Molecular Evidence for Two Domestication Events in the Pea Crop

 , ,
, , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. DArTseq Analysis
2.3. Bioinformatic Data Processing
2.3.1. Genetic Structure and Diversity Analysis
2.3.2. Seed Phenotyping
3. Results
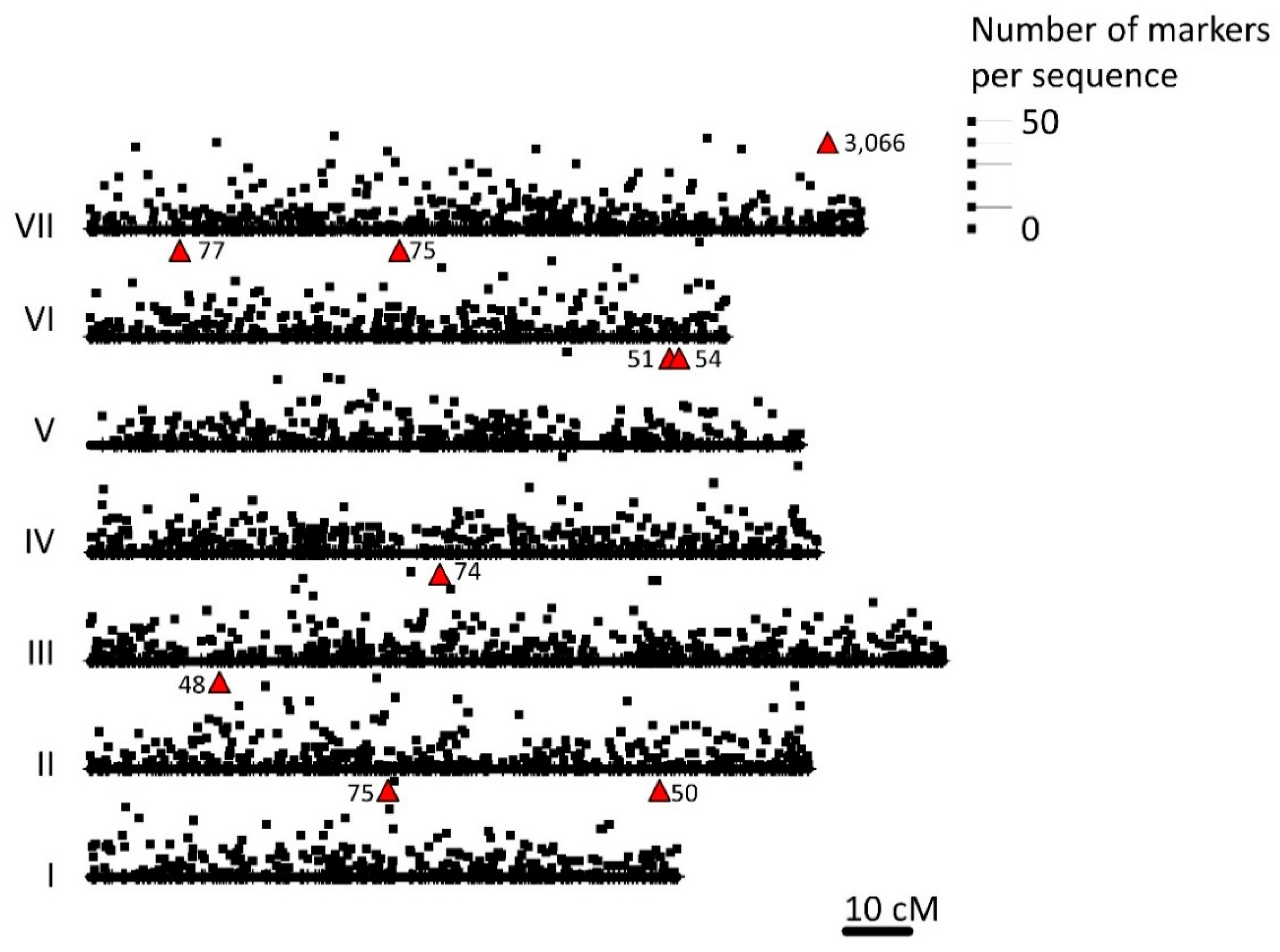
3.1. Diversity Array Technology Analysis and Bioinformatic Processing
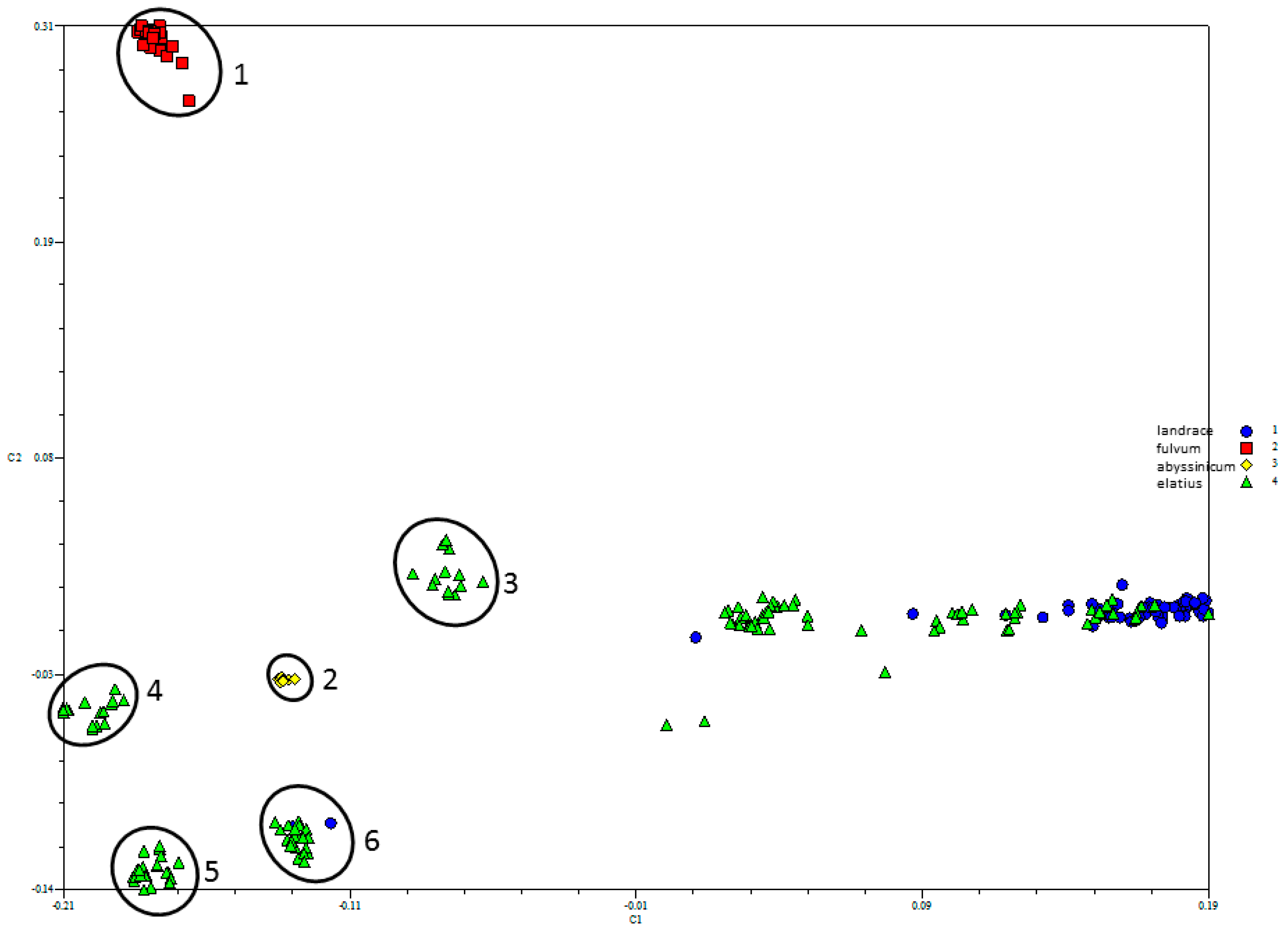
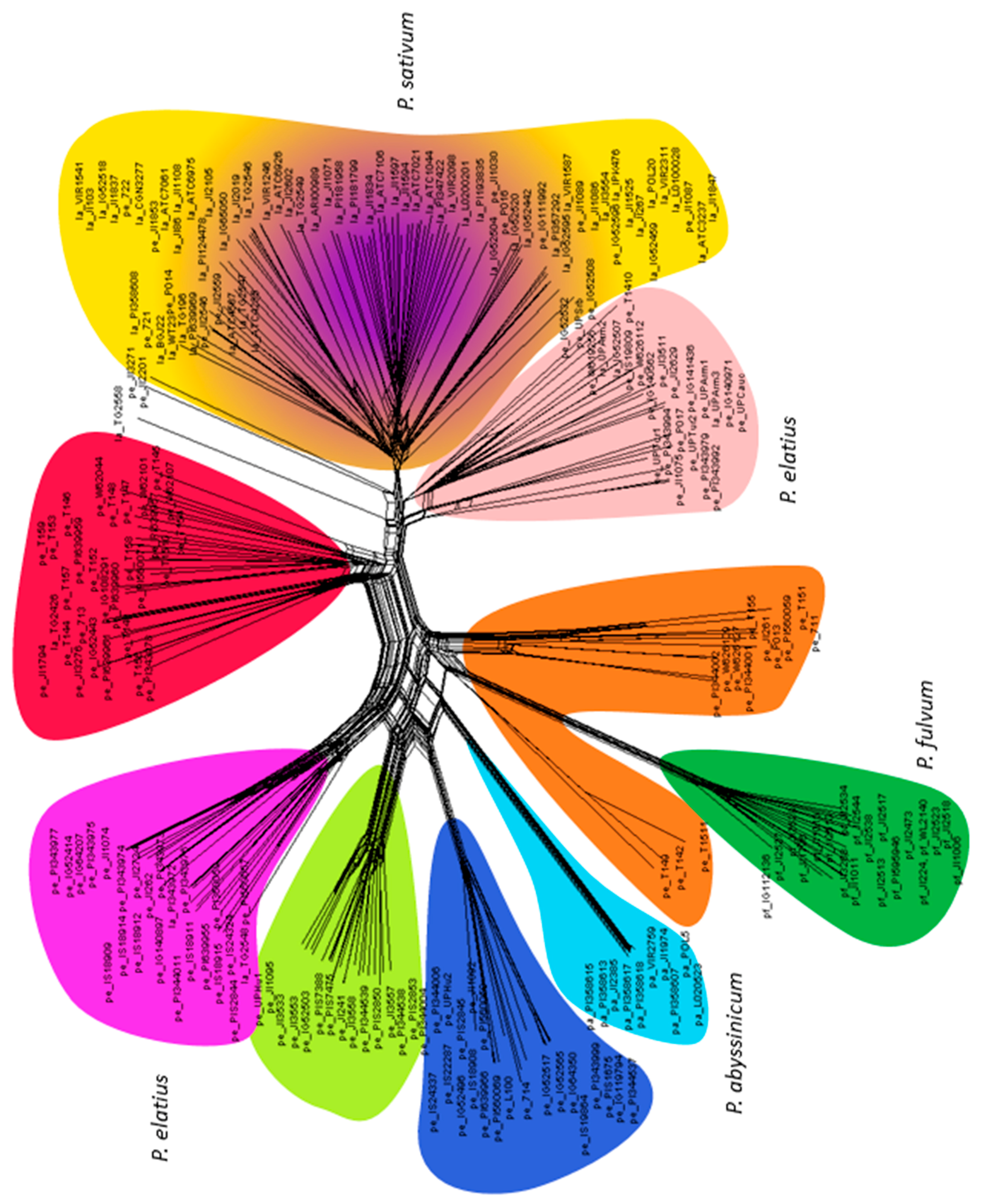
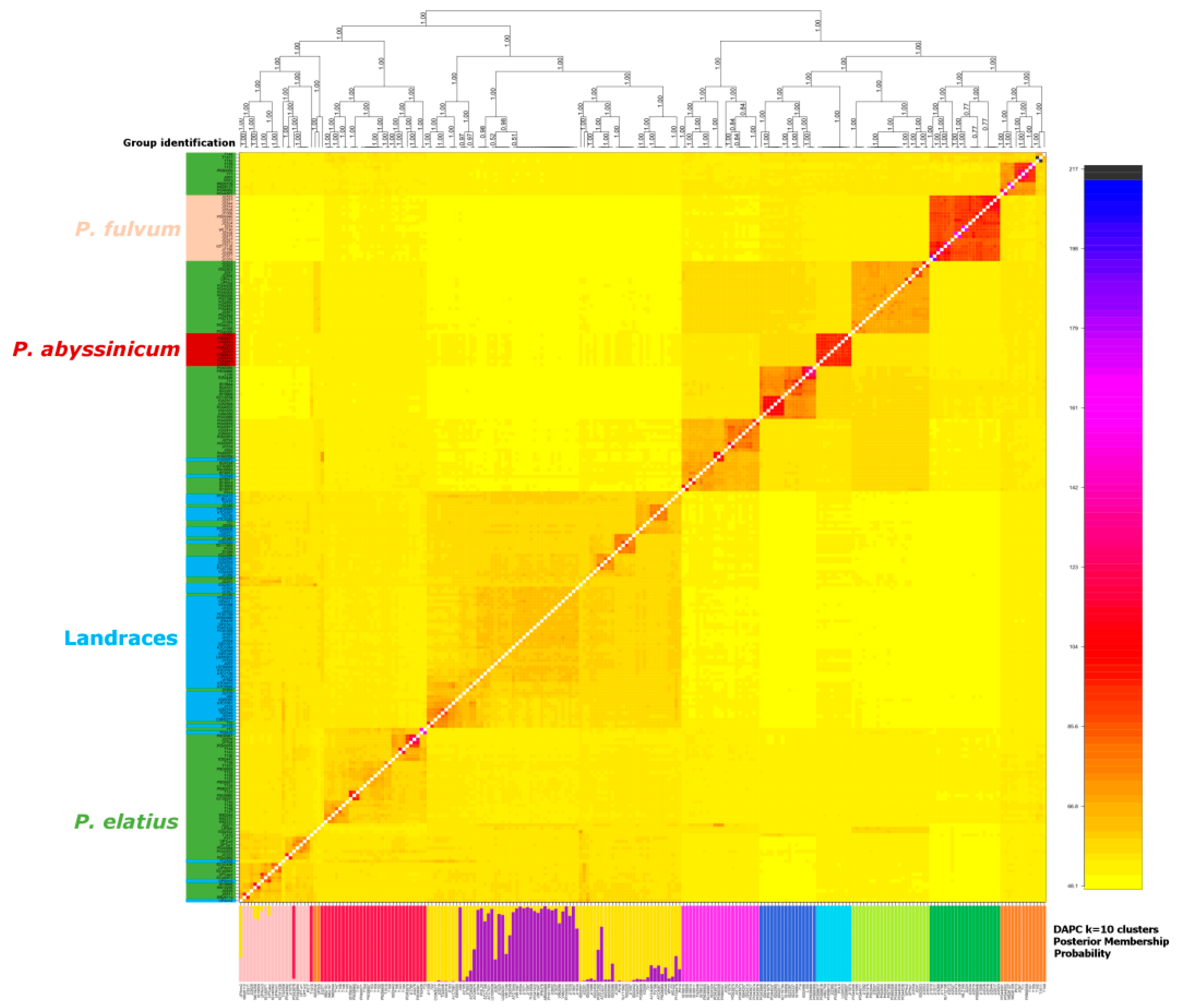
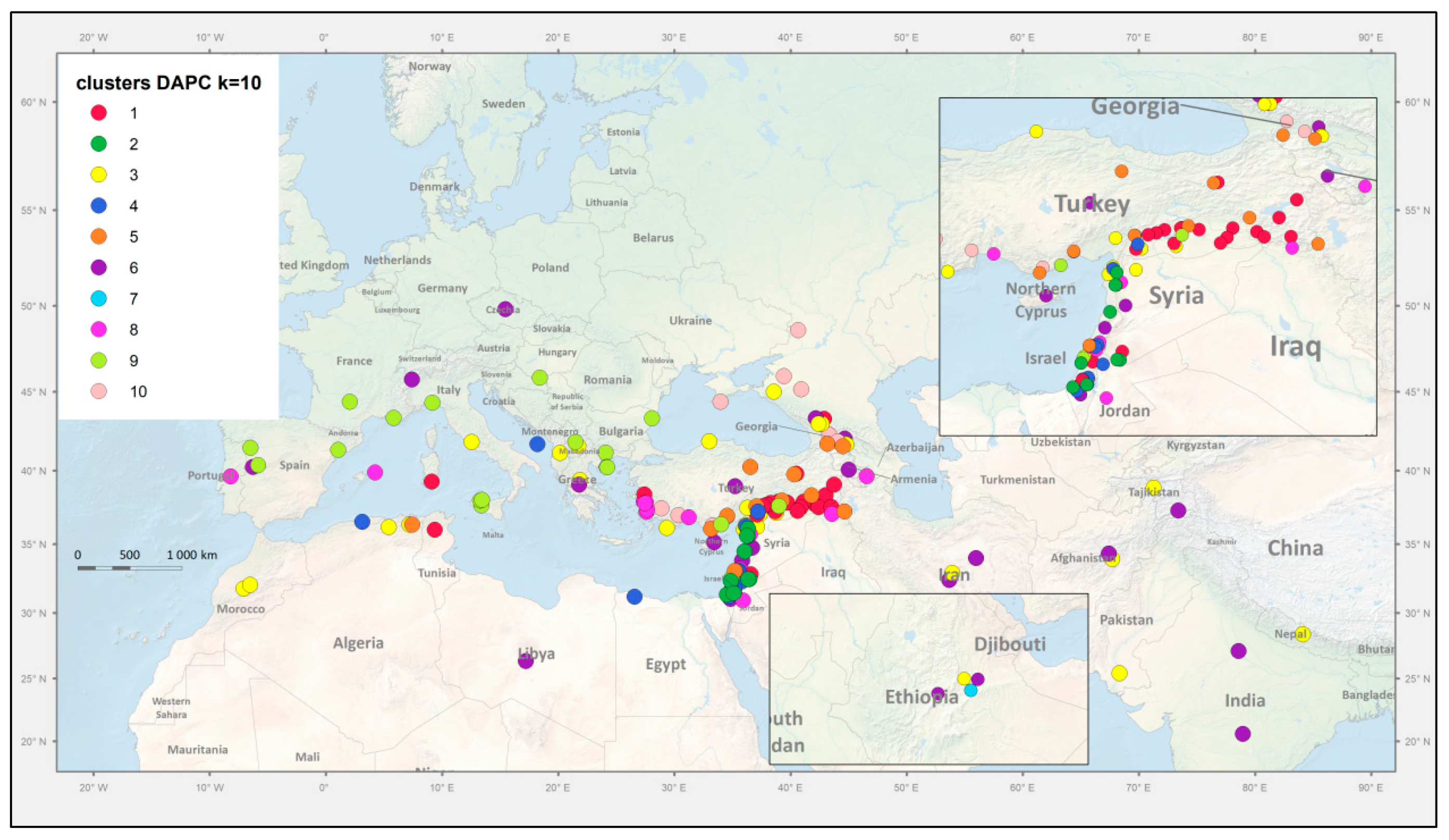
3.2. Genetic Structure and Diversity Analysis
3.3. Seed Phenotyping
4. Discussion
4.1. Pisum Taxonomy
4.2. Pisum Genetic Diversity
4.3. Pea Domestication
4.4. Independent Domestication of the Ethiopian (Pisum abyssinicum) Pea
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lev-Yadun, S.; Gopher, A.; Abbo, S. Archaeology. The cradle of agriculture. Science 2000, 288, 1602–1603. [Google Scholar] [CrossRef] [PubMed]
- Abbo, S.; Pinhasi van-Oss, R.; Gopher, A.; Saranga, Y.; Ofner, I.; Peleg, Z. Plant domestication versus crop evolution: A conceptual framework for cereals and grain legumes. Trends Plant Sci. 2014, 19, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Willcox, G.; Fornite, S.; Herveux, L. Early holocene cultivation before domestication in northern Syria. Veget. Hist. Archaeobot. 2008, 17, 313–325. [Google Scholar] [CrossRef]
- Tanno, K.-I.; Willcox, G. How fast was wild wheat domesticated? Science 2006, 311, 1886. [Google Scholar] [CrossRef] [PubMed]
- Weiss, E.; Kislev, M.E.; Hartmann, A. Anthropology: Autonomous cultivation before domestication. Science 2006, 312, 1608–1610. [Google Scholar] [CrossRef] [PubMed]
- Allaby, R.G.; Fuller, D.Q.; Brown, T.A. The genetic expectations of a protracted model for the origins of domesticated crops. Proc. Natl. Acad. Sci. USA 2008, 105, 13982. [Google Scholar] [CrossRef] [PubMed]
- Fuller, D.Q. Contrasting patterns in crop domestication and domestication rates: Recent archaeobotanical insights from the Old World. Ann. Bot. 2007, 100, 903–924. [Google Scholar] [CrossRef] [PubMed]
- Fuller, D.Q.; Asouti, E.; Purugganan, M.D. Cultivation as slow evolutionary entanglement: Comparative data on rate and sequence of domestication. Veget. Hist. Archaeobot. 2012, 21, 131–147. [Google Scholar] [CrossRef]
- Abbo, S.; Saranga, Y.; Peleg, Z.; Kerem, Z.; Lev-Yadun, S.; Gopher, A. Reconsidering domestication of legumes versus cereals in the ancient near east. Q. Rev. Biol. 2009, 84, 29–50. [Google Scholar] [CrossRef] [PubMed]
- Mithen, S. After the Ice: A Global Human History 20,000-5,000 BC; Weidenfield & Nicholson: London, UK, 2003; ISBN 13 978-0674019997. [Google Scholar]
- Ladizinsky, G. Plant Evolution under Domestication; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1998; ISBN 10 0412822105. [Google Scholar]
- Zohary, D.; Hopf, M.; Weiss, E. Domestication of Plants in the Old World: The Origin and Spread of Domesticated Plants in Southwest Asia, Europe, and the Mediterranean Basin, 4th ed.; Oxford University Press: Oxford, UK, 2012; ISBN 9780199549061. [Google Scholar]
- Zohary, D.; Hopf, M. Domestication of Pulses in the Old World: Legumes were companions of wheat and barley when agriculture began in the Near East. Science 1973, 182, 887–894. [Google Scholar] [CrossRef] [PubMed]
- De Candolle, A. Origin of Cultivated Plants; Kessinger Publishing: Whitefish, UK, 2006; ISBN 9781116495034. [Google Scholar]
- Vavilov, N.I. The Origin, Variation, Immunity and Breeding of Cultivated Plants; Translated from the Russian by K. Starchester; Chronica Botanica: Leyden, The Netherlands, 1951; Volume 13, pp. 1–364. [Google Scholar]
