The First Appearance of Symmetry in the Human Lineage: Where Perception Meets Art

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Symmetry Detection in Nonhuman Primates and Humans
3. Symmetry Detection, Acheulean Handaxes, and the Brain
4. Perceptual Fluency, Neural Synchrony, and Aesthetics
5. Implications of Symmetry for Understanding Visual Culture
6. Conclusions
References and Notes
- Lycett, S.J.; Gowlett, A.J. On questions surrounding the Acheulean “tradition”. World Archaeol. 2008, 40, 295–315. [Google Scholar] [CrossRef]
- Schmidt, R.R. The Dawn of the Human Min; Sidgwick & Jackson: London, UK, 1936. [Google Scholar]
- Schick, K.D.; Toth, N. Making Silent Stones Speak: Human Evolution and the Dawn of Human Technology; Weidenfeld and Nicolson: London, UK, 1993. [Google Scholar]
- Wynn, T. Archaeology and cognitive evolution. Behav. Brain Sci. 2002, 25, 389–438. [Google Scholar] [CrossRef] [PubMed]
- Wynn, T. The Large Cutting Tools from the South African Acheulean and the Question of Social Traditions. Curr. Anthropol. 2004, 45, 672–673. [Google Scholar]
- Nowell, A. Coincidental factors of handaxe morphology. Behav. Brain Sci. 2002, 25, 413–414. [Google Scholar] [CrossRef]
- Toth, N. The Prehistoric Roots of a Human Concept of Symmetry. Symmetry Cult. Sci. 1990, 1, 257–281. [Google Scholar]
- Rensch, B. Die Wirksankeit äesthetischer Faktoren bei Wirbeltieren. Z. Tierpsychol 1958, 15, 447–461. [Google Scholar] [CrossRef]
- Baylis, G.C.; Driver, J. Perception of symmetry and repetition within and across visual shapes: Part-descriptions and object-based-attention. Vis. Cogn. 2001, 8, 163–196. [Google Scholar] [CrossRef]
- Dragoi, V.; Turcu, C.M.; Sur, M. Stability of cortical responses and the statistics of natural scenes. Neuron 2001, 32, 1181–1192. [Google Scholar] [CrossRef]
- van der Helm, P.A.; Leeuwenberg, E.L.J. Goodness nontransformational approach. Psychol. Rev. 1996, 103, 429–456. [Google Scholar] [CrossRef] [PubMed]
- van der Helm, P.A.; Leeuwenberg, E.L.J. Accessibility, a criterion for regularity and hierarchy in visual pattern codes. J. Math. Psychol. 1991, 35, 151–213. [Google Scholar] [CrossRef]
