Exploring Interactions between the Gut Microbiota and Social Behavior through Nutrition



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
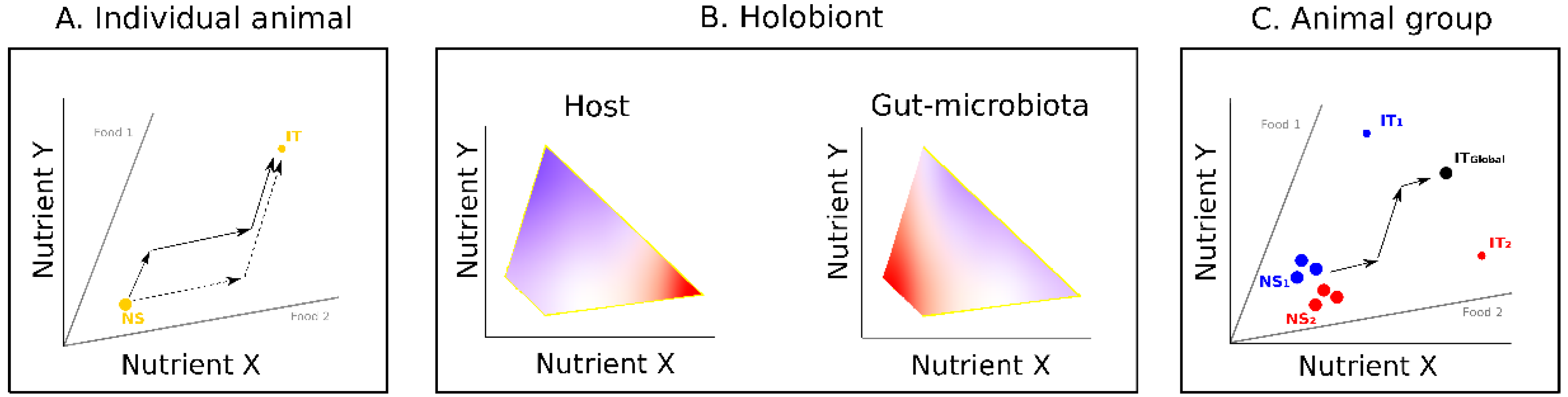
2. Influence of Gut Microbiota on Host Nutrition
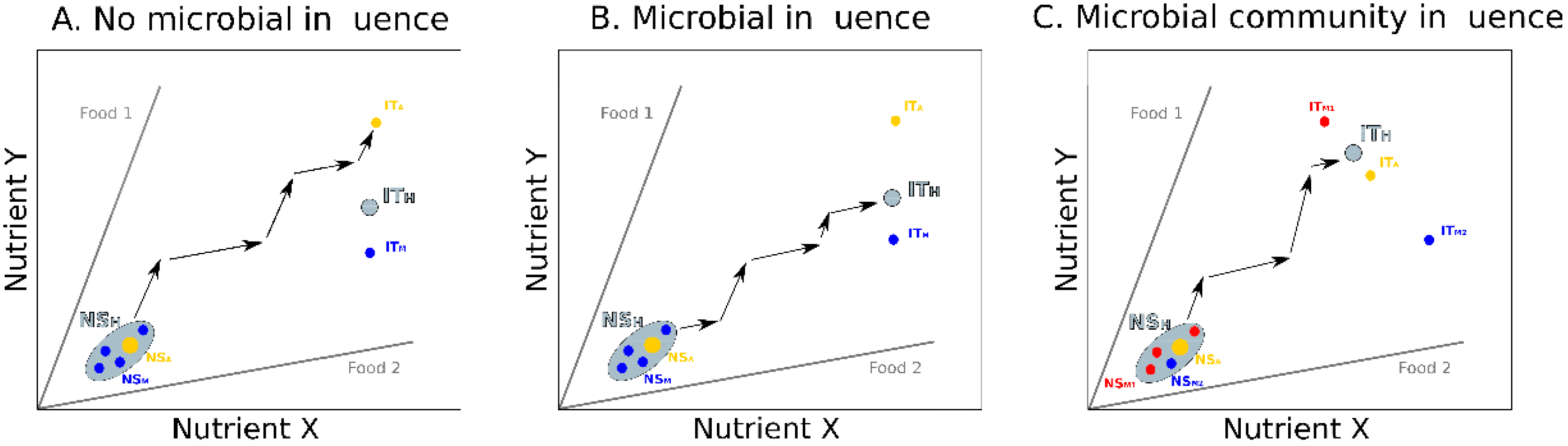
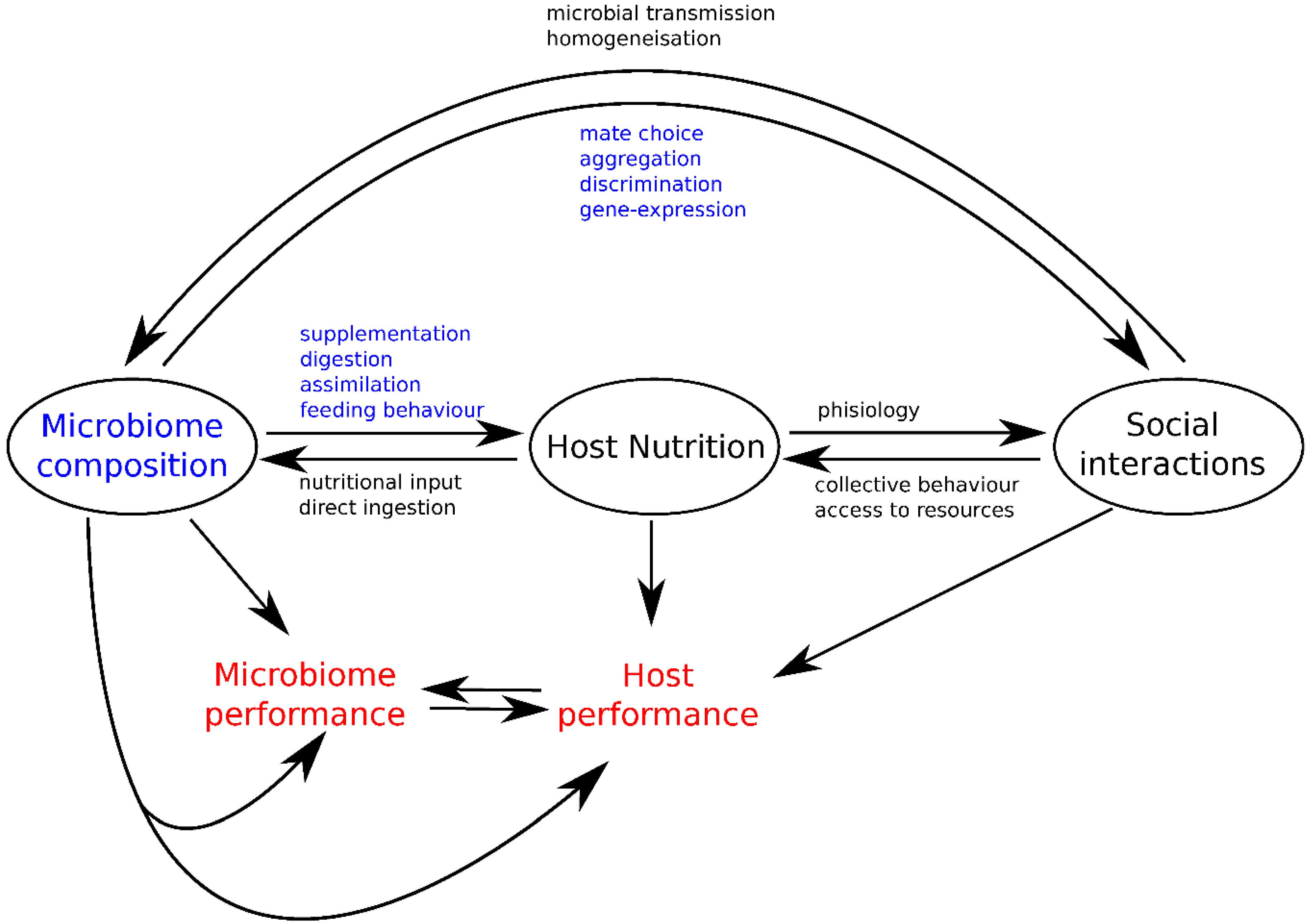
3. Integrating Gut Microbes, Host Nutrition and Social Behavior
4. Conclusions
Funding
Conflicts of Interest
References
- Margulis, L.; Fester, R. Symbiosis as a Source of Evolutionary Innovation: Speciation and Morphogenesis; MIT Press: Cambridge, MA, USA, 1991; ISBN 0-262-13269-9. [Google Scholar]
- Rohwer, F.; Seguritan, V.; Azam, F.; Knowlton, N. Diversity and distribution of coral-associated bacteria. Mar. Ecol. Prog. Ser. 2002, 243, 1–10. [Google Scholar] [CrossRef]
- Theis, K.R.; Dheilly, N.M.; Klassen, J.L.; Brucker, R.M.; Baines, J.F.; Bosch, T.C.; Cryan, J.F.; Gilbert, S.F.; Goodnight, C.J.; Lloyd, E.A. Getting the hologenome concept right: An eco-evolutionary framework for hosts and their microbiomes. Msystems 2016, 1. [Google Scholar] [CrossRef] [PubMed]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Ezenwa, V.O.; Gerardo, N.M.; Inouye, D.W.; Medina, M.; Xavier, J.B. Animal behavior and the microbiome. Science 2012, 338, 198–199. [Google Scholar] [CrossRef] [PubMed]
- Venu, I.; Durisko, Z.; Xu, J.; Dukas, R. Social attraction mediated by fruit flies’ microbiome. J. Exp. Biol. 2014, 217, 1346–1352. [Google Scholar] [CrossRef] [PubMed]
- Wada-Katsumata, A.; Zurek, L.; Nalyanya, G.; Roelofs, W.L.; Zhang, A.; Schal, C. Gut bacteria mediate aggregation in the German cockroach. Proc. Natl. Acad. Sci. USA 2015, 112, 15678–15683. [Google Scholar] [CrossRef] [PubMed]
- Sharon, G.; Segal, D.; Ringo, J.M.; Hefetz, A.; Zilber-Rosenberg, I.; Rosenberg, E. Commensal bacteria play a role in mating preference of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2010, 107, 20051–20056. [Google Scholar] [CrossRef] [PubMed]
- Theis, K.R.; Venkataraman, A.; Dycus, J.A.; Koonter, K.D.; Schmitt-Matzen, E.N.; Wagner, A.P.; Holekamp, K.E.; Schmidt, T.M. Symbiotic bacteria appear to mediate hyena social odors. Proc. Natl. Acad. Sci. USA 2013, 110, 19832–19837. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, K. Nestmate recognition mediated by intestinal bacteria in a termite, Reticulitermes speratus. Oikos 2001, 92, 20–26. [Google Scholar] [CrossRef]