- Smartt, J. Grain Legumes: Evolution and Genetic Resources; Cambridge University Press: Cambridge, UK, 1990. [Google Scholar]
- Ambrose, M.J. From Near East centre of origin the prized pea migrates thorough world. Diversity 1995, 11, 118–119. [Google Scholar]
- Kislev, M.E.; Bar-Yosef, O. The legumes: The earliest domesticated plants in the Near East? Curr. Anthropol. 1988, 29, 175–179. [Google Scholar] [CrossRef]
- Holtgrieve, G.W.; Schindler, D.E.; Hobbs, W.O.; Leavitt, P.R.; Ward, E.J.; Bunting, L.; Chen, G.; Finney, B.P.; Gregory-Eaves, I.; Holmgren, S.; et al. A coherent signature of anthropogenic nitrogen deposition to remote watersheds of the Northern Hemisphere. Science 2011, 334, 1545–1548. [Google Scholar] [CrossRef] [PubMed]
- Erisman, J.W.; Sutton, M.A.; Galloway, J.; Klimont, Z.; Winiwarter, W. How a century of ammonia synthesis changed the world. Nat. Geosci. 2008, 1, 636–639. [Google Scholar] [CrossRef]
- Diamond, J. Evolution, consequences and future of plant and animal domestication. Nature 2002, 418, 700–707. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT 2016. Available online: http://www.fao.org/faostat/en/ (accessed on 26 September 2018).
- Smýkal, P.; Coyne, C.; Ambrose, M.; Maxted, N.; Schaefer, H.; Blair, M.; Berger, J.; Greene, S.L.; Nelson, M.; Besharat, N.; et al. Legume crops phylogeny and genetic diversity for science and breeding. Crit. Rev. Plant Sci. 2014, 33, 43–104. [Google Scholar] [CrossRef]
- Lewis, G.; Schrire, B.; Mackinder, B.; Lock, M. Legumes of the World; Royal Botanic Gardens: London, UK, 2005; ISBN 1900347806. [Google Scholar]
- Schaefer, H.; Hechenleitner, P.; Santos-Guerra, A.; Menezes de Sequeira, M.; Pennington, R.T.; Kenicer, G.; Carine, M.A. Systematics, biogeography, and character evolution of the legume tribe Fabeae with special focus on the middle-Atlantic island lineages. BMC Evol. Biol. 2012, 12, 250. [Google Scholar] [CrossRef] [PubMed]
- Govorov, L. Pisum. In Flora of Cultivated Plants IV, Grain Leguminosae; Vavilovk, N.I., Wulff, E.V., Eds.; State Agricultural Publishing Company: Moscow/Leningrad, Russia, 1937; pp. 231–336. [Google Scholar]
- Makasheva, R.K. Gorokh (pea). In Kulturnaya Flora SSR; Korovina, O.N., Ed.; Kolos Publishers: Leningrad, Russia, 1979; pp. 1–324. [Google Scholar]
- Maxted, N.; Ambrose, M. Peas (Pisum L.). In Plant Genetic Resources of Legumes in the Mediterranean; Maxted, N., Bennett, S.J., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2001; pp. 181–190. ISBN 0792367073. [Google Scholar]
- Smýkal, P.; Kenicer, G.; Flavell, A.J.; Corander, J.; Kosterin, O.; Redden, R.J.; Ford, R.; Coyne, C.J.; Maxted, N.; Ambrose, M.J.; et al. Phylogeny, phylogeography and genetic diversity of the Pisum genus. Plant Genet. Res. 2011, 9, 4–18. [Google Scholar] [CrossRef]
- Smýkal, P.; Hradilová, I.; Trněný, O.; Brus, J.; Rathore, A.; Bariotakis, M.; Das, R.R.; Bhattacharyya, D.; Richards, C.; Coyne, C.J.; et al. Genomic diversity and macroecology of the crop wild relatives of domesticated pea. Sci. Rep. 2017, 7, 17384. [Google Scholar] [CrossRef] [PubMed]
- Jing, R.; Vershinin, A.; Grzebyta, J.; Shaw, P.; Smýkal, P.; Marshall, D.; Ambrose, M.J.; Ellis, T.H.N.; Flavell, A.J. The genetic diversity and evolution of field pea (Pisum) studied by high throughput retrotransposon based insertion polymorphism (RBIP) marker analysis. BMC Evol. Biol. 2010, 10, 44. [Google Scholar] [CrossRef] [PubMed]
