Trade in Zambian Edible Orchids—DNA Barcoding Reveals the Use of Unexpected Orchid Taxa for Chikanda

 , , , , , ,
, , , , , ,
Abstract

1. Introduction
2. Materials and Methods
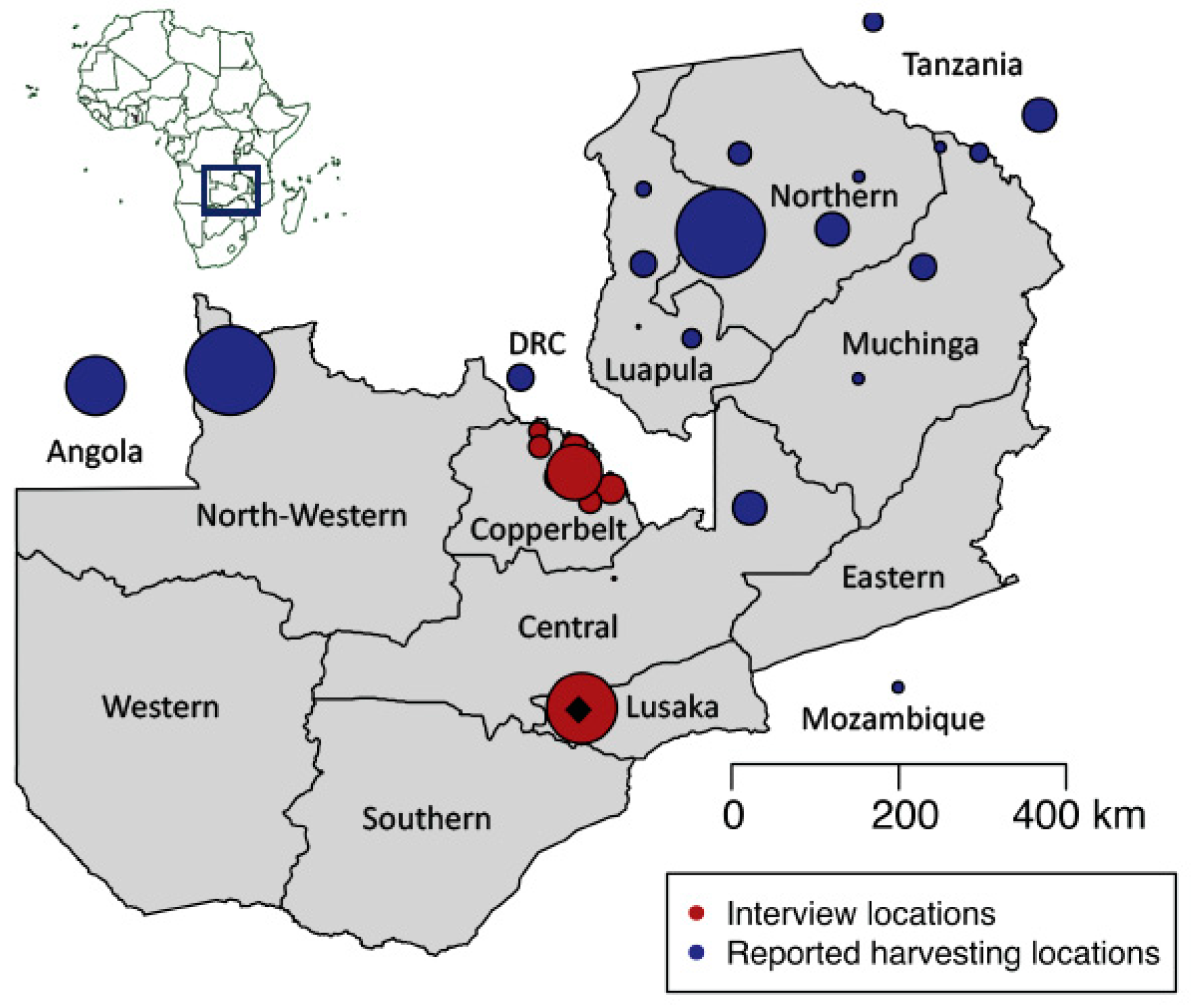
2.1. Interviews and Sample Collection
2.2. Reference Taxon Sampling
2.3. From Sample to Sequence
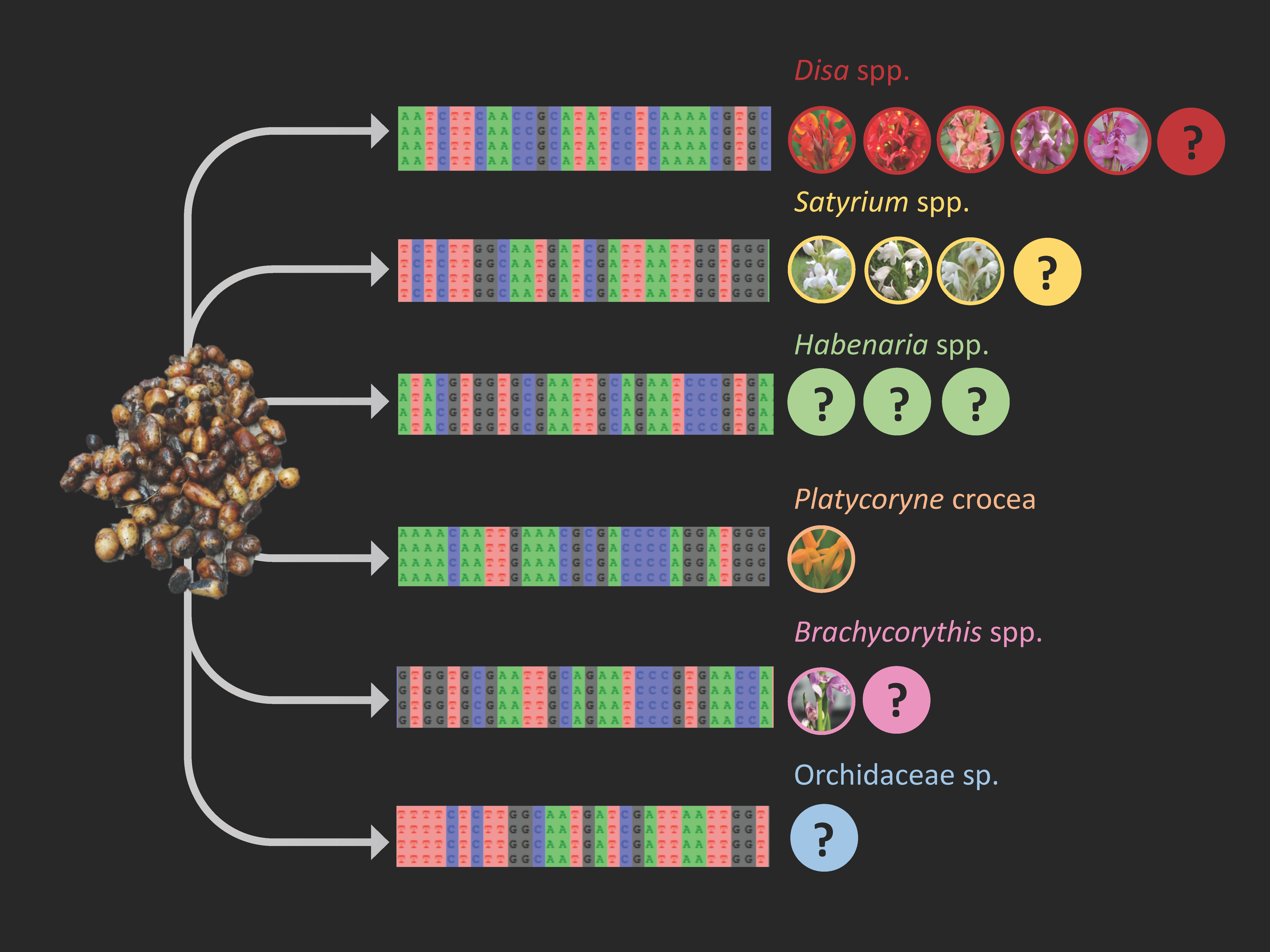
2.4. Phylogenetic Analysis and Species Identification
3. Results
3.1. Market Surveys and Interviews
3.2. Local Classification System
3.3. Chikanda Trade and Availability
3.4. Molecular Identification of Traded Orchids
4. Discussion
4.1. Species Used for Chikanda
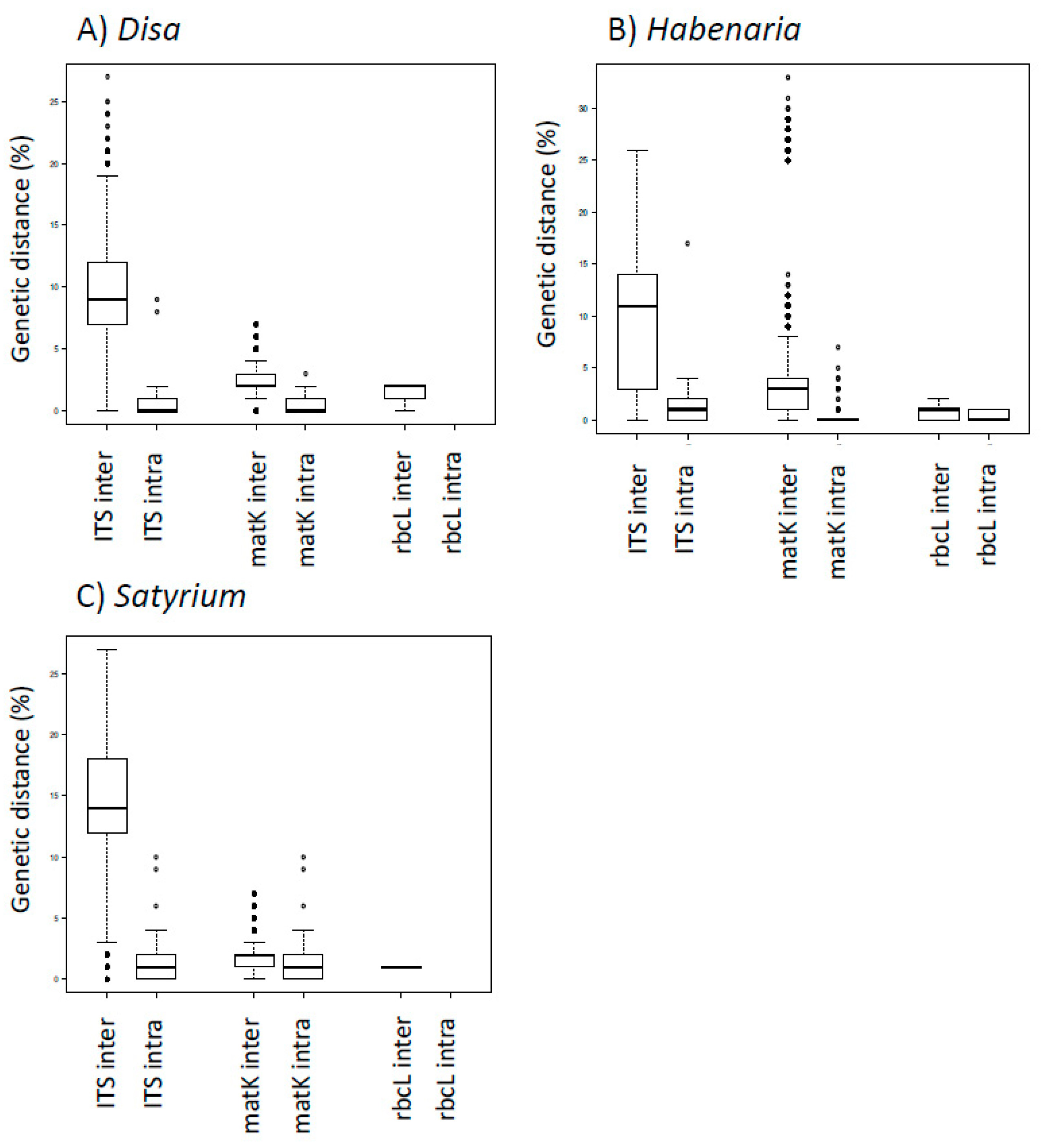
4.2. DNA Barcoding Performance
4.3. Local Versus Scientific Classification of Chikanda
4.4. Orchid Availability and Conservation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kasparek, M.; Grimm, U. European trade in Turkish Salep with special reference to Germany. Econ. Bot. 1999, 53, 396–406. [Google Scholar] [CrossRef]
- Ece Tamer, C.; Karaman, B.; Utku Copur, O. A traditional Turkish beverage: Salep. Food Rev. Int. 2006, 22, 43–50. [Google Scholar] [CrossRef]
- Kreziou, A.; de Boer, H.; Gravendeel, B. Harvesting of salep orchids in north-western Greece continues to threaten natural populations. Oryx 2015, 1–4. [Google Scholar] [CrossRef]
- Ghorbani, A.; Gravendeel, B.; Selliah, S.; Zarré, S.; Boer, H. DNA barcoding of tuberous Orchidoideae: a resource for identification of orchids used in Salep. Mol. Ecol. Resour. 2017, 17, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Davenport, T.R.B.; Ndangalasi, H.J. Orchid Harvest—An Assessment of the Harvesting and Trade of Orchid Tubers across Tanzania’s Southern Highlands; Wildlife Conservation Society: New York, NY, USA, 2001; p. 23. [Google Scholar]
- Bingham, M.G. Chikanda trade in Zambia. Orchid Conserv. News 2004, 4, 22–25. [Google Scholar]
- Veldman, S.; Otieno, J.N.; van Andel, T.; Gravendeel, B.; de Boer, H.J. Efforts urged to tackle thriving illegal orchid trade in Tanzania and Zambia for chikanda production. TRAFFIC Bull. 2014, 26, 47–50. [Google Scholar]
