Intracellular Behaviour of Three Legionella pneumophila Strains within Three Amoeba Strains, Including Willaertia magna C2c Maky


Abstract
1. Introduction
2. Results
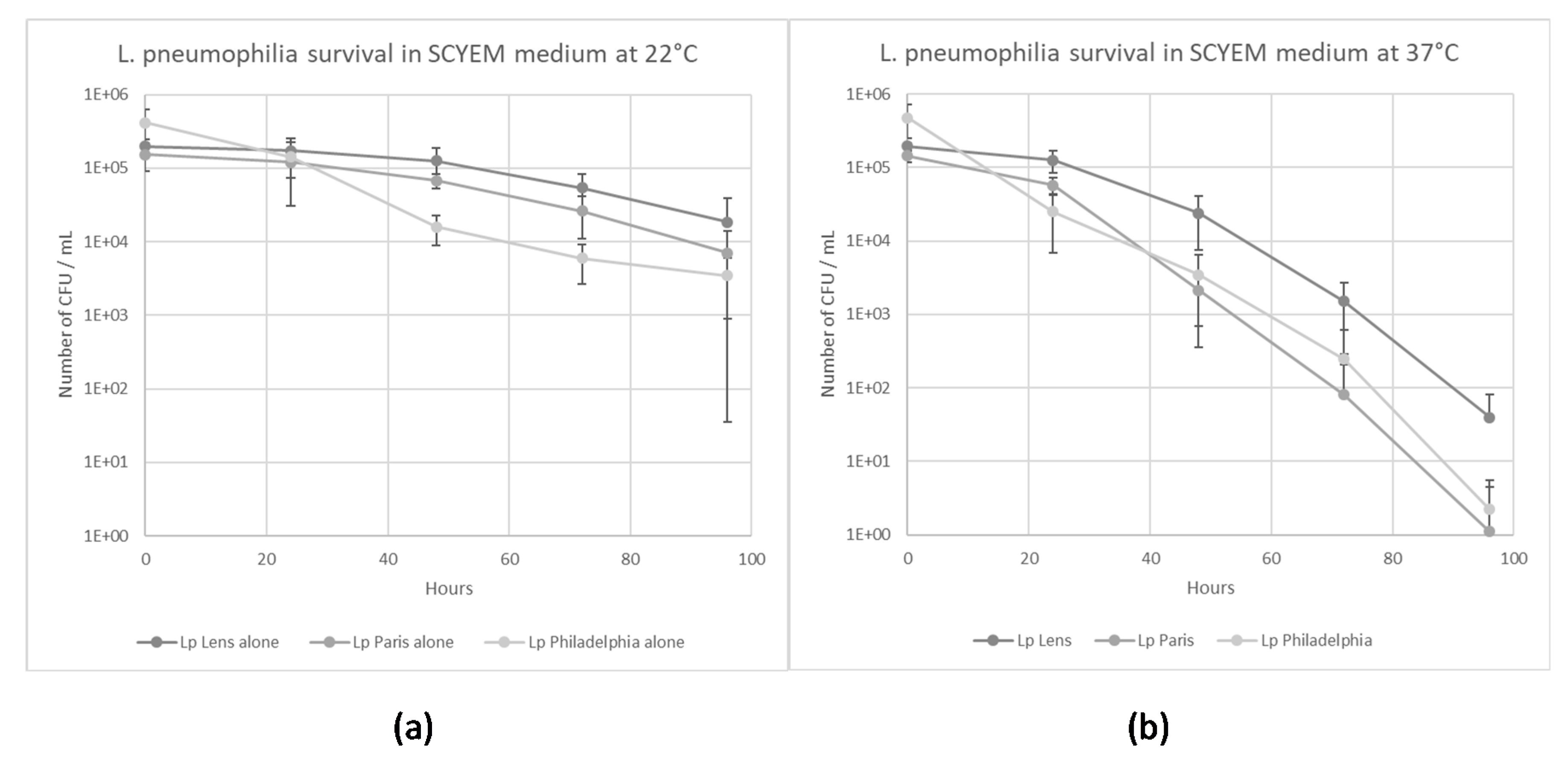
2.1. L. pneumophila Survival in Coculture Medium
2.2. Amoeba Survival in coculture Medium
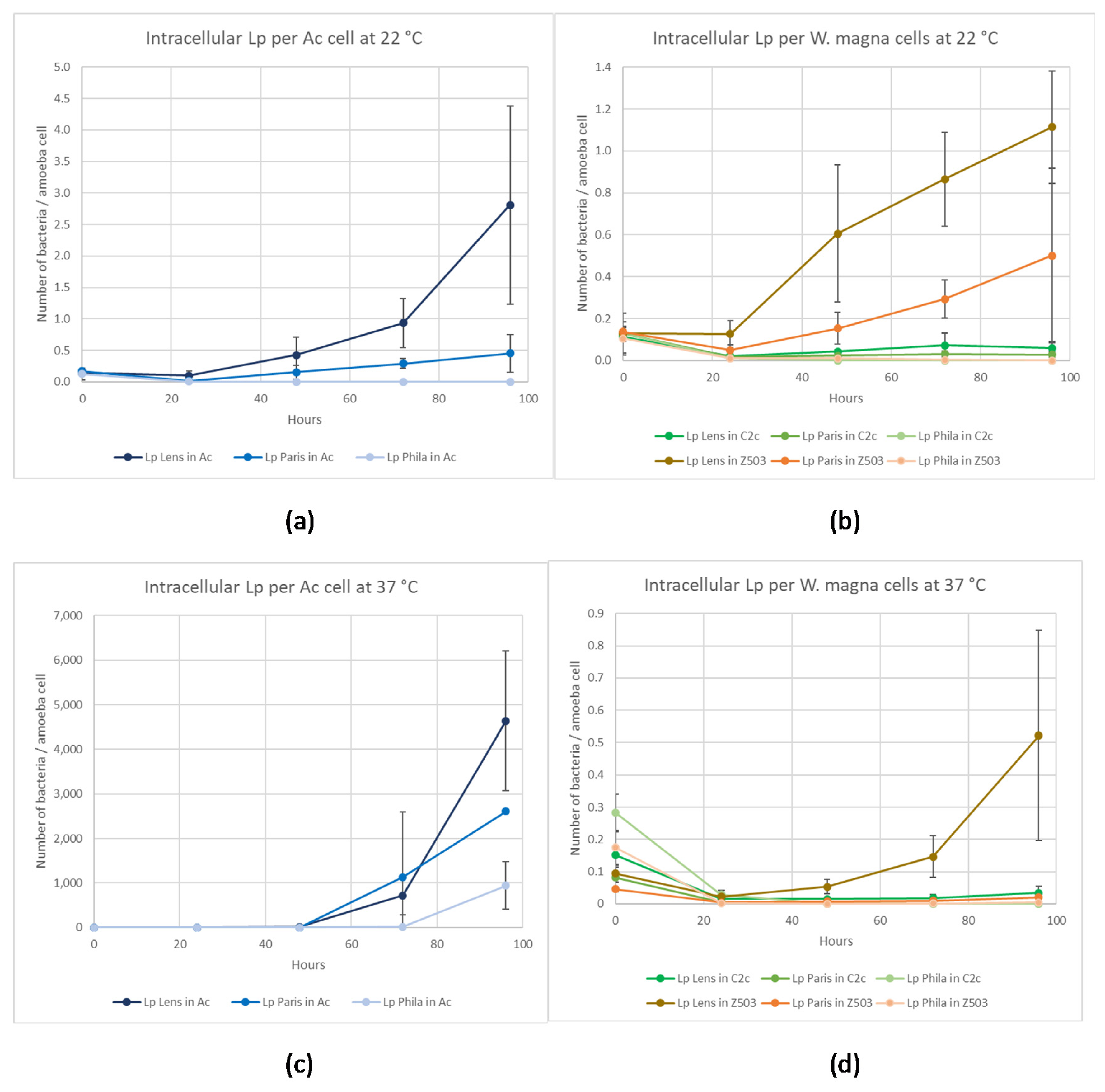
2.3. Co-Culture Experiments
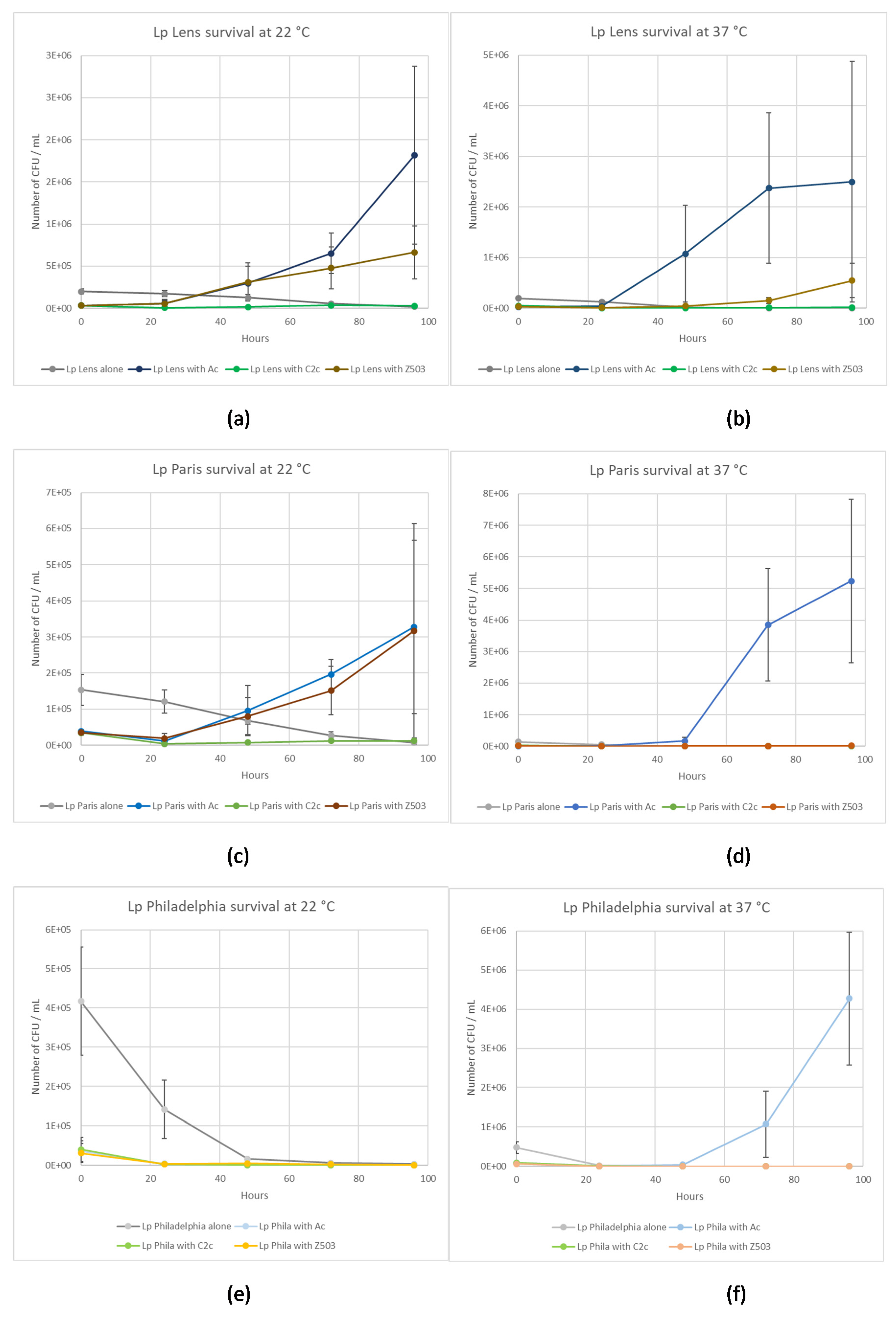
2.3.1. L. pneumophila Lens co-cultivated with Amoeba Strains
2.3.2. L. pneumophila Paris Co-Cultivated with Amoeba Strains
2.3.3. L. pneumophila Philadelphia Co-Cultivated with Amoeba Strains
2.4. Microscopic Observations of Intracellular L. pneumophila Philadelphia at 37 °C
2.5. Statistical Comparison of Amoeba Behavior
3. Discussion
4. Materials and Methods
4.1. Free-Living Amoebae Culture
4.2. Legionella Pneumophila Cultures
4.3. Bacterial Survival in the Coculture Medium (Control)
4.4. Amoeba Survival in the coculture Medium (Control)
4.5. Co-Culture Assays
4.6. L. pneumophila and Amoeba Quantifications in Co-Culture Assays from T0 to T0 + 96 h
4.7. Microscopic Observations in Co-Culture with L. pneumophila Philadelphia at 37 °C