- Butler, A. The ethiopian pea: Seeking the evidence for a separate domestication. In Food, Fuel and Fields: Progress in African Archaeobotany; African Archaeobotany: Frankfurt, Germany, 2003; pp. 37–47. ISBN 3927688207. [Google Scholar]
- Ellis, T.H.; Poyser, S.J.; Knox, M.R.; Vershinin, A.V.; Ambrose, M.J. Polymorphism of insertion sites of Ty1-copia class retrotransposons and its use for linkage and diversity analysis in pea. Mol. Gen. Genet. 1998, 260, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Vershinin, A.V.; Allnutt, T.R.; Knox, M.R.; Ambrose, M.J.; Ellis, T.H.N. Transposable elements reveal the impact of introgression, rather than transposition, in Pisum diversity, evolution, and domestication. Mol. Biol. Evol. 2003, 20, 2067–2075. [Google Scholar] [CrossRef] [PubMed]
- Jing, R.; Johnson, R.; Seres, A.; Kiss, G.; Ambrose, M.J.; Knox, M.R.; Ellis, T.H.N.; Flavell, A.J. Gene-based sequence diversity analysis of field pea (Pisum). Genetics 2007, 177, 2263–2275. [Google Scholar] [CrossRef] [PubMed]
- Polans, N.O.; Moreno, R.R. Microsatellite and ITS sequence variation in wild species and cultivars of pea. Pisum Genet. 2009, 41, 3–6. [Google Scholar]
- Ellis, T.H.N. Pisum. In Wild Crop Relatives, Genomic and Breeding Resources; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 237–248. ISBN 978-3-642-14387-8. [Google Scholar]
- Weeden, N.F. Domestication of pea (Pisum sativum L.): The case of the Abyssinian pea. Front. Plant Sci. 2018, 9, 515. [Google Scholar] [CrossRef] [PubMed]
- Smýkal, P.; Vernoud, V.; Blair, M.W.; Soukup, A.; Thompson, R.D. The role of the testa during development and in establishment of dormancy of the legume seed. Front. Plant Sci. 2014, 5, 351. [Google Scholar] [CrossRef] [PubMed]
- Baldev, B. Origin, distribution, taxonomy, and morphology. In Pulse Crops; Baldev, B., Ramanujam, S., Jain, H.K., Eds.; Oxford and IBH Publishing Co.: New Delhi, India, 1988; pp. 3–51. ISBN 9788120402782. [Google Scholar]
- Riehl, S.; Zeidi, M.; Conard, N.J. Emergence of agriculture in the foothills of the Zagros Mountains of Iran. Science 2013, 341, 65–67. [Google Scholar] [CrossRef] [PubMed]
- Mikić, A.; Medović, A.; Jovanović, Ž.; Stanisavljević, N. Integrating archaeobotany, paleogenetics and historical linguistics may cast more light onto crop domestication: The case of pea (Pisum sativum). Genet. Resour. Crop Evol. 2014, 61, 887–892. [Google Scholar] [CrossRef]
- Chimwamurombe, P.M.; Khulbe, R.K. Domestication. In Biology and Breeding of Food Legumes; Pratap, A., Kumar, J., Eds.; CABI: Cambridge, UK, 2011; pp. 19–34. ISBN 978-1-84593-766-9. [Google Scholar]
- Zong, X.; Guan, J.P.; Wang, S.M.; Liu, Q.; Redden, R.; Ford, R. Genetic diversity and core collection of alien Pisum sativum L. germplasm. Acta Agron. Sin. 2008, 34, 1518–1528. [Google Scholar] [CrossRef]
- Zong, X.; Redden, R.J.; Liu, Q.; Wang, S.; Guan, J.; Liu, J.; Xu, Y.; Liu, X.; Gu, J.; Yan, L.; et al. Analysis of a diverse global Pisum sp. collection and comparison to a Chinese local P. sativum collection with microsatellite markers. Theor. Appl. Genet. 2009, 118, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Redden, R.J.; Zong, X.; Berger, J.D.; Bennett, S.J. Ecogeographic analysis of pea collection sites from China to determine potential sites with abiotic stresses. Genet. Resour. Crop Evol. 2013, 60, 1801–1815. [Google Scholar] [CrossRef]
- Ben-Ze’ev, N.; Zohary, D. Species relationships in the genus Pisum L. Isr. J. Bot. 1973, 22, 73–91. [Google Scholar]
