
Review of the Influence of Climate Change on the Hydrologic Cycling and Gaseous Fluxes of Mercury in Boreal Peatlands: Implications for Restoration



Abstract
1. Introduction
2. Review of Boreal Peatland Climate Studies
3. Climate Change Influences on Peatland Mercury Cycling
3.1. Warming Effects on Peatland Hydrologic Cycling of Mercury
3.2. Water Balance Effects on Peatland Hydrologic Mercury Cycling
3.3. Elevated CO2 Effects on Peatland Hydrologic Mercury Cycling
3.4. Interacting Effects of Climate Change on Peatland Hydrologic Mercury Cycling
3.5. Warming Effects on the Gaseous Fluxes of Hg
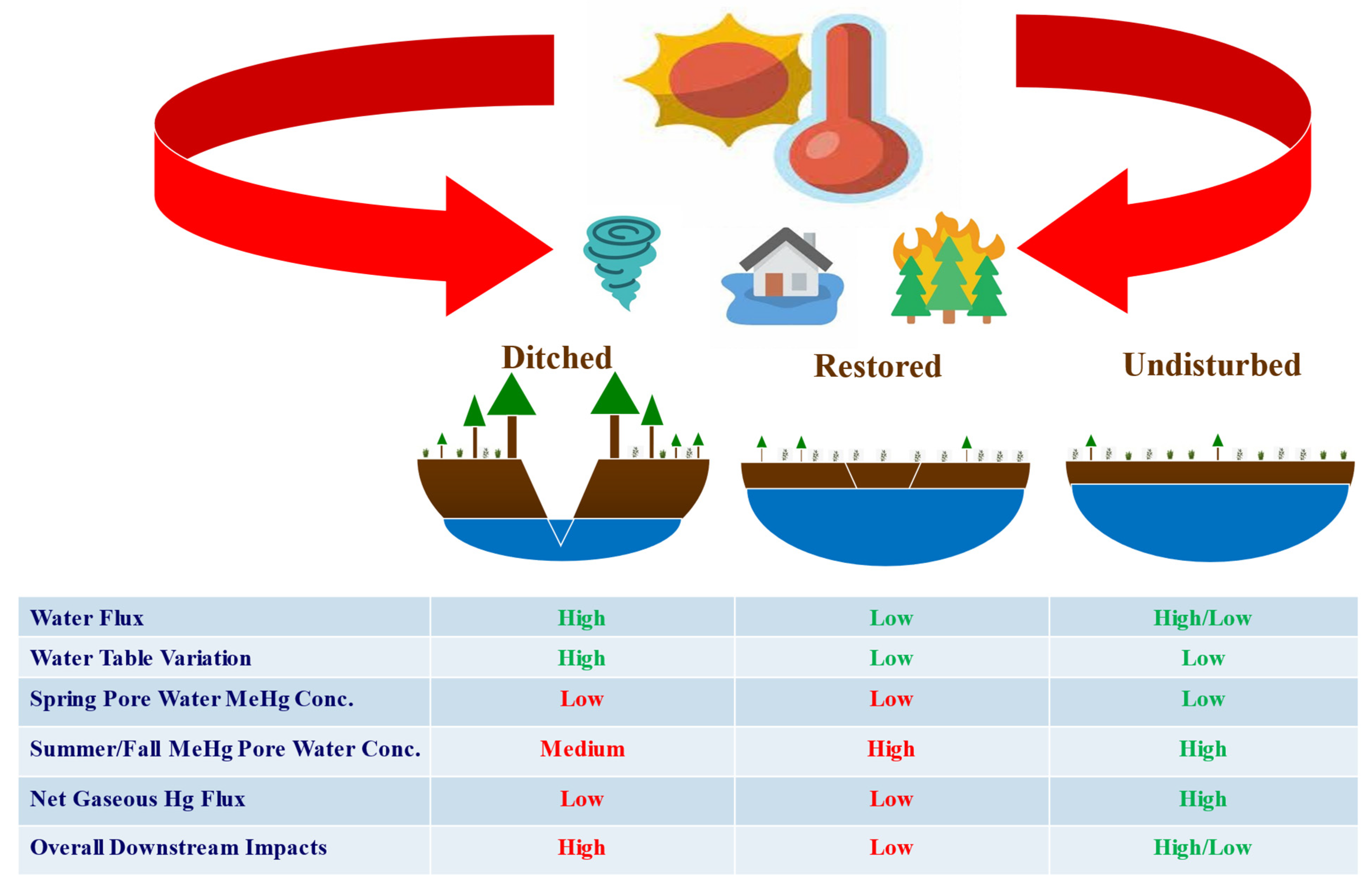
4. Implication of Peatland Restoration and Subsequent Climate Change on Hg Cycles
5. Conclusions
6. Future Directions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lavoie, R.A.; Jardine, T.D.; Chumchal, M.M.; Kidd, K.A.; Campbell, L.A. Biomagnification of mercury in aquatic food webs: A worldwide meta-analysis. Environ. Sci. Technol. 2013, 47, 13385–13394. [Google Scholar] [CrossRef] [PubMed]
- Rice, K.M.; Walker, E.M., Jr.; Wu, M.; Gillette, C.; Blough, E.R. Environmental mercury and its toxic effects. J. Prev. Med. Public Health 2014, 47, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Jiskra, M.; Nilsson, M.B.; Osterwalder, S.; Zhu, W.; Mauquoy, D.; Skyllberg, U.; Enrico, M.; Peng, H.; Song, Y.; et al. Mercury deposition and redox transformation processes in peatland constrained by mercury stable isotopes. Nat. Commun. 2023, 14, 7389. [Google Scholar] [CrossRef] [PubMed]
- Obrist, D.; Kirk, J.L.; Zhang, L.; Sunderland, E.M.; Jiskra, M.; Selin, N.E. A review of global environmental mercury processes in response to human and natural perturbations: Changes of emissions, climate, and land use. Ambio 2018, 47, 116–140. [Google Scholar] [CrossRef] [PubMed]
- Driscoll, C.T.; Mason, R.P.; Chan, H.M.; Jacob, D.J.; Pirrone, N. Mercury as a global pollutant: Sources, pathways, and effects. Environ. Sci. Technol. 2013, 47, 4967–4983. [Google Scholar] [CrossRef] [PubMed]