- Richards, A.I. Land, Labour and Diet in Northern Rhodesia: An Economic Study of the Bemba Tribe; Oxford University Press: Oxford, UK, 1939. [Google Scholar]
- Bingham, M.G. Chikanda an unsustainable industry. The Lowdown Magazine. 2007, 4. [Google Scholar]
- Temzie Bites Zambian Food|Chikanda African Ham (Polony) Recipe. 2017. Available online: https://www.youtube.com/watch?v=mDGKx-GOLoA (accessed on 24 October 2018).
- Davenport, T.R.B.; Ndangalasi, H.J. An escalating trade in orchid tubers across Tanzania’s Southern Highlands: assessment, dynamics and conservation implications. Oryx 2003, 37, 55–61. [Google Scholar] [CrossRef]
- Cribb, P.J.; Leedal, G.P. The mountain Flowers of Southern Tanzania: A Field Guide to the Common Flowers; AA Balkema: Rotterdam, NL, USA, 1982. [Google Scholar]
- Nyomora, A.M.S. Distribution and abundance of the edible orchids of the Southern Highlands of Tanzania. Tanzanian J. Sci. 2005, 31, 45–54. [Google Scholar] [CrossRef]
- Mapunda, L.N.D. Edible Orchids in Makete District, the Southern Highlands of Tanzania: Distribution, Population and Status. Master’s Thesis, Swedish Biodiversity Centre, Uppsala Universitet (Sweden), Uppsala, Sweden, 2007. [Google Scholar]
- Challe, J.F.; Price, L.L. Endangered edible orchids and vulnerable gatherers in the context of HIV/AIDS in the Southern Highlands of Tanzania. J. Ethnobiol. Ethnomedicine 2009, 5, 41. [Google Scholar] [CrossRef] [PubMed]
- Kasulo, V.; Mwabumba, L.; Munthali, C. A review of edible orchids in Malawi. J. Hortic. For. 2009, 1, 133–139. [Google Scholar]
- Hamisy, C.W. Development of conservation strategies for the wild edible orchid in Tanzania. Prog. Rep. Rufford Small Grants Found. 2010. [Google Scholar]
- Veldman, S.; Gravendeel, B.; Otieno, J.N.; Lammers, Y.; Duijm, E.; Nieman, A.; Bytebier, B.; Ngugi, G.; Martos, F.; van Andel, T.R.; et al. High-throughput sequencing of African chikanda cake highlights conservation challenges in orchids. Biodivers. Conserv. 2017, 1–18. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.; de Waard, J. Biological identifications through DNA barcodes. Proc. R. Soc. B 2003, 270, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Raclariu, A.C.; Heinrich, M.; Ichim, M.C.; Boer, H. Benefits and limitations of DNA barcoding and metabarcoding in herbal product authentication. Phytochem. Anal. 2017. [Google Scholar] [CrossRef] [PubMed]