- van der Helm, P.A. Natural selection of visual symmetries. Behav. Brain Sci. 2002, 25, 422–423; 432–438. [Google Scholar] [CrossRef]
- Enquist, M.E.; Arak, A. Symmetry, beauty and evolution. Nature 1994, 372, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Giurfa, M.; Eichmann, B.; Menzel, R. Symmetry perception in an insect. Nature 1996, 382, 458–546. [Google Scholar] [CrossRef] [PubMed]
- Wagemans, J. Characteristics and models of human symmetry detection. Trends Cogn. Sci. 1997, 1, 346–352. [Google Scholar] [CrossRef]
- Tyler, C.W.; Baseler, H.A.; Kontsevich, L.L.; Likova, L.T.; Wade, A.R.; Wandell, B.A. Predominantly extra-retinotopic cortical response to pattern symmetry. NeuroImage 2005, 24, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Redies, C.; Hasenstein, J.; Denzler, J. Fractal-like image statistics in visual art: similarity to natural scenes. Spatial Vis. 2007, 21, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Kersten, D. Three-dimensional symmetric shapes are discriminated more efficiently than asymmetric ones. J. Opt. Soc. Am. 2003, 20, 1331–1340. [Google Scholar] [CrossRef]
- Vetter, T.; Poggio, T.; Bültoff, H.H. The importance of symmetry and virtual views in three-dimensional object recognition. Curr. Biol. 1994, 4, 18–23. [Google Scholar] [CrossRef]
- Tyler, C.W. The Perception of Symmetry and its Computational Analysis; Erlbaum: NJ, USA, 1996. [Google Scholar]
- Tyler, C.W.; Baseler, H.A.; Kontsevich, L.L.; Likova, L.T.; Wade, A.R.; Wandell, B.A. Predominantly extra-retinotopic cortical response to pattern symmetry. NeuroImage 2005, 24, 306–314. [Google Scholar] [CrossRef]
- Sasaki, Y.; Vanduffel, W.; Knutsen, T.; Tyler, C.; Tootell, R. Symmetry activates extrastriate visual cortex in human and nonhuman primates. PNAS 2005, 102, 3159–3163. [Google Scholar] [CrossRef]
- Sasaki, Y. Processing local signals into global patterns. Curr. Opin. Neurobiol. 2007, 17, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Tyler, C.W. Empirical aspects of symmetry perception. Spatial Vis. 1995, 9, 1–7. [Google Scholar] [CrossRef]
- Barlow, H.B.; Reeves, B.C. The versatility and absolute efficiency of detecting mirror symmetry in random dot displays. Vision Res. 1979, 19, 783–793. [Google Scholar] [CrossRef]
