
Dairy Sheep Played a Minor Role in the 2005–2010 Human Q Fever Outbreak in The Netherlands Compared to Dairy Goats

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Historic Results
2.1.1. Historic Information on Dairy Sheep and Dairy Goat Farming in The Netherlands
2.1.2. Studies and Events on Dairy Goat and Dairy Sheep Farms
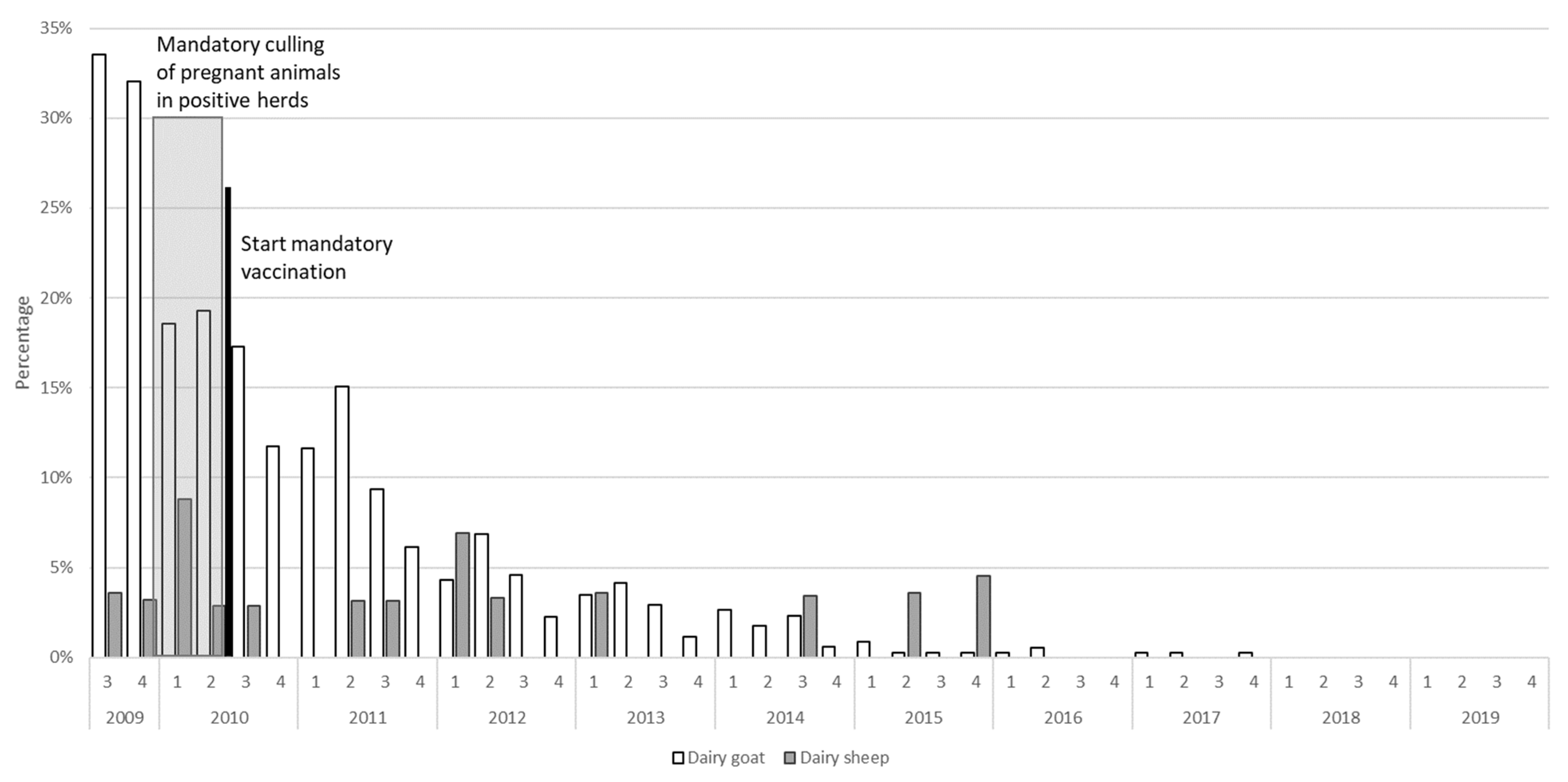
2.1.3. Measures Taken
2.2. Current Results
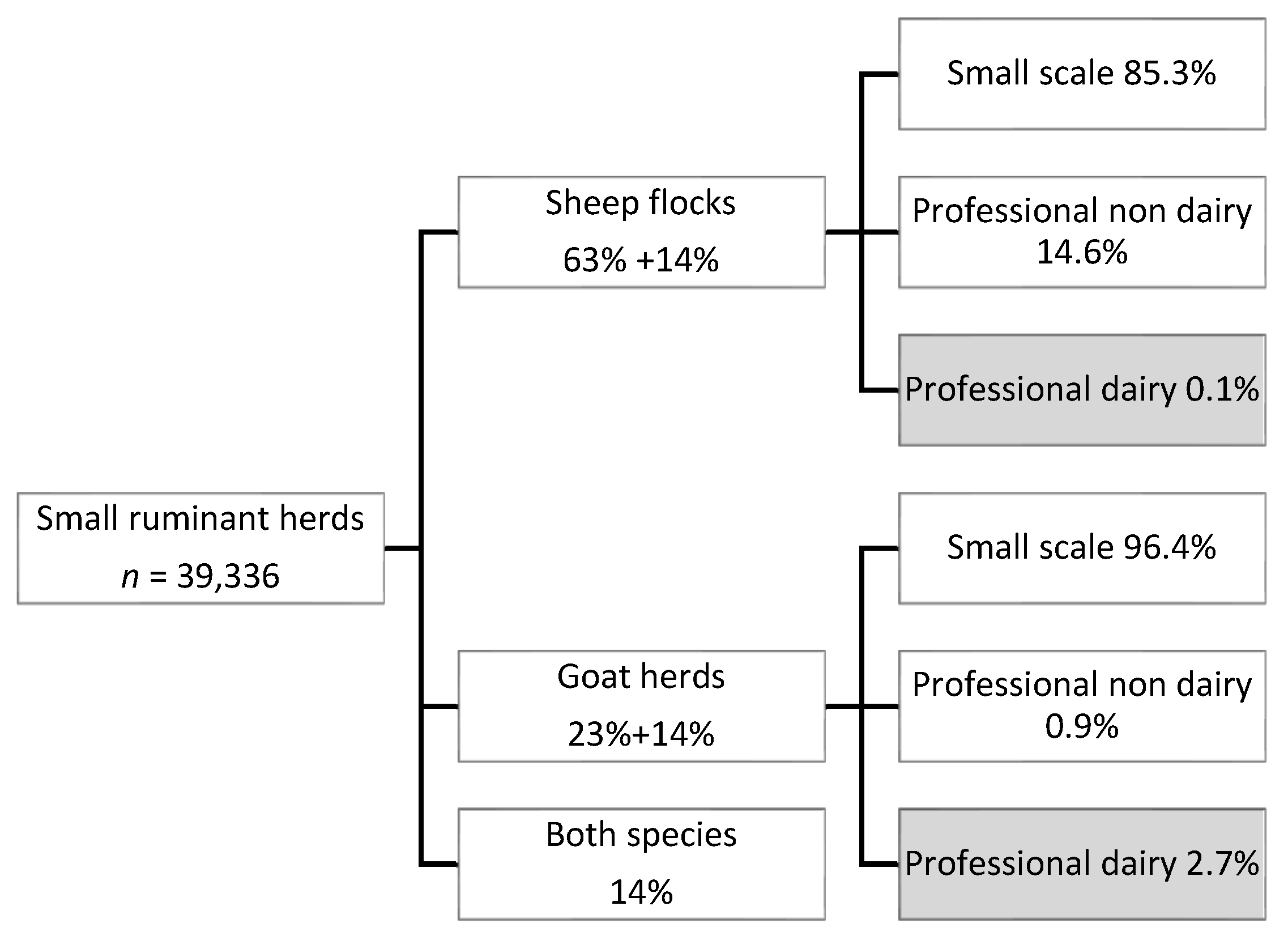
2.2.1. Goat and Sheep Farming in The Netherlands
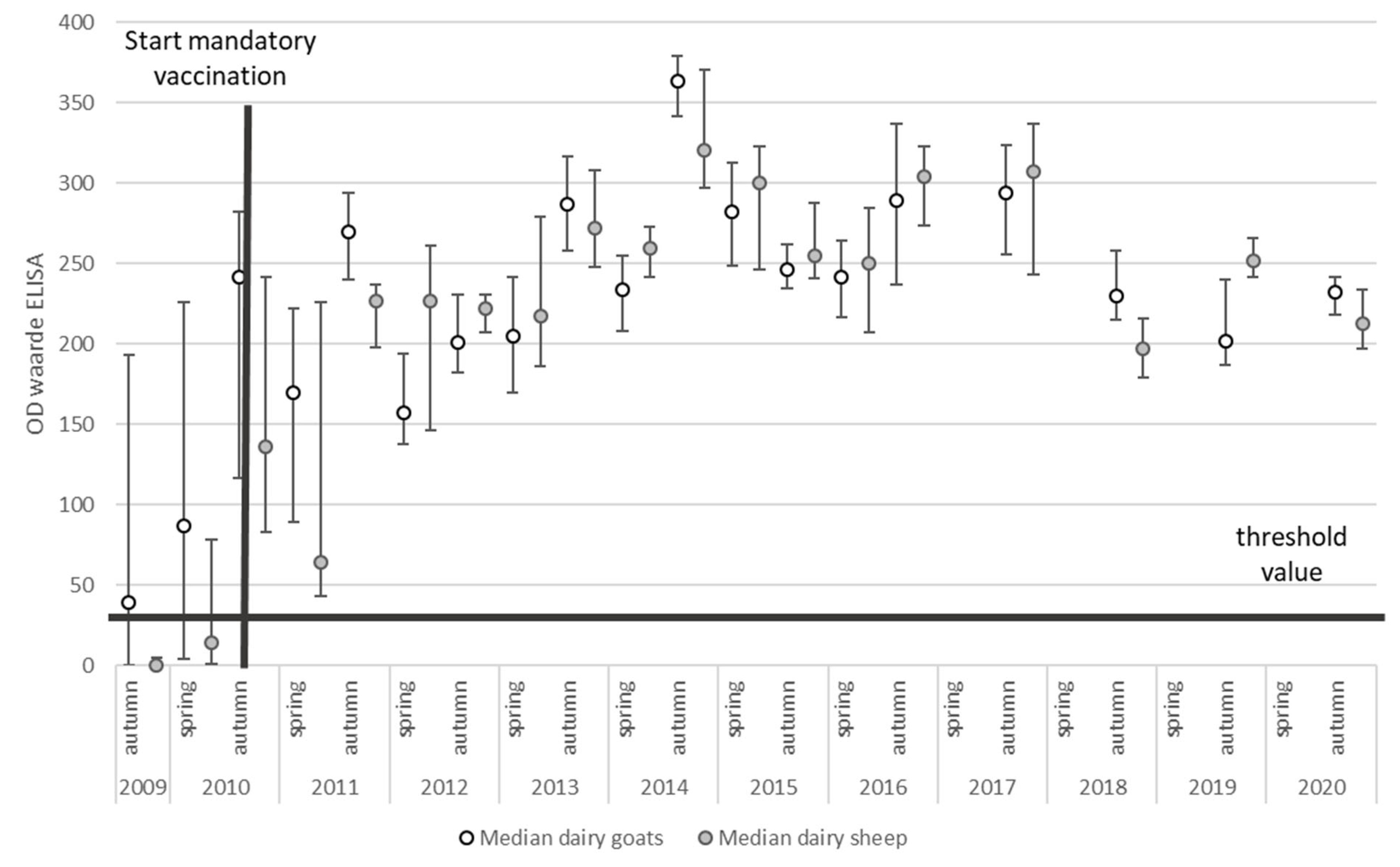
2.2.2. Bulk Tank Milk Monitoring
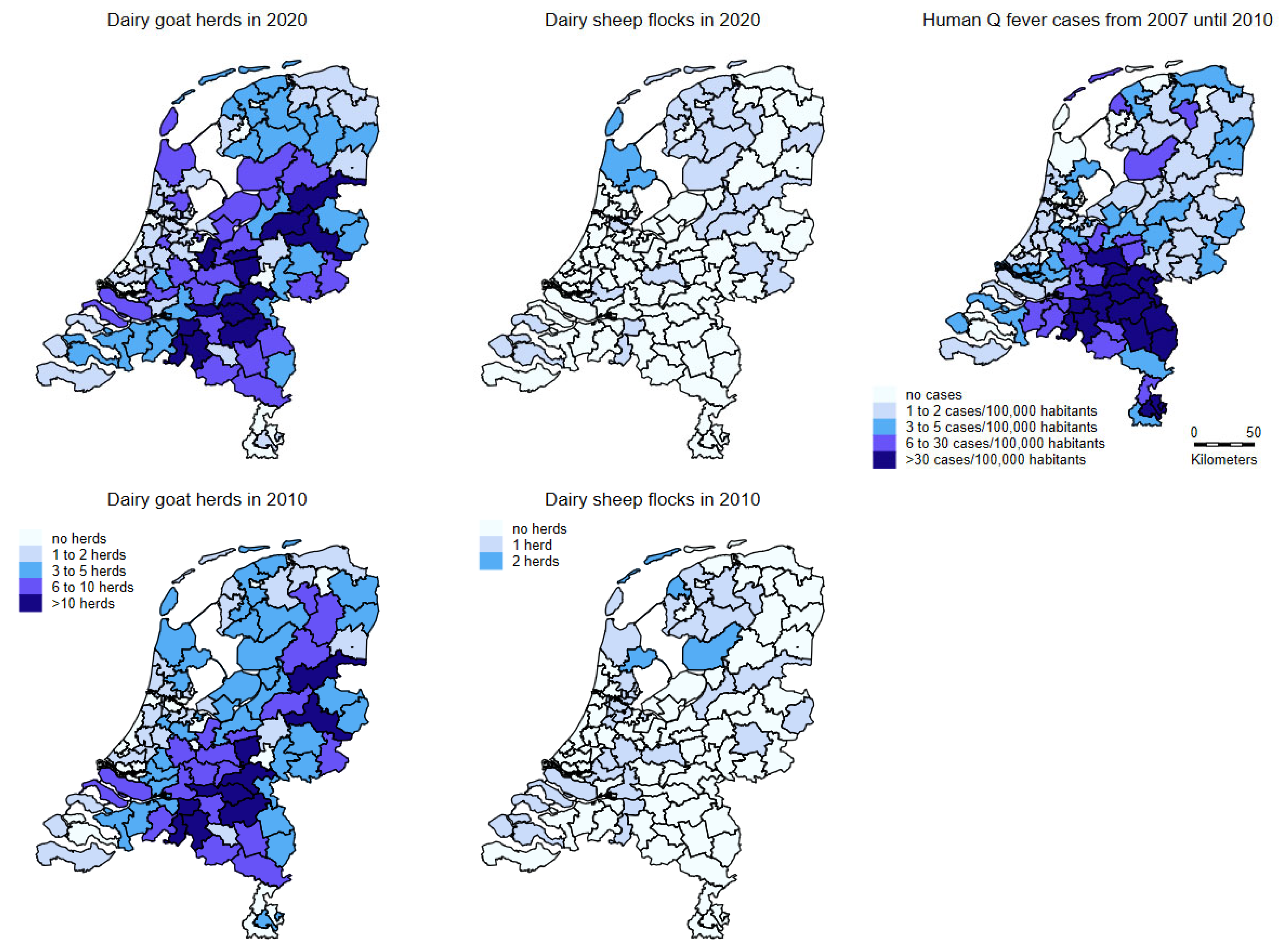
2.2.3. Human Notifications
3. Discussion
4. Materials and Methods
4.1. Historic Results
4.1.1. Historic Information on Dairy Sheep and Dairy Goat Farming in The Netherlands
4.1.2. Studies on Dairy Goat and Dairy Sheep Farms
4.2. Current Results
4.2.1. Goat and Sheep Farming in The Netherlands
4.2.2. Bulk Tank Milk Monitoring
4.2.3. PCR Testing
4.2.4. ELISA Testing
4.2.5. Human Notifications
4.2.6. Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Babudieri, B.; Moscovici, C. Experimental and natural infections of birds by Coxiella burnetii. Nature 1952, 169, 195–196. [Google Scholar] [CrossRef]
- Arricau-Bouvery, N.; Rodolakis, A. Is Q fever an emerging or re-emerging zoonosis? Vet. Res. 2005, 36, 327–349. [Google Scholar] [CrossRef]
- Zeman, D.H.; Kirkbride, C.A.; Leslie-Steen, P.; Duimstra, J.R. Ovine abortion due to Coxiella burnetii infection. J. Vet. Diagn Investig. 1989, 1, 178–180. [Google Scholar] [CrossRef] [PubMed]
- Damoser, J.; Hofer, E.; Müller, M. Abortions in a lower Austrian sheep facility caused by Coxiella burnetii. Berl. Munch. Tierarztl. Wochenschr. 1993, 106, 361–364. [Google Scholar] [PubMed]