- Errico, A.; Conicella, C.; Venora, G. Karyotype studies on Pisum fulvum and Pisum sativum, using a chromosome image analysis system. Genome 1991, 34, 105–108. [Google Scholar] [CrossRef]
- Pourkheirandish, M.; Hensel, G.; Kilian, B.; Senthil, N.; Chen, G.; Sameri, M.; Azhaguvel, P.; Sakuma, S.; Dhanagond, S.; Sharma, R.; et al. Evolution of the grain dispersal system in barley. Cell 2015, 162, 527–539. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, V.; Triska, P.; Pereira, J.B.; Alshamali, F.; Rito, T.; Machado, A.; Fajkošová, Z.; Cavadas, B.; Černý, V.; Soares, P.; et al. Genetic stratigraphy of key demographic events in Arabia. PLoS ONE 2015, 10, e0118625. [Google Scholar] [CrossRef] [PubMed]
- Gallego-Llorente, M.; Jones, E.R.; Eriksson, A.; Siska, V.; Arthur, K.W.; Arthur, J.W.; Curtis, M.C.; Stock, J.T.; Coltorti, M.; Pieruccini, P.; et al. Ancient Ethiopian genome reveals extensive Eurasian admixture throughout the African continent. Science 2015, 350, 820–822. [Google Scholar] [CrossRef] [PubMed]
- Smýkal, P.; Coyne, C.; Redden, R.; Maxted, N. Peas. In Genetic and Genomic Resources of Grain Legume Improvement; Singh, M., Upadhyaya, H.D., Bisht, I.S., Eds.; Elsevier: Amsterdam, The Netherlands, 2013; ISBN 9780123979353. [Google Scholar]
- Warkentin, T.; Smykal, P.; Coyne, C.; Weeden, N.; Domoney, C.; Bing, D.; Leonforte, T.; Xuxiao, Z.; Dixit, G.; Boros, L.; et al. Pea. In Grain Legumes. Handook of Plant Breeding; De Ron, A.M., Ed.; Springer: New York, NY, USA, 2015; Volume 10, pp. 37–85. ISBN 978-1-4939-2797-5. [Google Scholar]
- Tar’an, B.; Zhang, C.; Warkentin, T.; Tullu, A.; Vandenberg, A. Genetic diversity among varieties and wild species accessions of pea (Pisum sativum L.) based on molecular markers, and morphological and physiological characters. Genome 2005, 48, 257–272. [Google Scholar] [CrossRef] [PubMed]
- Siol, M.; Jacquin, F.; Chabert-Martinello, M.; Smýkal, P.; Le Paslier, M.-C.; Aubert, G.; Burstin, J. Patterns of genetic structure and linkage disequilibrium in a large collection of pea germplasm. G3 (Bethesda) 2017, 7, 2461–2471. [Google Scholar] [CrossRef] [PubMed]
- Burstin, J.; Salloignon, P.; Chabert-Martinello, M.; Magnin-Robert, J.-B.; Siol, M.; Jacquin, F.; Chauveau, A.; Pont, C.; Aubert, G.; Delaitre, C.; et al. Genetic diversity and trait genomic prediction in a pea diversity panel. BMC Genom. 2015, 16, 105. [Google Scholar] [CrossRef] [PubMed]
- Tayeh, N.; Aluome, C.; Falque, M.; Jacquin, F.; Klein, A.; Chauveau, A.; Bérard, A.; Houtin, H.; Rond, C.; Kreplak, J.; et al. Development of two major resources for pea genomics: The GenoPea 13.2K SNP Array and a high-density, high-resolution consensus genetic map. Plant J. 2015, 84, 1257–1273. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Li, N.; Hao, J.; Hu, J.; Zhang, X.; Blair, M.W. Genetic diversity of Chinese and global pea (Pisum sativum L.) collections. Crop Sci. 2017, 57, 1–11. [Google Scholar] [CrossRef]
- Baranger, A.; Aubert, G.; Arnau, G.; Lainé, A.L.; Deniot, G.; Potier, J.; Weinachter, C.; Lejeune-Hénaut, I.; Lallemand, J.; Burstin, J. Genetic diversity within Pisum sativum using protein- and PCR-based markers. Theor. Appl. Genet. 2004, 108, 1309–1321. [Google Scholar] [CrossRef] [PubMed]
- Tayeh, N.; Klein, A.; Le Paslier, M.-C.; Jacquin, F.; Houtin, H.; Rond, C.; Chabert-Martinello, M.; Magnin-Robert, J.-B.; Marget, P.; Aubert, G.; et al. Genomic prediction in pea: Effect of marker density and training population size and composition on prediction accuracy. Front. Plant Sci. 2015, 6, 941. [Google Scholar] [CrossRef] [PubMed]
