. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.C.C.; Chi, S.C.C. Persistence of betanodavirus in Barramundi brain (BB) cell line involves the induction of Interferon response. Fish Shellfish Immunol. 2006, 21, 540–547. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Ouyang, Z.; Wang, W.; Yu, Y.; Li, P.; Zhou, S.; Wei, S.; Wei, J.; Huang, X.; Qin, Q. Antiviral role of grouper STING against iridovirus infection. Fish Shellfish Immunol. 2015, 47, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Zhou, Q.; Shi, Y.; Zhang, J.; He, J.; Xie, J. Protein A from orange-spotted grouper nervous necrosis virus triggers type I interferon production in fish cell. Fish Shellfish Immunol. 2018, 79, 234–243. [Google Scholar] [CrossRef]
- Poisa-Beiro, L.; Dios, S.; Montes, A.; Aranguren, R.; Figueras, A.; Novoa, B. Nodavirus increases the expression of Mx and inflammatory cytokines in fish brain. Mol. Immunol. 2008, 45, 218–225. [Google Scholar] [CrossRef]
- Moreno, P.; Lopez-Jimena, B.; Randelli, E.; Scapigliati, G.; Buonocore, F.; Garcia-Rosado, E.; Borrego, J.J.; Alonso, M.C. Immuno-related gene transcription and antibody response in nodavirus (RGNNV and SJNNV)-infected European sea bass (Dicentrarchus labrax L.). Fish Shellfish Immunol. 2018, 78, 270–278. [Google Scholar] [CrossRef]
- Moreno, P.; Souto, S.; Leiva-rebollo, R.; Borrego, J.J.; Bandín, I.; Alonso, M.C. Capsid amino acids at positions 247 and 270 are involved in the virulence of betanodaviruses to European sea bass. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Álvarez-Torres, D.; Podadera, A.M.; Alonso, M.C.; Bandín, I.; Béjar, J.; García-Rosado, E. Molecular characterization and expression analyses of the Solea senegalensis interferon-stimulated gene 15 (isg15) following NNV infections. Fish Shellfish Immunol. 2017, 66, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhou, Y.; Cao, Z.; Sun, Y.; Chen, Y.; Xiang, Y.; Wang, L.; Zhang, S.; Guo, W. Comparative analysis of the expression patterns of IL-1β, IL-11, and IL-34 in golden pompano (Trachinotus ovatus) following different pathogens challenge. Fish Shellfish Immunol. 2019, 93, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Chaves-Pozo, E.; Guardiola, F.A.; Meseguer, J.; Esteban, M.A.; Cuesta, A. Nodavirus infection induces a great innate cell-mediated cytotoxic activity in resistant, gilthead seabream, and susceptible, European sea bass, teleost fish. Fish Shellfish Immunol. 2012, 33, 1159–1166. [Google Scholar] [CrossRef] [PubMed]
- Kai, Y.-H.H.; Wu, Y.-C.C.; Chi, S.-C.C. Immune gene expressions in grouper larvae (Epinephelus coioides) induced by bath and oral vaccinations with inactivated betanodavirus. Fish Shellfish Immunol. 2014, 40, 563–569. [Google Scholar] [CrossRef]
- Jung, J.W.; Lee, J.S.; Kim, J.; Im, S.P.; Kim, S.W.; Lazarte, J.M.S.; Kim, Y.R.; Chun, J.H.; Ha, M.W.; Kim, N.N.; et al. Involvement of CD4-1T cells in the cellular immune response of olive flounder (Paralichthys olivaceus) against viral hemorrhagic septicemia virus (VHSV) and nervous necrosis virus (NNV) infection. Dev. Comp. Immunol. 2020, 103, 1–8. [Google Scholar] [CrossRef]
- Ozsolak, F.; Milos, P.M. RNA sequencing: Advances, challenges and opportunities Fatih. Nat. Rev. Genet. 2011, 12, 87–98. [Google Scholar] [CrossRef]
- Lu, M.-W.W.; Ngou, F.-H.H.; Chao, Y.-M.M.; Lai, Y.-S.S.; Chen, N.-Y.Y.; Lee, F.-Y.Y.; Chiou, P.P. Transcriptome characterization and gene expression of Epinephelus spp in endoplasmic reticulum stress-related pathway during betanodavirus infection in vitro. BMC Genom. 2012, 13, 1–12. [Google Scholar] [CrossRef]
- Liu, P.; Wang, L.; Kwang, J.; Yue, G.H.; Wong, S.M. Transcriptome analysis of genes responding to NNV infection in Asian seabass epithelial cells. Fish Shellfish Immunol. 2016, 54, 342–352. [Google Scholar] [CrossRef]
- Chen, W.; Yi, L.; Feng, S.; Liu, X.; Asim, M.; Zhou, Y.; Lan, J.; Jiang, S.; Tu, J.; Lin, L. Transcriptomic profiles of striped snakehead fish cells (SSN-1) infected with red-spotted grouper nervous necrosis virus (RGNNV) with an emphasis on apoptosis pathway. Fish Shellfish Immunol. 2017, 60, 346–354. [Google Scholar] [CrossRef]
- Chaves-Pozo, E.; Valero, Y.; Esteve-Codina, A.; Gómez-Garrido, J.; Dabad, M.; Alioto, T.; Meseguer, J.; Esteban, M.Á.; Cuesta, A. Innate cell-mediated cytotoxic activity of European sea bass leucocytes against nodavirus-infected cells: A Functional and RNA-seq Study. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Chaves-Pozo, E.; Bandín, I.; Olveira, J.G.; Esteve-Codina, A.; Gómez-Garrido, J.; Dabad, M.; Alioto, T.; Ángeles Esteban, M.; Cuesta, A. European sea bass brain DLB-1cell line is susceptible to nodavirus: A transcriptomic study. Fish Shellfish Immunol. 2019, 86, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Jia, P.; Liu, W.; Yi, M.; Jia, K. Comparative transcriptome analysis reveals the role of p53 signalling pathway during red-spotted grouper nervous necrosis virus infection in Lateolabrax japonicus brain cells. J. Fish Dis. 2019, 42, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Krasnov, A.; Kileng, Ø.; Skugor, S.; Jørgensen, S.M.; Afanasyev, S.; Timmerhaus, G.; Sommer, A.I.; Jensen, I. Genomic analysis of the host response to nervous necrosis virus in Atlantic cod (Gadus morhua) brain. Mol. Immunol. 2013, 54, 443–452. [Google Scholar] [CrossRef]
- Wang, Y.D.; Rajanbabu, V.; Chen, J.Y. Transcriptome analysis of medaka following epinecidin-1 and TH1-5 treatment of NNV infection. Fish Shellfish Immunol. 2015, 42, 121–131. [Google Scholar] [CrossRef]
- Tso, C.-H.H.; Lu, M.-W.W. Transcriptome profiling analysis of grouper during nervous necrosis virus persistent infection. Fish Shellfish Immunol. 2018, 76, 224–232. [Google Scholar] [CrossRef]
- Labella, A.M.; Garcia-Rosado, E.; Bandín, I.; Dopazo, C.P.; Castro, D.; Alonso, M.C.; Borrego, J.J. Transcriptomic profiles of Senegalese sole infected with nervous necrosis virus reassortants presenting different degree of virulence. Front. Immunol. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Wang, L.; Tian, Y.; Cheng, M.; Li, Z.; Li, S.; Wu, Y.; Zhang, J.; Ma, W.; Li, W.; Pang, Z.; et al. Transcriptome comparative analysis of immune tissues from asymptomatic and diseased Epinephelus moara naturally infected with nervous necrosis virus. Fish Shellfish Immunol. 2019, 93, 99–107. [Google Scholar] [CrossRef]
- Kim, J.O.; Kim, J.O.; Kim, W.S.; Oh, M.J. Characterization of the transcriptome and gene expression of brain tissue in sevenband grouper (Hyporthodus septemfasciatus) in response to NNV infection. Genes 2017, 8, 31. [Google Scholar] [CrossRef]
- Barber, D.L.; Wherry, E.J.; Masopust, D.; Zhu, B.; Allison, J.P.; Sharpe, A.H.; Freeman, G.J.; Ahmed, R. Restoring function in exhausted CD8 T cells during chronic viral infection. Nature 2006, 439, 682–687. [Google Scholar] [CrossRef]
- Okagawa, T.; Konnai, S.; Deringer, J.R.; Ueti, M.W.; Scoles, G.A.; Murata, S.; Ohashi, K.; Brown, W.C. Cooperation of PD-1 and LAG-3 contributes to T-cell exhaustion in Anaplasma marginale-infected cattle. Infect. Immun. 2016, 84, 2779–2790. [Google Scholar] [CrossRef] [PubMed]
- Chiang, Y.-H.; Wu, Y.-C.; Chi, S.-C. Interleukin-1β secreted from betanodavirus-infected microglia caused the death of neurons in giant grouper brains. Dev. Comp. Immunol. 2017, 70, 19–26. [Google Scholar] [CrossRef]
- Snieszko, S.F. Recent advances in scientific knowledge and developments pertaining to diseases of fishes. Adv. Vet. Sci. Comp. Med. 1973, 17, 291–314. [Google Scholar]
