



Tackling the Phylogeny of Lampreys—Insight from the Croatia’s Danube Basin

Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Laboratory Protocols
2.3. Data Analyses
2.3.1. Sequence Alignment
2.3.2. Neutrality Test and Analyses of Population Genetic Diversity and Polymorphism
2.3.3. Phylogeographic and Evolutionary Analyses
3. Results
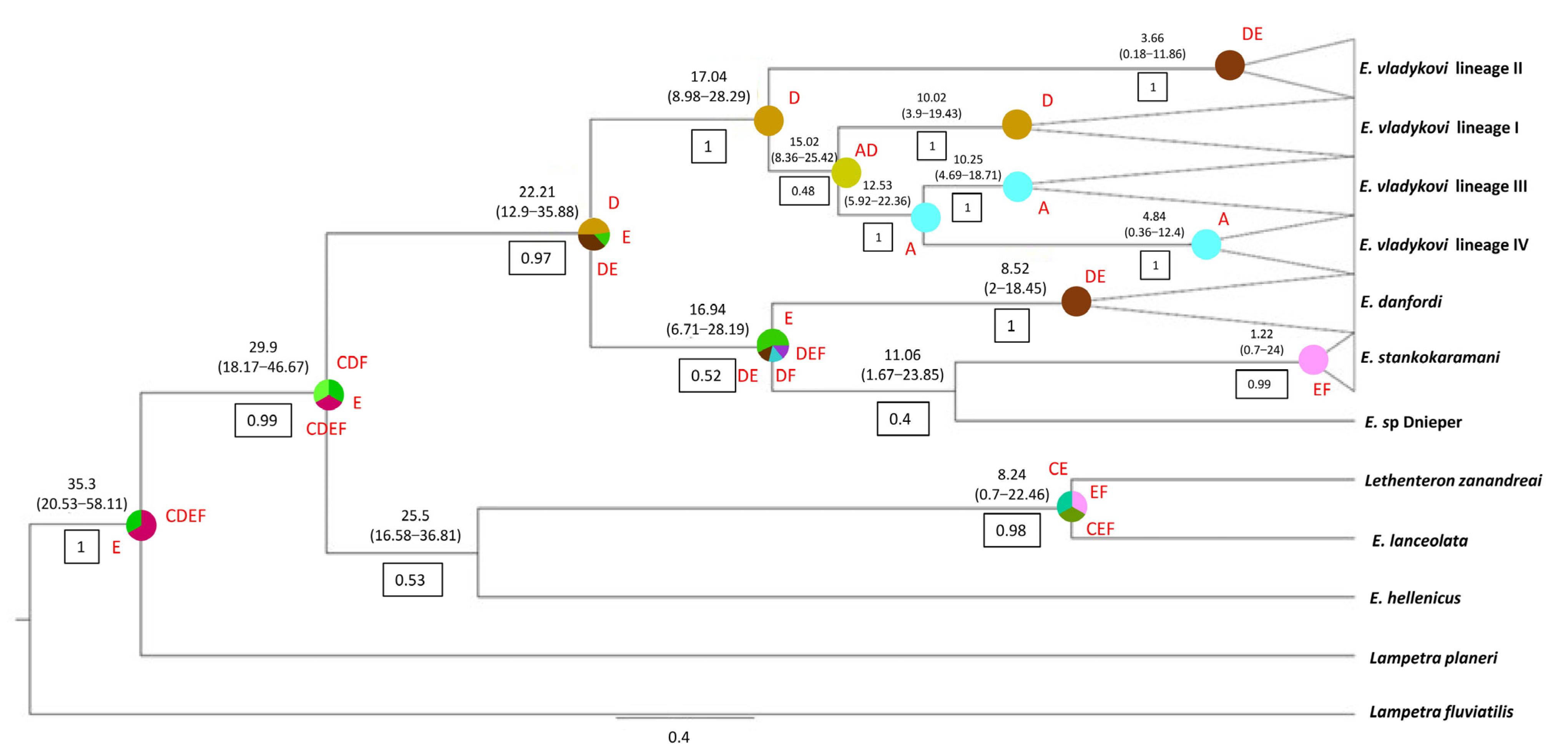
3.1. Phylogenetic Relationships and Distribution of Lampreys in Continental Croatia
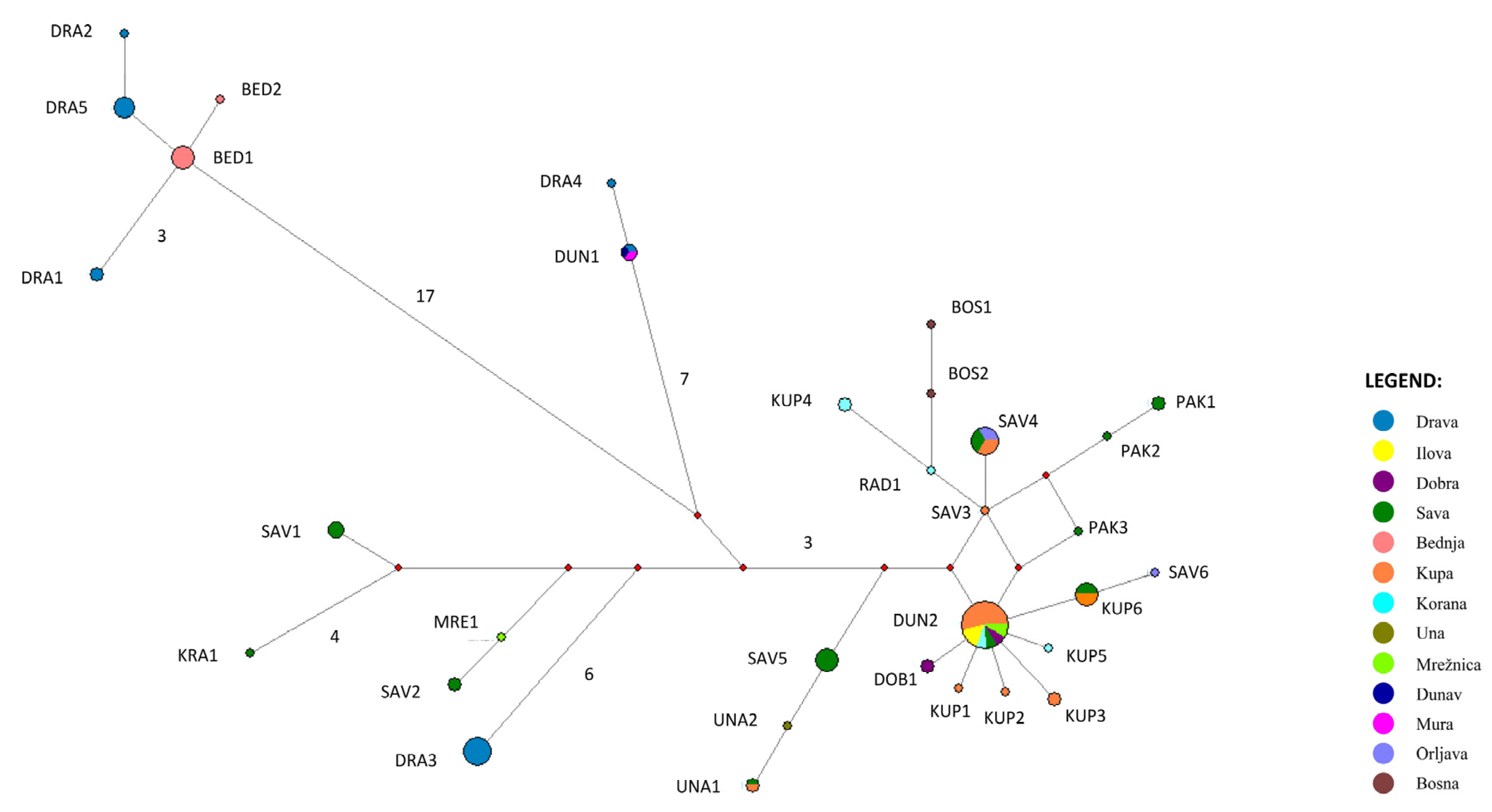
3.2. Inter- and Intraspecific Genetic Diversity and Differentiation
3.3. Molecular Identification of Species and Lineages
3.4. Evolutionary History of the Family Petromyzontidae in Europe
4. Discussion
4.1. Diversity of the Petromyzontidae Family in the Danube Basin in Croatia
4.2. Taxonomic Implications
4.3. Conservation Recommendations
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hewitt, G.M. Post-glacial re-colonization of European biota. Biol. J. Linn. Soc. 1999, 68, 87–112. [Google Scholar] [CrossRef]
- Buj, I.; Podnar, M.; Mrakovčić, M.; Choleva, L.; Šlechtova, V.; Tvrtković, N.; Ćaleta, M.; Mustafić, P.; Marčić, Z.; Zanella, D.; et al. Genetic diversity and phylogenetic relationships of spined loaches (genus Cobitis) in Croatia based on mtDNA and allozyme analyses. Folia Zool. 2008, 57, 71–82. [Google Scholar]
- Ćaleta, M.; Marčić, Z.; Buj, I.; Zanella, D.; Mustafić, P.; Duplić, A.; Horvatić, S. A review of extant Croatian freshwater fish and lampreys: Annotated list and distribution. Croat. J. Fish. 2019, 77, 137–234. [Google Scholar]
- Mrakovčić, M.; Brigić, A.; Buj, I.; Ćaleta, M.; Mustafić, P.; Zanella, D. Red Book of Freshwater Fish of Croatia; Ministry of Culture, State Institute for Nature Protection: Zagreb, Croatia, 2006. [Google Scholar]
- Raguž, L.; Buj, I.; Marčić, Z.; Veble, V.; Ivić, L.; Zanella, D.; Horvatić, S.; Mustafić, P.; Ćaleta, M.; Sabolić, M. First look into the evolutionary history, phylogeographic and population genetic structure of the Danube barbel in Croatia. Knowl. Manag. Aquat. Ecosyst. 2021, 422, 13. [Google Scholar] [CrossRef]
- Kottelat, M.; Freyhof, J. Handbook of European Freshwater Fishes, 1st ed.; Kottelat: Cornol, Switzerland; Freyhof: Berlin, Germany, 2007; p. 646. [Google Scholar]