4.8. Statistical Analyses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stout, J.E.; Yu, V.L. Legionellosis. N. Engl. J. Med. 1997, 337, 682–687. [Google Scholar] [CrossRef] [PubMed]
- Fraser, D.W.; Tsai, T.R.; Orenstein, W.; Parkin, W.E.; Beecham, H.J.; Sharrar, R.G.; Harris, J.; Mallison, G.F.; Martin, S.M.; McDade, J.E.; et al. Legionnaires’ Disease. N. Engl. J. Med. 1977, 297, 1189–1197. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, A.F.; McDade, J.E.; Patton, C.M.; Bennett, J.V.; Skaliy, P.; Feeley, J.C.; Anderson, D.C.; Potter, M.E.; Newhouse, V.F.; Gregg, M.B.; et al. Pontiac Fever: Isolation of the etiologic agent (legionella pneumophila) and demonstration of its mode of transmission. Am. J. Epidemiol. 1981, 114, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Benson, R.F.; Fields, B.S. Classification of the genus Legionella. Semin. Respir. Infect. 1998, 13, 90–99. [Google Scholar]
- Phin, N.; Parry-Ford, F.; Harrison, T.; Stagg, H.R.; Zhang, N.; Kumar, K.; Lortholary, O.; ZumLa, A.; Abubakar, I. Epidemiology and clinical management of Legionnaires’ disease. Lancet Infect. Dis. 2014, 14, 1011–1021. [Google Scholar] [CrossRef]