- Souto, S.; Mérour, E.; Biacchesi, S.; Brémont, M.; Olveira, J.G.; Bandín, I. In Vitro and in vivo characterization of molecular determinants of virulence in reassortant betanodavirus. J. Gen. Virol. 2015, 96, 1287–1296. [Google Scholar] [CrossRef] [PubMed]
- Doan, Q.K.; Vandeputte, M.; Chatain, B.; Morin, T.; Allal, F. Viral encephalopathy and retinopathy in aquaculture: A review. J. Fish Dis. 2017, 40, 717–742. [Google Scholar] [CrossRef] [PubMed]
- Gomez, D.K.; Baeck, G.W.; Kim, J.H.; Choresca, C.H.; Park, S.C. Genetic analysis of betanodaviruses in subclinically infected aquarium fish and invertebrates. Curr. Microbiol. 2008, 56, 499–504. [Google Scholar] [CrossRef]
- Ciulli, S.; Grodzki, M.; Bignami, G.; Serratore, P.; Prosperi, S. Molecular detection and genetic analysis of betanodaviruses in bivalve mollusks. J. Biotechnol. 2010, 150, 4. [Google Scholar] [CrossRef]
- Fichi, G.; Cardeti, G.; Cersini, A.; Mancusi, C.; Guarducci, M.; Di Guardo, G.; Terracciano, G. Bacterial and viral pathogens detected in sea turtles stranded along the coast of Tuscany, Italy. Vet. Microbiol. 2016, 185, 56–61. [Google Scholar] [CrossRef]
- Munday, B.L.; Langdon, J.S.; Hyatt, A.; Humphrey, J.D. Mass mortality associated with a viral-induced vacuolating encephalopathy and retinopathy of larval and juvenile barramundi, Lates calcarifer Bloch. Aquaculture 1992, 103, 197–211. [Google Scholar] [CrossRef]
- Chi, S.C.; Hu, W.W.; Lo, B.J. Establishment and characterization of a continuous cell line (GF-1) derived from grouper, Epinephelus coioides (Hamilton): A cell line susceptible to grouper nervous necrosis virus (GNNV). J. Fish Dis. 1999, 22, 173–182. [Google Scholar] [CrossRef]
- Wen, C.M.; Lee, C.W.; Wang, C.S.; Cheng, Y.H.; Huang, H.Y. Development of two cell lines from Epinephelus coioides brain tissue for characterization of betanodavirus and megalocytivirus infectivity and propagation. Aquaculture 2008, 278, 14–21. [Google Scholar] [CrossRef]
- Sarath Babu, V.; Abdul Majeed, S.; Nambi, K.S.N.; Taju, G.; Madan, N.; Sundar Raj, N.; Sahul Hameed, A.S. Comparison of betanodavirus replication efficiency in ten Indian fish cell lines. Arch. Virol. 2013, 158, 1367–1375. [Google Scholar] [CrossRef]
- Ma, H.; Cheng, C.; Su, Y.; Deng, Y.; Feng, J.; Guo, Z. Propagations of grouper (Epinephelus sp.) viruses in a new fibroblast-like cell line from orange spotted grouper (E. coioides) brain. Aquaculture 2019, 511, 1–7. [Google Scholar] [CrossRef]
- Liu, X.F.; Wu, Y.H.; Wei, S.N.; Wang, N.; Li, Y.Z.; Zhang, N.W.; Li, P.F.; Qin, Q.W.; Chen, S.L. Establishment and characterization of a brain-cell line from kelp grouper Epinephelus moara. J. Fish Biol. 2018, 92, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Le, Y.; Li, Y.; Jin, Y.; Jia, P.; Jia, K.; Yi, M. Establishment and characterization of a brain cell line from sea perch, Lateolabrax japonicus. Vitr. Cell. Dev. Biol. Anim. 2017, 53, 834–840. [Google Scholar] [CrossRef] [PubMed]
- Tu, J.; Chen, W.; Fu, X.; Lin, Q.; Chang, O.; Zhao, L.; Lan, J.; Li, N.; Lin, L. Susceptibility of Chinese perch brain (CPB) Cell and Mandarin fish to red-spotted grouper nervous necrosis virus (RGNNV) Infection. Int. J. Mol. Sci. 2016, 17, 740. [Google Scholar] [CrossRef] [PubMed]
- Adachi, K.; Sumiyoshi, K.; Ariyasu, R.; Yamashita, K.; Zenke, K.; Okinaka, Y. Susceptibilities of medaka (Oryzias latipes) cell lines to a betanodavirus. Virol. J. 2010, 7, 150. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Li, P.; Liu, J.; Ni, S.; Yu, Y.; Yang, M.; Wei, S.; Qin, Q. Establishment and characterization of a mid-kidney cell line derived from golden pompano Trachinotus ovatus, a new cell model for virus pathogenesis and toxicology studies. Vitr. Cell. Dev. Biol. Anim. 2017, 53, 320–327. [Google Scholar] [CrossRef]
- Wang, R.R.; Zhang, N.; Wang, R.R.; Wang, S.; Wang, N. Two skin cell lines from wild-type and albino Japanese flounder (Paralichthys olivaceus): Establishment, characterization, virus susceptibility, efficient transfection, and application to albinism study. Fish Physiol. Biochem. 2017, 43, 1477–1486. [Google Scholar] [CrossRef]
- Li, P.; Zhou, L.; Wei, S.; Yang, M.; Ni, S.; Yu, Y.; Cai, J.; Qin, Q. Establishment and characterization of a cell line from the head kidney of golden pompano Trachinotus ovatus and its application in toxicology and virus susceptibility. J. Fish Biol. 2017, 90, 1944–1959. [Google Scholar] [CrossRef]
- Sahul Hameed, A.S.; Parameswaran, V.; Shukla, R.; Bright Singh, I.S.; Thirunavukkarasu, A.R.; Bhonde, R.R. Establishment and characterization of India’s first marine fish cell line (SISK) from the kidney of sea bass (Lates calcarifer). Aquaculture 2006, 257, 92–103. [Google Scholar] [CrossRef]
- Parameswaran, V.; Shukla, R.; Bhonde, R.R.; Sahul Hameed, A.S. Splenic cell line from sea bass, Lates calcarifer: Establishment and characterization. Aquaculture 2006, 261, 43–53. [Google Scholar] [CrossRef]
- Chi, S.; Wu, Y.; Cheng, T. Persistent infection of betanodavirus in a novel cell line derived from the brain tissue of barramundi Lates calcarifer. Dis. Aquat. Organ. 2005, 65, 91–98. [Google Scholar] [CrossRef]
- Wen, C.M. Characterization and viral susceptibility of a brain cell line from brown-marbled grouper Epinephelus fuscoguttatus (Forsskål) with persistent betanodavirus infection. J. Fish Dis. 2016, 39, 1335–1346. [Google Scholar] [CrossRef] [PubMed]
- Adachi, K.; Ichinose, T.; Watanabe, K.; Kitazato, K.; Kobayashi, N. Potential for the replication of the betanodavirus redspotted grouper nervous necrosis virus in human cell lines. Arch. Virol. 2008, 153, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Takizawa, N.; Adachi, K.; Ichinose, T.; Kobayashi, N. Efficient propagation of betanodavirus in a murine astrocytoma cell line. Virus Res. 2008, 136, 206–210. [Google Scholar] [CrossRef]
- Chapter 2.2.2. Viral encephalopathy and retinopathy. In OIE Diagnostic Manual for Aquatic Animal Diseases; OIE: Paris, France, 2000; pp. 69–73.
- Kuo, H.C.; Wang, T.Y.; Chen, P.P.; Chen, Y.M.; Chuang, H.C.; Chen, T.Y. Real-time quantitative PCR assay for monitoring of nervous necrosis virus infection in grouper aquaculture. J. Clin. Microbiol. 2011, 49, 1090–1096. [Google Scholar] [CrossRef]
- Kim, J.-O.; Kim, J.-O.; Kim, W.-S.; Oh, M.-J. Development and application of quantitative detection method for nervous necrosis virus (NNV) isolated from sevenband grouper Hyporthodus septemfasciatus. Asian Pac. J. Trop. Med. 2016, 9, 742–748. [Google Scholar] [CrossRef]
- Nishizawa, T.; Muroga, K.; Arimoto, M. Failure of the Polymerase Chain Reaction (PCR) Method to Detect Striped Jack Nervous Necrosis Virus (SJNNV) in Striped Jack Pseudocaranx dentex Selected as Spawners. J. Aquat. Anim. Health 1996, 8, 332–334. [Google Scholar] [CrossRef]
- Thiery, R.; Raymond, J.-C.; Castric, J. Natural outbreak of viral encephalopathy and retinopathy in juvenile sea bass, Dicentrarchus labrax: Study by nested reverse transcriptase–polymerase chain reaction. Virus Res. 1999, 63, 11–17. [Google Scholar] [CrossRef]
- Nishizawa, T.; Mori, K.; Nakai, T.; Furusawa, I.; Muroga, K. Polymerase chain reaction (PCR) amplification of RNA of striped jack nervous necrosis virus (SJNNV). Dis. Aquat. Organ. 1994, 18, 103–107. [Google Scholar] [CrossRef]
- Dalla Valle, L.; Toffolo, V.; Lamprecht, M.; Maltese, C.; Bovo, G.; Belvedere, P.; Colombo, L. Development of a sensitive and quantitative diagnostic assay for fish nervous necrosis virus based on two-target real-time PCR. Vet. Microbiol. 2005, 110, 167–179. [Google Scholar] [CrossRef]
- Nerland, A.H.; Skaar, C.; Eriksen, T.B.; Bleie, H. Detection of nodavirus in seawater from rearing facilities for Atlantic halibut Hippoglossus hippoglossus larvae. Dis. Aquat. Organ. 2007, 73, 201–205. [Google Scholar] [CrossRef]
