
Controlling Factors of Spatiotemporal Variations in Transpiration on a Larch Plantation Hillslope in Northwest China

 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Meteorological and Soil Moisture Measurements
2.3. Sap Flow Measurements and Transpiration Estimates
2.4. Data Analysis
3. Results
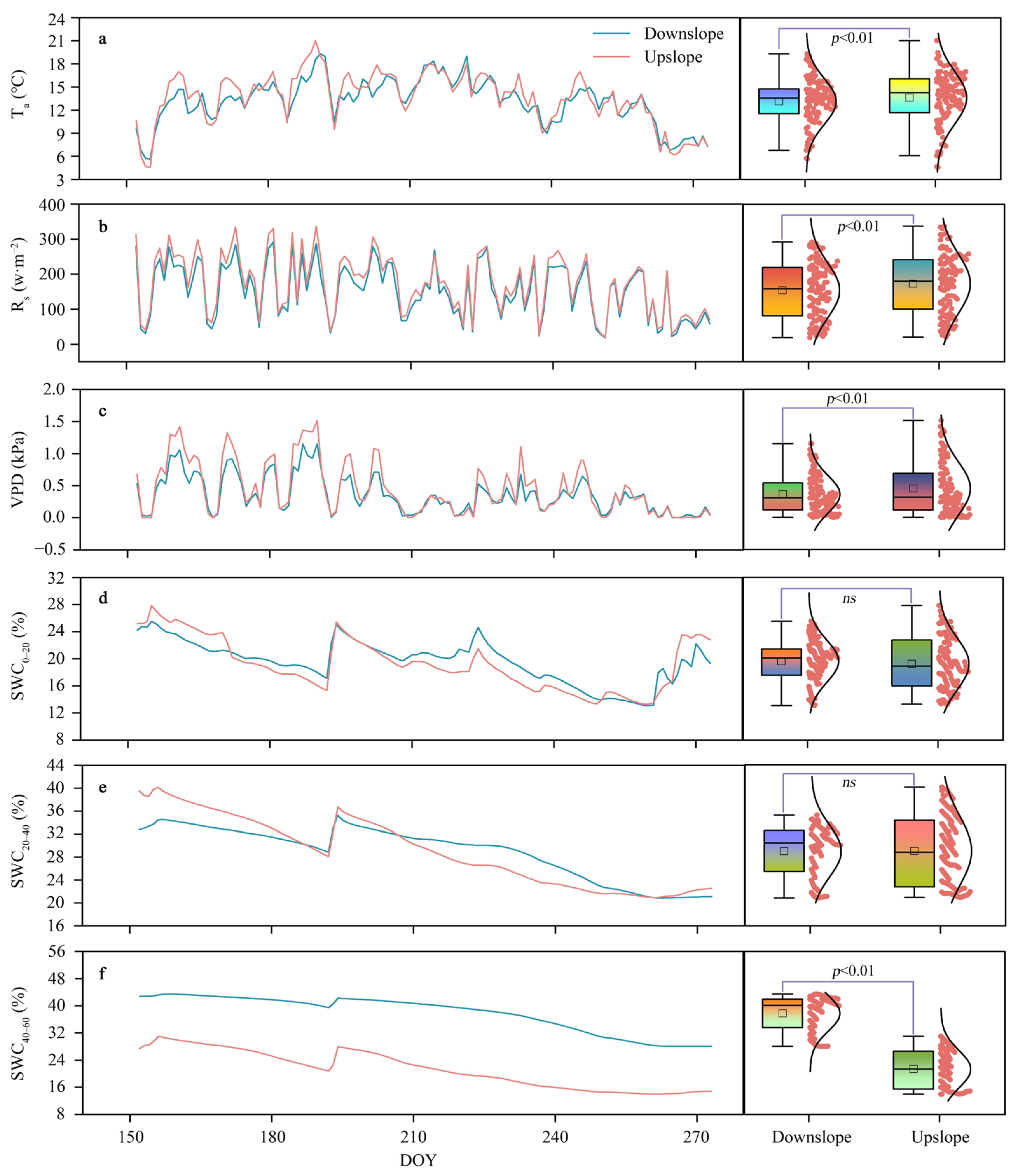
3.1. Variations in the Meteorological Conditions and Soil Moisture at the Two Slope Positions
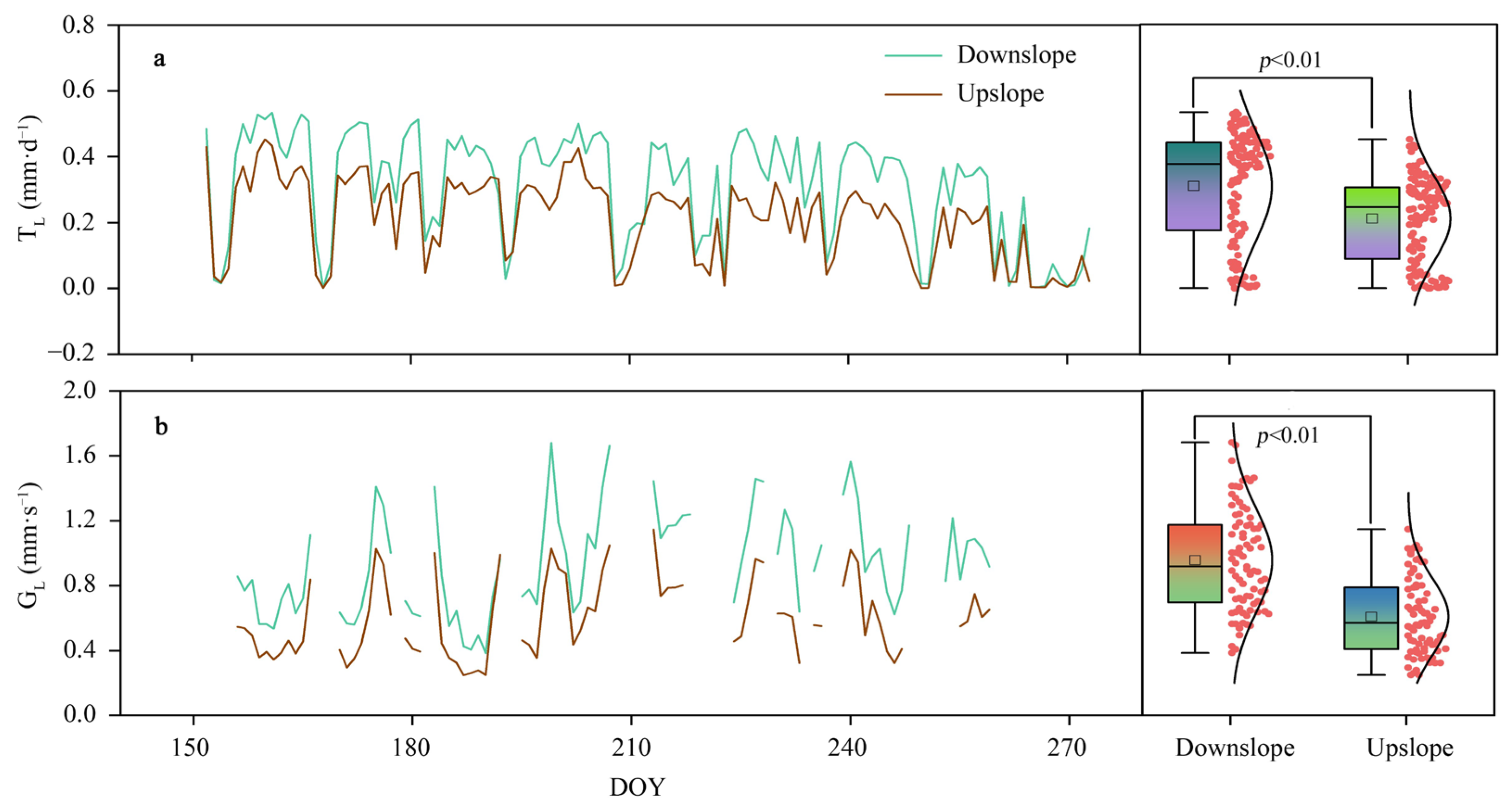
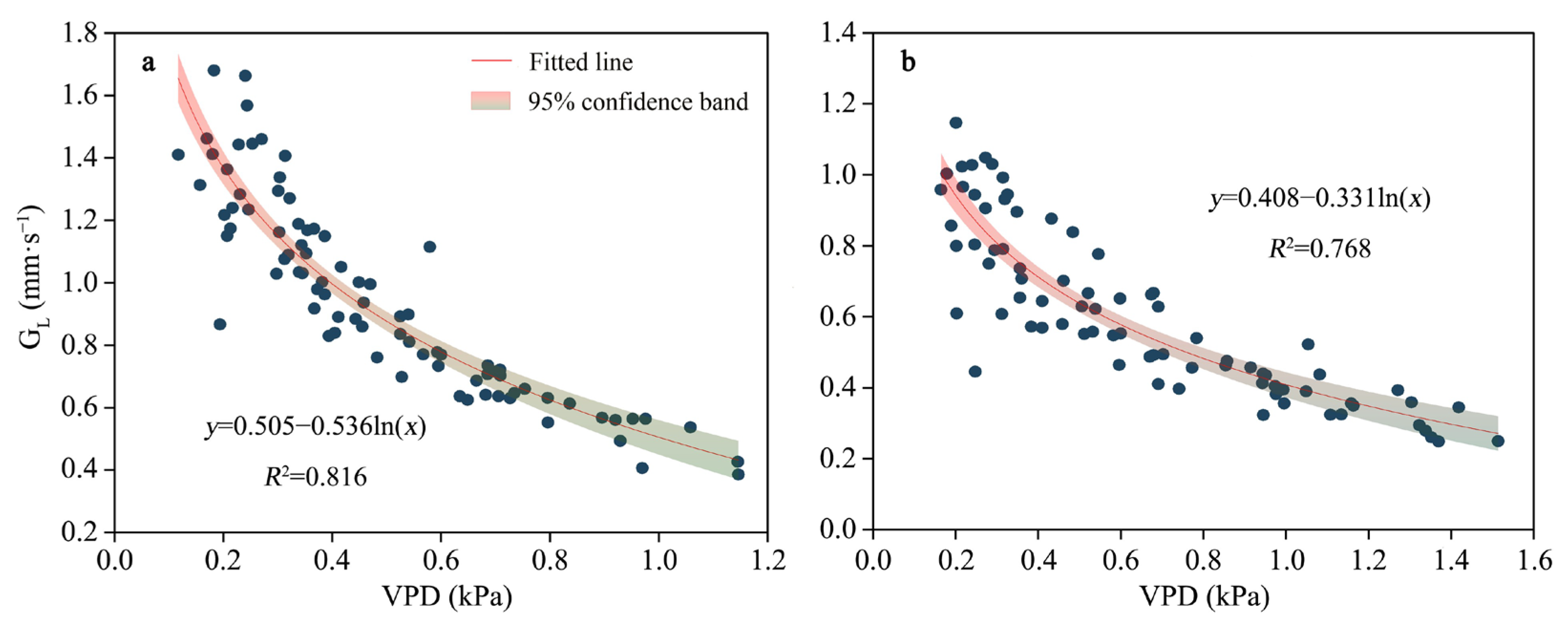
3.2. TL Variations and the Main Controlling Factors Affecting Slope Differences
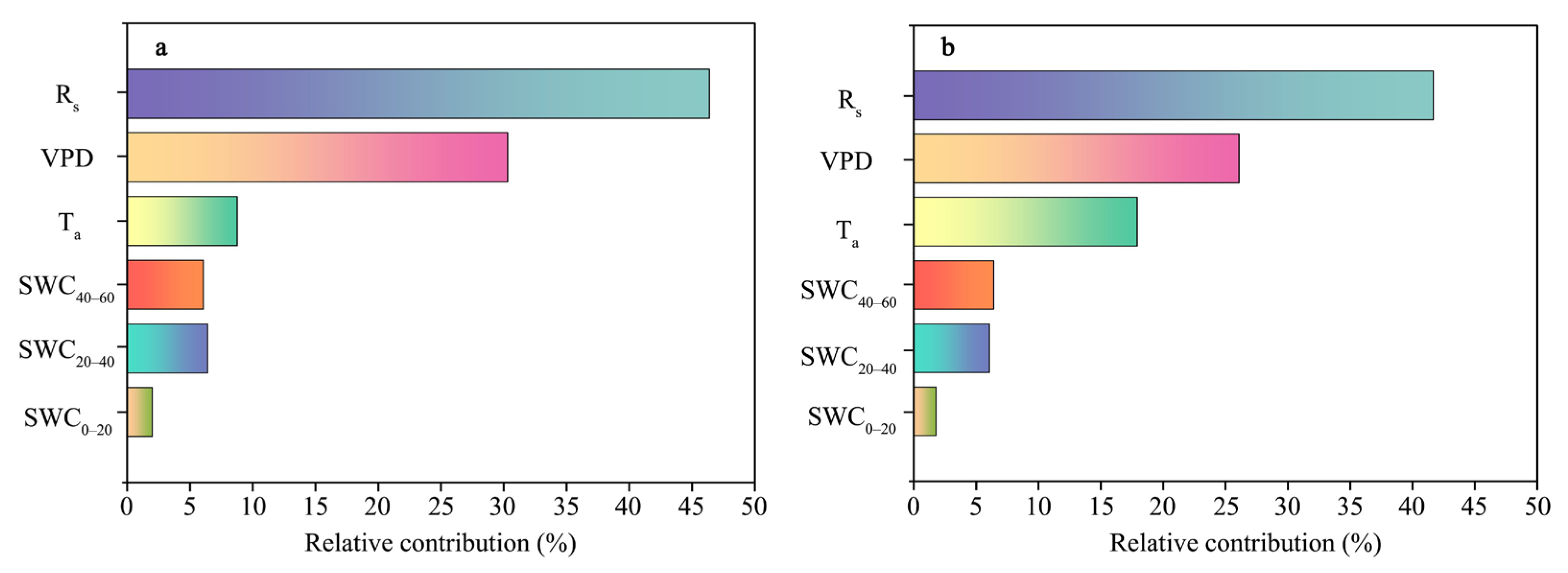
3.3. Impact and Contribution of the Dominant Factors to the Temporal Variations in the TL at the Two Positions
4. Discussion
4.1. Controls of Slope Variations in Canopy Transpiration
4.2. Controls of the Temporal Dynamics of Canopy Transpiration
4.3. Implications for Ecohydrological Studies and Forest Management
4.4. Limitations of This Study and Indications for Future Studies
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| TL | Transpiration per unit leaf area |
| GL | Canopy conductance per unit leaf area |
| VPD | Vapor pressure deficit |
| SWC | Soil water content |
| Ta | Air temperature |
| Rs | Solar radiation |
| LAI | Leaf area index |
| DOY | Day of the year |
References
- Wang, Y.H.; Yu, P.T.; Feger, K.H.; Wei, X.H.; Sun, G.; Bonell, M.; Xiong, W.; Zhang, S.L.; Xu, L.H. Annual runoff and evapotranspiration of forestlands and nonforestlands in selected basins of the Loess Plateau of China. Ecohydrology 2011, 4, 277–287. [Google Scholar] [CrossRef]
- Schwärzel, K.; Zhang, L.; Montanarella, L.; Wang, Y.; Sun, G. How afforestation affects the water cycle in drylands: A process-based comparative analysis. Glob. Change Biol. 2020, 26, 944–959. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.M.; Fu, B.J.; Piao, S.L.; Wang, S.; Ciais, P.; Zeng, Z.Z.; Lü, Y.H.; Zeng, Y.; Jiang, X.H.; Wu, B.F. Revegetation in China’s Loess Plateau is approaching sustainable water resource limits. Nat. Clim. Change 2016, 6, 1019–1022. [Google Scholar] [CrossRef]
- Liu, Z.B.; Yu, S.P.; Xu, L.H.; Wang, Y.H.; Yu, P.T.; Chao, Y. Differentiated responses of daytime and nighttime sap flow to soil water deficit in a larch plantation in Northwest China. Agric. Water Manag. 2023, 289, 108540. [Google Scholar] [CrossRef]
