Possible Increase of Vegetation Exposure to Spring Frost under Climate Change in Switzerland


Abstract
1. Introduction
2. Data and Methods
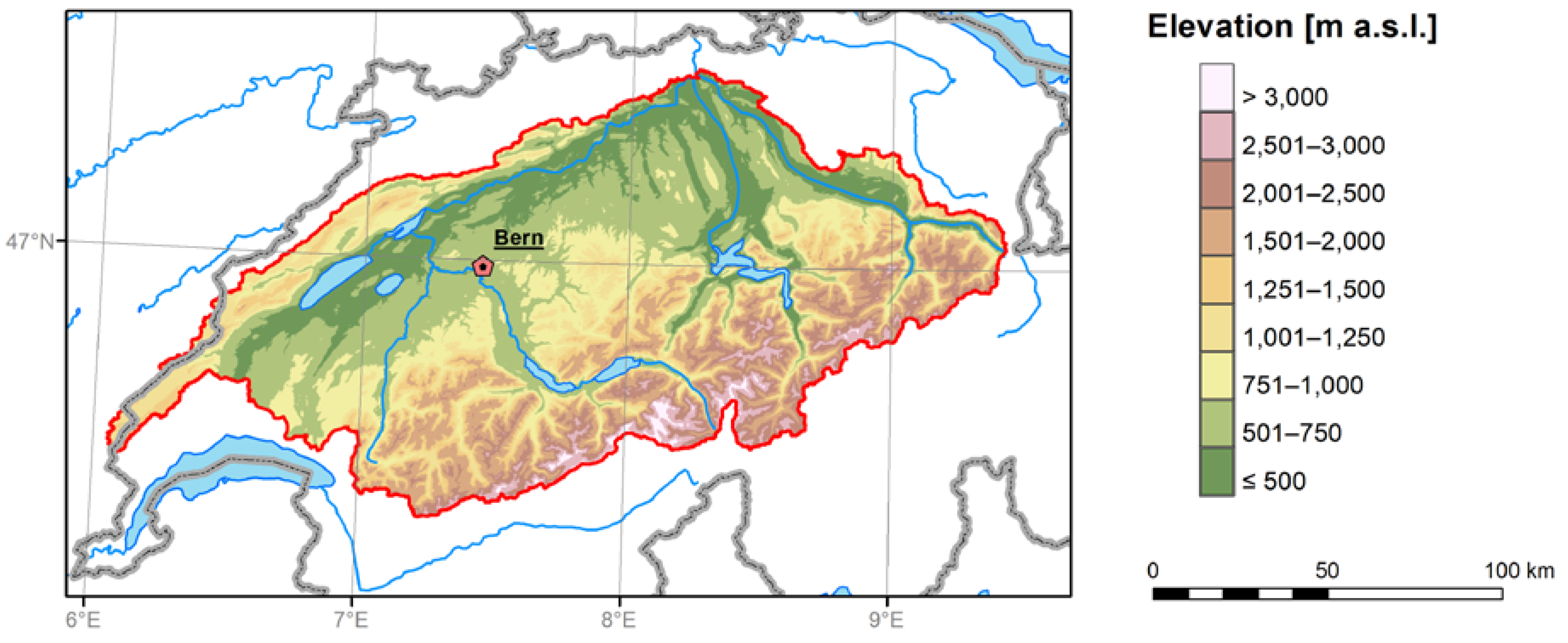
2.1. Study Domain and EXAR Data Set
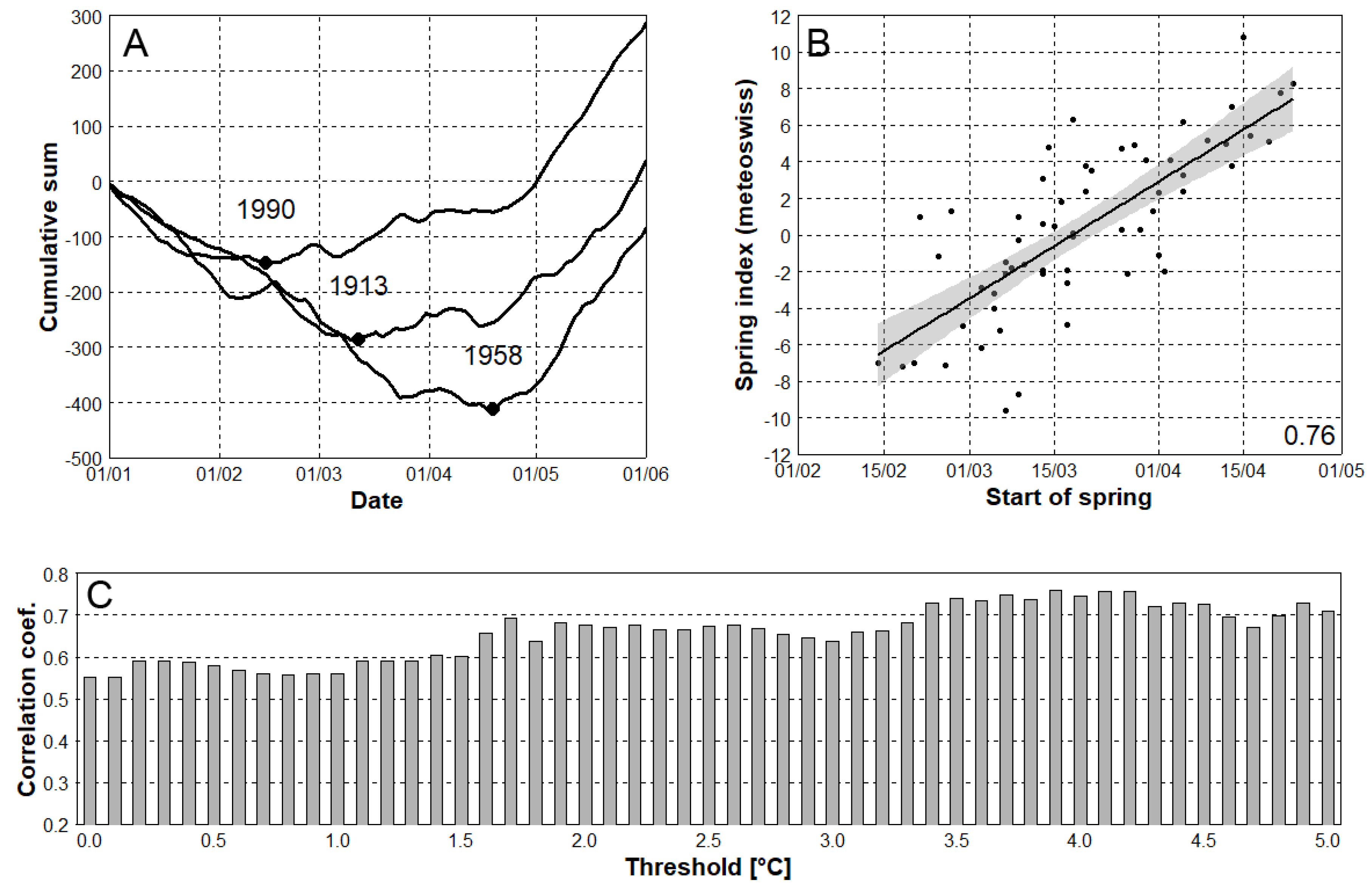