- Hick, P.; Whittington, R.J. Optimisation and validation of a real-time reverse transcriptase-polymerase chain reaction assay for detection of betanodavirus. J. Virol. Methods 2010, 163, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Baud, M.; Cabon, J.; Salomoni, A.; Toffan, A.; Panzarin, V.; Bigarré, L. First generic one step real-time Taqman RT-PCR targeting the RNA1 of betanodaviruses. J. Virol. Methods 2015, 211, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Toubanaki, D.K.; Margaroni, M.; Karagouni, E. Development of a novel allele-specific PCR method for rapid assessment of nervous necrosis virus genotypes. Curr. Microbiol. 2015, 71, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Toubanaki, D.K.; Karagouni, E. Genotype-specific real-time PCR combined with high-resolution melting analysis for rapid identification of red-spotted grouper nervous necrosis virus. Arch. Virol. 2017, 162, 2315–2328. [Google Scholar] [CrossRef] [PubMed]
- Thiéry, R.; Arnauld, C.; Delsert, C. Two isolates of sea bass, Dicentrarchus labrax L., nervous necrosis virus with distinct genomes. J. Fish Dis. 1999, 22, 201–207. [Google Scholar] [CrossRef]
- Bigarré, L.; Baud, M.; Cabon, J.; Crenn, K.; Castric, J. New PCR probes for detection and genotyping of piscine betanodaviruses. J. Fish Dis. 2010, 33, 907–912. [Google Scholar] [CrossRef]
- Toubanaki, D.K.; Margaroni, M.; Karagouni, E. Nanoparticle-based lateral flow biosensor for visual detection of fish nervous necrosis virus amplification products. Mol. Cell. Probes 2015, 29, 158–166. [Google Scholar] [CrossRef]
- Ferreira, I.A.; Costa, J.Z.; Macchia, V.; Dawn Thompson, K.; Baptista, T. Detection of Betanodavirus in experimentally infected European seabass (Dicentrarchus labrax, Linnaeus 1758) using non-lethal sampling methods. J. Fish Dis. 2019, 42, 13015. [Google Scholar] [CrossRef] [PubMed]
- Starkey, G.W.; Millar, R.M.; Jenkins, M.E.; Ireland, J.H.; Muir, K.F.; Richards, R.H. Detection of piscine nodaviruses by real-time nucleic acid sequence based amplification (NASBA). Dis. Aquat. Organ. 2004, 59, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.D.; Feng, J.; Guo, Z.X.; Ou, Y.J.; Wang, J.Y. Detection of red-spotted grouper nervous necrosis virus by loop-mediated isothermal amplification. J. Virol. Methods 2010, 163, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.D.; Shi, C.Y.; Huang, J.; Shen, G.M.; Li, J.; Wang, S.Q.; Fan, C. Establishment and application of cross-priming isothermal amplification coupled with lateral flow dipstick (CPA-LFD) for rapid and specific detection of red-spotted grouper nervous necrosis virus. Virol. J. 2015, 12, 149. [Google Scholar] [CrossRef]
- Mekata, T.; Satoh, J.; Inada, M.; Dinesh, S.; Harsha, P.; Itami, T.; Sudhakaran, R. Development of simple, rapid and sensitive detection assay for grouper nervous necrosis virus using real-time loop-mediated isothermal amplification. J. Fish Dis. 2015, 38, 873–879. [Google Scholar] [CrossRef]
- Huang, B.; Tan, C.; Chang, S.F.; Munday, B.; Mathew, J.A.; Ngoh, G.H.; Kwang, J. Detection of nodavirus in barramundi, Lates calcarifer (Bloch), using recombinant coat protein-based ELISA and RT-PCR. J. Fish Dis. 2001, 24, 135–141. [Google Scholar] [CrossRef]
- Shieh, J.; Chi, S. Production of monoclonal antibodies against grouper nervous necrosis virus (GNNV) and development of an antigen capture ELISA. Dis. Aquat. Organ. 2005, 63, 53–60. [Google Scholar] [CrossRef]
- Nuñez-Ortiz, N.; Pascoli, F.; Picchietti, S.; Buonocore, F.; Bernini, C.; Toson, M.; Scapigliati, G.; Toffan, A. A formalin-inactivated immunogen against viral encephalopathy and retinopathy (VER) disease in European sea bass (Dicentrarchus labrax): Immunological and protection effects. Vet. Res. 2016, 47, 89. [Google Scholar] [CrossRef]
- Gye, H.J.; Oh, M.-J.; Nishizawa, T. Lack of nervous necrosis virus (NNV) neutralizing antibodies in convalescent sevenband grouper Hyporthodus septemfasciatus after NNV infection. Vaccine 2018, 36, 1863–1870. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Jimena, B.; Garcia-Rosado, E.; Thompson, K.D.; Adams, A.; Infante, C.; Borrego, J.J.; Alonso, M.C. Distribution of red-spotted grouper nervous necrosis virus (RGNNV) antigens in nervous and non-nervous organs of European seabass (Dicentrarchus labrax) during the course of an experimental challenge. J. Vet. Sci. 2012, 13, 355–362. [Google Scholar] [CrossRef]
- Tarrab, K.; Ravid-Peretz, S.; Ucko, M. Immunoserology of European seabass (Dicentrarchus labrax) and white grouper (Epinephelus aeneus) as a non-lethal diagnostic tool for viral nervous necrosis. Aquac. Int. 2019, 27, 63–77. [Google Scholar] [CrossRef]
- Choi, B.; Gye, H.J.H.; Oh, M.M.-J.; Nishizawa, T. Cell culture medium inhibits antigen binding used in an ELISA for detection of antibodies against nervous necrosis virus. J. Aquat. Anim. Health 2014, 26, 168–172. [Google Scholar] [CrossRef] [PubMed]
- Jaramillo, D.; Hick, P.; Deece, K.; Tweedie, A.; Kirkland, P.; Arzey, E.; Whittington, R.J. Comparison of ELISA formats for detection of antibodies specific for nervous necrosis virus (Betanodavirus) in the serum of immunized barramundi Lates calcarifer and Australian bass Macquaria novemaculeata. Aquaculture 2016, 451, 33–38. [Google Scholar] [CrossRef]
- Gye, H.J.; Nishizawa, T. Reducing background optical density in enzyme-linked immunosorbent assay for detecting nervous necrosis virus (NNV)-specific IgM by immobilizing fish sera. Aquaculture 2018, 485, 93–100. [Google Scholar] [CrossRef]
- Zhou, L.; Li, P.; Ni, S.; Yu, Y.; Yang, M.; Wei, S.; Qin, Q. Rapid and sensitive detection of redspotted grouper nervous necrosis virus (RGNNV) infection by aptamer-coat protein-aptamer sandwich enzyme-linked apta-sorbent assay (ELASA). J. Fish Dis. 2017, 40, 1831–1838. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.O.; Kim, J.O.; Kim, S.J.; Kim, W.S.; Oh, M.J. Development of double labeling in situ hybridization using RNA probes for genome detection of nervous necrosis virus (NNV). Mol. Cell. Probes 2018, 42, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Eivazzadeh-Keihan, R.; Pashazadeh-Panahi, P.; Mahmoudi, T.; Chenab, K.K.; Baradaran, B.; Hashemzaei, M.; Radinekiyan, F.; Mokhtarzadeh, A.; Maleki, A. Dengue virus: A review on advances in detection and trends—From conventional methods to novel biosensors. Microchim. Acta 2019, 186, 329. [Google Scholar] [CrossRef]
- Lee, J.H.; Oh, B.K.; Choi, J.W. Development of a HIV-1 virus detection system based on nanotechnology. Sensors 2015, 15, 9915–9927. [Google Scholar] [CrossRef]
- Peng, X.; Luo, G.; Wu, Z.; Wen, W.; Zhang, X.; Wang, S. Fluorescent-magnetic-catalytic nanospheres for dual-modality detection of H9N2 avian influenza virus. ACS Appl. Mater. Interfaces 2019, 11, 41148–41156. [Google Scholar] [CrossRef]
- Ochmann, S.E.; Vietz, C.; Trofymchuk, K.; Acuna, G.P.; Lalkens, B.; Tinnefeld, P. Optical nanoantenna for single molecule-based detection of Zika Virus nucleic acids without molecular multiplication. Anal. Chem. 2017, 89, 13000–13007. [Google Scholar] [CrossRef]
- Sabzehparvar, F.; Rahmani Cherati, T.; Mohsenifar, A.; Roodbar Shojaei, T.; Tabatabaei, M. Immobilization of gold nanoparticles with rhodamine to enhance the fluorescence resonance energy transfer between quantum dots and rhodamine; new method for downstream sensing of infectious bursal disease virus. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2019, 212, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Takemura, K.; Lee, J.; Suzuki, T.; Hara, T.; Abe, F.; Park, E.Y. Ultrasensitive detection of norovirus using a magnetofluoroimmunoassay based on synergic properties of gold/magnetic nanoparticle hybrid nanocomposites and quantum dots. Sens. Actuators B Chem. 2019, 296, 126672. [Google Scholar] [CrossRef]