- Borella, P.; Guerrieri, E.; Marchesi, I.; Bondi, M.; Messi, P. Water ecology of Legionella and protozoan: Environmental and public health perspectives. Biotechnol. Annu. Rev. 2005, 11, 355–380. [Google Scholar]
- Hamilton, K.A.; Prussin, A.J.; Ahmed, W.; Haas, C.N. Outbreaks of Legionnaires’ Disease and Pontiac Fever 2006–2017. Curr. Environ. Health Rep. 2018, 5, 263–271. [Google Scholar] [CrossRef]
- Pine, L.; George, J.R.; Reeves, M.W.; Harrell, W.K. Development of a chemically defined liquid medium for growth of Legionella pneumophila. J. Clin. Microbiol. 1979, 9, 615–626. [Google Scholar]
- Rowbotham, T.J. Preliminary report on the pathogenicity of Legionella pneumophila for freshwater and soil amoebae. J. Clin. Pathol. 1980, 33, 1179–1183. [Google Scholar] [CrossRef]
- Yamamoto, H.; Sugiura, M.; Kusunoki, S.; Ezaki, T.; Ikedo, M.; Yabuuchi, E. Factors stimulating propagation of legionellae in cooling tower water. Appl. Environ. Microbiol. 1992, 58, 1394–1397. [Google Scholar] [CrossRef]
- Barbaree, J.M.; Fields, B.S.; Feeley, J.C.; Gorman, G.W.; Martin, W.T. Isolation of protozoa from water associated with a legionellosis outbreak and demonstration of intracellular multiplication of Legionella pneumophila. Appl. Environ. Microbiol. 1986, 51, 422–424. [Google Scholar] [CrossRef] [PubMed]