- Buffington, S.A.; Di Prisco, G.V.; Auchtung, T.A.; Ajami, N.J.; Petrosino, J.F.; Costa-Mattioli, M. Microbial Reconstitution Reverses Maternal Diet-Induced Social and Synaptic Deficits in Offspring. Cell 2016, 165, 1762–1775. [Google Scholar] [CrossRef] [PubMed]
- Münger, E.; Montiel-Castro, A.J.; Langhans, W.; Pacheco-López, G. Reciprocal Interactions Between Gut Microbiota and Host Social Behavior. Front. Integr. Neurosci. 2018, 12, 21. [Google Scholar] [CrossRef] [PubMed]
- Lax, S.; Smith, D.P.; Hampton-Marcell, J.; Owens, S.M.; Handley, K.M.; Scott, N.M.; Gibbons, S.M.; Larsen, P.; Shogan, B.D.; Weiss, S. Longitudinal analysis of microbial interaction between humans and the indoor environment. Science 2014, 345, 1048–1052. [Google Scholar] [CrossRef] [PubMed]
- Ponton, F.; Wilson, K.; Cotter, S.C.; Raubenheimer, D.; Simpson, S.J. Nutritional immunology: A multi-dimensional approach. PLoS Pathog. 2011, 7, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Feldhaar, H.; Straka, J.; Krischke, M.; Berthold, K.; Stoll, S.; Mueller, M.J.; Gross, R. Nutritional upgrading for omnivorous carpenter ants by the endosymbiont Blochmannia. BMC Biol. 2007, 5, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kohl, K.D.; Stengel, A.; Dearing, M.D. Inoculation of tannin-degrading bacteria into novel hosts increases performance on tannin-rich diets. Environ. Microbiol. 2016, 18, 1720–1729. [Google Scholar] [CrossRef] [PubMed]
- Douglas, A.E. Aphids and Their Symbiotic Bacteria Buchnera. Annu. Rev. Entomol. 1998, 17–37. [Google Scholar] [CrossRef] [PubMed]
- Nalepa, C.A.; Bignell, D.E.; Bandi, C. Detritivory, coprophagy, and the evolution of digestive mutualisms in Dictyoptera. Insectes Sociaux 2001, 48, 194–201. [Google Scholar] [CrossRef]
- Wong, A.C.N.; Wang, Q.P.; Morimoto, J.; Senior, A.M.; Lihoreau, M.; Neely, G.G.; Simpson, S.J.; Ponton, F. Gut Microbiota Modifies Olfactory-Guided Microbial Preferences and Foraging Decisions in Drosophila. Curr. Biol. 2017, 27, 2397–2404. [Google Scholar] [CrossRef] [PubMed]
- Chandler, J.A.; Lang, J.M.; Bhatnagar, S.; Eisen, J.A.; Kopp, A. Bacterial communities of diverse Drosophila species: Ecological context of a host-microbe model system. PLoS Genet. 2011, 7, e1002272. [Google Scholar] [CrossRef] [PubMed]
- Ravussin, Y.; Koren, O.; Spor, A.; LeDuc, C.; Gutman, R.; Stombaugh, J.; Knight, R.; Ley, R.E.; Leibel, R.L. Responses of gut microbiota to diet composition and weight loss in lean and obese mice. Obesity 2012, 20, 738–747. [Google Scholar] [CrossRef] [PubMed]