- De Cahsan, B.; Nagel, R.; Schedina, I.M.; King, J.J.; Bianco, P.G.; Tiedemann, R.; Ketmaier, V. Phylogeography of the European brook lamprey (Lampetra planeri) and the European river lamprey (Lampetra fluviatilis) species pair based on mitochondrial data. J. Fish Biol. 2020, 96, 905–912. [Google Scholar] [CrossRef]
- Mateus, C.S.; Stange, M.; Berner, D.; Roesti, M.; Quintella, B.R.; Alves, M.J.; Almeida, P.R.; Salzburger, W. Strong genome-wide divergence between sympatric European river and brook lampreys. Curr. Biol. 2013, 23, R649–R650. [Google Scholar] [CrossRef] [PubMed]
- Rüber, L.; Gandolfi, A.; Foresti, D.; Paltrinieri, L.; Splendiani, A.; Seehausen, O. Phylogenetic and biogeographic history of brook lampreys (Lampetra: Petromyzontidae) in the river basins of the Adriatic Sea based on DNA barcode data. Ecol. Evol. 2023, 13, e10496. [Google Scholar] [CrossRef] [PubMed]
- Zanandrea, G. Speciation among lampreys. Nature 1959, 184, 380. [Google Scholar] [CrossRef]
- Zanandrea, G.I.S. Recenti ricerche sulle forme appaiate di Lamprede dell’Italia e del Danubio. Bolletino Zool. 1959, 26, 545–554. [Google Scholar] [CrossRef]
- Zanandrea, G. Studies on European lampreys. Evolution 1961, 15, 523–534. [Google Scholar] [CrossRef]
- Boeker, C.; Geist, J. Lampreys as ecosystem engineers: Burrows of Eudontomyzon sp. and their impact on physical, chemical, and microbial properties in freshwater substrates. Hydrobiologia 2016, 777, 171–181. [Google Scholar] [CrossRef]
- Pough, F.H.; Janis, C.H.; Heiser, J.B. Vertebrate Life, 9th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 2013; pp. 1–69. [Google Scholar]
- Young, J.Z. The Life of Vertebrates, 3rd ed.; Clarendon Press: Oxford, UK, 1985; pp. 76–109. [Google Scholar]
- Gess, R.W.; Coates, M.I.; Rubidge, B.S. A lamprey from the Devonian period of South Africa. Nature 2006, 443, 981–984. [Google Scholar] [CrossRef] [PubMed]
- Tutman, P.; Freyhof, J.; Dulčić, J.; Glamuzina, B.; Geiger, M. Lampetra soljani, a new brook lamprey from the southern Adriatic Sea basin (Petromyzontiformes: Petromyzontidae). Zootaxa 2017, 4273, 531–548. [Google Scholar] [CrossRef] [PubMed]