- Wang, C.; Wang, C.; Wang, X.; Wang, K.; Zhu, Y.; Rong, Z.; Wang, W.; Xiao, R.; Wang, S. Magnetic SERS strip for sensitive and simultaneous detection of respiratory viruses. ACS Appl. Mater. Interfaces 2019, 11, 19495–19505. [Google Scholar] [CrossRef] [PubMed]
- Yadavalli, T.; Shukla, D. Role of metal and metal oxide nanoparticles as diagnostic and therapeutic tools for highly prevalent viral infections. Nanomed. Nanotechnol. Biol. Med. 2017, 13, 219–230. [Google Scholar] [CrossRef]
- Zhang, T.; Tian, F.; Long, L.; Liu, J.; Wu, X. Diagnosis of rubella virus using antigen- conjugated Au @ Pt nanorods as nanozyme probe. Int. J. Nanomed. 2018, 13, 4795–4805. [Google Scholar] [CrossRef]
- Cheng, X.; Chen, G.; Rodriguez, W.R. Micro- and nanotechnology for viral detection. Anal. Bioanal. Chem. 2009, 393, 487–501. [Google Scholar] [CrossRef]
- Tram, D.T.N.; Wang, H.; Sugiarto, S.; Li, T.; Ang, W.H.; Lee, C.; Pastorin, G. Advances in nanomaterials and their applications in point of care (POC) devices for the diagnosis of infectious diseases. Biotechnol. Adv. 2016, 34, 1275–1288. [Google Scholar] [CrossRef]
- Chavan, S.G.; Kim, D.; Hwang, J.; Choi, Y.; Hong, J.W.; Kim, J.; Lee, M.H.; Hwang, M.P.; Choi, J. Enhanced detection of infectious pancreatic necrosis virus via lateral flow chip and fluorometric biosensors based on self-assembled protein nanoprobes. ACS Sens. 2019, 4, 4–11. [Google Scholar] [CrossRef]
- Arimoto, M.; Sato, J.; Maruyama, K.; Mimura, G.; Furusawa, I. Effect of chemical and physical treatments on the inactivation of striped jack nervous necrosis virus (SJNNV). Aquaculture 1996, 143, 15–22. [Google Scholar] [CrossRef]
- Adachi, K.; Ichinose, T.; Takizawa, N.; Watanabe, K.; Kitazato, K.; Kobayashi, N. Inhibition of betanodavirus infection by inhibitors of endosomal acidification. Arch. Virol. 2007, 152, 2217–2224. [Google Scholar] [CrossRef]
- Kai, Y.H.; Su, H.M.; Tai, K.T.; Chi, S.C. Vaccination of grouper broodfish (Epinephelus tukula) reduces the risk of vertical transmission by nervous necrosis virus. Vaccine 2010, 28, 996–1001. [Google Scholar] [CrossRef]
- Wang, Y.D.; Kung, C.W.; Chen, J.Y. Antiviral activity by fish antimicrobial peptides of epinecidin-1 and hepcidin 1-5 against nervous necrosis virus in medaka. Peptides 2010, 31, 1026–1033. [Google Scholar] [CrossRef]
- Huang, Y.C.; Han, Y.S. Determining anti-betanodavirus compounds through a GF-1 cell-based screening platform. Antiviral Res. 2014, 105, 47–53. [Google Scholar] [CrossRef]
- Huang, Y.-C.; Lin, T.-S.; Peng, C.; Chan, N.-L.; Han, Y.-S. Strong inhibition of betanodavirus replication by ribavirin targeting RNA-dependent RNA polymerase. J. Fish Dis. 2016, 39, 619–623. [Google Scholar] [CrossRef]
- Zhou, L.; Li, P.; Yang, M.; Yu, Y.; Huang, Y.; Wei, J.; Wei, S.; Qin, Q. Generation and characterization of novel DNA aptamers against coat protein of grouper nervous necrosis virus (GNNV) with antiviral activities and delivery potential in grouper cells. Antiviral Res. 2016, 129, 104–114. [Google Scholar] [CrossRef]
- Zhou, Q.; Zhang, J.; Huang, R.; Huang, S.; Wu, Y.; Huang, L.; He, J.; Xie, J. An affinity peptide exerts antiviral activity by strongly binding nervous necrosis virus to block viral entry. Fish Shellfish Immunol. 2019, 86, 465–473. [Google Scholar] [CrossRef]
- Morick, D.; Saragovi, A. Inhibition of nervous necrosis virus by ribavirin in a zebrafish larvae model. Fish Shellfish Immunol. 2017, 60, 537–544. [Google Scholar] [CrossRef]
- Sushila, N.; Hameed, A.S.S.; Prasad, K.P.; Majeed, S.A.; Tripathi, G. In vitro screening of selected antiviral drugs against betanodavirus. J. Virol. Methods 2018, 259, 66–73. [Google Scholar] [CrossRef]
- Sieczkarski, S.B.; Whittaker, G.R. Influenza virus can enter and infect cells in the absence of clathrin-mediated endocytosis. J. Virol. 2002, 76, 10455–10464. [Google Scholar] [CrossRef]
- Zhu, S.; Li, J.; Huang, A.-G.; Huang, J.-Q.; Huang, Y.-Q.; Wang, G.-X. Anti-betanodavirus activity of isoprinosine and improved efficacy using carbon nanotubes based drug delivery system. Aquaculture 2019, 512, 734377. [Google Scholar] [CrossRef]
- Zhu, S.; Huang, A.G.; Luo, F.; Li, J.; Li, J.; Zhu, L.; Zhao, L.; Zhu, B.; Ling, F.; Wang, G.X. Application of Virus Targeting Nanocarrier Drug Delivery System in Virus-Induced Central Nervous System Disease Treatment. ACS Appl. Mater. Interfaces 2019, 11, 19006–19016. [Google Scholar] [CrossRef]
- Wang, Y.D.; Kung, C.W.; Chi, S.C.; Chen, J.Y. Inactivation of nervous necrosis virus infecting grouper (Epinephelus coioides) by epinecidin-1 and hepcidin 1-5 antimicrobial peptides, and downregulation of Mx2 and Mx3 gene expressions. Fish Shellfish Immunol. 2010, 28, 113–120. [Google Scholar] [CrossRef]
- Darmostuk, M.; Rimpelova, S.; Gbelcova, H.; Ruml, T. Current approaches in SELEX: An update to aptamer selection technology. Biotechnol. Adv. 2015, 33, 1141–1161. [Google Scholar] [CrossRef]
- Ramalingam, D.; Duclair, S.; Datta, S.A.K.; Ellington, A.; Rein, A.; Prasad, V.R. RNA aptamers directed to human immunodeficiency virus type 1 Gag polyprotein bind to the matrix and nucleocapsid domains and inhibit virus production. J. Virol. 2011, 85, 305. [Google Scholar] [CrossRef]
- Ditzler, M.A.; Bose, D.; Shkriabai, N.; Marchand, B.; Sarafianos, S.G.; Kvaratskhelia, M.; Burke, D.H. Broad-spectrum aptamer inhibitors of HIV reverse transcriptase closely mimic natural substrates. Nucleic Acids Res. 2011, 39, 8237–8247. [Google Scholar] [CrossRef]
- Yamashita, H.; Fujita, Y.; Kawakami, H.; Nakai, T. The efficacy of inactivated virus vaccine against viral nervous necrosis (VNN). Fish Pathol. 2005, 40, 15–21. [Google Scholar] [CrossRef][Green Version]
- Kai, Y.H.; Chi, S.C. Efficacies of inactivated vaccines against betanodavirus in grouper larvae (Epinephelus coioides) by bath immunization. Vaccine 2008, 26, 1450–1457. [Google Scholar] [CrossRef]
- Pakingking, R.; Bautista, N.B.; de Jesus-Ayson, E.G.; Reyes, O. Protective immunity against viral nervous necrosis (VNN) in brown-marbled grouper (Epinephelus fuscogutattus) following vaccination with inactivated betanodavirus. Fish Shellfish Immunol. 2010, 28, 525–533. [Google Scholar] [CrossRef]
- Cheng, Y.-K.K.; Wu, Y.-C.C.; Chi, S.-C.C. Humoral and cytokine responses in giant groupers after vaccination and challenge with betanodavirus. Dev. Comp. Immunol. 2017, 67, 385–394. [Google Scholar] [CrossRef]
- Pakingking, R.; de Jesus-Ayson, E.G.; Reyes, O.; Brian Bautista, N. Immunization regimen in Asian sea bass (Lates calcarifer) broodfish: A practical strategy to control vertical transmission of nervous necrosis virus during seed production. Vaccine 2018, 36, 5002–5009. [Google Scholar] [CrossRef]
- Valero, Y.; Mokrani, D.; Chaves-Pozo, E.; Arizcun, M.; Oumouna, M.; Meseguer, J.; Esteban, M.Á.; Cuesta, A. Vaccination with UV-inactivated nodavirus partly protects European sea bass against infection, while inducing few changes in immunity. Dev. Comp. Immunol. 2018, 86, 171–179. [Google Scholar] [CrossRef]
- Cho, S.Y.; Kim, H.J.; Lan, N.T.; Han, H.-J.; Lee, D.-C.; Hwang, J.Y.; Kwon, M.-G.; Kang, B.K.; Han, S.Y.; Moon, H.; et al. Oral vaccination through voluntary consumption of the convict grouper Epinephelus septemfasciatus with yeast producing the capsid protein of red-spotted grouper nervous necrosis virus. Vet. Microbiol. 2017, 204, 159–164. [Google Scholar] [CrossRef]