- Maurin, M.; Raoult, D. Q fever. Clin. Microbiol. Rev. 1999, 12, 518–553. [Google Scholar] [CrossRef] [PubMed]
- Hatchette, T.F.; Hudson, R.C.; Schlech, W.F.; Campbell, N.A.; Hatchette, J.E.; Ratnam, S.; Raoult, D.; Donovan, C.; Marrie, T.J. Goat-associated Q fever: A new disease in Newfoundland. Emerg. Infect. Dis. 2001, 7, 413–419. [Google Scholar] [CrossRef]
- Arricau-Bouvery, N.; Souriau, A.; Bodie, C.; Dufour, P.; Rousset, E.; Rodolakis, A. Effect of vaccination with phase 1 and phase Coxiella burnetii vaccines in pregnant goats. Vaccine 2005, 23, 4392–4402. [Google Scholar] [CrossRef]
- Wouda, W.; Dercksen, D.P. Abortion and stillbirth among dairy goats as a consequence of Coxiella burnetii. Tijdschr. Diergeneeskd. 2007, 132, 908–911. [Google Scholar]
- Roest, H.J.; van Gelderen, B.; Dinkla, A.; Frangoulidis, D.; van Zijderveld, F.G.; Rebel, J.; van Keulen, L. Q fever in pregnant goats: Pathogenesis and excretion of Coxiella burnetii. PLoS ONE 2012, 7, 14. [Google Scholar] [CrossRef]
- Marrie, T.J. Q fever—A review. Can. Vet. J. 1990, 31, 555–563. [Google Scholar]
- Schimmer, B.; ter Schegget, R.; Wegdam, M.; Zuchner, L.; de Bruin, A.; Schneeberger, P.M.; Veenstra, T.; Vellema, P.; van der Hoek, W. The use of a geographic information system to identify a dairy goat farm as the most likely source of an urban Q-fever outbreak. BMC Infect. Dis. 2010, 10, 69. [Google Scholar] [CrossRef]
- Derrick, E.H. Q fever, new fever entity: Clinical features, diagnosis and laboratory investigation. Med. J. Aust. 1937, 2, 282–299. [Google Scholar] [CrossRef]
- Burnet, F.M.; Freeman, M. Experimental studies on the virus of Qfever. Med. J. Aust. 1937, 2, 299–302. [Google Scholar] [CrossRef]