- Holdsworth, W.L.; Gazave, E.; Cheng, P.; Myers, J.R.; Gore, M.A.; Coyne, C.J.; McGee, R.J.; Mazourek, M. A community resource for exploring and utilizing genetic diversity in the USDA pea single plant plus collection. Hort. Res. 2017, 4, 17017. [Google Scholar] [CrossRef] [PubMed]
- Jing, R.; Ambrose, M.A.; Knox, M.R.; Smykal, P.; Hybl, M.; Ramos, Á.; Caminero, C.; Burstin, J.; Duc, G.; van Soest, L.J.M.; et al. Genetic diversity in European Pisum germplasm collections. Theor. Appl. Genet. 2012, 125, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.Y.; Platts, A.E.; Fuller, D.Q.; Hsing, Y.-I.; Wing, R.A.; Purugganan, M.D. The rice paradox: Multiple origins but single domestication in Asian rice. Mol. Biol. Evol. 2017, 34, 969–979. [Google Scholar] [CrossRef] [PubMed]
- Poets, A.M.; Fang, Z.; Clegg, M.T.; Morrell, P.L. Barley landraces are characterized by geographically heterogeneous genomic origins. Genome Biol. 2015, 16, 173. [Google Scholar] [CrossRef] [PubMed]
- Pankin, A.; Altmüller, J.; Becker, C.; von Korff, M. Targeted resequencing reveals genomic signatures of barley domestication. New Phytol. 2018, 218, 1247–1259. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.H.; Sun, D.; Nevo, E. Domestication evolution, genetics and genomics in wheat. Mol. Breed. 2011, 28, 281. [Google Scholar] [CrossRef]
- Civáň, P.; Ivaničová, Z.; Brown, T.A. Reticulated origin of domesticated emmer wheat supports a dynamic model for the emergence of agriculture in the Fertile Crescent. PLoS ONE 2013, 8, e81955. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, Y.; Vigouroux, Y.; Goodman, M.M.; Sanchez, G.J.; Buckler, E.; Doebley, J. A single domestication for maize shown by multilocus microsatellite genotyping. Proc. Natl. Acad. Sci. USA 2002, 99, 6080–6084. [Google Scholar] [CrossRef] [PubMed]
- von Wettberg, E.J.B.; Chang, P.L.; Başdemir, F.; Carrasquila-Garcia, N.; Korbu, L.B.; Moenga, S.M.; Bedada, G.; Greenlon, A.; Moriuchi, K.S.; Singh, V.; et al. Ecology and genomics of an important crop wild relative as a prelude to agricultural innovation. Nat. Commun. 2018, 9, 649. [Google Scholar] [CrossRef] [PubMed]
- Bertioli, D.J.; Cannon, S.B.; Froenicke, L.; Huang, G.; Farmer, A.D.; Cannon, E.K.S.; Liu, X.; Gao, D.; Clevenger, J.; Dash, S.; et al. The genome sequences of Arachis duranensis and Arachis ipaensis, the diploid ancestors of cultivated peanut. Nat. Genet. 2016, 48, 438–446. [Google Scholar] [CrossRef] [PubMed]
- Bitocchi, E.; Rau, D.; Bellucci, E.; Rodriguez, M.; Murgia, M.L.; Gioia, T.; Santo, D.; Nanni, L.; Attene, G.; Papa, R. Beans (Phaseolus ssp.) as a model for understanding crop evolution. Front. Plant Sci. 2017, 8, 722. [Google Scholar] [CrossRef] [PubMed]
- Sedivy, E.J.; Wu, F.; Hanzawa, Y. Soybean domestication: The origin, genetic architecture and molecular bases. New Phytol. 2017, 214, 539–553. [Google Scholar] [CrossRef] [PubMed]
- Linnaeus, C. Species Plantarum; Salvius: Stockholm, Sweden, 1753; Volume 2. [Google Scholar]
- Braun, A. Bemerkungen über die Flora von Abyssinien. Flora Oder Allgemeine Botanische Zeitung 1841, 1, 257–288. [Google Scholar]
- Bieberstein, M. Flora Taurico-Caucasica Exhibens Stirpes Phaenomagas in Chersoneso Taurica et regionibus Caucasicis Sponte Crescents; Typis Academicis: Charkouiae/Kharkov, Ukraine, 1808; Volume 2, p. 447. [Google Scholar]
- Sibthorp, J.M.D.; Smith, J.E. Flora Graeca Sibthorpiana; Richard Taylor: London, UK, 1813; Volume 2. [Google Scholar]