2.2. Definition of Spring Frost
2.3. Other Characteristics and Statistical Testing
3. Results
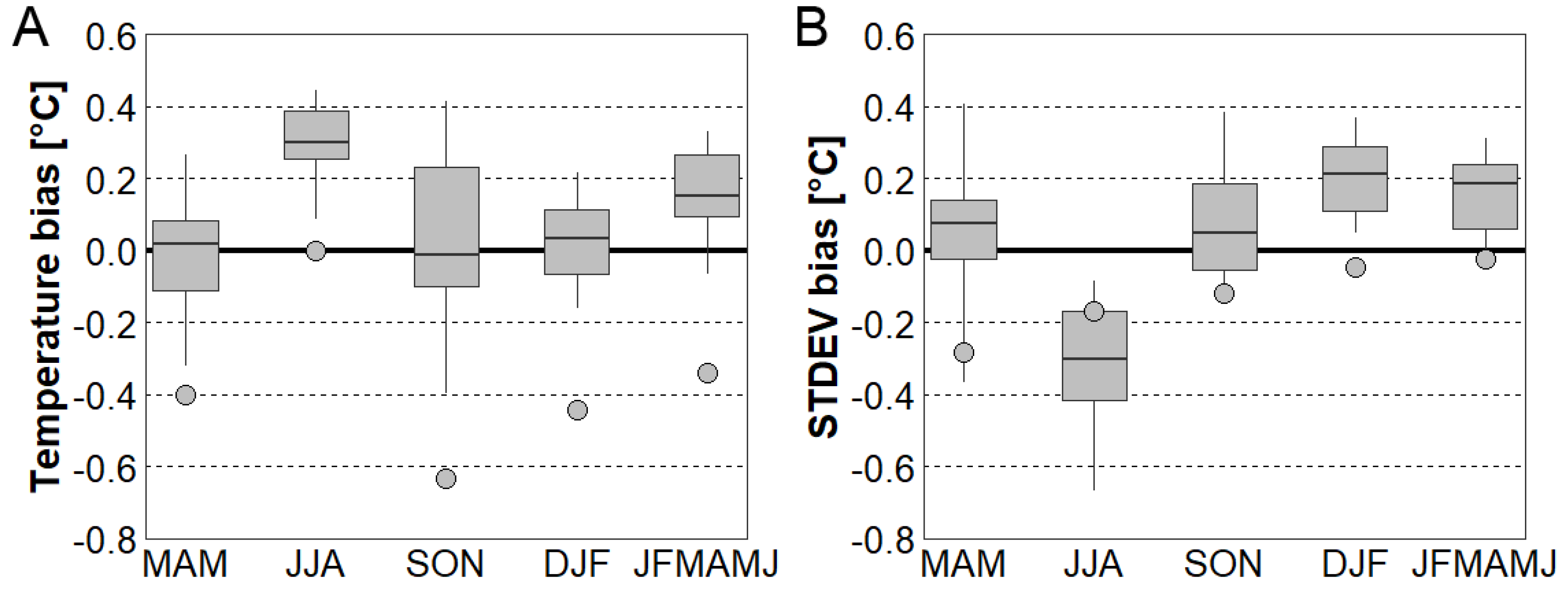
3.1. Evaluation of EXAR Dataset
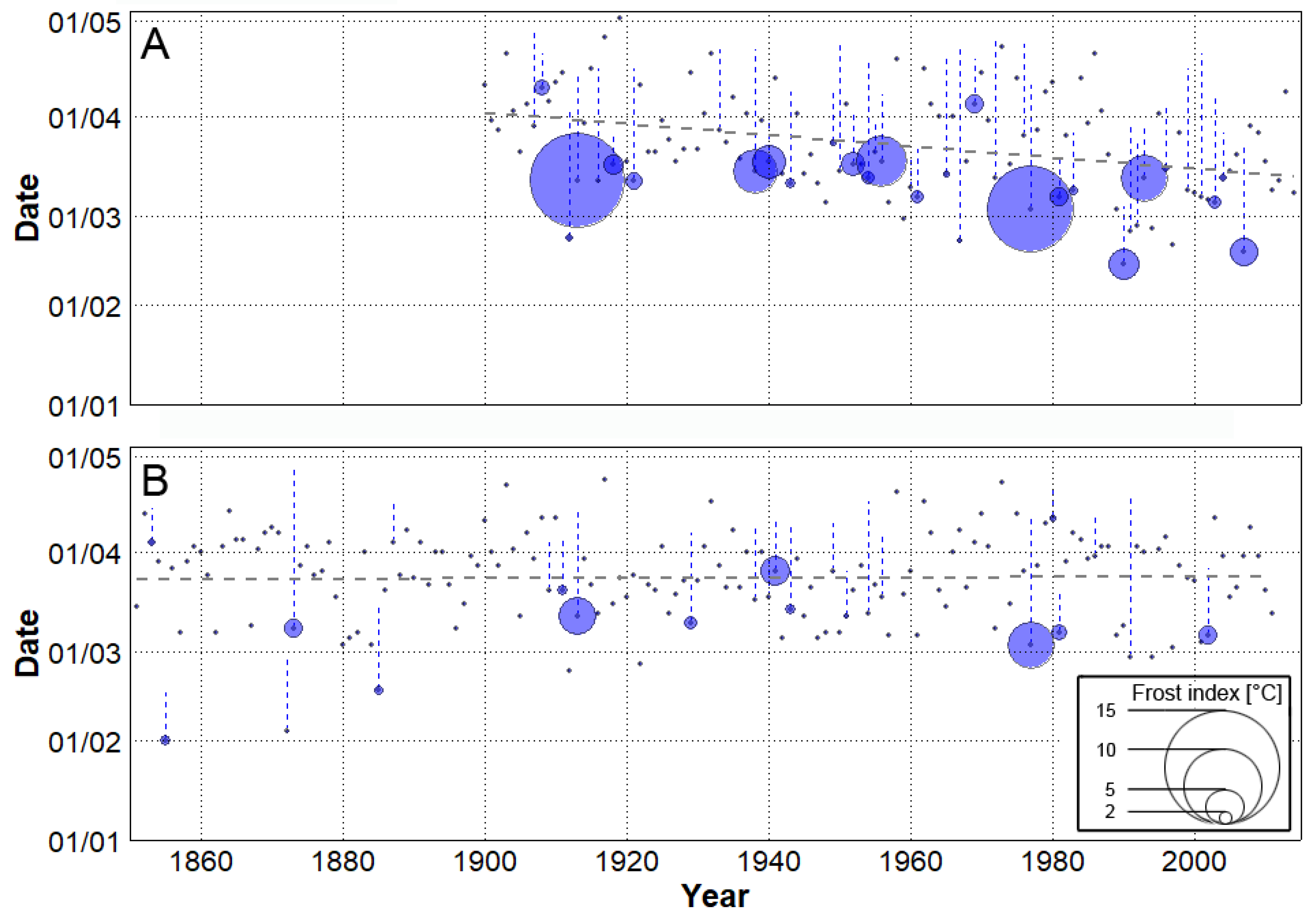
3.2. Spring Frost in the Historical Climate