- Wang, L.; Liu, Z.B.; Guo, J.B.; Wang, Y.H.; Ma, J.; Yu, S.P.; Yu, P.T.; Xu, L.H. Estimate canopy transpiration in larch plantations via the interactions among reference evapotranspiration, leaf area index, and soil moisture. For. Ecol. Manag. 2021, 481, 118749. [Google Scholar] [CrossRef]
- Méndez-Toribio, M.; Ibarra-Manríquez, G.; Navarrete-Segueda, A.; Paz, H. Topographic position, but not slope aspect, drives the dominance of functional strategies of tropical dry forest trees. Environ. Res. Lett. 2017, 12, 085002. [Google Scholar] [CrossRef]
- Metzen, D.; Sheridan, G.J.; Benyon, R.G.; Bolstad, P.V.; Griebel, A.; Lane, P.N. Spatio-temporal transpiration patterns reflect vegetation structure in complex upland terrain. Sci. Total Environ. 2019, 694, 133551. [Google Scholar] [CrossRef]
- Tsuruta, K.; Yamamoto, H.; Kosugi, Y.; Makita, N.; Katsuyama, M.; Kosugi, K.I.; Tani, M. Slope position and water use by trees in a headwater catchment dominated by Japanese cypress: Implications for catchment-scale transpiration estimates. Ecohydrology 2020, 13, e2245. [Google Scholar] [CrossRef]
- Kumagai, T.O.; Tateishi, M.; Shimizu, T.; Otsuki, K. Transpiration and canopy conductance at two slope positions in a Japanese cedar forest watershed. Agric. For. Meteorol. 2008, 148, 1444–1455. [Google Scholar] [CrossRef]
- Song, L.N.; Zhu, J.J.; Li, X.J.Y.; Wang, K.; Wang, G.C.; Sun, H.H. Transpiration of Pinus sylvestris var. mongolica trees at different positions of sand dunes in a semiarid sandy region of Northeast China. Trees 2022, 36, 749–762. [Google Scholar] [CrossRef]
- Li, Z.H.; Yu, P.T.; Wang, Y.H.; Webb, A.A.; He, C.; Wang, Y.B.; Yang, L.L. A model coupling the effects of soil moisture and potential evaporation on the tree transpiration of a semi-arid larch plantation. Ecohydrology 2017, 10, e1764. [Google Scholar] [CrossRef]
- Hayat, M.; Zha, T.S.; Jia, X.; Iqbal, S.; Qian, D.; Bourque, C.P.A.; Khan, A.; Tian, Y.; Bai, Y.J.; Liu, P.; et al. A multiple-temporal scale analysis of biophysical control of sap flow in Salix psammophila growing in a semiarid shrubland ecosystem of northwest China. Agric. For. Meteorol. 2020, 288, 107985. [Google Scholar] [CrossRef]