- Scheikl, U.; Sommer, R.; Kirschner, A.; Rameder, A.; Schrammel, B.; Zweimüller, I.; Wesner, W.; Hinker, M.; Walochnik, J. Free-living amoebae (FLA) co-occurring with legionellae in industrial waters. Eur. J. Protistol. 2014, 50, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Clarholm, M. Protozoan grazing of bacteria in soil—impact and importance. Microb. Ecol. 1981, 7, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Zaragoza, S. Ecology of Free-Living Amoebae. Crit. Rev. Microbiol. 1994, 20, 225–241. [Google Scholar] [CrossRef]
- Cirillo, J.D.; Falkow, S.; Tompkins, L.S. Growth of Legionella pneumophila in Acanthamoeba castellanii enhances invasion. Infect. Immun. 1994, 62, 3254–3261. [Google Scholar] [CrossRef]
- Greub, G.; Raoult, D. Microorganisms Resistant to Free-Living Amoebae. Clin. Microbiol. Rev. 2004, 17, 413–433. [Google Scholar] [CrossRef]
- Rowbotham, T.J. Isolation of Legionella pneumophila from clinical specimens via amoebae, and the interaction of those and other isolates with amoebae. J. Clin. Pathol. 1983, 36, 978–986. [Google Scholar] [CrossRef]