- Smýkal, P.; Trněný, O.; Brus, J.; Hanáček, P.; Rathore, A.; Roma, R.D.; Pechanec, V.; Duchoslav, M.; Bhattacharyya, D.; Bariotakis, M.; et al. Genetic structure of wild pea (Pisum sativum subsp elatius) populations in the northern part of the Fertile Crescent reflects moderate cross-pollination and strong effect of geographic but not environmental distance. PLoS ONE 2018, 13, e0194056. [Google Scholar] [CrossRef]
- Kilian, A.; Wenzl, P.; Huttner, E.; Carling, J.; Xia, L.; Blois, H.; Caig, V.; Heller-Uszynska, K.; Jaccoud, D.; Hopper, C.; et al. Diversity arrays technology: A generic genome profiling technology on open platforms. Methods Mol. Biol. 2012, 888, 67–89. [Google Scholar] [CrossRef] [PubMed]
- Cruz, V.M.V.; Kilian, A.; Dierig, D.A. Development of DArT marker platforms and genetic diversity assessment of the U.S. collection of the new oilseed crop Lesquerella and related species. PLoS ONE 2013, 8, e64062. [Google Scholar] [CrossRef] [PubMed]
- Raman, H.; Raman, R.; Kilian, A.; Detering, F.; Carling, J.; Coombes, N.; Diffey, S.; Kadkol, G.; Edwards, D.; McCully, M.; et al. Genome-wide delineation of natural variation for pod shatter resistance in Brassica napus. PLoS ONE 2014, 9, e101673. [Google Scholar] [CrossRef] [PubMed]
- Reeves, P.A.; Richards, C.M. Accurate inference of subtle population structure (and other genetic discontinuities) using principal coordinates. PLoS ONE 2009, 4, e4269. [Google Scholar] [CrossRef] [PubMed]
- Bryant, D.; Moulton, V. Neighbor-net: An agglomerative method for the construction of phylogenetic networks. Mol. Biol. Evol. 2004, 21, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Jombart, T.; Devillard, S.; Balloux, F. Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. BMC Genet. 2010, 11, 94. [Google Scholar] [CrossRef] [PubMed]
- Malinsky, M.; Trucchi, E.; Lawson, D.J.; Falush, D. RADpainter and fineRADstructure: Population inference from RADseq data. Mol. Biol. Evol. 2018, 35, 1284–1290. [Google Scholar] [CrossRef] [PubMed]
- Ranal, M.A.; Santana, D.G. How and why to measure the germination process? Braz. J. Bot. 2006, 29, 1–11. [Google Scholar] [CrossRef]
- Hradilová, I.; Trněný, O.; Válková, M.; Cechová, M.; Janská, A.; Prokešová, L.; Aamir, K.; Krezdorn, N.; Rotter, B.; Winter, P.; et al. A combined comparative transcriptomic, metabolomic, and anatomical analyses of two key domestication traits: Pod dehiscence and seed dormancy in pea (Pisum sp.). Front. Plant Sci. 2017, 8, 542. [Google Scholar] [CrossRef] [PubMed]
- Steele, K.P.; Wojciechowski, M.F. Phylogenetic analyses of tribes Trifolieae and Vicieae, based on sequences of the plastid gene matK (Papilionoideae: Leguminosae). In Advances in Legume Systematics, Part 10, Higher Level Systematics; Klitgaard, B.B., Bruneau, A., Eds.; Kew Royal Botanical Gardens: London, UK, 2003; pp. 355–370. [Google Scholar]
- de Lamarck, M.M. Flore Françoise, ou Description Succinte de Toutes des Plantes qui Croiffent Naturellement en France, disposées selon une nouvelle méthode d’analyse, et précédées par un exposé des principes élémentaires de la botanique, 3rd ed.; Desray: Paris, France, 1778. [Google Scholar]
- Coulot, P.; Rabaute, P. Monographie de Leguminosae de France. 4. Tribus des Fabeae, des Cicereae et des Genisteae. Bulletin de la Société Botanique du Centre-Ouest 2016, 46, 1–902. [Google Scholar]
- Kosterin, O. Abyssinian pea (Lathyrus schaeferi Kosterin nom. Nov. pro Pisum abyssinicum A. Br.) is a problematic taxon. Vavilov J. Genet. Breed. 2017, 21, 158–169. [Google Scholar] [CrossRef]
- Davis, P.H. Lathyrus. In Flora of Turkey; Davis, P.H., Ed.; Edinburgh University Press: Edinburgh, Scottland, UK, 1970; Volume 3, pp. 328–369. ISBN 10 0852245599. [Google Scholar]