- Martos, F.; Le Péchon, T.; Ngugi, G.; Pailler, T.; Bellstedt, D.U.; Linder, H.P.; Bytebier, B. Phylogenetic relationships amongst the African genera of the tribe Orchideae (Orchidaceae). Manuscript in prep. 2018. [Google Scholar]
- Goodman, L.A. Snowball sampling. Ann. Math. Stat. 1961, 148–170. [Google Scholar] [CrossRef]
- Dunning, L.T.; Savolainen, V. Broad-scale amplification of matK for DNA barcoding plants, a technical note: AMPLIFICATION OF matK FOR DNA BARCODING PLANTS. Bot. J. Linn. Soc. 2010, 164, 1–9. [Google Scholar] [CrossRef]
- Shepherd, L.D.; McLay, T.G. Two micro-scale protocols for the isolation of DNA from polysaccharide-rich plant tissue. J. Plant Res. 2011, 124, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Kress, W.J.; Wurdack, K.J.; Zimmer, E.A.; Weigt, L.A.; Janzen, D.H. Use of DNA barcodes to identify flowering plants. Proc. Natl. Acad. Sci. USA 2005, 102, 8369–8374. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Skinner, D.Z.; Liang, G.H.; Hulbert, S.H. Phylogenetic analysis of Sorghum and related taxa using internal transcribed spacers of nuclear ribosomal DNA. Theor. Appl. Genet. 1994, 89. [Google Scholar] [CrossRef] [PubMed]
- Bonfield, J.K.; Smith, K.F.; Staden, R. A new DNA sequence assembly program. Nucleic Acids Res. 1995, 23, 4992–4999. [Google Scholar] [CrossRef] [PubMed]
- Staden, R. The Staden sequence analysis package. Mol. Biotechnol. 1996, 5, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef] [PubMed]
- Davenport, T.R.B.; Bytebier, B. Kitulo Plateau, Tanzania-a first African park for orchids. Orchid Rev. 2004, 112, 161–165. [Google Scholar]
- Boessenkool, S.; Mcglynn, G.; Epp, L.S.; Taylor, D.; Pimentel, M.; Gizaw, A.; Nemomissa, S.; Brochmann, C.; Popp, M. Use of Ancient Sedimentary DNA as a Novel Conservation Tool for High-Altitude Tropical Biodiversity. Conserv. Biol. 2014, 28, 446–455. [Google Scholar] [CrossRef] [PubMed]