- Oogathoo, S.; Houle, D.; Duchesne, L.; Kneeshaw, D. Vapour pressure deficit and solar radiation are the major drivers of transpiration of balsam fir and black spruce tree species in humid boreal regions, even during a short-term drought. Agric. For. Meteorol. 2020, 291, 108063. [Google Scholar] [CrossRef]
- Ghimire, C.P.; van Meerveld, H.I.; Zwartendijk, B.W.; Bruijnzeel, L.A.; Ravelona, M.; Lahitiana, J.; Lubczynski, M.W. Vapour pressure deficit and solar radiation are the major drivers of transpiration in montane tropical secondary forests in eastern Madagascar. Agric. For. Meteorol. 2022, 326, 109159. [Google Scholar] [CrossRef]
- Song, X.W.; Lyu, S.D.; Wen, X.F. Limitation of soil moisture on the response of transpiration to vapor pressure deficit in a subtropical coniferous plantation subjected to seasonal drought. J. Hydrol. 2020, 591, 125301. [Google Scholar] [CrossRef]
- Yan, X.Y.; Zhang, Z.D.; Zhao, X.F.; Huang, M.B.; Wu, X.F.; Guo, T.Q. Differentiated responses of plant water use regulation to drought in Robinia pseudoacacia plantations on the Chinese Loess Plateau. Agric. Water Manag. 2024, 291, 108659. [Google Scholar] [CrossRef]
- Chen, S.N.; Zhang, Z.Q.; Chen, Z.S.N.; Xu, H.; Li, J.L. Responses of canopy transpiration and conductance to different drought levels in Mongolian pine plantations in a semiarid urban environment of China. Agric. For. Meteorol. 2024, 347, 109897. [Google Scholar] [CrossRef]
- Whitley, R.; Zeppel, M.; Armstrong, N.; Macinnis-Ng, C.; Yunusa, I.; Eamus, D. A modified Jarvis-Stewart model for predicting stand-scale transpiration of an Australian native forest. Plant Soil 2008, 305, 35–47. [Google Scholar] [CrossRef]
- Renner, M.; Hassler, S.K.; Blume, T.; Weiler, M.; Hildebrandt, A.; Guderle, M.; Schymanski, S.J.; Kleidon, A. Dominant controls of transpiration along a hillslope transect inferred from ecohydrological measurements and thermodynamic limits. Hydrol. Earth Syst. Sci. 2016, 20, 2063–2083. [Google Scholar] [CrossRef]
- Gwak, Y.; Kim, S. Factors affecting soil moisture spatial variability for a humid forest hillslope. Hydrol. Process. 2017, 31, 431–445. [Google Scholar] [CrossRef]