- Beck, D.M.; Pinsk, M.A.; Kastner, S. Symmetry perception in humans and macaques. Trends Cogn. Sci. 2005, 9, 405–406. [Google Scholar] [CrossRef] [PubMed]
- Julesz, B. Figure and ground perception in briefly presented isodipole textures. In Perceptual Organization; Kubouy, M., Pomerantz, J., Eds.; Erlbaum: NJ, USA, 1981; pp. 27–54. [Google Scholar]
- Rentschler, I.; Jüttner, M.; Unzicker, A.; Landis, T. Innate and learned components of human visual preference. Curr. Biol. 1999, 9, 665–671. [Google Scholar] [CrossRef]
- Locher, P.J.; Wagemans, J. Effects of element type and spatial grouping on symmetry detection. Perception 1993, 22, 565–587. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, G.K.; Humphrey, D.E. The role of structure in infant visual pattern perception. Can. J. Psychol. 1989, 43, 165–182. [Google Scholar] [CrossRef]
- Stucchi, N.; Graciô, V.; Toneatto, C.; Scocchi, L. The perceptual salience of symmetrical and asymmetrical sections of a line. Perception 2010, 39, 1026–1042. [Google Scholar] [CrossRef]
- Treder, M.S. Behind the Looking-Glass: A Review on Human Symmetry Perception. Symmetry 2010, 2, 1510–1543. [Google Scholar] [CrossRef]
- Goren-Inbar, N.; Sharon, G. Axe Age: Acheulian Tool-making from Quarry to Discard; Equinox: London, UK, 2006. [Google Scholar]
- McNabb, J.; Binyon, F.; Hazelwood, L. The Large Cutting Tools from the South African Acheulean and the Question of Social Traditions. Curr. Anthropol. 2004, 41, 653–677. [Google Scholar] [CrossRef]
- Brumm, A.; McLaren, A. Scraper reduction and “imposed form” at the Lower Palaeolithic site of High Lodge, England. J. Hum. Evol. 2010, 2, 185–204. [Google Scholar] [CrossRef] [PubMed]
- McPherron, S.P. Handaxes as a measure of the mental capabilities of early hominids. J. Archaeol. Sci. 2000, 27, 655–663. [Google Scholar] [CrossRef]
- White, M.J. On the significance of Acheulean bifaces variability in Southern Britain. Proc. Prehist. Soc. 1998, 64, 15–44. [Google Scholar] [CrossRef]