- Kilvington, S.; Price, J. Survival of Legionella pneumophila within cysts of Acanthamoeba polyphaga following chlorine exposure. J. Appl. Bacteriol. 1990, 68, 519–525. [Google Scholar] [CrossRef]
- Linder, J.W.-K. Ewert Free-living Amoebae Protecting Legionella in Water: The Tip of an Iceberg? Scand. J. Infect. Dis. 1999, 31, 383–385. [Google Scholar] [CrossRef]
- Dey, R.; Bodennec, J.; Mameri, M.O.; Pernin, P. Free-living freshwater amoebae differ in their susceptibility to the pathogenic bacterium Legionella pneumophila. FEMS Microbiol. Lett. 2009, 290, 10–17. [Google Scholar] [CrossRef]
- Aurell, H.; Etienne, J.; Forey, F.; Reyrolle, M.; Girardo, P.; Farge, P.; Decludt, B.; Campese, C.; Vandenesch, F.; Jarraud, S. Legionella pneumophila Serogroup 1 Strain Paris: Endemic Distribution throughout France. J. Clin. Microbiol. 2003, 41, 3320–3322. [Google Scholar] [CrossRef] [PubMed][Green Version]
- De Jonckheere, J.F.; Dive, D.G.; Pussard, M.; Vickerman, K. Willaertia Magna gen. nov. sp. nov. (Vahlkampfiidae), a Thermophilic Amoeba Found in Different Habitats. Available online: https://eurekamag.com/research/001/281/001281223.php (accessed on 6 February 2019).