- Petit, G.; von Arx, G.; Kiorapostolou, N.; Lechthaler, S.; Prendin, A.L.; Anfodillo, T.; Caldeira, M.C.; Cochard, H.; Copini, P.; Crivellaro, A.; et al. Tree differences in primary and secondary growth drive convergent scaling in leaf area to sapwood area across Europe. New Phytol. 2018, 218, 1383–1392. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.N.; Nie, Y.P.; Luo, Z.D.; Wang, Z.Y.; Huang, L.; He, F.; Chen, H.S. Transpiration rates decline under limited moisture supply along hillslopes in a humid karst terrain. Sci. Total Environ. 2023, 894, 164977. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.J.; Benyon, R.G.; Lane, P.N. Responses of evapotranspiration at different topographic positions and catchment water balance following a pronounced drought in a mixed species eucalypt forest, Australia. J. Hydrol. 2012, 440, 62–74. [Google Scholar] [CrossRef]
- Wang, X.M.; Zhang, C.X.; Hasi, E.; Dong, Z.B. Has the Three Norths Forest Shelterbelt Program solved the desertification and dust storm problems in arid and semiarid China? J. Arid Environ. 2010, 74, 13–22. [Google Scholar] [CrossRef]
- Tian, A.; Wang, Y.H.; Webb, A.A.; Liu, Z.B.; Yu, P.T.; Xiong, W.; Wang, X. Partitioning the causes of spatiotemporal variation in the sunny day sap flux density of a larch plantation on a hillslope in northwest China. J. Hydrol. 2019, 571, 503–515. [Google Scholar] [CrossRef]
- Li, J.M.; Yu, P.T.; Wan, Y.F.; Wang, Y.H.; Liu, B.B.; Yu, Y.P. Effects of topography and social position on the solar radiation of individual trees on a hillslope in Northwest China. Forests 2023, 14, 561. [Google Scholar] [CrossRef]
- Granier, A. Evaluation of transpiration in a Douglas-Fir stand by means of sap flow measurements. Tree Physiol. 1987, 3, 309–320. [Google Scholar] [CrossRef]
- Liu, Z.B.; Yu, S.P.; Xu, L.H.; Wang, Y.H.; Wang, M.F.; Yu, P.T. Spatiotemporal variations in sap flow in a larch plantation: Sampling size for stand scale estimates. J. For. Res. 2024, 35, 139. [Google Scholar] [CrossRef]
- Naithani, K.J.; Ewers, B.E.; Pendall, E. Sap flux-scaled transpiration and stomatal conductance response to soil and atmospheric drought in a semi-arid sagebrush ecosystem. J. Hydrol. 2012, 464–465, 176–185. [Google Scholar] [CrossRef]
- Yu, S.P.; Liu, Z.B.; Guo, J.B.; Wang, Y.H.; Yu, P.T.; Wang, L. Stand transpiration characteristics of Larix principis-rupprechtii plantation and their influencing factors in Liupan Mountain. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2021, 45, 131–140. [Google Scholar] [CrossRef]
- Wilks, D.S. Statistical Methods in the Atmospheric Sciences, 2nd ed.; Elsevier: Burlington, MA, USA, 2006; pp. 141–143. [Google Scholar]