- Van Geel, B.; Aptroot, A.; Baittinger, C.; Birks, H.H.; Bull, I.D.; Cross, H.B.; Evershed, R.P.; Gravendeel, B.; Kompanje, E.J.; Kuperus, P. The ecological implications of a Yakutian mammoth’s last meal. Quat. Res. 2008, 69, 361–376. [Google Scholar] [CrossRef]
- Kress, W.J.; Erickson, D.L. A two-locus global DNA barcode for land plants: the coding rbcL gene complements the non-coding trnH-psbA spacer region. PLOS ONE 2007, 2, e508. [Google Scholar] [CrossRef] [PubMed]
- Fazekas, A.J.; Burgess, K.S.; Kesanakurti, P.R.; Graham, S.W.; Newmaster, S.G.; Husband, B.C.; Percy, D.M.; Hajibabaei, M.; Barrett, S.C.H. Multiple multilocus DNA barcodes from the plastid genome discriminate plant species equally well. PLOS ONE 2008, 3, e2802. [Google Scholar] [CrossRef] [PubMed]
- Kool, A.; de Boer, H.J.; Krüger, Å.; Rydberg, A.; Abbad, A.; Björk, L.; Martin, G. Molecular Identification of Commercialized Medicinal Plants in Southern Morocco. PLoS ONE 2012, 7, e39459. [Google Scholar] [CrossRef] [PubMed]
- Elder, J.F.; Turner, B.J. Concerted Evolution of Repetitive DNA Sequences in Eukaryotes. Q. Rev. Biol. 1995, 70, 297–320. [Google Scholar] [CrossRef] [PubMed]
- Ganley, A.R.D.; Kobayashi, T. Highly efficient concerted evolution in the ribosomal DNA repeats: Total rDNA repeat variation revealed by whole-genome shotgun sequence data. Genome Res. 2007, 17, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Harpke, D.; Peterson, A. Non-concerted ITS evolution in Mammillaria (Cactaceae). Mol. Phylogenet. Evol. 2006, 41, 579–593. [Google Scholar] [CrossRef] [PubMed]
- Feliner, G.; Rosselló, J. Better the devil you know? Guidelines for insightful utilization of nrDNA ITS in species-level evolutionary studies in plants. Mol. Phylogenet. Evol. 2007, 44, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Cai, D.; Yao, L.; Teng, Y. Non-concerted ITS evolution, early origin and phylogenetic utility of ITS pseudogenes in Pyrus. Mol. Phylogenet. Evol. 2008, 48, 892–903. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Zeng, X.-M.; Gao, X.