- Vladykov, V.D.; Kott, E. List of the Northern Hemisphere Lampreys (Petromyzondidae) and Their Distribution, 3rd ed.; Miscellaneous Special Publications; Department Fisheries and Oceans: Ottawa, Canada, 1979; pp. 1–30. [Google Scholar]
- Gill, H.S.; Renaud, C.B.; Chapleau, F.; Mayden, R.L.; Potter, I.C. Phylogeny of living parasitic lampreys (Petromyzontiformes) based on morphological data. Copeia 2003, 687–703. [Google Scholar] [CrossRef]
- Krappe, M. Quantitative Analysen Populationsbiologischer Phänomene im Lebenszyklus des Bachneunauges Lampetra planeri (Bloch, 1784). Ph.D. Thesis, Fakultät der Universität Rostock, Rostock, Germany, 2005. [Google Scholar]
- Perea, S.; Böhme, M.; Zupančič, P.; Freyhof, J.; Šanda, R.; Özuluğ, M.; Abdoli, A.; Doadrio, I. Phylogenetic relationships and biogeographical patterns in Circum-Mediterranean subfamily Leuciscinae (Teleostei, Cyprinidae) inferred from both mitochondrial and nuclear data. BMC Evol. Biol. 2010, 10, 265. [Google Scholar] [CrossRef] [PubMed]
- Castresana, J. Cytochrome b phylogeny and the taxonomy of great apes and mammals. Mol. Biol. Evol. 2001, 18, 465–471. [Google Scholar] [CrossRef]
- Patwardhan, A.; Ray, S.; Roy, A. Molecular Markers in Phylogenetic Studies—A Review. J. Phylogen. Evol. Biol. 2014, 2, 131. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods), Version 4; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Nakhleh, L.; Guohua, J.; Fengmei, Z.; Mellor-Crummey, J. Reconstructing Phylogenetic Networks Using Maximum Parsimony. In Proceedings of the IEEE Computational Systems Bioinformatics Conference (CSB’05), Stanford, CA, USA, 8–11 August 2005. [Google Scholar]
- Lang, N.J.; Roe, K.J.; Renaud, C.B.; Gill, H.S.; Potter, I.C.; Freyhof, J.; Naseka, A.M.; Cochran, P.A.; Pérez, H.E.; Habit, E.M.; et al. Novel Relationships among Lampreys (Petromyzontiformes) Revealed by a Taxonomically Comprehensive Molecular Data Set. Am. Fish. Soc. Symp. 2009, 72, 41–55. [Google Scholar]
- Delarbre, C.; Escriva, H.; Gallut, C.; Barriel, V.; Kourilsky, P.; Janvier, P.; Laudet, V.; Gachelin, G. The complete nucleotide sequence of the mitochondrial DNA of the agnathan Lampetra fluviatilis: Bearings on the phylogeny of cyclostomes. Mol. Biol. Evol. 2000, 17, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Levin, B.; Ermakov, A.; Ermakov, O.; Levina, M.; Sarycheva, O.; Sarychev, V. Ukrainian