- Kupicha, F.K. Vicieae (Adans.) DC. (1825) nom conserv prop. In Advances in Legume Systematics; Polhill, R.M., Raven, P.H., Eds.; Royal Botanical Gardens: Kew, London, UK, 1981; Volume 1, pp. 377–381. ISBN 9780855212247. [Google Scholar]
- Schmalhausen, I. Flora Srednei y Yuzhnoj Rossii, Kryma i Severnogo Kavkaza; Botanicheskii Institut (Akademiia nauk SSSR): Kiev, Russia, 1895; Volume 1, p. 468. [Google Scholar]
- Ascheron, P.; Graebner, P. Synopsis der Mitteleuropaischen Flora Bd 6, Abt 2, IV; W. Engelmann: Leipzig, Germany, 1910. [Google Scholar]
- Boissier, E. Diagnoses plantarum orientalum novarum. Lipsie 1856, 3, 125. [Google Scholar]
- Miller, P. The Gardener’s Dictionary; Containing the Methods of Cultivating and Improving the Kitchen, Fruit and Flower Garden, 8th ed.; Printed by J. and J. Rivington, 1768; Verlag von J. Cramer: Leutershausen, Germany; London, UK, 1969. [Google Scholar]
- Berger, A. Systematic botany of peas and their allies. Peas of New York. In The Vegetables of New York; Hedrick, J.B., Ed.; Lyon Company: Albany, NY, USA, 1928; pp. 1–132. [Google Scholar]
- Lehmann, C. Das morphologische system der saaterbsen (Pisum sativum L. sensu lato Gov. subsp. sativum). Der Züchter 1954, 24, 316–337. [Google Scholar]
- Ladizinsky, G.; Abbo, S. The Search for Wild Relatives of Cool Season Legumes; Springer: Heidelberg, Germany, 2015. [Google Scholar]
- Bogdanova, V.S.; Mglinets, A.V.; Shatskaya, N.V.; Kosterin, O.E.; Solovyev, V.I.; Vasiliev, G.V. Cryptic divergences in the genus Pisum L. (peas), as revealed by phylogenetic analysis of plastid genomes. Mol. Phyl. Evol. 2018, 129, 280–290. [Google Scholar] [CrossRef] [PubMed]
- van Oss, R.; Abbo, S.; Eshed, R.; Sherman, A.; Coyne, C.J.; Vandemark, G.J.; Zhang, H.B.; Peleg, Z. Genetic relationship in Cicer sp. expose evidence for geneflow between the cultigen and its wild progenitor. PLoS ONE 2015, 10, e0139789. [Google Scholar] [CrossRef] [PubMed]
- Meyer, R.S.; DuVal, A.E.; Jensen, H.R. Patterns and processes in crop domestication: An historical review and quantitative analysis of 203 global food crops. New Phytol. 2012, 196, 29–48. [Google Scholar] [CrossRef] [PubMed]
- Varshney, R.K.; Song, C.; Saxena, R.K.; Azam, S.; Yu, S.; Sharpe, A.G.; Cannon, S.; Baek, J.; Rosen, B.D.; Tar’an, B.; et al. Draft genome sequence of chickpea (Cicer arietinum) provides a resource for trait improvement. Nat. Biotechnol. 2013, 31, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Parween, S.; Nawaz, K.; Roy, R.; Pole, A.K.; Venkata Suresh, B.; Misra, G.; Jain, M.; Yadav, G.; Parida, S.K.; Tyagi, A.K.; et al. An advanced draft genome assembly of a desi type chickpea (Cicer arietinum L.). Sci. Rep. 2015, 5, 12806. [Google Scholar] [CrossRef] [PubMed]
- Caracuta, V.; Vardi, J.; Paz, Y.; Boaretto, E. Farming legumes in the pre-pottery Neolithic: New discoveries from the site of Ahihud (Israel). PLoS ONE 2017, 12, e0177859. [Google Scholar] [CrossRef] [PubMed]
- Abbo, S.; Rachamim, E.; Zehavi, Y.; Zezak, I.; Lev-Yadun, S.; Gopher, A. Experimental growing of wild pea in Israel and its bearing on Near Eastern plant domestication. Ann. Bot. 2011, 107, 1399–1404. [Google Scholar] [CrossRef] [PubMed]
- Smýkal, P.; Nelson, M.; Berger, J.; Smýkal, P.; Nelson, M.N.; Berger, J.D.; von Wettberg, E.J.B. The impact of genetic changes during crop domestication. Agronomy 2018, 8, 119. [Google Scholar] [CrossRef]