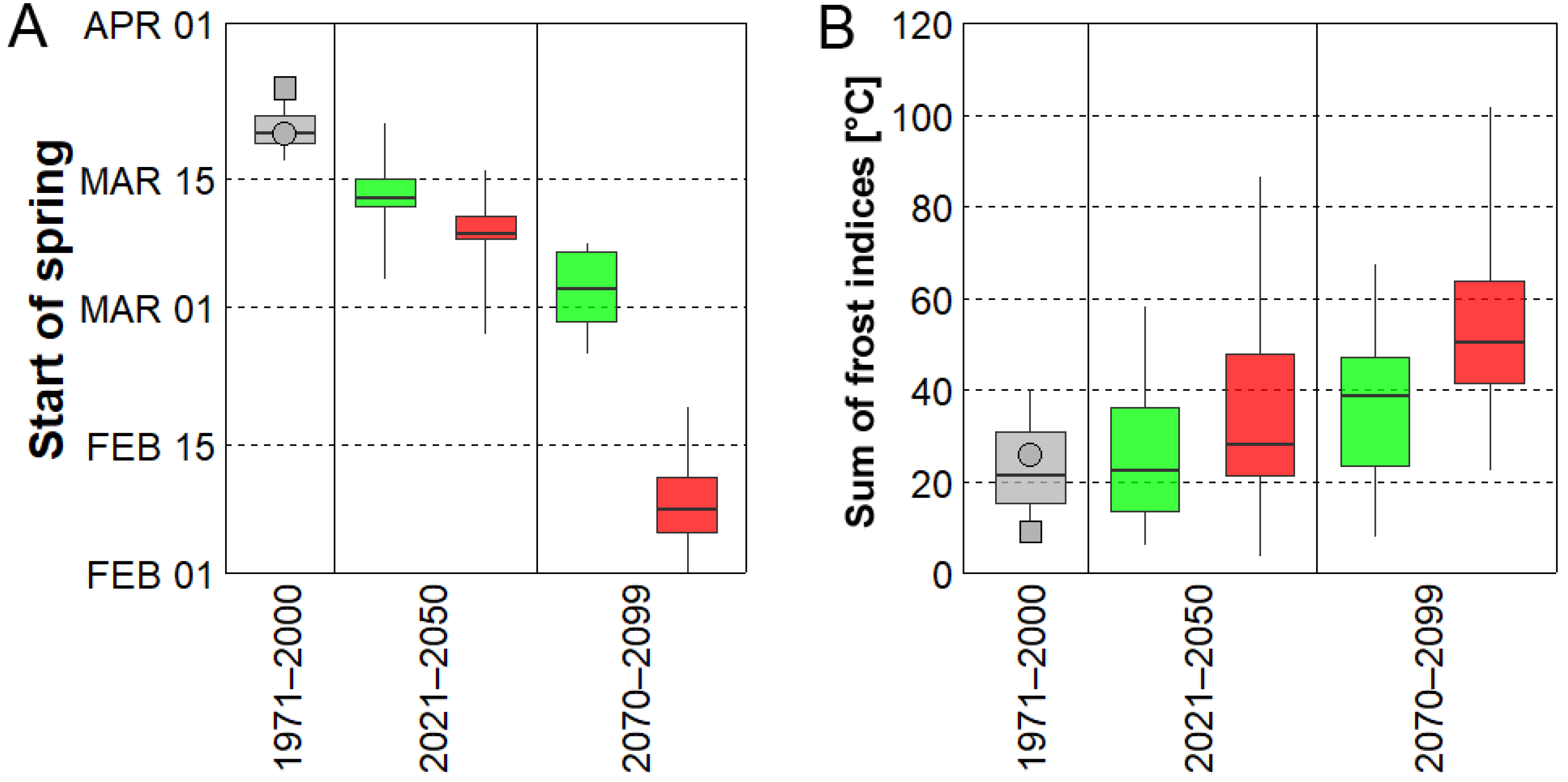
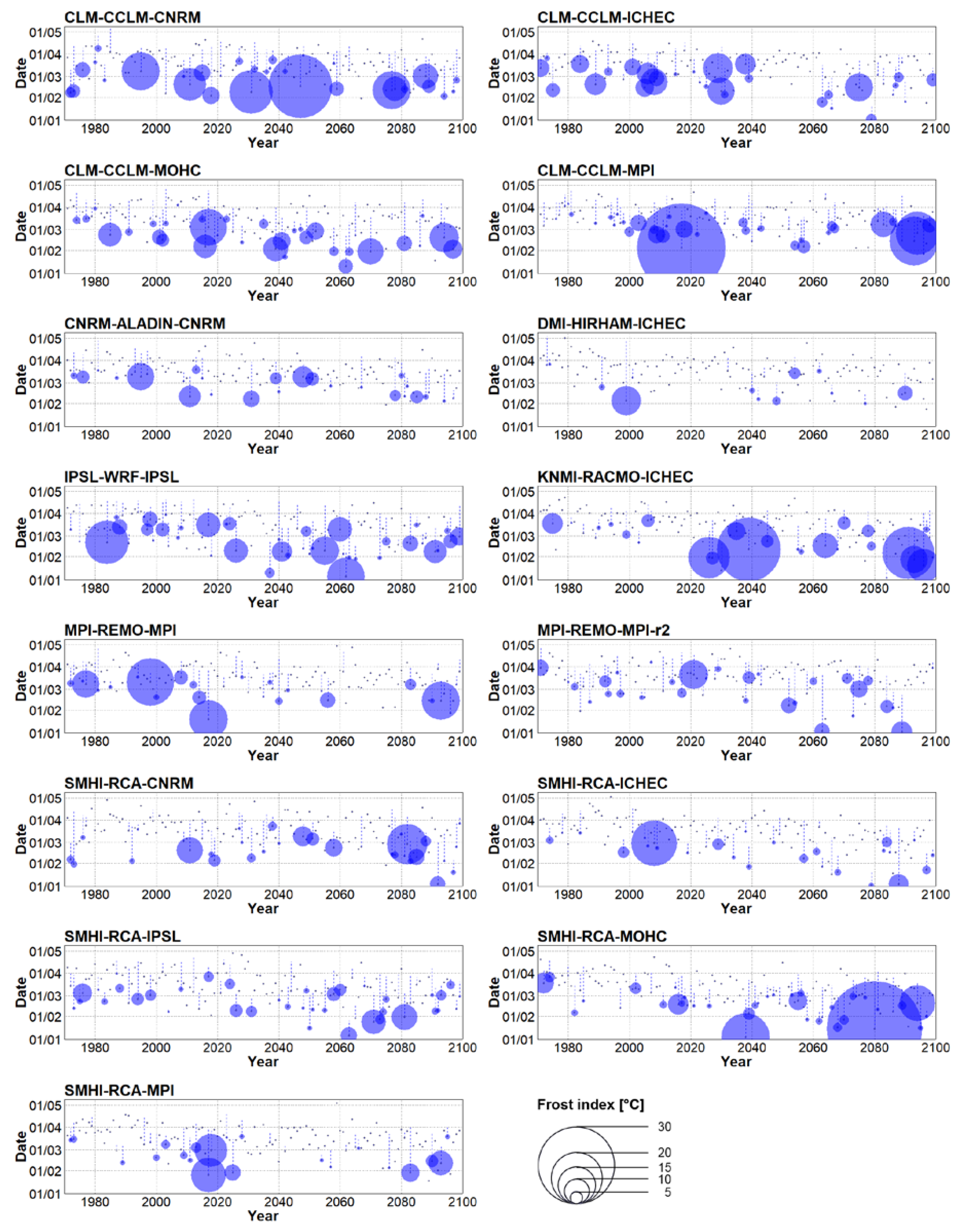
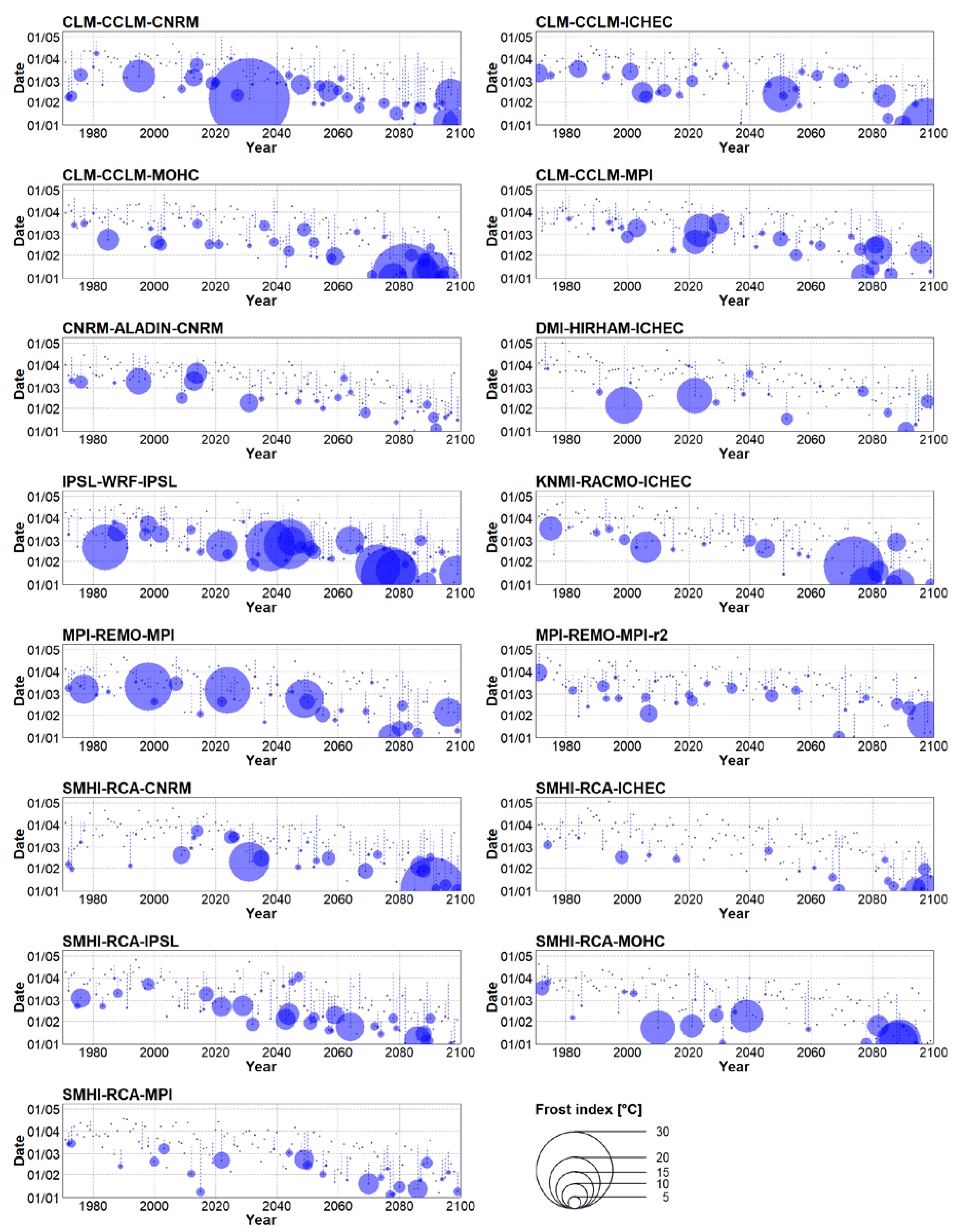
3.3. Future Changes of Spring Frost Risk
4. Discussion
5. Conclusions
- The methodology for estimating an onset of spring plant growth solely from temperature data was developed and tested. The estimated dates were well correlated (correlation coefficient = 0.76) with the Swiss spring index that was calculated from actual phenological observations.