- Krabbenhoft, D.P.; Sunderland, E.M. Global change and mercury. Science 2013, 341, 1457–1458. [Google Scholar] [CrossRef] [PubMed]
- Kolka, R.; Trettin, C.; Tang, W.; Krauss, K.; Bansal, S.; Drexler, J.; Wickland, K.; Chimner, R.; Hogan, D.; Pindilli, E.J.; et al. Chapter 13: Terrestrial wetlands. In Second State of the Carbon Cycle Report (SOCCR2): A Sustained Assessment Report; Cavallaro, N., Shrestha, G., Birdsey, R., Mayes, M.A., Najjar, R.G., Reed, S.C., Romero-Lankao, P., Zhu, Z., Eds.; U.S. Global Change Research Program: Washington, DC, USA, 2018; pp. 507–567. [Google Scholar] [CrossRef]
- Humpenöder, F.; Karstens, K.; Lotze-Campen, H.; Leifeld, J.; Menichetti, L.; Barthelmes, A.; Popp, A. Peatland protection and restoration are key for climate change mitigation. Environ. Res. Lett. 2020, 15, 104093. [Google Scholar] [CrossRef]
- Temmink, R.J.M.; Lamers, L.P.M.; Angelini, C.; Bouma, T.J.; Fritz, C.; van de Koppel, J.; Lexmond, R.; Rietkerk, M.; Silliman, B.R.; Joosten, H.; et al. Recovering wetland biogeomorphic feedbacks to restore the world’s biotic carbon hotspots. Science 2022, 376, eabn1479. [Google Scholar] [CrossRef]
- Price, J.; Heathwaite, A.; Baird, A. Hydrological processes in abandoned and restored peatlands: An overview of management approaches. Wetl. Ecol. Manag. 2003, 11, 65–83. [Google Scholar] [CrossRef]
- Krause, L.M.; McCullough, K.J.; Kane, E.S.; Kolka, R.K.; Chimner, R.A.; Lilleskov, E.A. Impacts of historical ditching on peat volume and carbon in northern Minnesota peatlands. J. Environ. Manag. 2021, 296, 113090. [Google Scholar] [CrossRef]
- Minnesota Pollution Control Agency. Greenhouse Gas Emissions in Minnesota (2005–2020). Report to the Legislature. 2023, pp. 1–25. Available online: https://www.pca.state.mn.us/sites/default/files/lraq-2sy23.pdf (accessed on 17 January 2024).