- Robinson, B.S.; Christy, P.E.; De Jonckheere, J.F. A temporary flagellate (mastigote) stage in the vahlkampfiid amoeba Willaertia magna and its possible evolutionary significance. Biosystems 1989, 23, 75–86. [Google Scholar] [CrossRef]
- Hasni, I.; Chelkha, N.; Baptiste, E.; Mameri, M.R.; Lachuer, J.; Plasson, F.; Colson, P.; La Scola, B. Investigation of potential pathogenicity of Willaertia magna by investigating the transfer of bacteria pathogenicity genes into its genome. Sci. Rep. 2019, 9, 18318. [Google Scholar] [CrossRef] [PubMed]
- Ohno, A.; Kato, N.; Sakamoto, R.; Kimura, S.; Yamaguchi, K. Temperature-Dependent Parasitic Relationship between Legionella pneumophila and a Free-Living Amoeba (Acanthamoeba castellanii). Appl. Environ. Microbiol. 2008, 74, 4585–4588. [Google Scholar] [CrossRef]
- Casini, B.; Baggiani, A.; Totaro, M.; Mansi, A.; Costa, A.L.; Aquino, F.; Miccoli, M.; Valentini, P.; Bruschi, F.; Lopalco, P.L.; et al. Detection of viable but non-culturable legionella in hospital water network following monochloramine disinfection. J. Hosp. Infect. 2018, 98, 46–52. [Google Scholar] [CrossRef]
- Arslan-Aydoğdu, E.Ö.; Kimiran, A. An investigation of virulence factors of Legionella pneumophila environmental isolates. Braz. J. Microbiol. Publ. Braz. Soc. Microbiol. 2018, 49, 189–199. [Google Scholar] [CrossRef]
- García, M.T.; Jones, S.; Pelaz, C.; Millar, R.D.; Abu Kwaik, Y. Acanthamoeba polyphaga resuscitates viable non-culturable Legionella pneumophila after disinfection. Environ. Microbiol. 2007, 9, 1267–1277. [Google Scholar] [CrossRef]
- Molmeret, M.; Jarraud, S.; Mori, J.P.; Pernin, P.; Forey, F.; Reyrolle, M.; Vandenesch, F.; Etienne, J.; Farge, P. Different growth rates in amoeba of genotypically related environmental and clinical Legionella pneumophila strains isolated from a thermal spa. Epidemiol. Infect. 2001, 126, 231–239. [Google Scholar] [CrossRef]
- Cazalet, C.; Rusniok, C.; Brüggemann, H.; Zidane, N.; Magnier, A.; Ma, L.; Tichit, M.; Jarraud, S.; Bouchier, C.; Vandenesch, F.; et al. Evidence in the Legionella pneumophila genome for exploitation of host cell functions and high genome plasticity. Nat. Genet. 2004, 36, 1165–1173. [Google Scholar] [CrossRef]
- Buse, H.Y.; Ashbolt, N.J. Differential growth of Legionella pneumophila strains within a range of amoebae at various temperatures associated with in-premise plumbing. Lett. Appl. Microbiol. 2011, 53, 217–224. [Google Scholar] [CrossRef]
- Rowbotham, T.J. Pontiac fever, amoebae, and legionellae. Lancet Lond. Engl. 1981, 1, 40–41. [Google Scholar] [CrossRef]
- Dupuy, M.; Binet, M.; Bouteleux, C.; Herbelin, P.; Soreau, S.; Héchard, Y. Permissiveness of freshly isolated environmental strains of amoebae for growth of Legionella pneumophila. FEMS Microbiol. Lett. 2016, 363, fnw022. [Google Scholar] [CrossRef] [PubMed]
- Atlan, D.; Coupat-Goutaland, B.; Risler, A.; Reyrolle, M.; Souchon, M.; Briolay, J.; Jarraud, S.; Doublet, P.; Pélandakis, M. Micriamoeba tesseris nov. gen. nov. sp.: A new taxon of free-living small-sized Amoebae non-permissive to virulent Legionellae. Protist 2012, 163, 888–902. [Google Scholar] [CrossRef]
- Temmerman, R.; Vervaeren, H.; Noseda, B.; Boon, N.; Verstraete, W. Necrotrophic growth of Legionella pneumophila. Appl. Environ. Microbiol. 2006, 72, 4323–4328. [Google Scholar] [CrossRef]