Brook Lamprey Eudontomyzon mariae (Berg): Phylogenetic position, genetic diversity, distribution, and some data on biology. In Jawless Fishes of the World, 2nd ed.; Orlov, A., Beamish, R., Eds.; Cambridge Scholar Publishing, Lady Stephenson Library: Newcastle upon Tyne, UK, 2016; Volume 1, pp. 154–196. [Google Scholar]
- Fox, A.M. A Molecular Phylogenetic Analysis of the Lamprey Genera Eudontomyzon, Lampetra, and Lethenteron. Ph.D. Thesis, University of Manitoba, Department of Biological Sciences, Winnipeg, MB, Canada, 29 August 2016. [Google Scholar]
- Bouckaert, R.; Heled, J.; Kuhnert, D.; Vaughan, T.; Wu, C.-H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef]
- Pereira, A.M.; Levy, A.; Vukić, J.; Šanda, R.; Levin, B.A.; Freyhof, J.; Geiger, M.; Choleva, L.; Francisco, S.M.; Robalo, J.I. Putting European lampreys into perspective: A global-scale multilocus phylogeny with a proposal for a generic structure of the Petromyzontidae. J. Zool. Syst. Evol. Res. 2021, 59, 1982–1993. [Google Scholar] [CrossRef]
- Yu, Y.; Harris, A.J.; He, X. S-DIVA (Statistical Dispersal-Vicariance Analysis): A tool for inferring biogeographic histories. Mol. Phylogenet. Evol. 2010, 56, 848–850. [Google Scholar] [CrossRef] [PubMed]
- Nei, M. Evolution of human races at the gene level. Prog. Clin. Biol. Res. 1982, 103 Pt A, 167–181. [Google Scholar]
- Lynch, M.; Crease, T.J. The analysis of population survey data on DNA sequence variation. Mol. Biol. Evol. 1990, 7, 377–394. [Google Scholar] [PubMed]
- Hudson, R.R.; Slatkin, M.; Maddison, W.P. Estimation of levels of gene flow from DNA sequence data. Genetics 1992, 132, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Krezsek, C.; Olariu, C. Filling of sedimentary basins and the birth of large rivers: The lower Danube network in the Dacian Basin, Romania. Glob. Planet. Chang. 2021, 197, 103391. [Google Scholar] [CrossRef]
- Magyar, I.; Radivojević, D.; Sztano, O.; Synak, R.; Ujszaszi, K.; Pocsik, M. Progradation of the paleo-Danube shelf margin across the Pannonian Basin during the Late Miocene and Early Pliocene. Glob. Planet. Chang. 2013, 103, 168–173. [Google Scholar] [CrossRef]