- Machin, A.J.; Hosfield, R.T.; Mithen, S.J. Why are some handaxes symmetrical? Testing the influence of handaxe morphology on effectiveness. J. Archaeol. Sci. 2007, 34, 883–893. [Google Scholar] [CrossRef]
- Le Tensorer, J.M. Les cultures acheuléennes et la question de l’émergence de la pensée symbolique chez Homo erectus à partir des données relatives à la forme symétrique et harmonique des bifaces. C. R. Palevol. 2006, 5, 135–172. [Google Scholar] [CrossRef]
- Wenban-Smith, F.F. Biface Typology and Lower Paleolithic Cultural Development: Ficrons, Cleavers and Two Giant Bifaces from Cuxton. Lithics 2004, 25, 11–21. [Google Scholar]
- Mallol, C. The Selection of Lithic Raw Material in the Lower and Middle Pleistocene Levels TD6 and TD10 of Gran Dolina (Sierra de Atapuerca, Burgos, Spain). J. Anthropol. Res. 1999, 55, 385–407. [Google Scholar] [CrossRef]
- Saragusti, I.; Karasik, A.; Sharon, I.; Smilansky, U. Symmetry and roughness of pre-historical tools: Archaeological evidence on the development of the human capability to produce symmetric and smooth-edged tools during the Lower Paleolithic. Presented at Kimmel Centre for archaeological studies, the Weizmann Institute of Science, Israel, 2004. [Google Scholar]
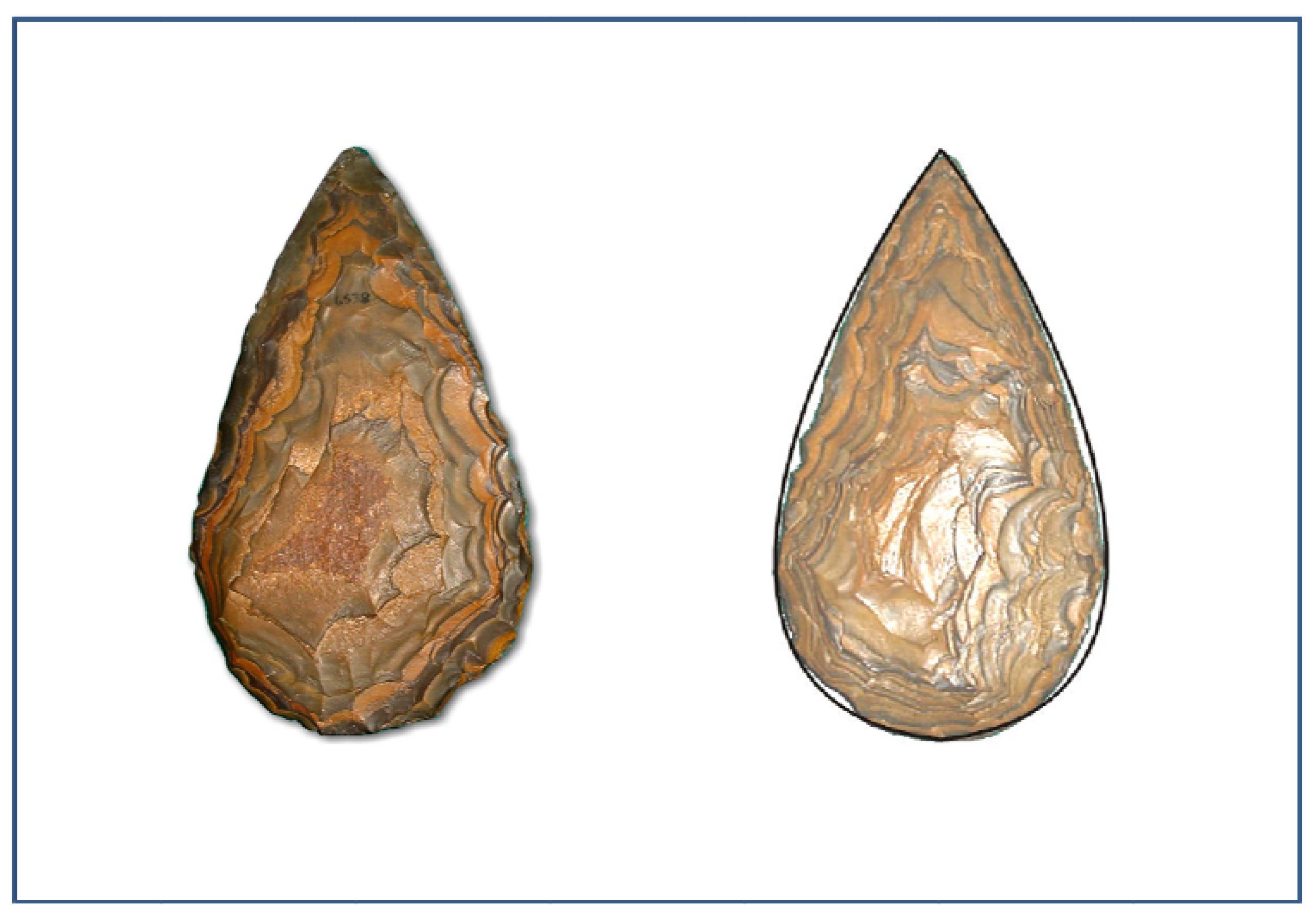
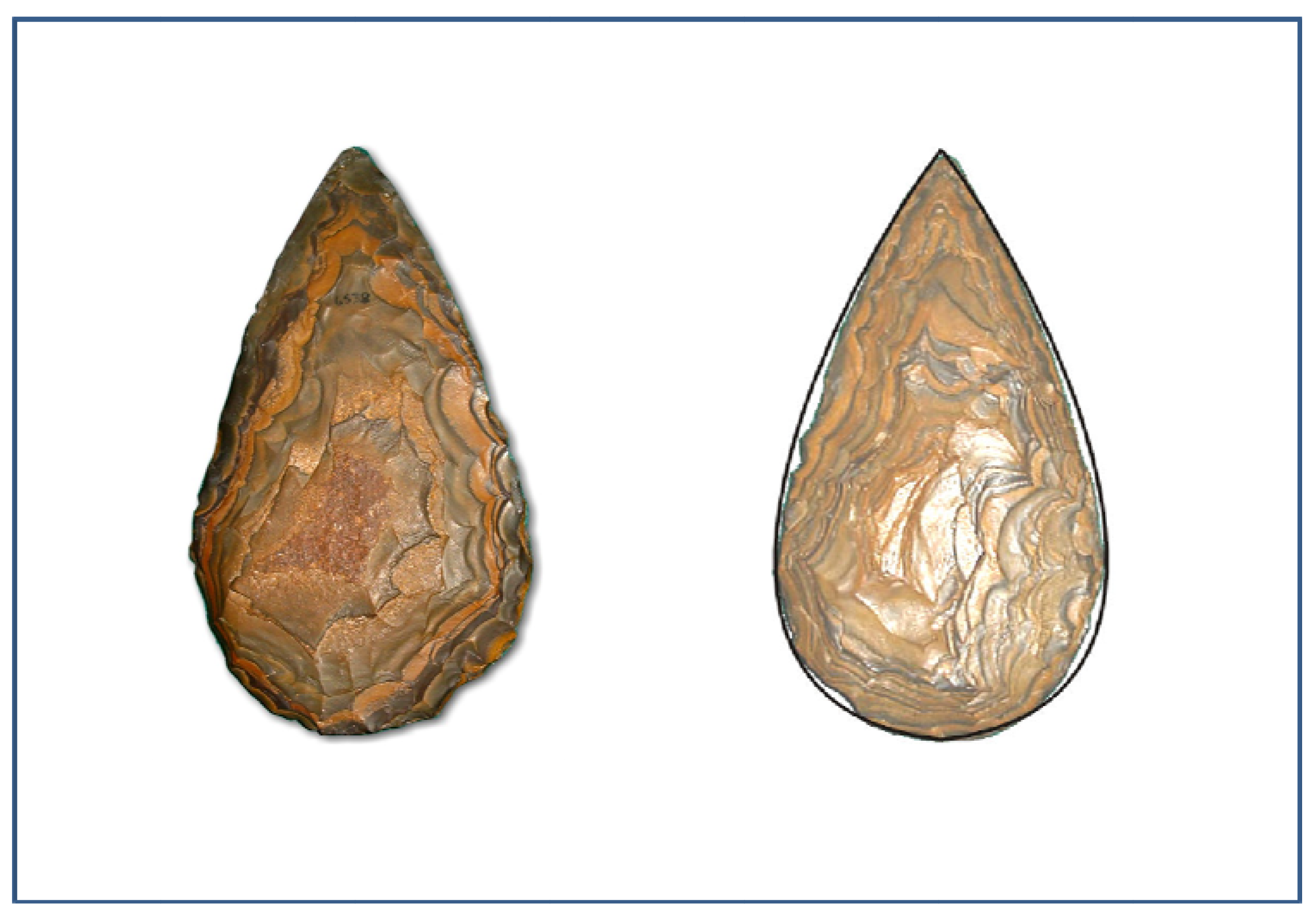
- Illustration from the McGregor Museum, Kimberley. Photo Michael Cope©
- Illustration from Marshall, G.D., Gamble, C.G., Roe, D.A., and Dupplaw, D. 2002. Acheulian biface database. Available at: http://ads.ahds.ac.uk/catalogue/specColl/bifaces/bf_query.cfm ADS, York (Accessed on 20 February 2011).
- Kohn, M.; Mithen, S. Handaxes: products of sexual selection? Antiquity 1999, 73, 518–526. [Google Scholar] [CrossRef]
- Machin, A.J. Why Bifaces Just Aren’t That Sexy: A Response to Kohn and Mithen (1999). Antiquity 2008, 82, 761–769. [Google Scholar] [CrossRef]
- 48 Nowell, A.; Chang, M.L. The case against sexual selection as an explanation of handaxe morphology. Paleoanthropology 2009, 77–78. Available at: http://www.paleoanthro.org/journal/contents_dynamic.asp?volume=2009 (Accessed on 3 November 2010).
- Hodgson, D. Symmetry and humans: Reply to Mithen’s “Sexy Handaxe Theory”. Antiquity 2009, 83, 195–198. [Google Scholar] [CrossRef]