- Davis, G.E.; Cox, H.R. A filter-passing infectious agent isolated from ticks. I. Isolation from Dermacentor andersoni, reactions in animals, and filtration experiments. Public Health Rep. 1938, 53, 9. [Google Scholar] [CrossRef]
- Cox, H.R. A filter-passing infectious agent isolated from ticks III. Description of organism and cultivation experiments. Public Health Rep. 1938, 53, 7. [Google Scholar]
- Cox, H.R. Studies of a filter-passing infectious agent isolated from ticks. V. Further attempts to cultivate in cell-free media. Suggested classification. Public Health Rep. 1939, 54, 1822–1827. [Google Scholar] [CrossRef]
- Dyer, R.E. A filter-passing infectious agent isolated from ticks IV. Human infection. Public Health Rep. 1938, 53, 6. [Google Scholar]
- Cox, H.R.; Bell, E.J. The cultivation of Rickettsia diaporica in tissue culture and in the tissues of developing chick embryos. Public Health Rep. 1939, 54, 2171–2176. [Google Scholar] [CrossRef]
- Derrick, E.H. Rickettsia burneti: The cause of ‘Q’ fever. Med. J. Aust. 1939, 1, 14. [Google Scholar] [CrossRef]
- Philip, C.B. Comments of the name of the Q fever organism. Public Health Rep. 1948, 63, 58. [Google Scholar] [CrossRef]
- Vellema, P.; Van den Brom, R. The rise and control of the 2007–2012 human Q fever outbreaks in The Netherlands. Small Rumin. Res. 2014, 118, 69–78. [Google Scholar] [CrossRef]
- Imhäuser, K. Viruspneumonien: Q-Fieber und Virusgrippe. Klin. Wochenschr. 1949, 27, 353–360. [Google Scholar] [CrossRef]
- Kaplan, M.M.; Bertagna, P. The geographical distribution of Q fever. Bull. World Health Organ. 1955, 13, 829–860. [Google Scholar] [PubMed]