- Matsoukis, A.; Kamoutsis, A.; Chronopoulou-Sereli, A. A note on the flowering of Ajuga orientalis L. in relation to air temperature in Mount Aenos (Cephalonia, Greece). Curr. Agric. Res. J. 2018, 6, 261–267. [Google Scholar] [CrossRef]
- Kamoutsis, A.; Chronopoulos, K.; Matsoukis, A. Altitude and canopy cover effects on air temperature in a mountainous region of Ionian Islands, Greece. Curr. World Environ. 2018, 13, 292–298. [Google Scholar] [CrossRef]
- Oren, R.; Sperry, J.S.; Katul, G.G.; Pataki, D.E.; Ewers, B.E.; Phillips, N.; Schäfer, K.V.R. Survey and synthesis of intra-and interspecific variation in stomatal sensitivity to vapour pressure deficit. Plant Cell Environ. 1999, 22, 1515–1526. [Google Scholar] [CrossRef]
- Schmidt, U.; Thoni, H.; Kaupenjohann, M. Using a boundary line approach to analyze N2O flux data from agricultural soils. Nutr. Cycl. Agroecosyst. 2000, 57, 119–129. [Google Scholar] [CrossRef]
- Yu, S.P.; Guo, J.B.; Liu, Z.B.; Wang, Y.H.; Ma, J.; Li, J.M.; Liu, F. Assessing the impact of soil moisture on canopy transpiration using a modified Jarvis-Stewart model. Water 2021, 13, 2720. [Google Scholar] [CrossRef]
- R, Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 14 June 2024).
- Grömping, U. Relative importance for linear regression in R: The package relaimpo. J. Stat. Softw. 2007, 17, 1–27. [Google Scholar] [CrossRef]
- Fabiani, G.; Klaus, J.; Penna, D. The influence of hillslope topography on beech water use: A comparative study in two different climates. Hydrol. Earth Syst. Sci. 2024, 28, 2683–2703. [Google Scholar] [CrossRef]
- Kume, T.; Tsuruta, K.; Komatsu, H.; Shinohara, Y.; Katayama, A.; Ide, J.I.; Otsuki, K. Differences in sap flux-based stand transpiration between upper and lower slope positions in a Japanese cypress plantation watershed. Ecohydrology 2016, 9, 1105–1116. [Google Scholar] [CrossRef]
- Du, J.L.; Dai, X.Q.; Huo, Z.L.; Wang, X.W.; Wang, S.; Wang, C.Z.; Zhang, C.L.; Huang, G.H. Stand transpiration and canopy conductance dynamics of Populus popularis under varying water availability in an arid area. Sci. Total Environ. 2023, 892, 164397. [Google Scholar] [CrossRef]
- Novick, K.A.; Ficklin, D.L.; Stoy, P.C.; Williams, C.A.; Bohrer, G.; Oishi, A.C.; Papuga, S.A.; Blanken, P.D.; Noormets, A.; Sulman, B.N.; et al. The increasing importance of atmospheric demand for ecosystem water and carbon fluxes. Nat. Clim. Change 2016, 6, 1023–1027. [Google Scholar] [CrossRef]