- Cho, H.S.; Seo, J.Y.; Park, S.I.; Kim, T.G.; Kim, T.J. Oral immunization with recombinant protein antigen expressed in tobacco against fish nervous necrosis virus. J. Vet. Med. Sci. 2018, 80, 272–279. [Google Scholar] [CrossRef]
- Gonzalez-Silvera, D.; Guardiola, F.A.; Espinosa, C.; Chaves-Pozo, E.; Esteban, M.Á.; Cuesta, A. Recombinant nodavirus vaccine produced in bacteria and administered without purification elicits humoral immunity and protects European sea bass against infection. Fish Shellfish Immunol. 2019, 88, 458–463. [Google Scholar] [CrossRef]
- Lin, C.-F.F.; Jiang, H.-K.K.; Chen, N.-C.C.; Wang, T.-Y.Y.; Chen, T.-Y.Y. Novel subunit vaccine with linear array epitope protect giant grouper against nervous necrosis virus infection. Fish Shellfish Immunol. 2018, 74, 551–558. [Google Scholar] [CrossRef]
- Luu, V.-T.; Moon, H.Y.; Hwang, J.Y.; Kang, B.-K.; Kang, H.A. Development of recombinant Yarrowia lipolytica producing virus-like particles of a fish nervous necrosis virus. J. Microbiol. 2017, 55, 655–664. [Google Scholar] [CrossRef]
- Yuasa, K.; Koesharyani, I.; Roza, D.; Mori, K.; Katata, M.; Nakai, T. Immune response of humpback grouper, Cromileptes altivelis (Valenciennes) injected with the recombinant coat protein of betanodavirus. J. Fish Dis. 2002, 25, 53–56. [Google Scholar] [CrossRef]
- Sommerset, I.; Skern, R.; Biering, E.; Bleie, H.; Fiksdal, I.U.; Grove, S.; Nerland, A.H. Protection against Atlantic halibut nodavirus in turbot is induced by recombinant capsid protein vaccination but not following DNA vaccination. Fish Shellfish Immunol. 2005, 18, 13–29. [Google Scholar] [CrossRef]
- Thiery, R.; Cozien, J.; Cabon, J.; Lamour, F.; Baud, M.; Schneemann, A. Induction of a protective immune response against viral nervous necrosis in the European sea bass Dicentrarchus labrax by using betanodavirus virus-like particles. J. Virol. 2006, 80, 10201–10207. [Google Scholar] [CrossRef]
- Wi, G.R.; Hwang, J.Y.; Kwon, M.-G.G.; Kim, H.J.H.-J.J.; Kang, H.A.; Kim, H.J.H.-J.J. Protective immunity against nervous necrosis virus in convict grouper Epinephelus septemfasciatus following vaccination with virus-like particles produced in yeast Saccharomyces cerevisiae. Vet. Microbiol. 2015, 177, 214–218. [Google Scholar] [CrossRef]
- Lin, K.; Zhu, Z.; Ge, H.; Zheng, L.; Huang, Z.; Wu, S. Immunity to nervous necrosis virus infections of orange-spotted grouper (Epinephelus coioides) by vaccination with virus-like particles. Fish Shellfish Immunol. 2016, 56, 136–143. [Google Scholar] [CrossRef]
- Chien, M.-H.H.; Wu, S.-Y.Y.; Lin, C.-H.H. Oral immunization with cell-free self-assembly virus-like particles against orange-spotted grouper nervous necrosis virus in grouper larvae, Epinephelus coioides. Vet. Immunol. Immunopathol. 2018, 197, 69–75. [Google Scholar] [CrossRef]
- Lai, Y.-X.X.; Jin, B.-L.L.; Xu, Y.; Huang, L.J.; Huang, R.-Q.Q.; Zhang, Y.; Kwang, J.; He, J.-G.G.; Xie, J.-F.F. Immune responses of orange-spotted grouper, Epinephelus coioides, against virus-like particles of betanodavirus produced in Escherichia coli. Vet. Immunol. Immunopathol. 2014, 157, 87–96. [Google Scholar] [CrossRef]
- Lan, N.T.; Kim, H.J.; Han, H.-J.; Lee, D.-C.; Kang, B.K.; Han, S.Y.; Moon, H.; Kim, H.-J. Stability of virus-like particles of red-spotted grouper nervous necrosis virus in the aqueous state, and the vaccine potential of lyophilized particles. Biologicals 2018, 51, 25–31. [Google Scholar] [CrossRef]
- Sommerset, I.; Lorenzen, E.; Lorenzen, N.; Bleie, H.; Nerland, A.H. A DNA vaccine directed against a rainbow trout rhabdovirus induces early protection against a nodavirus challenge in turbot. Vaccine 2003, 21, 4661–4667. [Google Scholar] [CrossRef]
- Vimal, S.; Abdul Majeed, S.; Nambi, K.S.N.; Madan, N.; Farook, M.A.; Venkatesan, C.; Taju, G.; Venu, S.; Subburaj, R.; Thirunavukkarasu, A.R.; et al. Delivery of DNA vaccine using chitosan–tripolyphosphate (CS/TPP) nanoparticles in Asian sea bass, Lates calcarifer (Bloch, 1790) for protection against nodavirus infection. Aquaculture 2014, 420–421, 240–246. [Google Scholar] [CrossRef]
- Valero, Y.; Awad, E.; Buonocore, F.; Arizcun, M.; Esteban, M.Á.; Meseguer, J.; Chaves-Pozo, E.; Cuesta, A. An oral chitosan DNA vaccine against nodavirus improves transcription of cell-mediated cytotoxicity and interferon genes in the European sea bass juveniles gut and survival upon infection. Dev. Comp. Immunol. 2016, 65, 64–72. [Google Scholar] [CrossRef]
- Chen, S.P.; Peng, R.H.; Chiou, P.P. Modulatory effect of CpG oligodeoxynucleotide on a DNA vaccine against nervous necrosis virus in orange-spotted grouper (Epinephelus coioides). Fish Shellfish Immunol. 2015, 45, 919–926. [Google Scholar] [CrossRef]
- Vimal, S.; Farook, M.A.; Madan, N.; Abdul Majeed, S.; Nambi, K.S.N.; Taju, G.; Sundar raj, N.; Venu, S.; Subburaj, R.; Thirunavukkarasu, A.R.; et al. Development, distribution and expression of a DNA vaccine against nodavirus in Asian Seabass, Lates calcarifier (Bloch, 1790). Aquac. Res. 2016, 47, 1209–1220. [Google Scholar] [CrossRef]
- Huang, J.N.; Lin, L.; Weng, S.P.; He, J.G. High expression of capsid protein of red-spotted grouper nervous necrosis virus in an avian cell line requires viral RNA2 non-coding regions. J. Fish Dis. 2007, 30, 439–444. [Google Scholar] [CrossRef]
- Gaafar, A.Y.; Yamashita, H.; Istiqomah, I.; Kawato, Y.; Ninomiya, K.; Younes, A.E.; Nakai, T. An oral vaccination method with the aid of Capsaicin against viral nervous necrosis (VNN). Fish Pathol. 2018, 53, 110–113. [Google Scholar] [CrossRef]
- Lin, C.-C.; Lin, J.H.-Y.; Chen, M.-S.; Yang, H.-L. An oral nervous necrosis virus vaccine that induces protective immunity in larvae of grouper (Epinephelus coioides). Aquaculture 2007, 268, 265–273. [Google Scholar] [CrossRef]
- Oh, M.J.; Gye, H.J.; Nishizawa, T. Assessment of the sevenband grouper Epinephelus septemfasciatus with a live nervous necrosis virus (NNV) vaccine at natural seawater temperature. Vaccine 2013, 31, 2025–2027. [Google Scholar] [CrossRef]
- Boutier, M.; Ronsmans, M.; Ouyang, P.; Fournier, G.; Reschner, A.; Rakus, K.; Wilkie, G.S.; Farnir, F.; Bayrou, C.; Lieffrig, F.; et al. Rational development of an attenuated recombinant Cyprinid herpesvirus 3 vaccine using prokaryotic mutagenesis and in vivo bioluminescent imaging. PLoS Pathog. 2015, 11, e1004690. [Google Scholar] [CrossRef]
- Rouxel, R.N.; Tafalla, C.; Mérour, E.; Leal, E.; Biacchesi, S.; Brémont, M. Attenuated infectious hematopoietic necrosis virus with rearranged gene order as potential vaccine. J. Virol. 2016, 90, 10857–10866. [Google Scholar] [CrossRef]



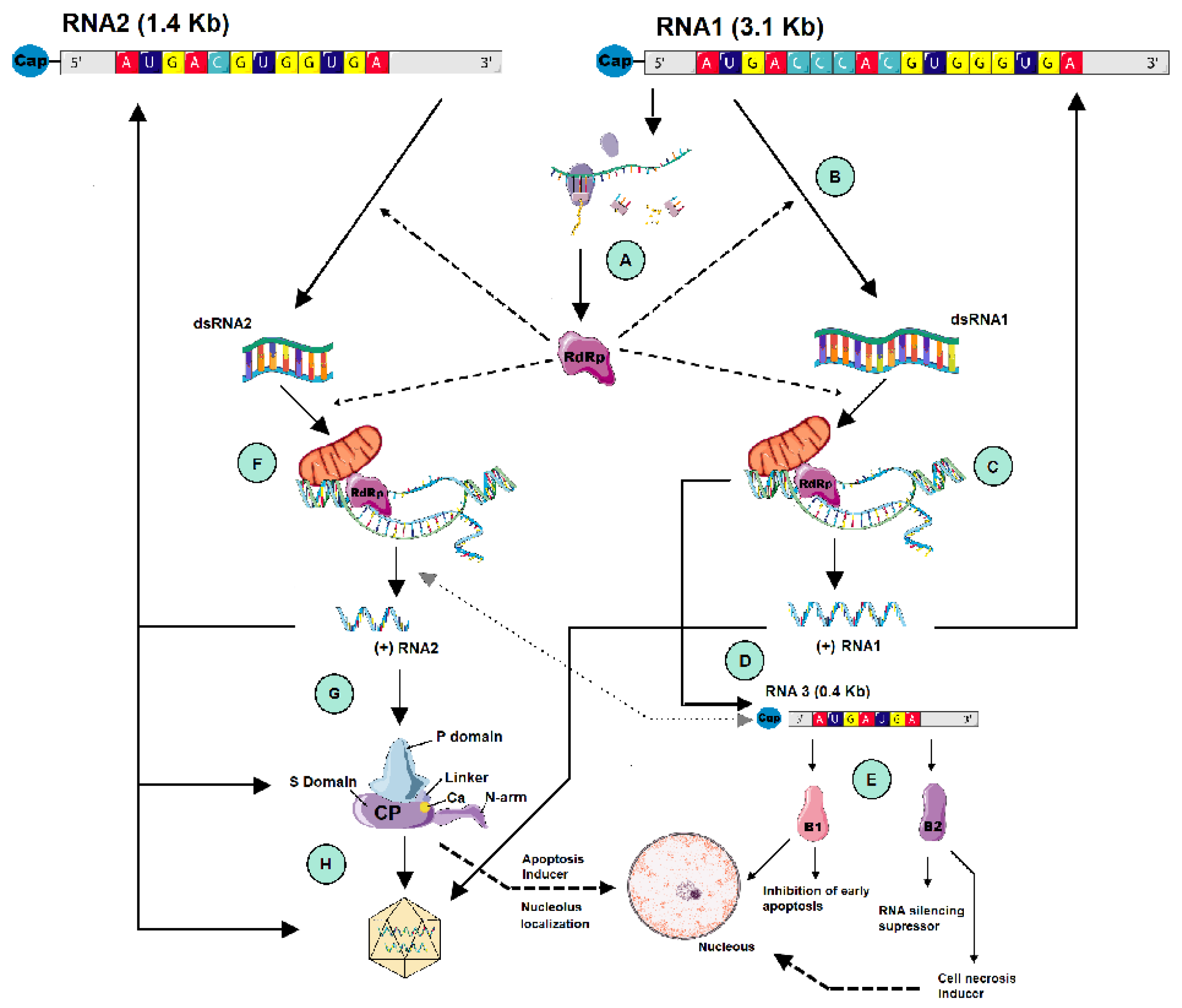{kind=link}
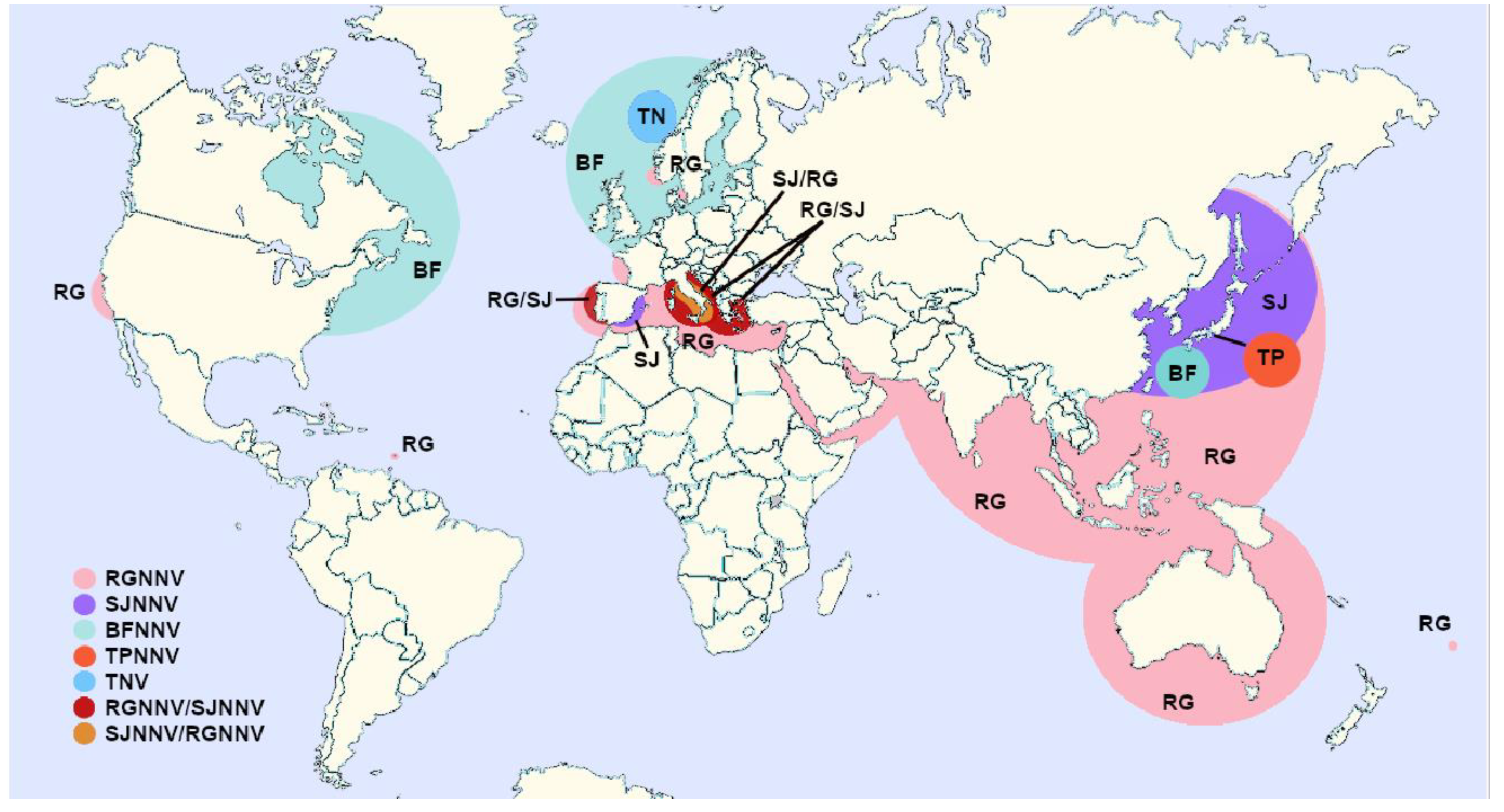{kind=link}
| Order | Family | Common Name | Latin Name | Status a | Genotype | References | Geographic Range |
|---|---|---|---|---|---|---|---|
| Centrarchiformes | Latridae | Striped trumpeteer | Latris lineata | F | ND b | [74] | Australia |
| Oplegnathidae | Japanese parrotfish | Oplegnathus fasciatus | F | ND | [75] | Japan | |
| Spotted knifejaw | O. punctatus | F | RGNNV | [76,77] | Japan | ||
| Australian bass | Macquaria novemaculeata | F | RGNNV | [46] | Australia | ||
| Gadiformes | Gadidade | Atlantic cod | Gadus morhua | F | BFNNV | [34,44,47,77,78] | Canada, USA, Norway, UK |
| Pacific cod | Gadus macrocephalus | F | BFNNV | [59,79] | Japan, China | ||
| Haddock | Melanogrammus aeglefinus | F | BFNNV | [44] | Canada, Atlantic USA | ||
| Gonorynchiformes | Chanidae | Milk fish | Chanos chanos | W | ND | [80] | India |
| Mugiliformes | Mugilidae | Flathead grey mullet | Mugil cephalus | F | RGNNV | [81] | Israel |