- Significant (at 1% level) advancement of spring start was found in the observed data, with a trend of 1.8 days/decade in the 1900–2014 period. The 20CRv2 reanalysis failed to reproduce this trend.
- In the observed data, springs with early start were significantly (at 1% level) more prone to experience frost events compared to spring that began later. This relationship was, in general, simulated also in a future climate, but in some RCMs, a substantial spring advancement was not linked to a large sum of the yearly frost indices and vice versa.
- Considering the 2021–2050 period, spring is projected to start 8 or 12 days earlier (depending on concentration scenario). This advancement is linked to larger sums of the yearly frost indices compared to historical climate, approximately by a factor of 1.3 or 1.6, respectively.
- Major differences between concentration scenarios were found at the end of the 21st century (2070–2099). The earliest starts of spring and the largest values of the sum of yearly frost indices were simulated under RCP 8.5, in which the mean date of spring start was February 9 (about 6 weeks earlier) and the sum of the yearly frost indices was larger by a factor of 2.9 compared to historical climate.
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kollas, C.; Körner, C.; Randin, C.F. Spring frost and growing season length co-control the cold range limits of broad-leaved trees. J. Biogeogr. 2014, 41, 773–783. [Google Scholar] [CrossRef]
- Meier, M.; Fuhrer, J.; Holzkämper, A. Changing risk of spring frost damage in grapevines due to climate change? A case study in the Swiss Rhone Valley. Int. J. Biometeorol. 2018, 62, 991–1002. [Google Scholar] [CrossRef] [PubMed]
- Vitasse, Y.; Rebetez, M. Unprecedented risk of spring frost damage in Switzerland and Germany in 2017. Clim. Chang. 2018, 149, 233–246. [Google Scholar] [CrossRef]
- Augspurger, C.K. Reconstructing patterns of temperature, phenology, and frost damage over 124 years: Spring damage risk is increasing. Ecology 2013, 94, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.-J.; Ho, C.-H.; Gim, H.-J.; Brown, M.E. Phenology shifts at start vs. end of growing season in temperate vegetation over the Northern Hemisphere for the period 1982–2008. Glob. Chang. Biol. 2011, 17, 2385–2399. [Google Scholar] [CrossRef]
- Menzel, A.; Sparks, T.H.; Estrella, N.; Koch, E.; Aasa, A.; Ahas, R.; Alm-Kübler, K.; Bissolli, P.; Braslavská, O.; Briede, A.; et al. European phenological response to climate change matches the warming pattern. Glob. Chang. Biol. 2006, 12, 1969–1976. [Google Scholar] [CrossRef]
- Lavalle, C.; Micale, F.; Houston, T.D.; Camia, A.; Hiederer, R.; Lazar, C.; Costanza, C.; Amatulli, G.; Genovese, G. Climate change in Europe. 3. Impact on agriculture and forestry. A review. Agron. Sustain. Dev. 2009, 29, 433–446. [Google Scholar] [CrossRef]
- Shen, M.; Cong, N.; Cao, R. Temperature sensitivity as an explanation of the latitudinal pattern of green-up date trend in Northern Hemisphere vegetation during 1982–2008. Int. J. Climatol. 2015, 35, 3707–3712. [Google Scholar] [CrossRef]
- Kolářová, E.; Nekovář, J.; Adamík, P. Long-term temporal changes in central European tree phenology (1946−2010) confirm the recent extension of growing seasons. Int. J. Biometeorol. 2014, 58, 1739–1748. [Google Scholar] [CrossRef]
- Legave, J.M.; Blanke, M.; Christen, D.; Giovannini, D.; Mathieu, V.; Oger, R. A comprehensive overview of the spatial and temporal variability of apple bud dormancy release and blooming phenology in Western Europe. Int. J. Biometeorol. 2013, 57, 317–331. [Google Scholar] [CrossRef]
- Wypych, A.; Ustrnul, Z.; Sulikowska, A.; Chmielewski, F.-M.; Bochenek, B. Spatial and temporal variability of the frost-free season in Central Europe and its circulation background. Int. J. Climatol. 2017, 37, 3340–3352. [Google Scholar] [CrossRef]
- Bigler, C.; Bugmann, H. Climate-induced shifts in leaf unfolding and frost risk of European trees and shrubs. Sci. Rep. 2018, 9865. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Piao, S.; Janssens, I.M.; Fu, Y.; Peng, S.; Lian, X.; Ciais, P.; Myneni, R.B.; Penuelas, J.; Wang, T. Extension of the growing season increases vegetation exposure to frost. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Kimball, J.S.; Didan, K.; Henebry, G.M. Response of vegetation growth and productivity to spring climate indicators in the conterminous United States derived from satellite remote sensing data fusion. Agric. For. Meteorol. 2014, 194, 132–143. [Google Scholar] [CrossRef]