- Woldehiwet, Z. Q fever (coxiellosis): Epidemiology and pathogenesis. Res. Vet. Sci. 2004, 77, 93–100. [Google Scholar] [CrossRef]
- Hilbink, F.; Penrose, M.; Kovacova, E.; Kazar, J. Q fever is absent from New Zealand. Int. J. Epidemiol. 1993, 22, 945–949. [Google Scholar] [CrossRef] [PubMed]
- Fox-Lewis, A.; Isteed, K.; Austin, P.; Thompson-Faiva, H.; Wolfgang, J.; Ussher, J.E. A case of imported Q fever in New Zealand. Int. J. Epidemiol. 1993, 22, 945–949. [Google Scholar] [PubMed]
- Westra, S.A.; Lopes Cardozo, E.; ten Berg, J. The first cases of Q fever in The Netherlands. Ned. Tijdschr. Geneeskd. 1958, 102, 69–72. [Google Scholar]
- Dijkstra, F.; van der Hoek, W.; Wijers, N.; Schimmer, B.; Rietveld, A.; Wijkmans, C.J.; Vellema, P.; Schneeberger, P.M. The 2007–Q fever epidemic in The Netherlands: Characteristics of notified acute Q fever patients and the association with dairy goat farming. FEMS Immunol. Med. Microbiol. 2012, 64, 3–12. [Google Scholar] [CrossRef]
- Van den Brom, R.; van Engelen, E.; Roest, H.I.; van der Hoek, W.; Vellema, P. Coxiella burnetii infections in sheep or goats: An opinionated review. Vet. Microbiol. 2015, 181, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Jansen, J. It Fryske Melkskiep. Skiednis en ûntjouwing; Fryske Akademy: Ljouwert, The Netherlands, 1985. [Google Scholar]
- Dercksen, D.P.; ter Laak, E.A.; Schreuder, B.E. Eradication programme for caseous lymphadenitis in goats in The Netherlands. Vet. Rec. 1996, 138, 237. [Google Scholar] [CrossRef]
- Santman-Berends, I.M.G.A.; Van den Brom, R.; van Schaik, G.; Vellema, P. Data-Analyses of the Small Ruminant Industry in 2012; GD Animal Health Service: Deventer, The Netherlands, 2013. [Google Scholar]