- Weeden, N.F. Genetic changes accompanying the domestication of Pisum sativum: Is there a common genetic basis to the “domestication syndrome” for legumes? Ann. Bot. 2007, 100, 1017–1025. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Vieira, F.G.; Crawford, J.E.; Chu, C.; Nielsen, R. Asian wild rice is a hybrid swarm with extensive gene flow and feralization from domesticated rice. Genome Res. 2017, 27, 1029–1038. [Google Scholar] [CrossRef] [PubMed]
- Li, L.F.; Li, Y.L.; Jia, Y.; Caicedo, A.L.; Olsen, K.M. Signatures of adaptation in the weedy rice genome. Nat. Genet. 2017, 49, 811–814. [Google Scholar] [CrossRef] [PubMed]
- Abbo, S.; Lev-Yadun, S.; Heun, M.; Gopher, A. On the “lost” crops of the neolithic Near East. J. Exp. Bot. 2013, 64, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Young, J.P.W.; Matthews, P. A distinct class of peas (Pisum sativum L.) from Afghanistan that show strain specificity for symbiotic Rhizobium. Heredity 1982, 48, 203. [Google Scholar] [CrossRef]
- Lamprecht, H. Die Enstehung der Arten und hohhren Kaategorien; Springer: Wien, Germany, 1966. [Google Scholar]
- Gebreslassie, B.; Abraha, B. Review: Distribution and productivity of dekoko (Pisum sativum var. abyssinicum A. Braun) in Ethiopia. Glob. J. Sci. Front. Res. C Biol. Sci. 2016, 16, 1–14. [Google Scholar]
- Baranyi, M.; Greilhuber, J. Flow cytometric analysis of genome size variation in cultivated and wild Pisum sativum (Fabaceae). Plant Syst. Evol. 1995, 194, 231–239. [Google Scholar] [CrossRef]
- Kloz, J. Serology of the Leguminosae. In Chemotaxonomy of the Leguminosae; Harborne, J.B., Boulter, D., Turner, B.L., Eds.; Academic Press: London, UK, 1971; pp. 309–365. ISBN 10-0123246520. [Google Scholar]
- Holden, D. Genetic studies of wide crosses between cultivated pea species, and the domestication of Pisum abyssinicum. Ph.D. Thesis, School of Biological Sciences, University of East Anglia, John Innes Centre & Unilever Research, Colworth, UK, 2009. [Google Scholar]
- Claussen, M.; Kubatzki, C.; Brovkin, V.; Ganopolski, A.; Hoelzmann, P.; Pachur, H.J. Simulation of an abrupt change in Saharan vegetation at the end of the Mid-Holocene. Geophys. Res. Lett. 1999, 24, 2037–2040. [Google Scholar] [CrossRef]
- Weeden, N.; Wolko, B. Allozyme analysis of Pisum sativum ssp. abyssinicum and the development of a genotype definition for this subspecies. Pisum Genet. 2001, 33, 21–25. [Google Scholar]
- Conicella, C.; Errico, A. Karyotype Variations in Pisum Sativum Ect. Abyssinicum. Caryologia 1990, 43, 87–97. [Google Scholar] [CrossRef]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trněný, O.; Brus, J.; Hradilová, I.; Rathore, A.; Das, R.R.; Kopecký, P.; Coyne, C.J.; Reeves, P.; Richards, C.; Smýkal, P. Molecular Evidence for Two Domestication Events in the Pea Crop. Genes 2018, 9, 535. https://doi.org/10.3390/genes9110535
Trněný O, Brus J, Hradilová I, Rathore A, Das RR, Kopecký P, Coyne CJ, Reeves P, Richards C, Smýkal P. Molecular Evidence for Two Domestication Events in the Pea Crop. Genes. 2018; 9(11):535. https://doi.org/10.3390/genes9110535
Chicago/Turabian StyleTrněný, Oldřich, Jan Brus, Iveta Hradilová, Abhishek Rathore, Roma R. Das, Pavel Kopecký, Clarice J. Coyne, Patrick Reeves, Christopher Richards, and Petr Smýkal. 2018. "Molecular Evidence for Two Domestication Events in the Pea Crop" Genes 9, no. 11: 535. https://doi.org/10.3390/genes9110535
APA StyleTrněný, O., Brus, J., Hradilová, I., Rathore, A., Das, R. R., Kopecký, P., Coyne, C. J., Reeves, P., Richards, C., & Smýkal, P. (2018). Molecular Evidence for Two Domestication Events in the Pea Crop. Genes, 9(11), 535. https://doi.org/10.3390/genes9110535