- Schaefer, K.; Elshorbany, Y.; Jafarov, E.; Schuster, P.F.; Striegl, R.G.; Wickland, K.P.; Sunderland, E.M. Potential impacts of mercury released from thawing permafrost. Nat. Commun. 2020, 11, 4650. [Google Scholar] [CrossRef] [PubMed]
- Turetsky, M.R.; Harden, J.W.; Friedli, H.R.; Flannigan, M.D.; Payne, N.; Crock, J.; Radke, L.F. Wildfires threaten mercury stocks in northern soils, Geophys. Res. Lett. 2006, 33, L16403. [Google Scholar] [CrossRef]
- Yang, Z.; Fang, W.; Lu, X.; Sheng, G.-P.; Graham, D.E.; Liang, L.; Wullschleger, S.D.; Gu, B. Warming increases methylmercury production in an Arctic soil. Environ. Pollut. 2016, 214, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.P.G.; Branfireun, B.A.; Kolka, R.K. Assessing sulfate and carbon controls on net methylmercury production in peatlands: An in situ mesocosm approach. Appl. Geochem. 2008, 23, 503–518. [Google Scholar] [CrossRef]
- Oswald, C.J.; Carey, S.K. Total and methyl mercury concentrations in sediment and water of a constructed wetland in the Athabasca Oil Sands Region. Environ. Pollut. 2016, 213, 628–637. [Google Scholar] [CrossRef]
- Waddington, J.M.; Day, S.M. Methane emissions from a peatland following restoration. J. Geophys. Res. 2007, 112, G03018. [Google Scholar] [CrossRef]
- McCarter, C.P.R.; Sebestyen, S.D.; Jeremiason, J.D.; Nater, E.A.; Kolka, R.K. Methylmercury export from a headwater peatland catchment decreased with cleaner emissions despite opposing effect of climate warming. Water Resour. Res. 2024; in press. [Google Scholar]
- Hanson, P.J.; Riggs, J.S.; Nettles, R.; Phillips, J.R.; Krassovski, M.B.; Hook, L.A.; Gu, L.; Richardson, A.D.; Aubrecht, D.M.; Ricciuto, D.M.; et al. Attaining whole-ecosystem warming using air and deep-soil heating methods with an elevated CO2 atmosphere. Biogeosciences 2017, 14, 861–883. [Google Scholar] [CrossRef]
- Sun, T.; Lindo, Z.; Branfireun, B.A. Ground warming releases inorganic mercury and increases net methylmercury production in two boreal peatland types. Front. Environ. Sci. 2023, 11, 1100443. [Google Scholar] [CrossRef]
- Potvin, L.; Kane, E.S.; Chimner, R.D.; Kolka, R.K.; Lilleskov, E.A. Effects of water table position and plant functional group on plant community, aboveground production, and peat properties in a peatland mesocosm experiment (PEATcosm). Plant Soil 2015, 387, 277–294. [Google Scholar] [CrossRef]
- Sirota, J.I.; Kolka, R.K.; Sebestyen, S.D.; Nater, E.A. Mercury dynamics in the pore water of peat columns during experimental freezing and thawing. J. Environ. Qual. 2020, 49, 404–416. [Google Scholar] [CrossRef]
- Rosset, T.; Binet, S.; Rigal, F.; Gandois, L. Peatland dissolved organic carbon export to surface waters: Global significance and effects of anthropogenic disturbance. Geophys. Res. Lett. 2022, 49, e2021GL096616. [Google Scholar] [CrossRef]
- Hudelson, K.E.; Drevnick, P.E.; Wang, F.; Armstrong, D.; Fisk, A.T. Mercury methylation and demethylation potentials in Arctic lake sediments. Chemosphere 2020, 248, 126001. [Google Scholar] [CrossRef] [PubMed]
- Pierce, C. The Effect of Climate Change on Mercury in Boreal Peatlands. Retrieved from the University of Minnesota Digital Conservancy. 2023. Available online: https://hdl.handle.net/11299/258880 (accessed on 17 January 2024).
- Nater, E. Assessing Release of Mercury and Sulfur on Aquatic Communities. Report to the Legislative Citizens Commission on Minnesota Resources. 2021; 42p. Available online: https://www.lccmr.mn.gov/projects/2017/finals/2017_04i.pdf (accessed on 17 January 2024).