- Holmes, A.J.; Chew, Y.V.; Colakoglu, F.; Cliff, J.B.; Klaassens, E.; Read, M.N.; Solon-Biet, S.M.; McMahon, A.C.; Cogger, V.C.; Ruohonen, K.; et al. Diet-Microbiome Interactions in Health Are Controlled by Intestinal Nitrogen Source Constraints. Cell Metab. 2017, 25, 140–151. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.; Ridaura, V.; Faith, J. The effect of diet on the human gut microbiome: A metagenomic analysis in humanized gnotobiotic mice. Sci. Transl. Med. 2009, 1. [Google Scholar] [CrossRef] [PubMed]
- Simpson, S.J.; Raubenheimer, D. A multi-level analysis of feeding behaviour: The geometry of nutritional decisions. Phil. Trans. R. Soc. Lond. B 1993, 342, 381–402. [Google Scholar] [CrossRef]
- Simpson, S.J.; Raubenheimer, D. The Nature of Nutrition: A Unifying Framework from Animal Adaptation to Human Obesity; Princeton University Press: Princeton, NJ, USA, 2012; ISBN 1-4008-4280-8. [Google Scholar]
- Wong, A.C.N.; Holmes, A.; Ponton, F.; Lihoreau, M.; Wilson, K.; Raubenheimer, D.; Simpson, S.J. Behavioral microbiomics: A multi-dimensional approach to microbial influence on behavior. Front. Microbiol. 2015, 6, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Steffan, S.A.; Chikaraishi, Y.; Currie, C.R.; Horn, H.; Gaines-Day, H.R.; Pauli, J.N.; Zalapa, J.E.; Ohkouchi, N. Microbes are trophic analogs of animals. Proc. Natl. Acad. Sci. USA 2015, 112, 15119–15124. [Google Scholar] [CrossRef] [PubMed]
- Lihoreau, M.; Buhl, J.; Charleston, M.A.; Sword, G.A.; Raubenheimer, D.; Simpson, S.J. Nutritional ecology beyond the individual: A conceptual framework for integrating nutrition and social interactions. Ecol. Lett. 2015, 18, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Lihoreau, M.; Buhl, J.; Charleston, M.A.; Sword, G.A.; Raubenheimer, D.; Simpson, S.J. Modelling nutrition across organizational levels: From individuals to superorganisms. J. Insect Physiol. 2014, 69, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Lihoreau, M.; Gómez-Moracho, T.; Pasquaretta, C.; Costa, J.T.; Buhl, J. Social nutrition: An emerging field in insect science. Curr. Opin. Insect Sci. 2018, 28, 73–80. [Google Scholar] [CrossRef]
- Buhl, J.; Sumpter, D.J.T.; Couzin, I.D.; Hale, J.J.; Despland, E.; Miller, E.R.; Simpson, S.J. From disorder to order in marching locusts. Science 2006, 312, 1402–1406. [Google Scholar] [CrossRef] [PubMed]