- Vitasse, Y.; Schneider, L.; Rixen, C.; Christen, D.; Rebetez, M. Increase in the risk of exposure of forest and fruit trees to spring frosts at higher elevations in Switzerland over the last four decades. Agric. For. Meteorol. 2018, 248, 60–69. [Google Scholar] [CrossRef]
- Bennie, J.; Kubin, E.; Wiltshire, A.; Huntley, B.; Baxter, R. Predicting spatial and temporal patterns of bud-burst and spring frost risk in north-west Europe: The implications of local adaptation to climate. Glob. Chang. Biol. 2010, 16, 1503–1514. [Google Scholar] [CrossRef]
- Molitor, D.; Caffarra, A.; Sinigoj, P.; Pertot, I.; Hoffmann, L.; Junk, J. Late frost damage risk for viticulture under future climate conditions: A case study for the Luxembourgish winegrowing region. Aust. J. Grape Wine Res. 2014, 20, 160–168. [Google Scholar] [CrossRef]
- Leolini, L.; Moriondo, M.; Fila, G.; Costafreda-Aumedes, S.; Ferrise, R.; Bindi, M. Late spring frost impacts on future grapevine distribution in Europe. Field Crop. Res. 2018, 222, 197–208. [Google Scholar] [CrossRef]
- Mosedale, J.R.; Wilson, R.J.; Maclean, I.M. Climate Change and Crop Exposure to Adverse Weather: Changes to Frost Risk and Grapevine Flowering Conditions. PLoS ONE 2015. [Google Scholar] [CrossRef]
- Spring Index. Available online: https://www.meteoswiss.admin.ch/home/climate/climate-change-in-switzerland/vegetation-development/spring-index.html (accessed on 12 October 2019).
- Brönnimann, S.; Rajczak, J.; Fischer, E.M.; Raible, C.C.; Rohrer, M.; Schär, C. Changing seasonality of moderate and extreme precipitation events in the Alps. Nat. Hazard. Earth Syst. 2018, 18, 2047–2056. [Google Scholar] [CrossRef]
- Rajczak, J.; Brönnimann, S.; Fischer, E.M.; Raible, C.C.; Rohrer, M.; Schär, C. Daily Precipitation and Temperature Time Series from Multiple Climate Model Simulations for the Aare River Catchment (Switzerland). Available online: https://www.pangaea.de (accessed on 12 October 2019). [CrossRef]
- Begert, M.; Schlegel, T.; Kirchhofer, W. Homogeneous Temperature and Precipitation Series of Switzerland from 1864 to 2000. Int. J. Climatol. 2005, 25, 65–80. [Google Scholar] [CrossRef]
- Maraun, D. Bias Correction, Quantile Mapping, and Downscaling: Revisiting the Inflation Issue. J. Clim. 2013, 26, 2137–2143. [Google Scholar] [CrossRef]
- Rajczak, J.; Schär, C. Projections of Future Precipitation Extremes Over Europe: A Multimodel Assessment of Climate Simulations. J. Geophys. Res. Atmos. 2017, 122, 10–773. [Google Scholar] [CrossRef]
- Compo, G.P.; Whitaker, J.S.; Sardeshmukh, P.D.; Matsui, N.; Allan, R.J.; Yin, X.; Gleason, B.E.; Vose, R.S.; Rutledge, G.; Bessemoulin, P.; et al. The Twentieth Century Reanalysis Project. Q. J. R. Meteorol. Soc. 2011, 137, 1–28. [Google Scholar] [CrossRef]
- Jacob, D.; Petersen, J.; Eggert, B.; Alias, A.; Christensen, O.B.; Bouwer, L.M.; Braun, A.; Colette, A.; Déqué, M.; Georgievski, G.; et al. EURO-CORDEX: New high-resolution climate change projections for European impact research. Reg. Environ. Chang. 2014, 14, 563–578. [Google Scholar] [CrossRef]
- Taylor, K.E.; Stouffer, R.J.; Meehl, G.A. An overview of CMIP5 and the experiment design. Bull. Am. Meteorol. Soc. 2012, 93, 485–498. [Google Scholar] [CrossRef]
- Thomson, A.M.; Calvin, K.V.; Smith, S.J.; Kyle, G.P.; Volke, A.; Patel, P.; Delgado-Arias, S.; Bond-Lamberty, B.; Wise, M.A.; Clarke, L.E.; et al. RCP 4.5: A pathway for stabilization of radiative forcing by 2100. Clim. Chang. 2011, 109, 77–94. [Google Scholar] [CrossRef]
- Riahi, K.; Rao, S.; Krey, V.; Cho, C.; Chirkov, V.; Fischer, G.; Nakicenovic, N.; Rafaj, P. RCP 8.5—A scenario of comparatively high greenhouse gas emissions. Clim. Chang. 2011, 109, 33–57. [Google Scholar] [CrossRef]
- Kotlarski, S.; Keuler, K.; Christensen, O.B.; Colette, A.; Déqué, M.; Gobiet, A.; Goergen, K.; Jacob, D.; Lüthi, D.; van Meijgaard, E.; et al. Regional climate modeling on European scales: A joint standard evaluation of the EURO-CORDEX RCM ensemble. Geosci. Model. Dev. 2014, 7, 1297–1333. [Google Scholar] [CrossRef]
- McMaster, G.S.; Wilhelm, W.W. Growing degree-days: One equation, two interpretations. Agric. For. Meteorol. 1997, 87, 291–300. [Google Scholar] [CrossRef]
- Campbell, G.S.; Norman, J. An Introduction to Environmental Biophysics; Springer: New York, NY, USA, 1998; p. 286. ISBN 978-0-387-94937-6. [Google Scholar]