| Golden mullet | Liza auratus | W | ND | [82] | Iran | ||
| Leaping mullet | L. saliens | W | ND | [82] | Iran | ||
| Perciformes | Centropomatidae | Asian sea bass/Barramundi | Lates calcarifer | F | RGNNV | [48,68,81,83,84,85,86] | Taiwan, India, Singapore, Malaysia, Australia, Israel, China, Indonesia |
| Japanese sea bass | Lateolabrax japonicus | F | ND | [87] | Japan | ||
| Ephippidae | Orbicular batfish | Platax orbicularis | F | RGNNV | [88] | French Polynesia | |
| Carangidae | Striped Jack | Pseudocaranx dentex | F | SJNNV c | [6] | Japan | |
| Purplish amberjack | Seriola dumerili | F | ND | [76] | Japan | ||
| Pompano | Trachinotus blochii | F | RGNNV | [89] | Malaysia | ||
| Permit | T. falcatus | F | RGNNV | [83,84] | Taiwan | ||
| Golden pompano | T. ovatus | F | RGNNV | [90] | China | ||
| Lutjanidae | Firespot snapper | Lutjanus erythropterus | F | RGNNV | [83,91] | Taiwan, Malaysia | |
| Moronidae | European sea bass | Dicentrarchus labrax | F W | RGNNV, SJ/RG d RGNNV | [3,4,63,77,81,92,93,94,95] [96] | Martinique, Spain, Portugal, Mediterranean Italy | |
| Pomacentridae | Clownfish, | Amphiprion sebae Bleeker | F | RGNNV | [97] | India | |
| Rachycentridae | Cobia | Rachycentron canadum | F | RGNNV | [84] | Taiwan | |
| Sciaenidae | Red drum | Sciaenops ocellatus | F | RGNNV | [81,98] | Korea, Israel | |
| Shi drum | Umbrina cirrosa | W/F | RGNNV | [77,95,99,100] | France, Italy, Adriatic Sea | ||
| White seabass | Atractoscion nobilis | F | RGNNV | [73] | California (USA) | ||
| Scombridae | Pacific bluefin tuna | Thunnus orientalis | F | RGNNV | [101] | Japan | |
| Serranidae | White grouper | Epinephelus aeneus | F | RGNNV | [81] | Israel | |
| Red spotted grouper | E. akaara | F | ND | [102,103] | Taiwan, Japan | ||
| Yellow grouper | E. awooara | F | RGNNV | [104] | Taiwan, China | ||
| Duskytail grouper | E. bleekeri | F | RGNNV | [69] | China | ||
| Orange-spotted grouper | E. coioides | F | RGNNV | [84,105] | Philippines, Taiwan | ||
| Golden grouper | E. costae | W | RGNNV | [96,106] | Italy, Algeria | ||
| Blackspotted grouper | E. fuscogutatus | F | RGNNV | [85,91,102] | Taiwan, Malaysia | ||
| Dragon/Giant grouper | E. lanceolatus | F | RGNNV | [68,107,108] | China, Taiwan, Australia | ||
| Malabar grouper | E. malabaricus | F | RGNNV | [107] | Taiwan | ||
| Dusky grouper | E. marginatus | W | RGNNV | [96,106,109] | Italy, Algeria, Spain | ||
| Kelp grouper | E. moara | F | ND | [110] | Japan | ||
| Sevenband grouper | E. septemfasciatus | F | ND | [111,112] | Japan, Korea | ||
| Greasy grouper | E. tauvina | F | RGNNV | [15,113,114] | Singapore | ||
| Hybrid grouper | E. fuscoguttatus x E. lanceolatus | F | RGNNV | [115] | China, Indonesia | ||
| Humpback grouper | Chromileptes altivelis | F | RGNNV | [46,83,84,116,117,118] | Taiwan, Indonesia, Malasia, Vietnam, Australia, | ||
| Spottet coral grouper | Plectropomus maculatus | F | ND | [119] | Thailand | ||
| Siganidae | Dusky sinefoot | Siganus fuscescens | F | RGNNV | [68] | China | |
| Sparidae | Gilthead seabream | Sparus aurata | F | RG/SJ d | [55,57,120] | Iberian Peninsula, Italy, Arabian Gulf | |
| Sobaity seabream | Sparidentex hasta | F | ND | [120] | Arabian Gulf | ||
| Sparidae | White seabream | Dioplodus sargus | F | RGNNV | [95] | Italia, Adriatic sea | |
| Pleuronectiformes | Paralicthyidae | Japanese flounder | Paralichthys olivaceus | F | RGNNV | [16,42,121] | Japan, Korea |
| Pleuronectidae | Barfin flounder | Verasper moseri | F | BFNNV | [42,76] | Japan | |
| Atlantic halibut | Hippoglossus hippoglossus | F | BFNNV | [122,123,124] | Norway, United Kingdom | ||
| Scophthalmidae | Turbot | Scophthalmus maximus | F | TNNV | [43] | Norway | |
| Soleidae | Dover sole | Solea solea | F | BFNNV | [77] | United Kingdom | |
| Senegalese sole | S. senegalensis | F | SJNNV, RG/SJ | [53,55] | Iberian Peninsula | ||
| Scorpaeniformes | Sebastidae | Oblong rockfish | Sebastes oblongus | F | RGNNV | [121] | Korea |
| Tetraodontiformes | Monacanthidae | Thread-sail filefish | Stephanolepis cirrhifer | F | ND | [125] | Thailand |
| Tetraodontidae | Tiger puffer | Takifugu rubripes | F | TPNNV c | [110] | Japan |
| Order | Family | Common Name | Latin Name | Habitat | Geno-type | Detection | Reference | Geographic Range |
|---|---|---|---|---|---|---|---|---|
| Anguilliformes | Muraenidae | Blue ribbon eel | Rhinomuraena quaesita | Marine a | ND b | PCR | [126] | Korea/Indonesia d |
| Beryciformes | Monocentridae | Pinecone fish | Monocentris japonica | Marine a | ND | PCR | [126] | Korea/Japan d |
| Centrarchiformes | Oplegnathidae | Japanese parrotfish | Oplegnathus fasciatus | Marine | ND | PCR | [66,127] | Japan, Korea |
| Clupeiformes | Engraulidae | Japanese anchovy | Engraulis japonicus | Marine a | RGNNV | PCR | [126] | Korea/ Japand |
| Gonorynchiformes | Chanidae | Milk fish | Chanos chanos | Marine/ freshwater a | ND | PCR | [126] | Korea/Japan d |
| Mugiliformes | Mugilidae | Flathead grey mullet | Mugil cephalus | Farmed | RGNNV | PCR | [95] | Italy, Adriatic sea |
| Perciformes | Acanthuridae | Yellow tang | Zebrasoma flavescens | Marine a | ND | PCR | [126] | Korea/Singapore d |
| Carangidae | Look down fish | Selene vomer | Marine a | ND | PCR | [126] | Korea/N. America d | |
| Golden pompano | Trachinotus ovatus | Marine | RGNNV | PCR | [68] | China | ||
| Lateolabracidae | Chinese seabass | Lateolabrax sp | Marine | RGNNV | PCR | [66] | Japan | |
| Japanese sea perch | Lateolabrax japonicus | Marine | RGNNV | PCR | [68] | China | ||
| Moronidae | European sea bass | Dicentrarchus labrax | Marine | RGNNV | PCR, CC c, | [128] | Italy | |
| Mullidae | Red mullet | Mullus barbatus barbatus | Marine | RGNNV | PCR, CC | [95] | Italy, Adriatic Sea | |
| Polycentridae | South American leaf fish | Monocirrhus polyacanthus | Freshwater a | RGNNV | PCR, CC | [126] | Korea/Amazon d | |
| Pomacentridae | Three spot damsel | Dascyllus trimaculatus | Marine a | RGNNV | PCR, CC | [126] | Korea/Singapore d | |
| Serranidae | Brown-spotted grouper | E. chlorostigma | Marine | ND | PCR | [120] | Arabian Gulf | |
| Giant grouper | E. lanceolatus | Marine | RGNNV | PCR | [68] | China | ||
| Redspot grouper | Epinephelus akaara | Marine | RGNNV | PCR | [66] | Japan | ||
| Yellow grouper | E. awooara | Marine | RGNNV | PCR | [68] | China | ||
| Serrasalmidae | Red piranha | Pygocentrus nattereri | Freshwater a | RGNNV | PCR, CC | [126] | Korea/Amazon d | |
| Siganidae | Dusky sinefoot | Siganus fuscescens | Marine | RGNNV | PCR | [68] | China | |
| Sparidae | Gilthead seabream | Sparus aurata | Marine | ND, RGNNV | PCR | [63,95,129,130] | France (Atlantic),Mediterranean | |
| Red seabream | Pagrus major | Marine | RGNNV | PCR | [66,131] | Japan, Korea | ||
| Stromateidae | Silver pomfret | Pampus argenteus | Marine | RGNNV | PCR | [68] | China | |
| Pleuronectiformes | Paralicthyidae | Japanese flounder | Paralichthys olivaceus | Marine | RGNNV | PCR | [66,127] | Japan, Korea |
| Pleuronectidae | Marbled sole | Pleuronectes yokohamae | Marine | RGNNV | PCR | [66] | Japan | |
| Stone flounder | Kareius bicoloratus | Marine | RGNNV | PCR | [131] | Korea | ||
| Sciaenidae | Red drum | Sciaenops ocellatus | Marine | RGNNV | PCR | [68] | China | |
| Scopththalmidae | Turbot | Scophthalmus maximus | Marine | RGNNV | PCR, CC | [132] | Spain | |
| Scorpaeniformes | Sebastidae | Black rockfish | Sebastes inermis | Marine | RGNNV | PCR | [66] | Japan |
| Scorpaeniformes | Oblong rockfish | S. oblongus | Marine | RGNNV | PCR | [66] | Japan | |
| Schlegel’s black rockfish | S. schlegelii | Marine | RGNNV | PCR | [131] | Korea | ||