- Dussutour, A.; Simpson, S.J. Communal nutrition in ants. Curr. Biol. 2009, 19, 740–744. [Google Scholar] [CrossRef] [PubMed]
- Salomon, M.; Mayntz, D.; Lubin, Y. Colony nutrition skews reproduction in a social spider. Behav. Ecol. 2008, 19, 605–611. [Google Scholar] [CrossRef]
- Senior, A.M.; Nakagawa, S.; Lihoreau, M.; Simpson, S.J.; Raubenheimer, D. An overlooked consequence of dietary mixing: A varied diet reduces interindividual variance in fitness. Am. Nat. 2015, 186, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Senior, A.M.; Charleston, M.A.; Lihoreau, M.; Buhl, J.; Raubenheimer, D.; Simpson, S.J. Evolving nutritional strategies in the presence of competition: A geometric agent-based model. PLoS Comput. Biol. 2015, 11, e1004111. [Google Scholar] [CrossRef] [PubMed]
- Senior, A.M.; Lihoreau, M.; Charleston, M.A.; Buhl, J.; Raubenheimer, D.; Simpson, S.J. Adaptive collective foraging in groups with conflicting nutritional needs. R. Soc. Open Sci. 2016, 3, 150638. [Google Scholar] [CrossRef] [PubMed]
- Poissonnier, L.-A.; Lihoreau, M.; Gomez-Moracho, T.; Dussutour, A.; Buhl, J. A theoretical exploration of dietary collective medication in social insects. J. Insect Physiol. 2018, 106, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Behmer, S.T. Animal behaviour: Feeding the superorganism. Curr. Biol. 2009, 19, R366–R368. [Google Scholar] [CrossRef] [PubMed]
- Dosmann, A.; Bahet, N.; Gordon, D.M. Experimental modulation of external microbiome affects nestmate recognition in harvester ants (Pogonomyrmex barbatus). PeerJ 2016, 4, e1566. [Google Scholar] [CrossRef] [PubMed]
- Wagner, D.; Tissot, M.; Cuevas, W.; Gordon, D.M. Harvester ants utilize cuticular hydrocarbons in nestmate recognition. J. Chem. Ecol. 2000, 26, 2245–2257. [Google Scholar] [CrossRef]
- Wagner, D.; Tissot, M.; Gordon, D. Task-related environment alters the cuticular hydrocarbon composition of harvester ants. J. Chem. Ecol. 2001, 27, 1805–1819. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.W.; Blomquist, G.J. Ecological, Behavioral, and Biochemical Aspects of Insect Hydrocarbons. Annu. Rev. Entomol. 2005, 50, 371–393. [Google Scholar] [CrossRef] [PubMed]