- Richards, A.M.; Von Dwingelo, J.E.; Price, C.T.; Abu Kwaik, Y. Cellular microbiology and molecular ecology of Legionella-amoeba interaction. Virulence 2013, 4, 307–314. [Google Scholar] [CrossRef]
- McDade, J.E. Legionella and the Prevention of Legionellosis; Bartram, J., Chartier, Y., Lee, J.V., Bond, K., Surman-Lee, S., Eds.; World Health Organization: Geneva, Switzerland, 2007; ISBN 978-92-4-156297-3. [Google Scholar]
- Carducci, A.; Verani, M.; Battistini, R. Legionella in industrial cooling towers: Monitoring and control strategies. Lett. Appl. Microbiol. 2010, 50, 24–29. [Google Scholar] [CrossRef]
- Canals, O.; Serrano-Suárez, A.; Salvadó, H.; Méndez, J.; Cervero-Aragó, S.; Ruiz de Porras, V.; Dellundé, J.; Araujo, R. Effect of chlorine and temperature on free-living protozoa in operational man-made water systems (cooling towers and hot sanitary water systems) in Catalonia. Environ. Sci. Pollut. Res. Int. 2015, 22, 6610–6618. [Google Scholar] [CrossRef]
- Heimberger, T.; Birkhead, G.; Bornstein, D.; Same, K.; Morse, D. Control of nosocomial Legionnaires’ disease through hot water flushing and supplemental chlorination of potable water. J. Infect. Dis. 1991, 163, 413. [Google Scholar] [CrossRef]
- Thomas, V.; Bouchez, T.; Nicolas, V.; Robert, S.; Loret, J.F.; Lévi, Y. Amoebae in domestic water systems: Resistance to disinfection treatments and implication in Legionella persistence. J. Appl. Microbiol. 2004, 97, 950–963. [Google Scholar] [CrossRef]
- Abdel-Wahab, A.; Batchelor, B. Chloride Removal from Recycled Cooling Water Using Ultra-High Lime with Aluminum Process. Water Environ. Res. 2002, 74, 256–263. [Google Scholar] [CrossRef]
- Pagnier, I.; Merchat, M.; La Scola, B. Potentially pathogenic amoeba-associated microorganisms in cooling towers and their control. Future Microbiol. 2009, 4, 615–629. [Google Scholar] [CrossRef]
- Iervolino, M.; Mancini, B.; Cristino, S. Industrial Cooling Tower Disinfection Treatment to Prevent Legionella spp. Int. J. Environ. Res. Public. Health 2017, 14, 1125. [Google Scholar] [CrossRef]
- Farhat, M.; Moletta-Denat, M.; Frère, J.; Onillon, S.; Trouilhé, M.-C.; Robine, E. Effects of disinfection on Legionella spp., eukarya, and biofilms in a hot water system. Appl. Environ. Microbiol. 2012, 78, 6850–6858. [Google Scholar] [CrossRef]
- Maillard, J.-Y. Resistance of Bacteria to Biocides. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef]
- Wéry, N.; Bru-Adan, V.; Minervini, C.; Delgénes, J.-P.; Garrelly, L.; Godon, J.-J. Dynamics of Legionella spp. and bacterial populations during the proliferation of L. pneumophila in a cooling tower facility. Appl. Environ. Microbiol. 2008, 74, 3030–3037. [Google Scholar] [CrossRef]
- Rivera, F.; Lares, F.; Gallegos, E.; Ramirez, E.; Bonilla, P.; Calderon, A.; Martinez, J.J.; Rodriguez, S.; Alcocer, J. Pathogenic amoebae in natural thermal waters of three resorts of Hidalgo, Mexico. Environ. Res. 1989, 50, 289–295. [Google Scholar] [CrossRef]
- De Jonckheere, J. Use of an axenic medium for differentiation between pathogenic and nonpathogenic Naegleria fowleri isolates. Appl. Environ. Microbiol. 1977, 33, 751–757. [Google Scholar] [CrossRef]
- Gimenez, D.F. Staining rickettsiae in yolk-sac cultures. Stain Technol. 1964, 39, 135–140. [Google Scholar] [CrossRef]