- Van den Brom, R.; Vellema, P. Q fever outbreaks in small ruminants and people in The Netherlands. Small Rumin. Res. 2009, 86, 74–79. [Google Scholar] [CrossRef]
- Van den Wijngaard, C.C.; Dijkstra, F.; van Pelt, W.; van Asten, L.; Kretzschmar, M.; Schimmer, B.; Nagelkerke, N.J.; Vellema, P.; Donker, G.A.; Koopmans, M.P. In search of hidden Q-fever outbreaks: Linking syndromic hospital clusters to infected goat farms. Epidemiol. Infect. 2011, 139, 19–26. [Google Scholar] [CrossRef]
- van den Brom, R.; Santman-Berends, M.; Dijkman, R.; Vellema, P.; Dijkman, R.; van Engelen, E. An Accessible Diagnostic Toolbox to Detect Bacterial Causes of Ovine and Caprine Abortion. Pathogens 2021, 10, 1147. [Google Scholar] [CrossRef]
- Van den Brom, R.; Moll, L.; van Schaik, G.; Vellema, P. Demography of Q fever seroprevalence in sheep and goats in The Netherlands in 2008. Prev. Vet. Med. 2013, 109, 76–82. [Google Scholar] [CrossRef]
- Van den Brom, R.; van Engelen, E.; Luttikholt, S.; Moll, L.; van Maanen, K.; Vellema, P. Coxiella burnetii in bulk tank milk samples from dairy goat and dairy sheep farms in The Netherlands in 2008. Vet. Rec. 2012, 170, 310. [Google Scholar] [CrossRef]
- Ter Bogt-Kappert, C.; van den Brom, R.; Dijkstra, E.; Gongrijp, M.; Heuvelink, A.; Peterson, K.; Dijkman, R.; Vellema, P.; Werkman, A. Jaarverslag Monitoring Diergezondheid Kleine Herkauwers 2020; Royal GD: Deventer, The Netherlands, 2020. [Google Scholar]
- Georgiev, M.; Afonso, A.; Neubauer, H.; Needham, H.; Thiery, R.; Rodolakis, A.; Roest, H.; Stark, K.; Stegeman, J.; Vellema, P.; et al. Q fever in humans and farm animals in four European countries, 1982 to 2010. Eurosurveillance 2013, 18, 20407. [Google Scholar] [CrossRef] [PubMed]