- Hodgson, D. Another side of symmetry: the Acheulean biface debate. Antiquity 2010, 84, 325. [Google Scholar]
- Enquist, M.; Johnstone, R.A. Generalization and the evolution of symmetry preferences. Proc. R. Soc. Lond. B 1997, 264, 1345–1348. [Google Scholar] [CrossRef]
- Grill-Spector, K.; Kushnir, T.; Edelman, S.; Avidan, G.; Itzchak, Y.; Malach, R. Differential processing of objects under various viewing conditions in the human lateral occipital complex. Neuron 1999, 24, 187–203. [Google Scholar] [CrossRef]
- Denys, K.; Vanduffel, W.; Fize, D.; Nelissen, K.; Peuskens, H.; Van Essen, D.; Orban, G.A. The Processing of Visual Shape in the Cerebral Cortex of Human and Nonhuman Primates: A Functional Magnetic Resonance Imaging Study. J. Neurosci. 2004, 24, 2551–2565. [Google Scholar] [CrossRef] [PubMed]
- Orban, G.A.; Van Essen, D.; Vanduffel, W. Comparative mapping of higher visual areas in monkeys and humans. Trends Cogn. Sci. 2004, 8, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Orban, G.A.; Sunaert, S.; Todd, J.T.; van Hecke, P.; Marchal, G. Human cortical regions involved in extracting depth from motion. Neuron 1999, 24, 929–940. [Google Scholar] [CrossRef]
- Orban, G.A.; Claeys, K.; Nelissen, K.; Smans, R.; Sunaert, S.; Todd, J.T. Mapping the parietal cortex of human and non-human primates. Neuropsychol 2006, 44, 2647–2667. [Google Scholar] [CrossRef]
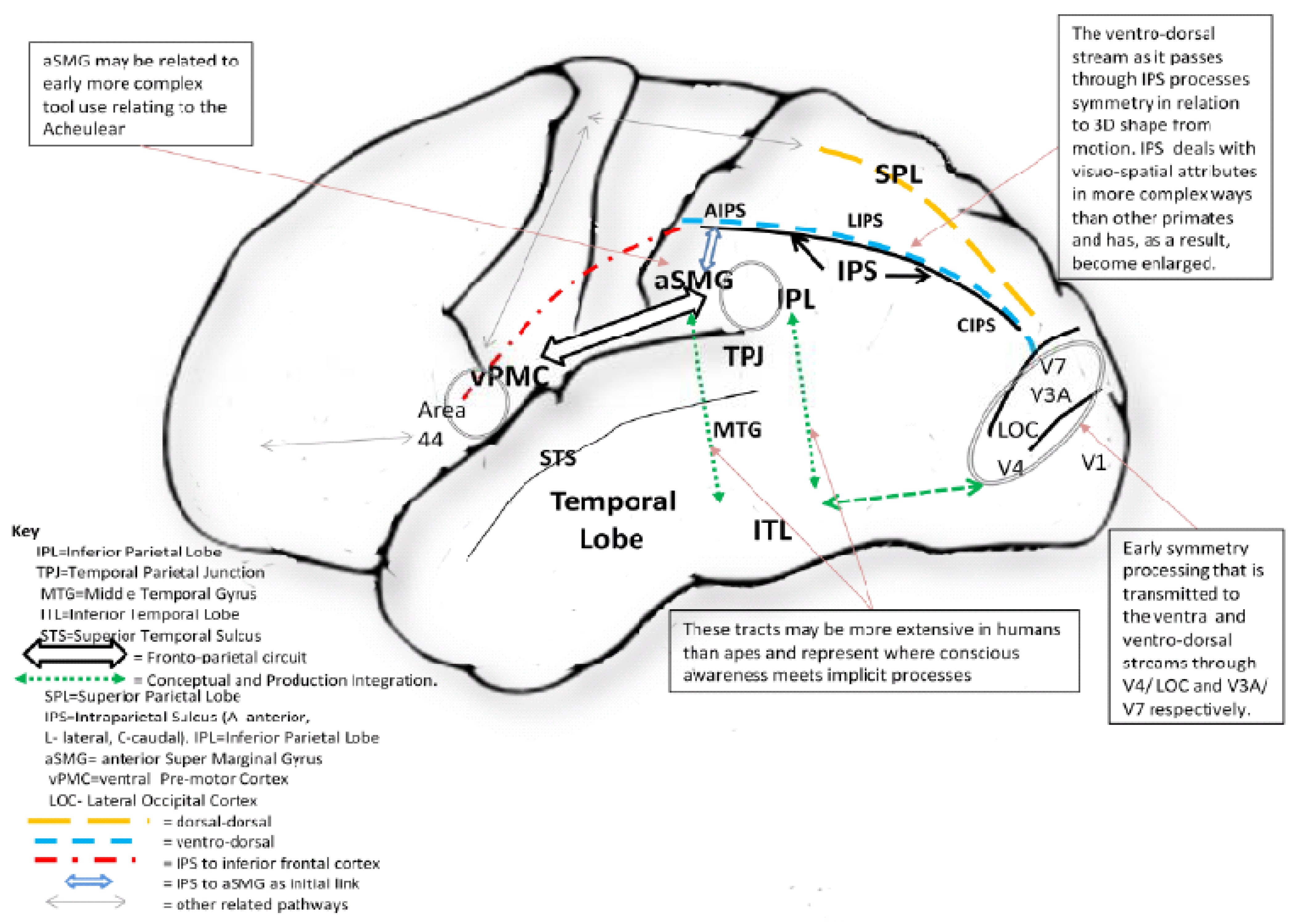
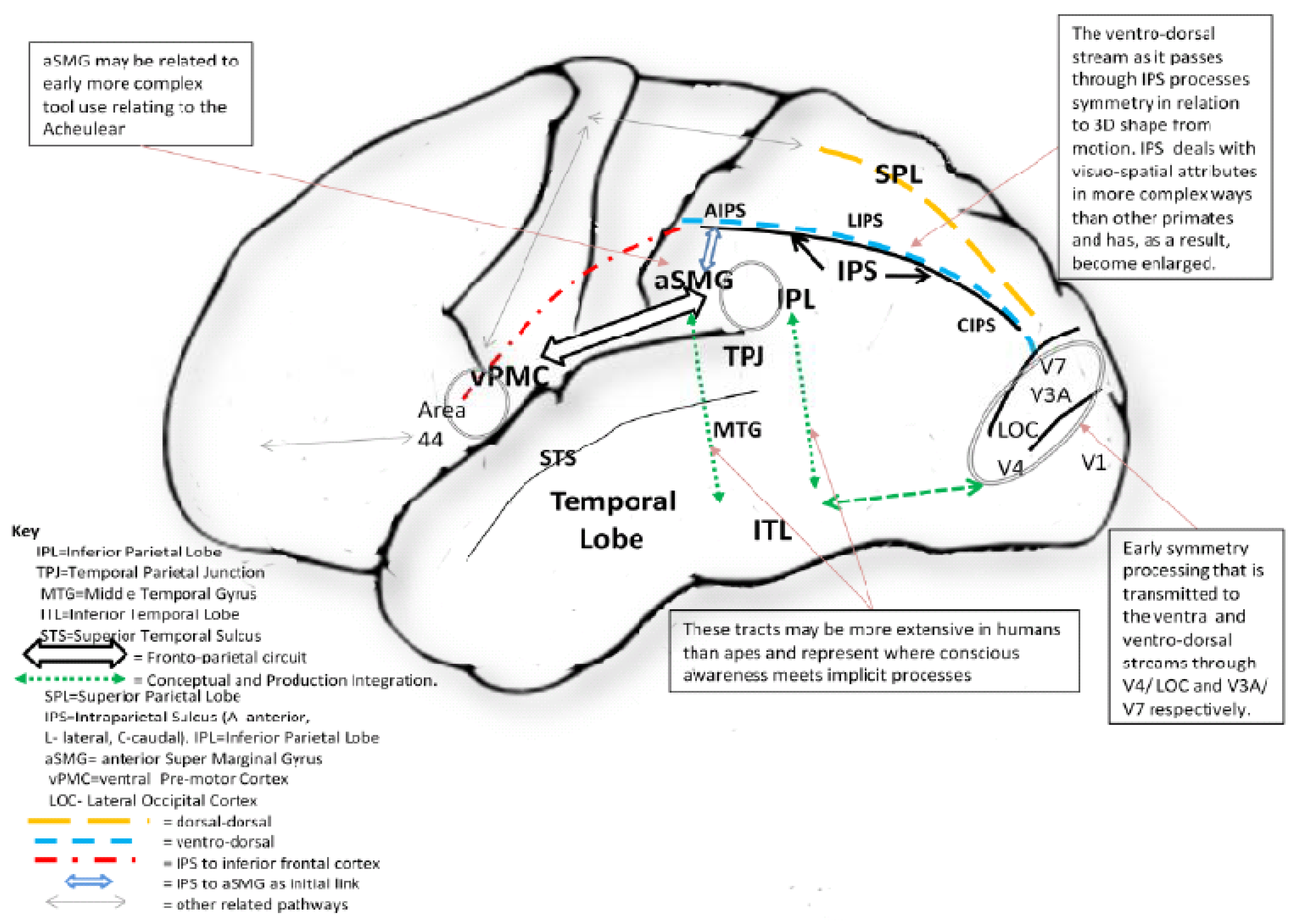
- Hodgson, D. Hominin Tool Production, Neural Integration, and the Social Brain. Hum. Origins 2011, in press. [Google Scholar]
- Vanduffel, W.; Fize, D.; Peuskens, H.; Denys, K.; Sunaert, S.; Todd, J.T.; Orban, G.A. Extracting 3D from motion: Differences in human and monkey intraparietal cortex. Science 2002, 298, 413–415. [Google Scholar] [CrossRef]
- Schendan, H.E.; Stern, C.E. Mental rotation and object categorization share a common network of prefrontal and dorsal and ventral regions of posterior cortex. NeuroImage 2007, 35, 1264–1277. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, T.; Schubotz, R.I.; Höfel, L.; van Cramon, D.Y. Brain correlates of aesthetic judgment of beauty. NeuroImage 2006, 29, 276–285. [Google Scholar] [CrossRef] [PubMed]