- Buj, I.; Šanda, R.; Marčić, Z.; Ćaleta, M.; Mrakovčić, M. Combining morphology and genetics in resolving taxonomy—a systematic revision of spined loaches (genus Cobitis; Cypriniformes, Actinopterygii) in the Adriatic Watershed. PLoS ONE 2014, 9, e99833. [Google Scholar] [CrossRef]
- Meyer, L.; Brunken, H. Historical occurrence and current distribution of migrating fishes and lampreys (Osteichthyes et Cyclostomata) in the drainage system of the River Aller (Lower Saxony) with an evaluation of future developments of their stocks. Braunschweiger Naturkundliche Schriften 1997, 5, 281–303. (in German). [Google Scholar]
- Ojutkangas, E.; Aronen, K.; Laukkanen, E. Distribution and abundance of river lamprey (Lampetra fluviatilis) ammocoetes in the regulated river Perhonjoki. Regul. Rivers Res. Manag. 1995, 10, 239–245. [Google Scholar] [CrossRef]
- Kelly, F.L.; King, J.J. A review of the ecology and distribution of three lamprey species, Lampetra fluviatilis (L.), Lampetra planeri (Bloch) and Petromyzon marinus (L.): A context for conservation and biodiversity considerations in Ireland. In Biology and Environment: Proceedings of the Royal Irish Academy; Royal Irish Academy: Dublin, Ireland, 2001; Volume 101, p. 101. [Google Scholar]
- Clemens, B.J.; Arakawa, H.; Baker, C.; Coghlan, S.; Kucheryavyy, A.; Lampman, R.; Lança, M.J.; Mateus, C.S.; Miller, A.; Nazari, H.; et al. Management of anadromous lampreys: Common threats, different approaches. J. Great Lakes Res. 2021, 47 (Suppl. S1), S129–S146. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality (Name & Number) | Drainage System | Number of Samples (n) | cyt b Haplotype | GenBank Accession Numbers |
|---|---|---|---|---|
| Maslenjača (1) | Ilova | 1 | DUN2 | PP661384 |
| Toplica, Daruvar (2) | Ilova | 2 | DUN2 | PP661384 |
| Lešće (3) | Dobra | 7 | DOB1, DUN2 | PP661393, PP661384 |
| Sunja (4) | Sava | 8 | KUP6, SAV5 | PP661399, PP661390 |
| Osijek (5) | Drava | 1 | DRA1 | PP661378 |
| Voćin (6) | Drava | 7 | DRA2, DRA3, DRA5 | PP661379, PP661380, PP661382 |
| Štorgač (7) | Drava | 1 | DUN1 | PP661383 |
| Repaški most (8) | Drava | 1 | DRA4 | PP661381 |
| Petrijevci (9) | Drava | 1 | DRA1 | PP661378 |
| Ivanečka Željeznica (10) | Bednja | 1 | BED1 | PP661376 |
| Bednja (11) | Bednja | 2 | BED1 | PP661376 |