- Van den Brom, R.; Roest, H.J.; de Bruin, A.; Dercksen, D.; Santman-Berends, I.; van der Hoek, W.; Dinkla, A.; Vellema, J.; Vellema, P. A probably minor role for land-applied goat manure in the transmission of Coxiella burnetii to humans in the 2007–Dutch Q fever outbreak. PLoS ONE 2015, 10, e0121355. [Google Scholar] [CrossRef] [PubMed]
- Hermans, T.; Jeurissen, L.; Hackert, V.; Hoebe, C. Land-applied goat manure as a source of human Q fever in The Netherlands, 2006–2010. PLoS ONE 2014, 9, e96607. [Google Scholar] [CrossRef] [PubMed]
- Van den Brom, R.; Santman-Berends, I.; Luttikholt, S.; Moll, L.; Van Engelen, E.; Vellema, P. Bulk tank milk surveillance as a measure to detect Coxiella burnetii shedding dairy goat herds in The Netherlands between 2009 and 2014. J. Dairy Sci. 2015, 98, 3814–3825. [Google Scholar] [CrossRef]
- Hogerwerf, L.; van den Brom, R.; Roest, H.I.; Bouma, A.; Vellema, P.; Pieterse, M.; Dercksen, D.; Nielen, M. Reduction of Coxiella burnetii prevalence by vaccination of goats and sheep, The Netherlands. Emerg. Infect. Dis. 2011, 17, 379–386. [Google Scholar] [CrossRef]
- Roest, H.I.J.; Tilburg, J.J.H.C.; Van der Hoek, W.; Vellema, P.; Van Zijderveld, F.G.; Klaassen, C.H.; Raoult, D. The Q fever epidemic in The Netherlands: History, onset, response and reflection. Epidemiol. Infect. 2010, 5, 1–12. [Google Scholar] [CrossRef]
- Roest, H.I.; van Solt, C.B.; Tilburg, J.J.; Klaassen, C.H.; Hovius, E.K.; Roest, F.T.; Vellema, P.; van den Brom, R.; van Zijderveld, F.G. Search for possible additional reservoirs for human Q fever, The Netherlands. Emerg. Infect. Dis. 2013, 19, 834–835. [Google Scholar] [CrossRef]
- RIVM. Available online: https://www.rivm.nl/q-koorts (accessed on 30 September 2021).