-F.; Jin, D.-P.; Zhang, L.-B. ITS non-concerted evolution and rampant hybridization in the legume genus Lespedeza (Fabaceae). Sci. Rep. 2017, 7, 40057. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yao, H.; Han, J.; Liu, C.; Song, J.; Shi, L.; Zhu, Y.; Ma, X.; Gao, T.; Pang, X.; et al. Validation of the ITS2 region as a novel DNA barcode for identifying medicinal plant species. PLoS ONE 2010, 5, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Mezzasalma, V.; Bruni, I.; Fontana, D.; Galimberti, A.; Magoni, C.; Labra, M. A DNA barcoding approach for identifying species in Amazonian traditional medicine: The case of Piri-Piri. Plant Gene 2017, 9, 1–5. [Google Scholar] [CrossRef]
- Bytebier, B.; Dirk, U.; Bellstedt, H. Peter Linder A New Phylogeny-Based Sectional Classification for the Large African Orchid Genus Disa. Taxon 2008, 57, 1233–1251. [Google Scholar]
- Van der Niet, T.; Linder, H.P. Dealing with incongruence in the quest for the species tree: A case study from the orchid genus Satyrium. Mol. Phylogenet. Evol. 2008, 47, 154–174. [Google Scholar] [CrossRef] [PubMed]
- Whitten, W.M.; Blanco, M.A.; Williams, N.H.; Koehler, S.; Carnevali, G.; Singer, R.B.; Endara, L.; Neubig, K.M. Molecular phylogenetics of Maxillaria and related genera (Orchidaceae: Cymbidieae) based on combined molecular data sets. Am. J. Bot. 2007, 94, 1860–1889. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.A.; Soltis, D.E. matK DNA Sequences and Phylogenetic Reconstruction in Saxifragaceae s. str. Syst. Bot. 1994, 19, 143. [Google Scholar] [CrossRef]
- Pridgeon, A.M.; Solano, R.; Chase, M.W. Phylogenetic relationships in Pleurothallidinae (Orchidaceae): Combined evidence from nuclear and plastid DNA sequences. Am. J. Bot. 2001, 88, 2286–2308. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, A.B. Applied Ethnobotany: People, Wild Plant Use and Conservation; Earthscan: London, UK, 2001; ISBN 1853836974. [Google Scholar]
- Martin, G.J. Ethnobotany: A Methods Manual; Earthscan: London, UK, 2004. [Google Scholar]