- Greer, P.W.; Chandler, F.W.; Hicklin, M.D. Rapid demonstration of Legionella pneumophila in unembedded tissue. An adaptation of the Giménez stain. Am. J. Clin. Pathol. 1980, 73, 788–790. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 22 °C | 37 °C | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| L. pneumophila Lens | T0 | 24 h | 48 h | 72 h | 96 h | T0 | 24 h | 48 h | 72 h | 96 h |
| With A. castellanii | A | A | A | A | A | A | A | A | A | A |
| With W. magna Z503 | A | A | A | A | A | AB | AB | A | A | A |
| With W. magna C2c Maky | A | B | B | B | B | A | B | B | C | B |
| L. pneumophila Paris | T0 | 24 h | 48 h | 72 h | 96 h | T0 | 24 h | 48 h | 72 h | 96 h |
| With A. castellanii | A | A | A | A | A | C | A | A | A | A |
| With W. magna Z503 | A | A | A | A | A | B | A | B | B | B |
| With W. magna C2c Maky | A | B | B | B | B | A | B | C | C | C |
| L. pneumophila Philadelphia | T0 | 24 h | 48 h | 72 h | 96 h | T0 | 24 h | 48 h | 72 h | 96 h |
| With A. castellanii | A | A | A | AB | AB | AB | A | A | AB | AB |
| With W. magna Z503 | A | A | A | A | A | B | B | B | A | A |
| With W. magna C2c Maky | A | A | A | B | B | A | A | B | C | C |
| Co-Culture | AWS 1 Volume | BWS 2 Volume |
|---|---|---|
| L.p. Philadelphia + W. magna C2c Maky | 10 mL AWSC2C | 0.1 mL BWSPhila |
| L.p. Philadelphia + W. magna Z503 | 10 mL AWSZ503 | 0.1 mL BWSPhila |
| L.p. Philadelphia + A. castellanii. | 10 mL AWSAC | 0.1 mL BWSPhila |
| L.p. Paris + W. magna C2c Maky | 10 mL AWSC2C | 0.1 mL BWSParis |
| L.p. Paris + W. magna Z503 | 10 mL AWSZ503 | 0.1 mL BWSParis |
| L.p. Paris + A. castellanii. | 10 mL AWSAC | 0.1 mL BWSParis |
| L.p. Lens + W. magna C2c Maky | 10 mL AWSC2C | 0.1 mL BWSLens |
| L.p. Lens + W. magna Z503 | 10 mL AWSZ503 | 0.1 mL BWSLens |
| L.p. Lens + A. castellanii. | 10 mL AWSAC | 0.1 mL BWSLens |
| Control L.p. Philadelphia | 10 mL SCYEM | 0.1 mL BWSPhila |
| Control L.p. Paris | 10 mL SCYEM | 0.1 mL BWSParis |
| Control L.p. Lens | 10 mL SCYEM | 0.1 mL BWSLens |
| Control W. magna C2c Maky | 10 mL AWSC2C | 0 mL |
| Control W. magna Z503 | 10 mL AWSZ503 | 0 mL |
| Control A. castellanii | 10 mL AWSAC | 0 mL |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hasni, I.; Jarry, A.; Quelard, B.; Carlino, A.; Eberst, J.-B.; Abbe, O.; Demanèche, S. Intracellular Behaviour of Three Legionella pneumophila Strains within Three Amoeba Strains, Including Willaertia magna C2c Maky. Pathogens 2020, 9, 105. https://doi.org/10.3390/pathogens9020105
Hasni I, Jarry A, Quelard B, Carlino A, Eberst J-B, Abbe O, Demanèche S. Intracellular Behaviour of Three Legionella pneumophila Strains within Three Amoeba Strains, Including Willaertia magna C2c Maky. Pathogens. 2020; 9(2):105. https://doi.org/10.3390/pathogens9020105
Chicago/Turabian StyleHasni, Issam, Antoine Jarry, Benjamin Quelard, Antoine Carlino, Jean-Baptiste Eberst, Olivier Abbe, and Sandrine Demanèche. 2020. "Intracellular Behaviour of Three Legionella pneumophila Strains within Three Amoeba Strains, Including Willaertia magna C2c Maky" Pathogens 9, no. 2: 105. https://doi.org/10.3390/pathogens9020105
APA StyleHasni, I., Jarry, A., Quelard, B., Carlino, A., Eberst, J.-B., Abbe, O., & Demanèche, S. (2020). Intracellular Behaviour of Three Legionella pneumophila Strains within Three Amoeba Strains, Including Willaertia magna C2c Maky. Pathogens, 9(2), 105. https://doi.org/10.3390/pathogens9020105