- Garrioch, I.; Behrens, K.; Nater, E.; Kolka, R. Understanding Mercury Gaseous Fluxes at Marcell and the SPRUCE Experiment; National Atmospheric Deposition Program Fall Meeting and Scientific Symposium: Madison, WI, USA, 2023. [Google Scholar]
- Haynes, K.M.; Kane, E.; Potvin, L.; Lilleskov, E.; Kolka, R.; Mitchell, C.P.J. Mobility and transport of mercury and methylmercury in peat as a function of changes in water table regime and plant functional groups. Glob. Biogeochem. Cycles 2017, 31, 233–244. [Google Scholar] [CrossRef]
- Haynes, K.M.; Kane, E.; Potvin, L.; Lilleskov, E.; Kolka, R.K.; Mitchell, C.P.J. Gaseous mercury fluxes in peatlands and the potential influence of climate change. Atmos. Environ. 2017, 154, 247–259. [Google Scholar] [CrossRef]
- Berkeley Earth. Global Temperature Report for 2023. Online: Global Temperature Report for 2023—Berkeley Earth 2024. Available online: https://berkeleyearth.org/global-temperature-report-for-2023/ (accessed on 17 January 2024).
- Pierce, C.E.; Furman, O.S.; Nicholas, S.L.; Wasik, J.C.; Gionfriddo, C.M.; Wymore, A.M.; Sebestyen, S.D.; Kolka, R.K.; Mitchell, C.P.J.; Griffiths, N.A.; et al. The role of ester sulfate and organic disulfide in the mercury methylation in peat soils. Environ. Sci. Technol. 2022, 56, 1433–1444. [Google Scholar] [CrossRef]
- Coleman Wasik, J.K.; Engstrom, D.R.; Mitchell, C.P.J.; Swain, E.B.; Monson, B.A.; Balogh, S.J.; Jeremiason, J.D.; Branfireun, B.A.; Kolka, R.K.; Almendinger, J.E. Hydrologic fluctuations and sulfate regeneration increase methylmercury in an experimental peatland. J. Geophys. Res. Biogeosci. 2015, 120, 1697–2017. [Google Scholar] [CrossRef]
- Ward, E.J.; Warren, J.M.; McLennan, D.A.; Dusenge, M.E.; Way, D.A.; Wullschleger, S.D.; Hanson, P.J. Photosynthetic and respiratory responses of two bog shrub species to whole ecosystem warming and elevated CO2 at the Boreal-Temperate ecotone. Front. For. Glob. Chang. 2019, 2, 54. [Google Scholar] [CrossRef]
- Iversen, C.M.; Latimer, J.M.; Brice, D.J.; Childs, J.; Stel, H.V.; Defrenne, C.E.; Graham, J.D.; Griffiths, N.A.; Malhotra, A.; Norby, R.J.; et al. Whole-ecosystem warming increases plant available nitrogen and phosphorus in an ombrotrophic bog. Ecosystems 2022, 26, 86–113. [Google Scholar] [CrossRef]
- Haynes, K.; Kane, E.S.; Potvin, L.; Lilleskov, E.A.; Kolka, R.K.; Mitchell, C. Impacts of experimental alteration of water table regime and vascular plant community composition on peat mercury profiles and methylmercury production. Sci. Total Environ. 2019, 682, 611–622. [Google Scholar] [CrossRef]
- Antala, M.; Juszczak, R.; van der Tol, C.; Rastogi, A. Impact of climate change-induced alterations in peatland vegetation phenology and composition on carbon balance. Sci. Total Environ. 2022, 827, 154294. [Google Scholar] [CrossRef]
- Dieleman, C.M.; Branfireun, B.A.; McLaughlin, J.W.; Lindo, Z. Climate change drives a shift in peatland ecosystem plant community: Implications for ecosystem function and stability. Glob Chang. Biol. 2015, 21, 388–395. [Google Scholar] [CrossRef] [PubMed]
- McPartland, M.Y.; Montgomery, R.A.; Hanson, P.J.; Phillips, J.R.; Kolka, R.; Palik, B. Vascular plant species response to warming and elevated carbon dioxide in a boreal peatland. Environ. Res. Lett. 2020, 15, 124066. [Google Scholar] [CrossRef]
- Norby, R.J.; Childs, J.; Hanson, P.J.; Warren, J.M. Rapid loss of an ecosystem engineer: Sphagnum decline in an experimentally warmed bog. Ecol. Evol. 2019, 22, 12571–12585. [Google Scholar] [CrossRef] [PubMed]
- Arp, W.J.; Drake, B.G.; Pockman, W.T.; Curtis, P.S.; Whigham, D.F. Interactions between C3 and C4 salt marsh plant species during four years of exposure to elevated atmospheric CO2. Vegetatio 1993, 104–105, 133–143. [Google Scholar] [CrossRef]