- Bingham, M.G.; Smith, P.P. Southern African Plant Red Data Lists: Zambia; Golding, J.S., Bandeira, S.O., Eds.; SABONET: Pretoria, South Africa, 2002. [Google Scholar]
- IUCN IUCN Red List of Threatened Species. Version 2017-3. 2018.
- Bone, R.E.; Wightman, N.; Vinya, R.; Veldman, S.; Yokoya, K.; Hargreaves, S.; Kendon, J.; Crous, H. Edible wild orchid trade: sustaining livelihoods and biodiversity in Zambia. Darwin Initiative Main Project Annual Report. 2017. [Google Scholar]
- Filer, D. BRAHMS V5. 6202 Botanical Research and Herbarium Management System (Software). 2007. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vernacular Name | Collections | Reported Origin | Barcoding IDs | # Samples |
|---|---|---|---|---|
| Fungulwe | SJK16 | unknown | Disa robusta | 1 |
| Iringe | SJK17 | Tanzania | Satyrium buchananii | 1 |
| Satyrium carsonii | 1 | |||
| John White | SJK39 | Mporokoso, Zambia | Satyrium buchananii | 1 |
| Kabula seke | SJK46 | Serenje, Zambia | Habenaria sp. (Clade H. schimperiana, H. kyimbilae, H. microsaccos | 1 |
| Platycoryne crocea | 7 | |||
| Kapapa | SJK44 | Mporokoso, Zambia | Platycoryne crocea | 6 |
| Kasebelela, John White and Myala | SJK41 | Chinsali and Mporokoso, Zambia and Tanzania | Habenaria cf sp. DO122 (Clade H. schimperiana, H. kyimbilae, H. microsaccos | 4 |
| Platycoryne crocea | 4 | |||
| Platycoryne sp./Habenaria sp. | 2 | |||
| Kasebulela and Kapapa | SJK11 | Luwingu, Zambia | Satyrium kitimboense | 6 |
| Satyrium carsonii | 5 | |||
| Mbwelenge | SJK5 | Luwingu, Zambia | Satyrium buchananii | 11 |
| Satyrium sp. | 1 | |||
| SJK32 | Serenje, Zambia | Satyrium buchananii | 6 | |
| Mshilamshila | SJK7 | Luwingu, Zambia | Brachycorythis sp. | 1 |
| SJK12 | Kawamba, Zambia | Brachycorythis sp. | 1 | |
| Brachycorythis cf. friesii | 1 | |||
| Myala | SJK4 | Mwinilunga, Zambia; | Disa robusta | 4 |
| Disa welwitschii | 1 | |||
| Satyrium buchananii | 4 | |||
| SJK18 | Sumbawanga, Tanzania | Disa robusta | 4 | |
| Satyrium buchananii | 1 | |||
| Myala | SJK37 | Kawambwa, Zambia | Satyrium buchananii | 1 |
| Myala and nampanda | SJK21 | Luapula, Zambia | Disa welwitschii | 2 |
| Satyrium buchananii | 1 | |||