| Voća (12) | Bednja | 4 | BED1, BED2 | PP661376, PP661377 |
| Bubnjarci (13) | Kupa | 1 | DUN 2 | PP661384 |
| Brod na Kupi (14) | Kupa | 2 | KUP1, KUP2 | PP661394, PP661395 |
| Lazina (15) | Kupa | 1 | KUP3 | PP661396 |
| Pribanjci (16) | Kupa | 1 | DUN2 | PP661384 |
| Furjašnica (17) | Korana | 2 | KUP4 | PP661397 |
| Brezova Glava (18) | Korana | 2 | KUP5, DUN2 | PP661398, PP661384 |
| Stubička Slatina (19) | Sava | 1 | KRA1 | PP661385 |
| Brzaja (20) | Sava | 5 | SAV4 | PP661389 |
| Pakra (21) | Sava | 7 | PAK1, PAK2, PAK3, DUN2 | PP661402, PP661403, PP661404, PP661384 |
| Bručina (22) | Kupa | 3 | DUN2 | PP661384 |
| Ruševnica (23) | Kupa | 1 | KUP6 | PP661399 |
| Glinica (24) | Kupa | 1 | KUP6 | PP661399 |
| Čemernica (25) | Kupa | 1 | UNA1 | PP661400 |
| Buzeta, Šibine (26) | Kupa | 4 | KUP6, DUN2 | PP661399, PP661384 |
| Žirovnica (27) | Una | 2 | UNA1, UNA2 | PP661400, PP661401 |
| Petrinjčica (28) | Kupa | 5 | SAV3, SAV4, DUN2 | PP661388, PP661389, PP661384 |
| Lupinjak (29) | Sava (Sutla) | 3 | SAV1, SAV2 | PP661386 PP661387 |
| Mostanje (30) | Mrežnica | 2 | MRE1, DUN2 | PP661392, PP661384 |
| Batina (31) | Dunav | 1 | DUN1 | PP661383 |
| Goričan (32) | Mura | 1 | DUN1 | PP661383 |
| Orljava (33) | Orljava | 4 | SAV4, SAV6 | PP661389, PP661391 |
| Živković kosa (34) | Korana | 1 | RAD1 | PP661407 |
| Ušće Lašve (35) | Bosna | 2 | BOS1, BOS2 | PP661405, PP661406 |
| Terezino polje (36) | Drava | 5 | DRA3 | PP661380 |
| Bukvik (37) | Drava | 5 | DRA3 | PP661380 |
| Jaševnica (38) | Kupa | 2 | KUP3, DUN2 | PP661396, PP661384 |
| Ozalj (39) | Kupa | 2 | DUN2 | PP661384 |
| Genetic Marker | cyt b |
|---|---|
| PCR conditions | 35 cycles of 45 s at 92 °C, following 90 s at 48 °C, following 105 s at 72 °C |
| PCR primers | ProK (5′TTATTTAATGTTAAGATRCTAGCTTTGG3′) Pak-Glu F (5′CACCGTTGTAGAATTCAA CTATAAG3′) |
| Species | bp (Number of Base Pairs) | Locality | GenBank Accession Number | Reference |
|---|---|---|---|---|
| Eudontomyzon danfordi Regan, 1911 | 1133 | Zdychava River (SVK) | GQ206158.1 | [28] |
| Eudontomyzon mariae (Berg, 1931) | 1133 | Ivianka River (UKR) | GQ206162.1 | [28] |
| Eudontomyzon stankokaramani (Karaman, 1974) | 1133 | Zeta River (MNE) | GQ206189.1 | [28] |