- Tzagarakis, C.; Trenton, A.; Jerde, T.A.; Lewis, S.M.; Uğurbil, K.; Georgopoulos, A.P. Cerebral cortical mechanisms of copying geometrical shapes: a multidimensional scaling analysis of fMRI patterns of activation. Exp. Brain Res. 2009, 194, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Nakahara, K.; Adachi, Y.; Osada, T.; Miyashita, Y. Exploring the neural basis of cognition: multi-modal links between human fMRI and macaque neurophysiology. Trends Cogn. Sci. 2007, 11, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Stout, D.; Chaminade, T. The evolutionary neuroscience of toolmaking. Neuropsychologia 2007, 45, 1091–1100. [Google Scholar] [CrossRef] [PubMed]
- Peeters, R.; Simone, L.; Nelissen, K.; Fabbri-Destro, M.; Vanduffel, W.; Rizzolatti, G.; Orban, G.A. The Representation of Tool Use in Humans and Monkeys: Common and Uniquely Human Features. J. Neurosci. 2009, 29, 11523–11539. [Google Scholar] [CrossRef] [PubMed]
- Gallese, V. The Inner Sense of Action. J. Conscious. Stud. 2000, 10, 23–40. [Google Scholar]
- Johnson-Frey, S.H.; Newman-Norland, R.; Grafton, S.T. A distributed Left Hemisphere Network Active During Planning of Everyday Tool Use Skills. Cereb. Cortex 2005, 15, 681–695. [Google Scholar] [CrossRef] [PubMed]
- Frey, S.H. Tool use, communicative gesture and cerebral asymmetries in the modern human brain. Phil. Trans. R. Soc. Lond. B. Biol Sci. 2008, 363, 1951–1957. [Google Scholar] [CrossRef] [PubMed]
- Ramayya, A.G.; Glasser, M.F.; Rilling, J.K. A DTI Investigation of Neural Substrates Supporting Tool Use. Cereb. Cortex 2010, 20, 507–516. [Google Scholar] [CrossRef]
- Read, D.; van der Leeuw, S. Biology is only part of the story. Philos. T. Roy. Soc. B 2008, 363, 1959–1968. [Google Scholar] [CrossRef] [PubMed]
- Keller, O. Aux Origines de la Géométrie—Le Paléolithique; Vuibert: Paris, France, 2004. [Google Scholar]
- Reber, R.; Winkielman, P.; Schwarz, N. Effects of perceptual fluency on affective judgements. Psychol. Sci. 1998, 9, 45–48. [Google Scholar] [CrossRef]