- Brugnara, Y.; Auchmann, R.; Rutishauser, T.; Gehrig, R.; Pietragalla, B.; Begert, M.; Sigg, C.; Knechtl, V.; Konzelmann, T.; Calpini, B.; et al. Homogeneity assessment of phenological records from the Swiss Phenology Network. Int. J. Biometeorol. 2020, 64, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Venn, S.E.; Green, K. Evergreen alpine shrubs have high freezing resistance in spring, irrespective of snowmelt timing and exposure to frost: An investigation from the Snowy Mountains, Australia. Plant Ecol. 2017, 219, 209–216. [Google Scholar] [CrossRef]
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agric. For. Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- Eccel, E.; Rea, R.; Caffarra, A.; Crisci, A. Risk of spring frost to apple production under future climate scenarios: The role of phenological acclimation. Int. J. Biometeorol. 2009, 53, 273–286. [Google Scholar] [CrossRef]
- Wang, T.; Ottlé, C.; Peng, S.; Janssens, I.A.; Lin, X.; Poulter, B.; Yue, C.; Ciais, P. The influence of local spring temperature variance on temperature sensitivity of spring phenology. Glob. Chang. Biol. 2014, 20, 1473–1480. [Google Scholar] [CrossRef] [PubMed]
- Breitenmoser, P.; Brönnimann, S.; Frank, D. Forward modelling of tree-ring width and comparison with a global network of tree-ring chronologies. Clim. Past 2014, 10, 437–449. [Google Scholar] [CrossRef]
- Kleiber, W.; Katz, R.W.; Rajagopalan, B. Daily minimum and maximum temperature simulation over complex terrain. Ann. Appl. Stat. 2013, 7, 588–612. [Google Scholar] [CrossRef]
- Giaccone, E.; Luoto, M.; Vittoz, P.; Guisan, A.; Mariéthoz, G.; Lambiel, C. Influence of microclimate and geomorphological factors on alpine vegetation in the Western Swiss Alps. Earth Surf. Process. Landf. 2019, 44, 3093–3107. [Google Scholar] [CrossRef]
- Cattiaux, J.; Yiou, P.; Vautard, R. Dynamics of future seasonal temperature trends and extremes in Europe: A multi-model analysis from CMIP3. Clim. Dynam. 2012, 38, 1949–1964. [Google Scholar] [CrossRef]
- Ustrnul, Z.; Wypych, A.; Winkler, J.A.; Czekierda, D. Late Spring Freezes in Poland in Relation to Atmospheric Circulation. QuaGeo 2014, 33, 165–172. [Google Scholar] [CrossRef][Green Version]
- Yao, Y.; Luo, D.; Dai, A.; Simmonds, I. Increased quasi stationarity and persistence of winter Ural blocking and Eurasian extreme cold events in response to arctic warming. Part I: Insights from Observational Analyses. J. Clim. 2017, 30, 3569–3587. [Google Scholar] [CrossRef]
- Masato, G. Winter and Summer Northern Hemisphere Blocking in CMIP5 Models. J. Clim. 2013, 26, 7044–7059. [Google Scholar] [CrossRef]
- Dunn-Sigouin, E.; Son, S.-W. Northern Hemisphere blocking frequency and duration in the CMIP5 models. J. Geophys. Res. Atmos. 2013, 118, 1179–1188. [Google Scholar] [CrossRef]
- Scaife, A.A.; Woollings, T.; Knight, J.; Martin, G.; Hinton, T. Atmospheric Blocking and Mean Biases in Climate Models. J. Clim. 2010, 23, 6143–6152. [Google Scholar] [CrossRef]
- Lhotka, O.; Kyselý, J. Circulation-Conditioned Wintertime Temperature Bias in EURO-CORDEX Regional Climate Models over Central Europe. J. Geophys. Res. Atmos. 2018, 16, 8661–8673. [Google Scholar] [CrossRef]
- Lorenz, P.; Jacob, D. Validation of temperature trends in the ENSEMBLES regional climate model runs driven by ERA40. Clim. Res. 2010, 44, 167–177. [Google Scholar] [CrossRef]
- Kalnay, E.; Kanamitsu, M.; Kistler, R.; Collins, W.; Deaven, D.; Gandin, L.; Iredell, M.; Saha, S.; White, G.; Woollen, J.; et al. The NCEP/NCAR 40-year reanalysis project. Bull. Am. Meteorol. Soc. 1996, 77, 437–471. [Google Scholar] [CrossRef]
- Cong, N.; Shen, M.; Piao, S.; Chen, X.; An, S.; Yang, W.; Fu, Y.H.; Meng, F.; Wang, T. Little change in heat requirement for vegetation green-up on the Tibetan Plateau over the warming period of 1998–2012. Agric. For. Meteorol. 2017, 232, 650–658. [Google Scholar] [CrossRef]
- Asse, D.; Chuine, I.; Vitasse, Y.; Yocco, N.G.; Delpierre, N. Warmer winters reduce the advance of tree spring phenology induced by warmer springs in the Alps. Agric. For. Meteorol. 2018, 252, 220–230. [Google Scholar] [CrossRef]
- Güsewell, S.; Furrer, R.; Gehrig, R.; Pietragalla, B. Changes in temperature sensitivity of spring phenology with recent climate warming in Switzerland are related to shifts of the preseason. Glob. Chang. Biol. 2017, 23, 5189–5202. [Google Scholar] [CrossRef]
- Körner, C.; Basler, D. Phenology under Global Warming. Science 2010, 327, 1461–1462. [Google Scholar] [CrossRef] [PubMed]