- Wu, Z.; Behzad, H.M.; He, Q.F.; Wu, C.; Bai, Y.; Jiang, Y.J. Seasonal transpiration dynamics of evergreen Ligustrum lucidum linked with water source and water-use strategy in a limestone karst area, southwest China. J. Hydrol. 2021, 597, 126199. [Google Scholar] [CrossRef]
- Tromp-van Meerveld, H.J.; McDonnell, J.J. On the interrelationships between topography soil depth, soil moisture, transpiration rates and species distributions at the hillslope scale. Adv. Water Res. 2006, 29, 293–310. [Google Scholar] [CrossRef]
- Berry, Z.C.; Gotsch, S.G.; Holwerda, F.; Muñoz-Villers, L.E.; Asbjornsen, H. Slope position influences vegetation-atmosphere interactions in a tropical montane cloud forest. Agric. For. Meteorol. 2016, 221, 207–218. [Google Scholar] [CrossRef]
- She, D.L.; Xia, Y.Q.; Shao, M.G.; Peng, S.Z.; Yu, S.G. Transpiration and canopy conductance of Caragana korshinskii trees in response to soil moisture in sand land of China. Agroforest. Syst. 2013, 87, 667–678. [Google Scholar] [CrossRef]
- Chen, S.N.; Wei, W.; Huang, Y. Biophysical controls on canopy transpiration of Pinus tabulaeformis under different soil moisture conditions in the Loess Plateau of China. J. Hydrol. 2024, 631, 130799. [Google Scholar] [CrossRef]
- Song, L.N.; Zhu, J.J.; Li, M.C.; Zhang, J.X.; Zheng, X.; Wang, K. Canopy transpiration of Pinus sylvestris var. mongolica in a sparse wood grassland in the semiarid sandy region of Northeast China. Agric. For. Meteorol. 2018, 250, 192–201. [Google Scholar] [CrossRef]
- Flo, V.; Martínez-Vilalta, J.; Granda, V.; Mencuccini, M.; Poyatos, R. Vapour pressure deficit is the main driver of tree canopy conductance across biomes. Agric. For. Meteorol. 2022, 322, 109029. [Google Scholar] [CrossRef]
- Klein, T.; Ramon, U.; Cohen, S.; Yakir, D. Diurnal dynamics of water transport, storage and hydraulic conductivity in pine trees under seasonal drought. iForest 2021, 14, 71–79. [Google Scholar] [CrossRef]
- Wieser, G.; Leo, M.; Oberhuber, W. Transpiration and canopy conductance in an inner alpine Scots pine (Pinus sylvestris) forest. Flora 2014, 209, 491–498. [Google Scholar] [CrossRef]
- Wang, H.L.; Tetzlaff, D.; Dick, J.J.; Soulsby, C. Assessing the environmental controls on Scots pine transpiration and the implications for water partitioning in a boreal headwater catchment. Agric. For. Meteorol. 2017, 240, 58–66. [Google Scholar] [CrossRef]