| Ntonkonshi | SJK25 | Democratic Republic of Congo | Disa robusta | 1 |
| Sumbawanga | SJK20 | Sumbawanga, Tanzania | Disa satyriopsis | 1 |
| Mixed | SJK31 | Serenje, Zambia | Disa caffra | 1 |
| Disa robusta | 1 | |||
| Habenaria cf sp. DO122 (Clade H. schimperiana, H. kyimbilae, H. microsaccos | 1 | |||
| Satyrium buchananii | 6 | |||
| Unknown-mixed | SJK19 | Luwingu, Zambia | Satyrium carsonii | 1 |
| Habenaria aff. helicoplectrum (BB3151) | 1 | |||
| Unknown | SJK8 | Mwinilunga, Zambia | Disa miniata | 1 |
| Disa robusta | 2 | |||
| Disa welwitschii | 1 | |||
| Satyrium buchananii | 2 | |||
| SJK9 | Luwingu, Zambia | Satyrium carsonii | 1 | |
| SJK13 | Kawamba, Zambia | Disa celata | 1 | |
| Disa welwitschii | 1 | |||
| Satyrium buchananii | 3 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veldman, S.; Kim, S.-J.; Van Andel, T.R.; Bello Font, M.; Bone, R.E.; Bytebier, B.; Chuba, D.; Gravendeel, B.; Martos, F.; Mpatwa, G.; et al. Trade in Zambian Edible Orchids—DNA Barcoding Reveals the Use of Unexpected Orchid Taxa for Chikanda. Genes 2018, 9, 595. https://doi.org/10.3390/genes9120595
Veldman S, Kim S-J, Van Andel TR, Bello Font M, Bone RE, Bytebier B, Chuba D, Gravendeel B, Martos F, Mpatwa G, et al. Trade in Zambian Edible Orchids—DNA Barcoding Reveals the Use of Unexpected Orchid Taxa for Chikanda. Genes. 2018; 9(12):595. https://doi.org/10.3390/genes9120595
Chicago/Turabian StyleVeldman, Sarina, Seol-Jong Kim, Tinde R. Van Andel, Maria Bello Font, Ruth E. Bone, Benny Bytebier, David Chuba, Barbara Gravendeel, Florent Martos, Geophat Mpatwa, and et al. 2018. "Trade in Zambian Edible Orchids—DNA Barcoding Reveals the Use of Unexpected Orchid Taxa for Chikanda" Genes 9, no. 12: 595. https://doi.org/10.3390/genes9120595
APA StyleVeldman, S., Kim, S.-J., Van Andel, T. R., Bello Font, M., Bone, R. E., Bytebier, B., Chuba, D., Gravendeel, B., Martos, F., Mpatwa, G., Ngugi, G., Vinya, R., Wightman, N., Yokoya, K., & De Boer, H. J. (2018). Trade in Zambian Edible Orchids—DNA Barcoding Reveals the Use of Unexpected Orchid Taxa for Chikanda. Genes, 9(12), 595. https://doi.org/10.3390/genes9120595