- Migliavacca, M.; Sonnentag, O.; Keenan, T.F.; Cescatti, A.; O’keefe, J.; Richardson, A.D. On the uncertainty of phenological responses to climate change, and implications for a terrestrial biosphere model. Biogeosciences 2012, 9, 2063–2083. [Google Scholar] [CrossRef]
- Chen, X.; An, S.; Inouye, D.W.; Schwartz, M.D. Temperature and snowfall trigger alpine vegetation green-up on the world’s roof. Glob. Chang. Biol. 2015, 21, 3635–3646. [Google Scholar] [CrossRef] [PubMed]
- Young, S.L. As Climate Shifts, So Do Pests: A National Forum and Assessment. Bull. Ecol. Soc. Am. 2017, 98, 165–172. [Google Scholar] [CrossRef]
- CH2018. CH2018—Climate Scenarios for Switzerland, Technical Report; National Centre for Climate Services: Zürich, Switzerland, 2018; p. 271. ISBN 978-3-9525031-4-0. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Institute | Acronym | RCM | GCM |
|---|---|---|---|
| Climate Limited-Area Modelling Community | CLM | CCLM | CNRM |
| ICHEC | |||
| MOHC | |||
| MPI | |||
| National Centre for Meteorological Research | CNRM | ALADIN | CNRM |
| Danish Meteorological Institute | DMI | HIRHAM | ICHEC |
| Institute Pierre Simon Laplace | IPSL | WRF | IPSL |
| Royal Netherlands Meteorological Institute | KNMI | RACMO | ICHEC |
| Max Planck Institute for Meteorology | MPI | REMO | MPI* |
| Swedish Meteorological and Hydrological Institute | SMHI | RCA | CNRM |
| HadGEM | |||
| ICHEC | |||
| IPSL | |||
| MPI |
| Historical | RCP 4.5 | RCP 8.5 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1971–2000 | 2021–2050 | 2070–2099 | 2021–2050 | 2070–2099 | ||||||
| S | If [°C] | S | If [°C] | S | If [°C] | S | If [°C] | S | If [°C] | |
| CLM-CCLM-CNRM | 19/03 | 36.3 | 14/03 | 58.1 (1.6) | 07/03 | 49.6 (1.4) | 11/03 | 54.1 (1.5) | 12/02 | 63.8 (1.8) |
| CLM-CCLM-ICHEC | 19/03 | 33.9 | 12/03 | 40.2 (1.2) | 01/03 | 28.8 (0.8) | 11/03 | 27.9 (0.8) | 18/02 | 50.8 (1.5) |
| CLM-CCLM-MOHC | 20/03 | 22.0 | 07/03 | 35.8 (1.6) | 04/03 | 38.8 (1.8) | 06/03 | 24.2 (1.1) | 01/02 | 101.6 (4.6) |
| CLM-CCLM-MPI | 20/03 | 15.0 | 12/03 | 15.7 (1.0) | 08/03 | 57.4 (3.8) | 10/03 | 46.8 (3.1) | 08/02 | 59.7 (4.0) |
| CNRM-ALADIN-CNRM | 21/03 | 21.1 | 16/03 | 21.1 (1.0) | 03/03 | 17.7 (0.8) | 12/03 | 15.1 (0.7) | 08/02 | 25.6 (1.2) |
| DMI-HIRHAM-ICHEC | 24/03 | 15.4 | 14/03 | 6.1 (0.4) | 07/03 | 7.9 (0.5) | 09/03 | 23.1 (1.5) | 19/02 | 22.5 (1.5) |
| IPSL-WRF-IPSL | 20/03 | 40.1 | 12/03 | 36.5 (0.9) | 03/03 | 40.1 (1.0) | 09/03 | 86.5 (2.2) | 06/02 | 98.4 (2.5) |
| KNMI-RACMO-ICHEC | 23/03 | 16.3 | 09/03 | 58.2 (3.6) | 25/02 | 60.3 (3.7) | 09/03 | 16.3 (1.0) | 06/02 | 75.8 (4.7) |
| MPI-REMO-MPI | 19/03 | 39.5 | 16/03 | 8.7 (0.2) | 07/03 | 22.5 (0.6) | 08/03 | 50.4 (1.3) | 11/02 | 42.7 (1.1) |
| MPI-REMO-MPI-r2 | 17/03 | 23.9 | 21/03 | 23.8 (1.0) | 08/03 | 30.5 (1.3) | 16/03 | 19.6 (0.8) | 17/02 | 33.4 (1.4) |
| SMHI-RCA-CNRM | 18/03 | 10.6 | 18/03 | 19.5 (1.8) | 29/02 | 44.4 (4.2) | 09/03 | 37.5 (3.5) | 08/02 | 63.9 (6.0) |
| SMHI-RCA-ICHEC | 25/03 | 9.0 | 12/03 | 11.1 (1.2) | 25/02 | 21.6 (2.4) | 11/03 | 3.4 (0.4) | 10/02 | 40.3 (4.5) |
| SMHI-RCA-IPSL | 19/03 | 27.6 | 13/03 | 22.2 (0.8) | 28/02 | 43.6 (1.6) | 08/03 | 49.1 (1.8) | 02/02 | 43.0 (1.6) |
| SMHI-RCA-MOHC | 21/03 | 16.6 | 04/03 | 33.1 (2.0) | 25/02 | 67.5 (4.1) | 27/02 | 36.1 (2.2) | 02/02 | 50.2 (3.0) |
| SMHI-RCA-MPI | 23/03 | 9.3 | 13/03 | 9.5 (1.0) | 04/03 | 24.6 (2.6) | 11/03 | 23.0 (2.5) | 05/02 | 42.6 (4.6) |
| ensemble mean | 21/03 | 22.4 | 13/03 | 26.6 (1.3) | 03/03 | 37.0 (2.0) | 09/03 | 34.2 (1.6) | 09/02 | 54.3 (2.9) |
| station data | 20/03 | 25.8 | ||||||||
| 20CRv2 reanalysis | 25/03 | 9.0 | ||||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lhotka, O.; Brönnimann, S. Possible Increase of Vegetation Exposure to Spring Frost under Climate Change in Switzerland. Atmosphere 2020, 11, 391. https://doi.org/10.3390/atmos11040391
Lhotka O, Brönnimann S. Possible Increase of Vegetation Exposure to Spring Frost under Climate Change in Switzerland. Atmosphere. 2020; 11(4):391. https://doi.org/10.3390/atmos11040391
Chicago/Turabian StyleLhotka, Ondřej, and Stefan Brönnimann. 2020. "Possible Increase of Vegetation Exposure to Spring Frost under Climate Change in Switzerland" Atmosphere 11, no. 4: 391. https://doi.org/10.3390/atmos11040391
APA StyleLhotka, O., & Brönnimann, S. (2020). Possible Increase of Vegetation Exposure to Spring Frost under Climate Change in Switzerland. Atmosphere, 11(4), 391. https://doi.org/10.3390/atmos11040391