- Holstege, M.; Gongrijp, M.; van den Brom, R.; Dijkstra, E.; ter Bogt-Kappert, C.; Santman-Berends, I.; Vellema, P. Data-Analysis of the Small Ruminant Industry in 2020; Royal GD: Deventer, The Netherlands, 2021. [Google Scholar]
- Martinov, S.P.; Neikov, P.; Popov, G.V. Experimental Q fever in sheep. Eur. J. Epidemiol. 1989, 5, 428–431. [Google Scholar] [CrossRef]
- Van den Brom, R.; van Engelen, E.; Vos, J.; Luttikholt, S.J.M.; Moll, L.; Roest, H.I.J.; van der Heijden, H.M.J.F.; Vellema, P. Detection of Coxiella burnetii in the bulk tank milk from a farm with vaccinated goats, by using a specific PCR technique. Small Rumin. Res. 2013, 110, 150–154. [Google Scholar] [CrossRef]
- Stuen, S.; Longbottom, D. Treatment and control of chlamydial and rickettsial infections in sheep and goats. Vet. Clin. N. Am. Food Anim. Pract. 2011, 27, 213–233. [Google Scholar] [CrossRef]
- Whelan, J.; Schimmer, B.; Schneeberger, P.; Meekelenkamp, J.; IJff, A.; Van Der Hoek, W.; van Beest, M.; Mirna Robert, D.R. Q Fever among Culling Workers, The Netherlands, 2009–2010. Emerg. Infect. Dis. 2011, 17, 1719–1723. [Google Scholar] [CrossRef]
- Porten, K.; Rissland, J.; Tigges, A.; Broll, S.; Hopp, W.; Lunemann, M.; van Treeck, U.; Kimmig, P.; Brockmann, S.O.; Wagner-Wiening, C.; et al. A super-spreading ewe infects hundreds with Q fever at a farmers’ market in Germany. BMC Infect. Dis. 2006, 6, 147. [Google Scholar] [CrossRef]
- Gilsdorf, A.; Kroh, C.; Grimm, S.; Jensen, E.; Wagner-Wiening, C.; Alpers, K. Large Q fever outbreak due to sheep farming near residential areas, Germany, 2005. Epidemiol. Infect. 2008, 136, 1084–1087. [Google Scholar] [CrossRef]
- De Lange, M.M.; Schimmer, B.; Vellema, P.; Hautvast, J.L.; Schneeberger, P.M.; Van Duijnhoven, Y.T. Coxiella burnetii seroprevalence and risk factors in sheep farmers and farm residents in The Netherlands. Epidemiol. Infect. 2014, 142, 1231–1244. [Google Scholar] [CrossRef] [PubMed]
- Koene, R.P.M.; Schimmer, B.; Rensen, H.; Biesheuvel, M.; de Bruin, A.; Lohuis, A.; Horrevorts, A.; Verduyn Lunel, F.; Delsing, C.E.; Hautvast, J.L.A. A Q fever outbreak in a psychiatric care institution in The Netherlands. Epidemiol. Infect. 2010, 139, 13–18. [Google Scholar] [CrossRef]
- Whelan, J.; Schimmer, B.; de Bruin, A.; van Beest Holle, M.R.; van der Hoek, W.; ter Schegget, R. Visits on ‘lamb-viewing days’ at a sheep farm open to the public was a risk factor for Q fever in 2009. Epidemiol. Infect. 2012, 140, 858–864. [Google Scholar] [CrossRef] [PubMed]
- Schimmer, B.; Dijkstra, F.; Vellema, P.; Schneeberger, P.M.; Hackert, V.; ter Schegget, R.; Wijkmans, C.; van Duynhoven, Y.; van der Hoek, W. Sustained intensive transmission of Q fever in the south of The Netherlands, 2009. Eurosurveillance 2009, 14, 19210. [Google Scholar] [CrossRef] [PubMed]
- Van der Hoek, W.; van de Kassteele, J.; Bom, B.; de Bruin, A.; Dijkstra, F.; Schimmer, B.; Vellema, P.; ter Schegget, R.; Schneeberger, P.M. Smooth incidence maps give valuable insight into Q fever outbreaks in The Netherlands. Geospat. Health. 2012, 7, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Ladbury, G.A.; Van Leuken, J.P.; Swart, A.; Vellema, P.; Schimmer, B.; Ter Schegget, R.; Van der Hoek, W. Integrating interdisciplinary methodologies for One Health: Goat farm re-implicated as the probable source of an urban Q fever outbreak, The Netherlands, 2009. BMC Infect. Dis. 2015, 15, 372. [Google Scholar] [CrossRef] [PubMed]
- Huijsmans, C.J.J.; Schellekens, J.J.A.; Wever, P.C.; Toman, R.; Savelkoul, P.H.M.; Janse, I.; Hermans, M.H.A. Single-nucleotide-polymorphism genotyping of Coxiella burnetii during a Q fever outbreak in The Netherlands. Appl. Environ. Microbiol. 2011, 77, 2051–2057. [Google Scholar] [CrossRef]