| Eudontomyzon vladykovi Oliva and Zanandrea, 1959 | 1133 | Studenec Creek (SVK) | GQ206161.1 | [28] |
| Lampetra fluviatilis (Linnaeus, 1758) | 16,159 | Garonne Estuary (FRA) | NC001131.1 | [29] |
| Eudontomyzon lanceolata (Kux and Steiner, 1972) | 1133 | Chakhtsutsyr River (RUS) | GQ206176.1 | [28] |
| Lampetra planeri (Bloch, 1784) | 1133 | Kalte Moldau River (DEU) | GQ206149.1 | [28] |
| Lethenteron zanandreai (Vladykov, 1955) | 1133 | Vipava River (SLO) | GQ206184.1 | [28] |
| Eudontomyzon sp. Dnieper | 1164 | Ugra River (RUS) | KP135487.1 | [30] |
| Eudontomyzon sp. Dnieper | 1164 | Rudyanka River (RUS) | KP135483.1 | [30] |
| Eudontomyzon sp. Dnieper | 1164 | Sigosa River (RUS) | KP135485.1 | [30] |
| Eudontomyzon sp. Dnieper | 1164 | Vyazma River (RUS) | KP135482.1 | [30] |
| Eudontomyzon hellenicus (Vladykov, Renaud, Kott and Economidis, 1982) | 1133 | Strymon River (GRC) | GQ206160.1 | [28] |
| Lampetra fluviatilis (Linnaeus, 1758) | 1133 | Luga River (RUS) | GQ206175.1 | [28] |
| Eudontomyzon stankokaramani (Karaman, 1974) | 1191 | unknown river (SLO) | KX787432.1 | [31] |
| Eudontomyzon lanceolata (Kux and Steiner, 1972) | 1191 | unknown river (TUR) | KX787431.1 | [31] |
| Species/Lineage | N | h | S | Hd | k | π | ƞ |
|---|---|---|---|---|---|---|---|
| E. danfordi | 15 | 5 | 6 | 0.752 | 1.524 | 0.00128 | 6 |
| E. vladykovi | 88 | 30 | 53 | 0.895 | 7.158 | 0.00601 | 54 |
| vladykovi I | 16 | 5 | 19 | 0.667 | 6.983 | 0.00586 | 19 |
| vladykovi II | 4 | 2 | 1 | 0.5 | 0.5 | 0.00042 | 1 |
| vladykovi III | 59 | 20 | 23 | 0.8 | 2.414 | 0.00203 | 23 |
| vladykovi IV | 9 | 3 | 2 | 0.556 | 0.722 | 0.00061 | 2 |
| Species/Lineage | Vladykovi I | Vladykovi II | Vladykovi III | Vladykovi IV |
|---|---|---|---|---|
| E. danfordi | 0 | 0 | 1 | 0 |
| vladykovi I | - | 0 | 3 | 0 |
| vladykovi II | - | - | 1 | 0 |
| vladykovi III | - | - | - | 0 |
| Species/Lineage | E. danfordi | Vladykovi I | Vladykovi II | Vladykovi III | Intraspecific/Intralineage |
|---|---|---|---|---|---|
| E. danfordi | 0.08–0.41 (0.20) | ||||
| vladykovi I | 1.84–2.68 (2.26) | 0.08–1.25 (0.70) | |||
| vladykovi II | 2.01–2.35 (2.18) | 1.09–1.51 (1.30) | / (0.10) | ||
| vladykovi III | 1.93–2.35 (2.14) | 0.83–1.59 (1.21) | 1.09–1.42 (1.255) | 0.80–0.58 (0.30) | |