- Mitchell, C.P.G.; Branfireun, B.A.; Kolka, R.K. Total mercury and methylmercury dynamics in upland-peatland watersheds during snowmelt. Biogeochemistry 2008, 90, 225–241. [Google Scholar] [CrossRef]
- Kreyling, J. The ecological importance of winter in temperate, boreal, and Arctic ecosystems in times of climate change. In Progress in Botany; Cánovas, F.M., Ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2020; Volume 81, pp. 377–399. [Google Scholar] [CrossRef]
- Friesen, H.; Slesak, R.A.; Karwan, D.L.; Kolka, R.K. Effects of snow and climate on soil temperature and frost development in forested peatlands in Minnesota, USA. Geoderma 2021, 394, 115015. [Google Scholar] [CrossRef]
- Henry, H.A.L. Climate change and soil freezing dynamics: Historical trends and projected changes. Clim. Chang. 2008, 87, 421–434. [Google Scholar] [CrossRef]
- Dai, A. Increasing drought under global warming in observations and models. Nat. Clim. Chang. 2013, 3, 52–58. [Google Scholar] [CrossRef]
- Lehmann, J.; Coumou, D.; Frieler, K. Increased record-breaking precipitation events under global warming. Clim. Chang. 2015, 132, 501–515. [Google Scholar] [CrossRef]
- Jeremiason, J.D.; Engstrom, D.R.; Swain, E.B.; Nater, E.A.; Johnson, B.M.; Almendinger, J.A.; Monson, B.A.; Kolka, R.K. Sulfate addition increases methylmercury export in an experimental wetland. Environ. Sci. Technol. 2006, 40, 3800–3806. [Google Scholar] [CrossRef]
- Golden, H.E.; Knightes, C.D.; Conrads, P.A.; Feaster, T.D.; Davis, G.M.; Benedict, S.T.; Bradley, P.M. Climate change and watershed mercury export: A multiple projection and model analysis. Environ. Toxicol. Chem. 2013, 32, 2165–2174. [Google Scholar] [CrossRef] [PubMed]
- Hararuk, O.; Obrist, D.; Luo, Y. Modelling the sensitivity of soil mercury storage to climate-induced changes in soil carbon pools. Biogeosciences 2013, 10, 2393–2407. [Google Scholar] [CrossRef]


{kind=link}
| Name | Location | Manipulation/Source | Hg Data | Time Period |
|---|---|---|---|---|
| MEF | Minnesota | Long-term records | Streamwater | 2002–2017 [19] |
| SPRUCE | Minnesota | Warming and eCO2 | Pore water, passive gas | 2017–2019 [26,27,28] |
| BRACE | Canada | Warming | Pore water | 2017–2019 [21] |
| PEATCOSM | Michigan * | Hydrology and plants | Pore water, dynamic gas | 2012–2014 [29,30] |
| Jennie’s Bog | Minnesota | Freeze/thaw | Pore water | 2015 [23] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kolka, R.; Pierce, C.; Garrioch, I.; Behrens, K.; Toner, B.M. Review of the Influence of Climate Change on the Hydrologic Cycling and Gaseous Fluxes of Mercury in Boreal Peatlands: Implications for Restoration. Water 2024, 16, 1154. https://doi.org/10.3390/w16081154
Kolka R, Pierce C, Garrioch I, Behrens K, Toner BM. Review of the Influence of Climate Change on the Hydrologic Cycling and Gaseous Fluxes of Mercury in Boreal Peatlands: Implications for Restoration. Water. 2024; 16(8):1154. https://doi.org/10.3390/w16081154
Chicago/Turabian StyleKolka, Randy, Caroline Pierce, Isabella Garrioch, Kevin Behrens, and Brandy M. Toner. 2024. "Review of the Influence of Climate Change on the Hydrologic Cycling and Gaseous Fluxes of Mercury in Boreal Peatlands: Implications for Restoration" Water 16, no. 8: 1154. https://doi.org/10.3390/w16081154
APA StyleKolka, R., Pierce, C., Garrioch, I., Behrens, K., & Toner, B. M. (2024). Review of the Influence of Climate Change on the Hydrologic Cycling and Gaseous Fluxes of Mercury in Boreal Peatlands: Implications for Restoration. Water, 16(8), 1154. https://doi.org/10.3390/w16081154