- Esponda, F.; Gordon, D.M. Distributed nestmate recognition in ants. Proc. R. Soc. B 2015, 282. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, S.; Heeb, P. Social and sexual behaviours aid transmission of bacteria in birds. Behav. Process. 2007, 74, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.H.; Pothoulakis, C.; Mayer, E.A. Principles and clinical implications of the brain-gut-enteric microbiota axis. Nat. Rev. Gastroenterol. Hepatol. 2009, 6, 306. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.A. Gut feelings: The emerging biology of gut-brain communication. Nat. Rev. Neurosci. 2011, 12, 453. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; Dinan, T.G. Mind-altering microorganisms: The impact of the gut microbiota on brain and behaviour. Nat. Rev. Neurosci. 2012, 13, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Kaelberer, M.M.; Buchanan, K.L.; Klein, M.E.; Barth, B.B.; Montoya, M.M.; Shen, X.; Bohórquez, D.V. A gut-brain neural circuit for nutrient sensory transduction. Science 2018, 361, eaat5236. [Google Scholar] [CrossRef] [PubMed]
- Montiel-Castro, A.J.; González-Cervantes, R.M.; Bravo-Ruiseco, G.; Pacheco-López, G. The microbiota-gut-brain axis: Neurobehavioral correlates, health and sociality. Front. Integr. Neurosci. 2013, 7, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Konturek, S.; Konturek, P.; Pawlik, T.; Brzozowski, T. Brain-gut axis and its role in the control of food intake. J. Physiol. Pharmacol. 2004, 55, 137–154. [Google Scholar] [PubMed]
- Alcock, J.; Maley, C.C.; Aktipis, C.A. Is eating behavior manipulated by the gastrointestinal microbiota? Evolutionary pressures and potential mechanisms. BioEssays 2014, 36, 940–949. [Google Scholar] [CrossRef] [PubMed]
- Kleiman, S.C.; Watson, H.J.; Bulik-Sullivan, E.C.; Huh, E.Y.; Tarantino, L.M.; Bulik, C.M.; Carroll, I.M. The intestinal microbiota in acute anorexia nervosa and during renourishment: Relationship to depression, anxiety, and eating disorder psychopathology. Psychosom. Med. 2015, 77, 969. [Google Scholar] [CrossRef] [PubMed]
- Duca, F.A.; Swartz, T.D.; Sakar, Y.; Covasa, M. Increased oral detection, but decreased intestinal signaling for fats in mice lacking gut microbiota. PLoS ONE 2012, 7, e39748. [Google Scholar] [CrossRef] [PubMed]
- Swartz, T.D.; Duca, F.; De Wouters, T.; Sakar, Y.; Covasa, M. Up-regulation of intestinal type 1 taste receptor 3 and sodium glucose luminal transporter-1 expression and increased sucrose intake in mice lacking gut microbiota. Br. J. Nutr. 2012, 107, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Foster, K.R.; Schluter, J.; Coyte, K.Z.; Rakoff-Nahoum, S. The evolution of the host microbiome as an ecosystem on a leash. Nature 2017, 548, 43. [Google Scholar] [CrossRef] [PubMed]