| vladykovi IV | 1.84–2.09 (1.97) | 0.83–1.34 (1.09) | 0.92–1.09 (1.01) | 0.33–0.75 (0.54) | 0.08–0.16 (0.10) |
| Species/Lineage | Nm [35] | Nm [36] | Nm [37] |
|---|---|---|---|
| E. danfordi:E. vladykovi | 0.61 | 0.1 | 0.11 |
| vladykoviI:vladykoviII | 0.67 | 0.15 | 0.15 |
| vladykoviI:vladykoviIII | 0.53 | 0.26 | 0.26 |
| vladykoviI:vladykoviIV | 0.51 | 0.21 | 0.21 |
| vladykoviII:vladykoviIII | 0.74 | 0.6 | 0.6 |
| vladykoviII:vladykoviIV | 0.06 | 0.03 | 0.03 |
| vladykoviIII:vladykoviIV | 1.19 | 0.2 | 0.2 |
| Species/Lineage | Vladykovi I | Vladykovi II | Vladykovi III | Vladykovi IV |
|---|---|---|---|---|
| E. danfordi | 19 | 24 | 21 | 21 |
| vladykovi I | - | 9 | 6 | 6 |
| vladykovi II | - | - | 12 | 10 |
| vladykovi III | - | - | - | 3 |
| Species/Lineage | |||||
|---|---|---|---|---|---|
| Nucleotide Spot | E. danfordi | Vladykovi I | Vladykovi II | Vladykovi III | Vladykovi IV |
| 36 | T | C | C | C | C |
| 57 | C | T | T | T | T |
| 72 | C | T | T | T | T |
| 156 | G | A | A | A | A |
| 157 | C | T | T | T | T |
| 162 | T | C | C | C | C |
| 312 | A | G | A | A | A |
| 378 | A | A | G | A | A |
| 390 | T | T | C | T | T |
| 480 | A | G | G | G | G |
| 483 | A | G | G | G | G |
| 531 | C | C | C | C | T |
| 588 | A | A | G | A | A |
| 630 | T | T | C | T | T |
| 639 | G | A | A | A | A |
| 675 | T | C | C | C | C |
| 726 | C | T | T | T | T |
| 771 | A | C | C | C | C |
| 807 | G | A | G | G | G |
| 813 | C | C | C | T | C |
| 909 | G | A | A | A | A |
| 1155 | G | G | A | G | G |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pleše, S.; Buj, I. Tackling the Phylogeny of Lampreys—Insight from the Croatia’s Danube Basin. Water 2024, 16, 1153. https://doi.org/10.3390/w16081153
Pleše S, Buj I. Tackling the Phylogeny of Lampreys—Insight from the Croatia’s Danube Basin. Water. 2024; 16(8):1153. https://doi.org/10.3390/w16081153
Chicago/Turabian StylePleše, Sara, and Ivana Buj. 2024. "Tackling the Phylogeny of Lampreys—Insight from the Croatia’s Danube Basin" Water 16, no. 8: 1153. https://doi.org/10.3390/w16081153
APA StylePleše, S., & Buj, I. (2024). Tackling the Phylogeny of Lampreys—Insight from the Croatia’s Danube Basin. Water, 16(8), 1153. https://doi.org/10.3390/w16081153