- Reber, R.; Schwarz, N.; Winkielman, P. Processing Fluency and Aesthetic Pleasure: Is Beauty in the Perceiver’s Processing Experience? Pers. Soc. Psychol. Rev. 2004, 8, 364. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, D. Evolution of the visual cortex and the emergence of symmetry in the Acheulean techno-complex. C. R. Palevol. 2009, 8, 93–97. [Google Scholar] [CrossRef]
- Verpooten, J.; Nelissen, M. Sensory exploitation and cultural transmission: the late emergence of iconic representations in human evolution. Theory Biosci. 2010, 129, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Grill-Spector, K.; Henson, R.; Martin, A. Repetition and the brain: neural models of stimulus-specific effects. Trends Cogn. Sci. 2006, 10, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Supér, H.; van der Togt, C.; Spekreijse, H.; Lamme, V.A.F. Internal State of Monkey Primary Visual Cortex (V1) Predicts Figure–Ground Perception. J. Neurosci. 2003, 23, 3407–3414. [Google Scholar] [CrossRef]
- Posner, M.I.; Keele, S.W. On the genesis of abstract ideas. J. Exp. Psychol. 1968, 77, 353–363. [Google Scholar] [CrossRef]
- Zajonc, R.B. Attitudinal effects of mere exposure. J. Pers. Soc. Psychol. 1968, 9, 1–27. [Google Scholar] [CrossRef]
- Zajonc, R.B. Emotions. In The Handbook of Social Psychology; Gilbert, D.T., Fiske, S.T., Lindzey, G., Eds.; McGraw-Hill: Boston, MA, USA, 1998; pp. 591–632. [Google Scholar]
- Tinio, P.P.L.; Leder, H. Just how stable are stable aesthetic features? Symmetry, complexity, and the jaws of massive familiarization. Acta Psychologica. 2009, 130, 241–250. [Google Scholar] [CrossRef]
- Zaidel, D.W.; Hessamian, M. Asymmetry and Symmetry in the Beauty of Human Faces. Symmetry 2010, 2, 136–149. [Google Scholar] [CrossRef]
- Fechner, G.T. Vorschule der Äesthtik; Breitkopf & Härtel: Leipzig, Germany, 1876. [Google Scholar]
- Berlyne, D.E. Conflict, Arousal, and Curiosity; McGraw-Hill: New York, NY, USA, 1960. [Google Scholar]
- Abraham, F.D.; Sprott, J.C.; Mitina, O.; Osorio, M.; Dequito, E.A.; Pinili, J.M. Judgments of Time, Aesthetics, and Complexity as a Function of The Fractal Dimension of Images formed by Chaotic Attractors. Available at: http://www.blueberry-brain.org/silliman/JEM%20ms4.htm (Accessed on December 2010).