- Lhomme, J.P.; Elguero, E.; Chehbouni, A.; Boulet, G. Stomatal control of transpiration: Examination of Monteith’s formulation of canopy resistance. Water Resour. Res. 1998, 34, 2301–2308. [Google Scholar] [CrossRef]
- Hayat, M.; Xiang, J.; Yan, C.H.; Xiong, B.W.; Wang, B.; Qin, L.J.; Saeed, S.; Hussain, A.; Zou, Z.D.; Qiu, G.Y. Environmental control on transpiration and its cooling effect of Ficus concinna in a subtropical city Shenzhen, southern China. Agric. For. Meteorol. 2022, 312, 108715. [Google Scholar] [CrossRef]
- Sun, S.J.; Meng, P.; Zhang, J.S.; Wan, X. Variation in soil water uptake and its effect on plant water status in Juglans regia L. during dry and wet seasons. Tree Physiol. 2011, 31, 1378–1389. [Google Scholar] [CrossRef]
- Gessler, A.; Bächli, L.; Rouholahnejad Freund, E.; Treydte, K.; Schaub, M.; Haeni, M.; Weiler, M.; Seeger, S.; Marshall, J.; Christian, H.; et al. Drought reduces water uptake in beech from the drying topsoil, but no compensatory uptake occurs from deeper soil layers. New Phytol. 2022, 233, 194–206. [Google Scholar] [CrossRef]
- Brinkmann, N.; Eugster, W.; Buchmann, N.; Kahmen, A. Species-specific differences in water uptake depth of mature temperate trees vary with water availability in the soil. Plant Biology 2019, 21, 71–81. [Google Scholar] [CrossRef]
- Wang, Y.N.; Cao, G.X.; Wang, Y.H.; Webb, A.A.; Yu, P.T.; Wang, X.J. Response of the daily transpiration of a larch plantation to variation in potential evaporation, leaf area index and soil moisture. Sci. Rep. 2019, 9, 4697. [Google Scholar] [CrossRef]
- Ma, J.; Guo, J.B.; Wang, Y.H.; Liu, Z.B.; Gao, D.; Hong, L.; Zhang, Z.Y. Variations in stem radii of Larix principis-rupprechtii to environmental factors at two slope locations in the Liupan Mountains, northwest China. J. For. Res. 2021, 32, 513–527. [Google Scholar] [CrossRef]
- Cunningham, S.C. Stomatal sensitivity to vapour pressure deficit of temperate and tropical evergreen rainforest trees of Australia. Trees 2004, 18, 399–407. [Google Scholar] [CrossRef]
- Fabiani, G.; Schoppach, R.; Penna, D.; Klaus, J. Transpiration patterns and water use strategies of beech and oak trees along a hillslope. Ecohydrology 2021, 15, e2382. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Downslope | Upslope |