- Moeller, A.H.; Foerster, S.; Wilson, M.L.; Pusey, A.E.; Hahn, B.H.; Ochman, H. Social behavior shapes the chimpanzee pan-microbiome. Sci. Adv. 2016, 2. [Google Scholar] [CrossRef] [PubMed]
- Tung, J.; Barreiro, L.B.; Burns, M.B.; Grenier, J.C.; Lynch, J.; Grieneisen, L.E.; Altmann, J.; Alberts, S.C.; Blekhman, R.; Archie, E.A. Social networks predict gut microbiome composition in wild baboons. eLife 2015, 2015, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Leclaire, S.; Nielsen, J.F.; Drea, C.M. Bacterial communities in meerkat anal scent secretions vary with host sex, age, and group membership. Behav. Ecol. 2014, 25, 996–1004. [Google Scholar] [CrossRef]
- Lucas, F.S.; Heeb, P. Environmental factors shape cloacal bacterial assemblages in great tit Parus major and blue tit P. caeruleus nestlings. J. Avian Biol. 2005, 36, 510–516. [Google Scholar] [CrossRef]
- White, J.; Mirleau, P.; Danchin, E.; Mulard, H.; Hatch, S.A.; Heeb, P.; Wagner, R.H. Sexually transmitted bacteria affect female cloacal assemblages in a wild bird. Ecol. Lett. 2010, 13, 1515–1524. [Google Scholar] [CrossRef] [PubMed]
- Koch, H.; Schmid-Hempel, P. Socially transmitted gut microbiota protect bumble bees against an intestinal parasite. Proc. Natl. Acad. Sci. USA 2011, 108, 19288–19292. [Google Scholar] [CrossRef] [PubMed]
- Bright, M.; Bulgheresi, S. A complex journey: Transmission of microbial symbionts. Nat. Rev. Microbiol. 2010, 8, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Salem, H.; Florez, L.; Gerardo, N.; Kaltenpoth, M. An out-of-body experience: The extracellular dimension for the transmission of mutualistic bacteria in insects. Proc. R. Soc. B Biol. Sci. 2015, 282. [Google Scholar] [CrossRef] [PubMed]
- Caspi-Fluger, A.; Inbar, M.; Mozes-Daube, N.; Katzir, N.; Portnoy, V.; Belausov, E.; Hunter, M.S.; Zchori-Fein, E. Horizontal transmission of the insect symbiont Rickettsia is plant-mediated. Proc. R. Soc. B Biol. Sci. 2012, 279, 1791–1796. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, M.P. Access to mutualistic endosymbiotic microbes: An underappreciated benefit of group living. Behav. Ecol. Sociobiol. 2008, 62, 479–497. [Google Scholar] [CrossRef]
- Barber, I.; Wright, H.A. How strong are familiarity preferences in shoaling fish? Anim. Behav. 2001, 61, 975–979. [Google Scholar] [CrossRef]