| Sebastidae | Spotbelly rockfish | S. pachycephalus | Marine | RGNNV | PCR | [66] | Japan | |
| Synanceiidae | Devil stinger | Inimicus japonicus | Marine | RGNNV | PCR | [66] | Japan | |
| Tetraodontiformes | Monacanthidae | Black scraper | Thamnaconus modestus | Marine | ND | PCR | [66] | Japan |
| Tetraodontidae | Tiger puffer | Takifugu rubripes | Marine | RGNNV | PCR | [66] | Japan | |
| Syngnathiformes | Centriscidae | Shrimp fish | Aeoliscus strigatus | Marine a | ND | PCR | [126] | Korea/Japan c |
| Order | Family | Common Name | Latin Name | Status | Genotype | Reference | Geographic Range |
|---|---|---|---|---|---|---|---|
| Acipenseriformes | Acipenseridae | Danube sturgeon | Acipenser gueldenstaedtii | Farmed | ND a | [133] | Greece |
| Anguilliformes | Anguillidae | European eel | Anguilla anguilla | Farmed | RGNNV | [83,84] | Taiwan |
| Cypriniformes | Cyprinidae | Goldfish | Carassius auratus | Ornamental | ND | [134] | India |
| Rainbow sharkminnow | Epalzeorhynchos frenatum | Ornamental | ND | [134] | India | ||
| Cyprinodontiformes | Poeciliidae | Guppy | Poecilia reticulata | Farmed | RGNNV | [17] | Singapore |
| Perciformes | Blenniidae | Freshwater blenny | Salaria flubiatilis | Wild | RGNNV | [135] | Spain |
| Centrarchidae | Largemouth black bass | Micropterus salmoides | Farmed | RGNNV | [136] | Italy | |
| Cichlidae | Tilapia | Oreochromis niloticus | Farmed | RGNNV | [49,137] | Europe, Indonesia | |
| Eleotridae | Sleepy cod | Oxyeleotris lineolatus | Farmed | RGNNV | [46] | Australia | |
| Moronidae | Hybrid striped bass | Morone saxatilis x M. chrysops | Farmed | RGNNV | [136] | Italy | |
| Percidae | Pike-perch | Sander lucioperca | Farmed | RGNNV | [136] | Italy | |
| Siluriformes | Plotosidae | Catfish | Tandanus tandanus | Farmed | ND | [74] | Australia |
| Siluridae | Chinese catfish | Parasilurus asotus | Farmed | RGNNV | [84] | Taiwan |
| Order | Family | Common Name | Latin Name | Geno- type | Detection | Reference | Geographic Range |
|---|---|---|---|---|---|---|---|
| Anguilliformes | Anguillidae | European eel | Anguilla anguilla | SJNNV | PCR | [138] | Spain |
| Congridae | Conger eel | Rhynchocymba nystromi | RGNNV | PCR | [68] | China | |
| Atheriniformes | Atherinidae | Mediterranean sand smelt | Atherina hepsetus | RGNNV | qPCR a | [58] | Greece |
| Ring-tailed cardinalfish | A. aureus | RGNNV | PCR | [68] | China | ||
| Aulopiformes | Synodontidae | Threadfin saury | Saurida filamentosa | RGNNV | PCR | [68] | China |
| Brushtooth lizardfish | S. lessepsianus | RGNNV | PCR | [130] | Mediterranean | ||
| Snakefish | Trachinocephalus myops | RGNNV | PCR | [68] | China | ||
| Bathrachoidiformes | Bathrachoididae | Lusitania toadfish | Halobatrachus didactylus | RGNNV | PCR, CC b | [64] | Spain |
| Beloniformes | Belonidae | Garpike | Belone belone | RGNNV | PCR, CC | [62] | Italy |
| Exocoetidae | Mediterranean flyingfish | Cheilopogon heterurus | RGNNV | PCR | [58] | Greece | |
| Centrarchiformes | Latridae | Striped trumpeter | Latris lineata | RGNNV | PCR | [46] | Australia |
| Oplegnathidae | Japanese parrotfish | Oplegnathus fasciatus | RGNNV | PCR | [66] | Japan | |
| Clupeiformes | Clupeidae | Pilchard | Sardina pilchardus | RGNNV | PCR, CC | [58,62,65] | Italy. Tunisia |
| Round sardinella | Sardinella aurita | RGNNV | PCR | [65,130] | Mediterranean | ||
| Sardine | S. jussieui | RGNNV | PCR | [68] | China | ||
| Dussumieriidae | Slender rainbow sardine | Dussumieria elopsoides | RGNNV | PCR | [130] | Mediterranean | |
| Engraulidae | Commerson´s anchovy | Tolephorus commersonnii | RGNNV | PCR | [68] | China | |
| European anchovy | Engraulis encrasicolus | RGNNV | PCR | [58] | Greece | ||
| Gadiformes | Gadidae | Atlantic cod | Gadus morhua | BFNNV | PCR | [47] | Norway |
| Poor cod | Trisopterus minutus | RGNNV | PCR, qPCR | [56,58] | Italy, Greece | ||
| Pollock | Pollachius pollachius | ND | PCR | [47] | Norway | ||
| Saithe | P. virens | ND | PCR | [47] | Norway | ||
| Whiting | Merlangius merlangus | RGNNV | PCR | [62] | Italy | ||
| Merluciidae | European hake | Merlucius merlucius | RGNNV | PCR, CC | [58,62] | Italy, Greece | |
| Macrouridae | Glasshead grenadier | Hymenocephalus italicus | ND | PCR | [139] | Italy | |
| Heterodontiformes | Heterodontidae | Japanese bullhead shark | Heterodontus japonicus | RGNNV | PCR | [66] | Japan |
| Kurtiformes | Apogonidae | Red stoplight cardinalfish | Apogon erythrinus | RGNNV | PCR | [68] | China |
| Mugiliformes | Mugilidae | Flathead grey mullet | Mugil cephalus | RGNNV | PCR, CC | [58,62,66,69] | Japan, Italy, China, Greece |
| Golden mullet | Liza auratus | RGNNV | PCR | [65] | Tunisia | ||
| Thicklip grey mullet | Chelon labrosus | RGNNV, SJNNV | PCR | [64] | Spain | ||
| Notacanthiformes | Notacanthidae | Shorfin spiny eel | Notacanthus bonaparte | ND | PCR | [139] | Italy |
| Perciformes | Carangidae | Atlantic horse mackerel | Trachurus thrachurus | RGNNV | PCR, CC | [58,62,65] | Italy, Tunisia |
| Japanese jack mackerel | T. japonicus | RGNNV, SJNNV | PCR, CC | [66,67,68,70,71] | Japan, China | ||
| Mediterranean horse mackerel | T. mediterraneus | RGNNV, RG/SJ | PCR, CC | [58,62] | Italy, Greece | ||
| Blue Jack mackerel | T. picturatus | ND | PCR, CC | [128] | Italy | ||
| Bigeye trevalli | Caranx oshimai | RGNNV | PCR | [69] | China | ||
| Greater amberjack | Seriola dumerili | RGNNV, SJNNV | PCR | [58,67] | Japan, Greece | ||
| Japanese amberjack | S. quinqueradiata | SJNNV | PCR | [67] | Japan | ||
| Indian threadfish | Alectis indica | RGNNV | PCR | [69] | China | ||
| Japanese scad | Decapterus maruadsi | RGNNV | PCR | [66] | Japan | ||
| Shrimp scad | Caranx djedaba | RGNNV | PCR | [69] | China | ||
| Yellowstripe scad | Selaroides leptolepis | RGNNV | PCR | [68,69] | China | ||
| Whitefin trevally | Carangoides equula | RGNNV, SJNNV | nPCR c | [67] | Japan | ||
| Centrolophidae | Pacific rudderfish | Psenopsis anomala | RGNNV | PCR | [68] | China | |
| Centropomatidae | Asian sea bass/ Barramundi | Lates calcarifer | RGNNV | PCR | [48] | India | |
| Epigonidae | Cardinal fish | Epigonus telescopus | ND | PCR | [139] | Italy | |
| Gobiidae | Black goby | Gobius niger | RGNNV | PCR, CC | [62] | Italy | |
| Haemulidae | Trout sweetlips | Plectorhynchus pictus | RGNNV | PCR | [69] | China | |
| Kyphosidae | Stripey fish | Microcanthus strigatus | RGNNV | PCR | [66] | Japan | |
| Labridae | Ballan wrasse | Labrus bergylta | BFNNV, RGNNV | PCR | [50] | Sweden | |
| Corkwing wrasse | Symphodus melops | BFNNV, RGNNV | PCR | [50] | Norway | ||
| Goldsinny wrasse | Ctenolabrus rupestris | BFNNV | PCR | [50] | Norway, Sweden | ||
| Leiognathidae | Pugnose ponyfish | Leigognathus insidiator | RGNNV | PCR | [69] | China | |
| Berber ponyfish | L. berbis | RGNNV | PCR | [69] | China | ||
| Slender ponyfish | L. elongatus | RGNNV | PCR | [68] | China | ||
| Lutjanidae | Dory snapper | Lutjanus fulviflamma | RGNNV | PCR | [69] | China | |
| Humphead snapper | L. sanguineus | RGNNV | PCR | [69] | China | ||
| John’s snapper | L. johni | RGNNV | PCR | [69] | China | ||
| Mangrove red snapper | L. argentimaculatus | RGNNV | PCR | [69] | China | ||
| Russell’s snapper | L. russelli | RGNNV | PCR | [69] | China | ||
| Moronidae | European sea bass | Dicentrarchus labrax | ND | qPCR | [128] | Italy | |
| Mullidae | Red mullet | Mullus barbatus barbatus | RGNNV | PCR, CC | [62,64,130] | Mediterranean | |
| Surmullet | M. surmuletus | RGNNV | PCR, qPCR | [56,58,130] | Mediterranean | ||
| Japanese goatfish | Upeneus japonicus | RGNNV | PCR | [68] | China | ||