|---|---|---|
| Stand density (stems·ha−1) | 933 | 944 |
| Mean DBH (cm) | 18.9 | 20.6 |
| Mean tree height (m) | 16.3 | 16.9 |
| Mean leaf area index (m2·m−2) | 2.96 | 3.59 |
| Soil depth (m) | 1.2 | 1.0 |
| Soil bulk density (g·cm−3) | 1.10 | 0.94 |
| Soil water holding capacity (%) | 33.5 | 37.7 |
| Slope Position | Tree Code | DBH (cm) | H (m) | Sapwood Area (cm2) |
|---|---|---|---|---|
| Downslope | 1 | 12.9 | 13.9 | 72.5 |
| 2 | 14.5 | 14.5 | 87.2 | |
| 3 | 14.9 | 14.7 | 91.1 | |
| 4 | 19.2 | 16.4 | 136.0 | |
| 5 | 24.1 | 18.3 | 194.7 | |
| 6 | 28.2 | 19.9 | 249.6 | |
| Upslope | 1 | 13.9 | 14.2 | 72.6 |
| 2 | 16.3 | 15.3 | 98.5 | |
| 3 | 19.0 | 16.4 | 132.2 | |
| 4 | 21.0 | 17.1 | 160.2 | |
| 5 | 24.2 | 18.0 | 210.4 | |
| 6 | 27.4 | 18.8 | 267.0 |
| Slope Positions | Quantitative Relationship | R2 | p | n |
|---|---|---|---|---|
| Downslope | TL = (−0.100 + 0.101 × (1 − exp(−1.685 × Ta))) × (1.073 × ln(Rs) + 7.365) × (0.100 × (1 − exp(−6.093 × VPD))) × (7.327 × SWC0–20 + 1.330 × SWC20–40 + 2.817 × SWC40–60 + 5.233) | 0.889 | <0.01 | 122 |
| Upslope | TL = (−0.664 + 0.665 × (1 − exp(−4.105 × Ta))) × (2.644 × ln(Rs) − 6.430) × (0.276 × (1 − exp(−8.329 × VPD))) × (1.358 × SWC0–20 + 12.097 × SWC20–40 + 1.130 × SWC40–60 + 16.140) | 0.888 | <0.01 | 122 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Wang, M.; Wang, Y.; Liu, S.; Yu, S.; Ma, J.; Xu, L. Controlling Factors of Spatiotemporal Variations in Transpiration on a Larch Plantation Hillslope in Northwest China. Water 2025, 17, 1756. https://doi.org/10.3390/w17121756
Liu Z, Wang M, Wang Y, Liu S, Yu S, Ma J, Xu L. Controlling Factors of Spatiotemporal Variations in Transpiration on a Larch Plantation Hillslope in Northwest China. Water. 2025; 17(12):1756. https://doi.org/10.3390/w17121756
Chicago/Turabian StyleLiu, Zebin, Mengfei Wang, Yanhui Wang, Shan Liu, Songping Yu, Jing Ma, and Lihong Xu. 2025. "Controlling Factors of Spatiotemporal Variations in Transpiration on a Larch Plantation Hillslope in Northwest China" Water 17, no. 12: 1756. https://doi.org/10.3390/w17121756
APA StyleLiu, Z., Wang, M., Wang, Y., Liu, S., Yu, S., Ma, J., & Xu, L. (2025). Controlling Factors of Spatiotemporal Variations in Transpiration on a Larch Plantation Hillslope in Northwest China. Water, 17(12), 1756. https://doi.org/10.3390/w17121756