- Jacobsen, T.; Höfel, L. Descriptive and evaluative judgment processes: Behavioral and electrophysiological indices of processing symmetry and aesthetics. Cogn. Affect Behav. Neurosci. 2003, 3, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Redies, C. A universal model of esthetic perception based on the sensory coding of natural stimuli. Spatial Vis. 2007, 21, 97–117. [Google Scholar] [CrossRef] [PubMed]
- Halberstadt, J. The generality and ultimate origins of the attractiveness of prototypes. Pers. Soc. Psychol. Rev. 2006, 10, 166–183. [Google Scholar] [CrossRef] [PubMed]
- Machin, A. The role of the individual agent in Acheulean biface variability—a multi-factorial model. J. Soc. Archaeol. 2009, 9, 35–58. [Google Scholar] [CrossRef]
- McNabb, J. The visual display hypothesis. Hum. Origins 2011, in press. [Google Scholar]
- Pope, M.; Russel, K.; Watson, K. Biface form and structured behaviour in the Acheulean. Lithic Studies Soc. 2006, 27, 44–57. [Google Scholar]
- Pope, M.I.; Roberts, M.B. Observations on the relationship between individuals and artefact scatters at the Middle Palaeolithic site of Boxgrove, West Sussex. In The Hominid Individual in Context: Archaeological Investigations of Lower & Middle Palaeolithic Landscapes, Locales & Artefacts; Gamble, C., Porr, M., Eds.; Routledge: London, UK, 2005; pp. 81–97. [Google Scholar]
- Höfel, L.; Jacobsen, T. Electrophysiological Indices of Processing Symmetry and Aesthetics: A Result of Judgment Categorization or Judgment Report? J. Psychophysiol. 2007, 21, 9–21. [Google Scholar] [CrossRef]
- Wymer, J. Lower Palaeolithic archaeology in Britain; John Baker: London, UK, 1968. [Google Scholar]
- Wymer, J. The Palaeolithic Age; Croom Helm: London, UK, 1983. [Google Scholar]
- Roe, D. The Lower and Middle Palaeolithic periods in Britain; Routledge & Kegan Paul: London, UK, 1981. [Google Scholar]
- Photo, J. Wymer© 1982. Plate XIII. The palaeolithic age. New York: St. Martins (with permission of the Natural History Museum, London).
- Radomsky, A.S.; Rachman, S. Symmetry, ordering and arranging compulsive behaviour. Behav. Res. Therapy 2004, 42, 893–913. [Google Scholar] [CrossRef]
- Dulaney, S.; Fiske, A.P. Cultural rituals and obsessive-compulsive disorder: Is there a common psychological mechanism? Ethos 1994, 22, 243–283. [Google Scholar] [CrossRef]
- Fiske, A.P.; Haslam, N. Is obsessive-compulsive disorder of the human disposition to perform socially meaningful rituals? Evidence of similar content. J. Nerv. Ment. Dis. 1997, 185, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, D. Art, Perception and Information Processing: An evolutionary Perspective. Rock Art Res. 2000, 17, 3–34. [Google Scholar]
- From, Marshall, G.D., Gamble, C.G., Roe, D.A. and Dupplaw, D. 2002. Acheulian biface database. Available at: http://ads.ahds.ac.uk/catalogue/specColl/bifaces/bf_query.cfm, ADS, York.
- Batchelor, R.K. Evolution, Artefacts, Meaning and Design: The extent to which evolutionary theory can explain how and why humans attribute significance and meaning to the material world and the consequences of this for understanding design. Doctoral Dissertation, London, UK, 2004. Available at: http://dspace.bucks.ac.uk/dspace/bitstream/10239/127/1/01Prelims.pdf (Accessed on December 2010).
- Dissanayake, E. Homo Aestheticus: Where Art Comes From and Why; Free Press: New York, NY, USA, 1992. [Google Scholar]
- Dissanyake, E. The Deep Structure of Pleistocene Rock Art: The “Artification Hypothesis” IFRAO Congress. Pleistocene art of the world—Symposium: Signs, symbols, myth, ideology (Pre-Acts). 2010. Available at: http://www.ifraoariege2010.fr/docs/Articles/Dissanayake-Signes.pdf (Accessed on 1 November 2010).
- Oakley, K. Skill as a human possession. In Perspectives on Human Evolution; Washburn, S.L., Dolhinow, P., Eds.; Holt, Rinehart, and Winston: New York, NY, USA, 1972; pp. 14–50. [Google Scholar]
- Edwards, S.W. Acheulean Evidence. Rock Art Res. 2003, 20, 109–111. [Google Scholar]



© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hodgson, D. The First Appearance of Symmetry in the Human Lineage: Where Perception Meets Art. Symmetry 2011, 3, 37-53. https://doi.org/10.3390/sym3010037
Hodgson D. The First Appearance of Symmetry in the Human Lineage: Where Perception Meets Art. Symmetry. 2011; 3(1):37-53. https://doi.org/10.3390/sym3010037
Chicago/Turabian StyleHodgson, Derek. 2011. "The First Appearance of Symmetry in the Human Lineage: Where Perception Meets Art" Symmetry 3, no. 1: 37-53. https://doi.org/10.3390/sym3010037
APA StyleHodgson, D. (2011). The First Appearance of Symmetry in the Human Lineage: Where Perception Meets Art. Symmetry, 3(1), 37-53. https://doi.org/10.3390/sym3010037