| Goldband goatfish | U. moluccensis | RGNNV | PCR | [130] | Mediterranean | ||
| Nemipteridae | Japanese threadfin bream | Nemipterus japonicus | RGNNV | PCR | [68] | China | |
| Randall’s threadfin bream | N. randalli | RGNNV | PCR | [130] | Mediterranean | ||
| Percophidae | Ray finned fish | Chrionema chlorotaenia | RGNNV | PCR | [68] | China | |
| Pomacentridae | Heavenly damselfish | Pomacentrus coelestis | RGNNV | PCR | [66] | Japan | |
| Priacanthidae | Red bigeye | Priacanthus macracanthus | RGNNV | PCR | [68] | China | |
| Sciaenidae | Goatee croaker | Umbrina russelli | RGNNV | PCR | [69] | China | |
| Big head penah croaker | Argyrosomus macrocephalus | RGNNV | PCR | [68,69] | China | ||
| Silver croaker | A. argentatus | RGNNV | PCR | [69] | China | ||
| Meagre | A. regius | RGNNV, SJNNV | PCR | [140] | Spain | ||
| Brown meagre | Sciaena umbra | RGNNV | PCR, qPCR | [58,141] | Italy, Greece | ||
| Hoki | Johnius belengerii | RGNNV | PCR | [68] | China | ||
| Shy drum | Umbrina cirrosa | RGNNV | PCR | [43] | Italy | ||
| Scombridae | Atlantic mackerel | Scomber scombrus | ND | PCR | [47,58] | Norway, Greece | |
| Chub mackerel | S. japonicus | RGNNV, SJNNV | PCR | [67] | Japan | ||
| Atlantic bluefin tuna | Thunnus thynnus | RGNNV | PCR, qPCR | [58,142] | Japan, Greece | ||
| Serranidae | Areolate grouper | Epinephelus. areolatus | RGNNV | PCR | [69] | China | |
| Longfin grouper | E. megachir | RGNNV | PCR | [69] | China | ||
| Longspine grouper | E. fario | RGNNV | PCR | [69] | China | ||
| Dotted grouper | E. epistictus | RGNNV | PCR | [69] | China | ||
| Honeycumb grouper | E. merra | RGNNV | PCR | [69] | China | ||
| Kelp grouper | E. moara | RGNNV | PCR | [69] | China | ||
| Rock grouper | E. fasciatomaculatus | RGNNV | PCR | [69] | China | ||
| Siganidae | White-spotted spinefoot | Siganus oramin | RGNNV | PCR | [69] | China | |
| Sparidae | Bogue | Boops boops | RGNNV | qPCR | [58,128] | Italy, Greece | |
| Axillary seabream | Pagellus acarne | RGNNV | PCR, CC | [64] | Spain | ||
| Black seabream | Spondyliosoma cantharus | RGNNV | PCR, CC | [64] | Spain | ||
| Common two-banded seabream | Diploidus vulgaris | RGNNV | PCR, CC | [64] | Spain | ||
| Sharpsnout seabream | D. puntazzo | RGNNV | PCR, qPCR | [58] | Greece | ||
| Annular seabream | D. annularis | ND | qPCR | [198] | Italy | ||
| Red porgy | Pagrus pagrus | RGNNV | PCR, qPCR | [58,143] | Spain, Greece | ||
| Salema | Sarpa salpa | RGNNV | qPCR | [58] | Greece | ||
| Picarel | Spicara smaris | RGNNV | qPCR | [58] | Greece | ||
| Blotched picarel | S. maena | RGNNV | qPCR | [58] | Greece | ||
| Gilthead seabream | Sparus aurata | RGNNV | PCR, qPCR | [58,95] | Italy, Greece | ||
| Striped seabream | Lithognathus mormyrus | RGNNV | PCR | [130] | Mediterranean | ||
| Sphyraenidae | European barracuda | Sphiraena sphiraena | RGNNV | qPCR, CC | [58,128] | Italy, Greece | |
| Terapontidae | Fourlined terapon | Pelates quadrilineatus | RGNNV | PCR | [69] | China | |
| Trumpeter perch | P. quadrilineatus | RGNNV | PCR | [68] | China | ||
| Jarbua terapon | Terapon jarbua | RGNNV | PCR | [69] | China | ||
| Zanclidae | Moorish idol | Zanclus cornutus | RGNNV | PCR | [66] | Japan | |
| Pleuronectiformes | Citharidae | Branched ray flounder | Citharoides macrolepidotus | RGNNV | PCR | [68] | China |
| Pleuronectidae | European plaice | Pleuronectes platessa | ND | PCR | [47] | Norway | |
| Winter flounder | Pseudopleuronectes americanus | BFNNV | PCR, CC | [61] | Canada | ||
| Citharidae | Branched ray flounder | Citharoides macrolepidotus | RGNNV | PCR | [68] | China | |
| Siluriformes | Plotosidae | Striped catfish eel | Plotosus lineatus | RGNNV | PCR | [66] | Japan |
| Scorpaeniformes | Scorpaenidae | Luna lionfish | Pterois lunulata | RGNNV | PCR | [66] | Japan |
| Red scorpionfish | Scorpaena scrofa | RGNNV | qPCR | [58] | Greece | ||
| Sebastidae | Black rockfish | Sebastes inermis | ND | PCR | [66] | Japan | |
| Schlegel’s black rockfish | S. schlegelii | RGNNV | PCR | [66] | Japan | ||
| Marbled rockfish | Sebastiscus marmoratus | RGNNV | PCR | [66] | Japan | ||
| Triglidae | Gurnard | Tigla lyra | RGNNV | PCR | [62] | Italy | |
| Tub gurnard | Chelidonichtys lucerna | RGNNV | PCR, CC | [64] | Spain | ||
| Triglidae | Spiny gurnard | Lepidotrigla dieuzeidei | RGNNV | qPCR | [58] | Greece | |
| Syngnathiformes | Fistuliridae | Red cornetfish | Fistularia villosa | RGNNV | PCR | [68] | China |
| Trachichthyiformes | Trachichthyidae | Mediterranean slimehead | Hoplostethus mediterraneus | ND | PCR | [139] | Italy |
| Tetraodontiformes | Diodontidae | Freckled porcupine fish | Diodon holocanthus | RGNNV | PCR | [66] | Japan |
| Monacanthidae | Threadsail filefish | Stephanolepis cirrhifer | RGNNV | PCR | [66] | Japan | |
| Black scraper | Thamnaconus modestus | RGNNV | PCR | [66] | Japan | ||
| Tetraodontidae | Panther puffer | Takifugu pardalis | RGNNV | PCR | [66] | Japan |
| Order | Family | Common Name | Latin Name | Genotype | Detection | References | Geografic Range |
|---|---|---|---|---|---|---|---|
| Arcida | Arcidae | Granular ark | Tegillarca granosa | RGNNV, BFNNV | PCR | [131] | Korea, Japan |
| Mytiloida | Mytilidae | Mussel | Mytilus edulis | RGNNV, BFNNV | PCR | [131] | Korea |
| Mediterranean mussel | M. galloprovincialis | RGNNV | PCR | [58,200,201] | Korea, Italy, Greece | ||
| Oegopsida | Ommastrephidae | Japanese common squid | Todarodes pacificus | RGNNV | PCR, CC a | [201] | Japan |
| Ostreoida | Ostreidae | European flat oyster | Ostrea edulis | RGNNV | qPCR b | [58] | Greece |
| Pacific oyster | Cassostrea gigas | RGNNV, BFNNV | PCR | [131,200] | Korea, France | ||
| Octopoda | Octopodidae | Octopus | Octopus vulgaris | ND c | PCR, CC | [202] | Italy |
| Decapoda | Palinuridae | Spiny lobster | Pamulirus versicolor | ND | PCR | [126] | Japan |
| Pandalidae | Southern humpback shrimp | Pandalus hypsinotus | RGNNV | PCR | [201] | Korea | |
| Penaeidae | Kuruma prawn | Marsupenaeus japonicus | RGNNV | PCR | [130] | Mediterranean | |
| Portunidae | Charybdid crab | Charybdis bimaculata) | RGNNV | PCR | [201] | Korea | |
| Blue crab | Portunus pelagicus | RGNNV | PCR | [130] | Mediterranean | ||
| Pectinida | Pectinidae | Scallop | Patinopecten yessoensis | RGNNV, BFNNV | PCR | [131] | China, Japan |
| Veneroida | Veneridae | Clam | Ruditapes philipinarum | RGNNV | PCR | [200] | Italy |
| Common orient clam | Meretrix lusoria | RGNNV, BFNNV | PCR | [131] | Korea | ||
| Chinese cyclina | Cyclina sinensis | BFNNV | PCR | [131] | China | ||
| Manila clam | Venerupis philippinarum | RGNNV, BFNNV | PCR | [131] | Korea, China, Japan | ||
| Venus clam | Mercenaria mercenaria | BFNNV | PCR | [131] | China | ||
| Wrinkled venus clam | Callista brevisiphonata | BFNNV | PCR | [131] | China | ||
| Warty venus | Venus verrucosa | RGNNV | qPCR | [58] | Greece | ||
| Neogastropoda | Muricidae | Red-mouthed rockshell | Stramonita haemastoma | RGNNV | qPCR | [58] | Greece |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bandín, I.; Souto, S. Betanodavirus and VER Disease: A 30-year Research Review. Pathogens 2020, 9, 106. https://doi.org/10.3390/pathogens9020106
Bandín I, Souto S. Betanodavirus and VER Disease: A 30-year Research Review. Pathogens. 2020; 9(2):106. https://doi.org/10.3390/pathogens9020106
Chicago/Turabian StyleBandín, Isabel, and Sandra Souto. 2020. "Betanodavirus and VER Disease: A 30-year Research Review" Pathogens 9, no. 2: 106. https://doi.org/10.3390/pathogens9020106
APA StyleBandín, I., & Souto, S. (2020). Betanodavirus and VER Disease: A 30-year Research Review. Pathogens, 9(2), 106. https://doi.org/10.3390/pathogens9020106