- Roest, H.I.J.; Ruuls, R.C.; Tilburg, J.J.H.C.; Nabuurs-Franssen, M.H.; Klaassen, C.H.W.; Vellema, P.; van den Brom, R.; Dercksen, D.; Wouda, W.; Spierenburg, M.A.H.; et al. Molecular epidemiology of Coxiella burnetii from ruminants in Q fever outbreak, The Netherlands. Emerg. Infect. Dis. 2011, 17, 668–675. [Google Scholar] [CrossRef]
- Tilburg, J.J.H.C.; Rossen, J.W.A.; van Hannen, E.J.; Melchers, W.J.G.; Hermans, M.A.H.; van de Bovenkamp, J.; Roest, H.I.J.; de Bruin, A.; Nabuurs-Franssen, M.H.; Horrevorts, A.M.; et al. Genotypic Diversity of Coxiella burnetii in the 2007-Q Fever Outbreak Episodes in The Netherlands. J. Clin. Microbiol. 2011, 50, 1076–1078. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tilburg, J.J.H.C.; Roest, H.I.J.; Buffet, S.; Nabuurs-Franssen, M.H.; Horrevorts, A.M.; Raoult, D.; Klaassen, C.H.W. Epidemic genotype of Coxiella burnetii among goats, sheep, and humans in The Netherlands. Emerg. Infect. Dis. 2012, 18, 887–889. [Google Scholar] [CrossRef]
- Hemsley, C.M.; Essex-Lopresti, A.; Norville, I.H.; Titball, R.W. Correlating Genotyping Data of Coxiella burnetii with Genomic Groups. Pathogens 2021, 10, 604. [Google Scholar] [CrossRef] [PubMed]
- Hemsley, C.M.; O’Neill, P.A.; Essex-Lopresti, A.; Norville, I.H.; Atkins, T.P.; Titball, R.W. Extensive genome analysis of Coxiella burnetii reveals limited evolution within genomic groups. BMC Genom. 2019, 20, 1–17. [Google Scholar] [CrossRef]
- Guatteo, R.; Beaudeau, F.; Berri, M.; Rodolakis, A.; Joly, A.; Seegers, H. Shedding routes of Coxiella burnetii in dairy cows: Implications for detection and control. Vet. Res. 2006, 37, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Guatteo, R.; Joly, A.; Beaudeau, F. Shedding and serological patterns of dairy cows following abortions associated with Coxiella burnetii DNA detection. Vet. Microbiol. 2012, 155, 430–433. [Google Scholar] [CrossRef] [PubMed]
- Rodolakis, A.; Berri, M.; Héchard, C.; Caudron, C.; Souriau, A.; Bodier, C.C.; Blanchard, B.; Camuset, P.; Devillechaise, P.; Natorp, J.C.; et al. Comparison of Coxiella burnetii shedding in milk of dairy bovine, caprine, and ovine herds. J. Dairy Sci. 2007, 90, 5352–5360. [Google Scholar] [CrossRef]
- Coxevac: EPAR Scientific Discussion. Available online: https://www.ema.europa.eu/medicines/veterinary/EPAR/coxevac (accessed on 30 September 2021).




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vellema, P.; Santman-Berends, I.; Dijkstra, F.; van Engelen, E.; Aalberts, M.; ter Bogt-Kappert, C.; van den Brom, R. Dairy Sheep Played a Minor Role in the 2005–2010 Human Q Fever Outbreak in The Netherlands Compared to Dairy Goats. Pathogens 2021, 10, 1579. https://doi.org/10.3390/pathogens10121579
Vellema P, Santman-Berends I, Dijkstra F, van Engelen E, Aalberts M, ter Bogt-Kappert C, van den Brom R. Dairy Sheep Played a Minor Role in the 2005–2010 Human Q Fever Outbreak in The Netherlands Compared to Dairy Goats. Pathogens. 2021; 10(12):1579. https://doi.org/10.3390/pathogens10121579
Chicago/Turabian StyleVellema, Piet, Inge Santman-Berends, Frederika Dijkstra, Erik van Engelen, Marian Aalberts, Carlijn ter Bogt-Kappert, and René van den Brom. 2021. "Dairy Sheep Played a Minor Role in the 2005–2010 Human Q Fever Outbreak in The Netherlands Compared to Dairy Goats" Pathogens 10, no. 12: 1579. https://doi.org/10.3390/pathogens10121579
APA StyleVellema, P., Santman-Berends, I., Dijkstra, F., van Engelen, E., Aalberts, M., ter Bogt-Kappert, C., & van den Brom, R. (2021). Dairy Sheep Played a Minor Role in the 2005–2010 Human Q Fever Outbreak in The Netherlands Compared to Dairy Goats. Pathogens, 10(12), 1579. https://doi.org/10.3390/pathogens10121579