- Wiszniewski, J.; Lusseau, D.; Möller, L.M. Female bisexual kinship ties maintain social cohesion in a dolphin network. Anim. Behav. 2010, 80, 895–904. [Google Scholar] [CrossRef]
- Kurvers, R.H.J.M.; Adamczyk, V.M.A.P.; Kraus, R.H.S.; Hoffman, J.I.; van Wieren, S.E.; van der Jeugd, H.P.; Amos, W.; Prins, H.H.T.; Jonker, R.M. Contrasting context dependence of familiarity and kinship in animal social networks. Anim. Behav. 2013, 1–9. [Google Scholar] [CrossRef]
- Nalepa, C.A. Nourishment and the origin of termite eusociality. In Nourishment and Evolution in Insect Societies; Westview Press: Boulder, CO, USA, 1994; pp. 57–104. [Google Scholar]
- Moran, N.A.; Hansen, A.K.; Powell, J.E.; Sabree, Z.L. Distinctive gut microbiota of honey bees assessed using deep sampling from individual worker bees. PLoS ONE 2012, 7, e36393. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.C.; Fruciano, C.; Hildebrand, F.; Al Toufalilia, H.; Balfour, N.J.; Bork, P.; Engel, P.; Ratnieks, F.L.W.; Hughes, W.O.H. Gut microbiota composition is associated with environmental landscape in honey bees. Ecol. Evol. 2018, 8, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Wilson, E.O. The Insect Societies; Harvard University Press: Cambridge, MA, USA, 1971. [Google Scholar]
- Simpson, S.J.; Raubenheimer, D.; Charleston, M.A.; Clissold, F.J. Modelling nutritional interactions: From individuals to communities. Trends Ecol. Evol. 2010, 25, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Simpson, S.J.; Clissold, F.J.; Lihoreau, M.; Ponton, F.; Wilder, S.M.; Raubenheimer, D. Recent Advances in the Integrative Nutrition of Arthropods. Annu. Rev. Entomol. 2015, 60, 293–311. [Google Scholar] [CrossRef] [PubMed]
- Stilling, R.M.; Moloney, G.M.; Ryan, F.J.; Hoban, A.E.; Bastiaanssen, T.F.; Shanahan, F.; Clarke, G.; Claesson, M.J.; Dinan, T.G.; Cryan, J.F. Social interaction-induced activation of RNA splicing in the amygdala of microbiome-deficient mice. eLife 2018, 7, e33070. [Google Scholar] [CrossRef] [PubMed]
- Elgar, M.A. Predator vigilance and group size in mammals and birds: A critical review of the empirical evidence. Biol. Rev. Camb. Philos. Soc. 1989, 64, 13–33. [Google Scholar] [CrossRef] [PubMed]
- Kurvers, R.H.J.M.; van Oers, K.; Nolet, B.A.; Jonker, R.M.; van Wieren, S.E.; Prins, H.H.T.; Ydenberg, R.C. Personality predicts the use of social information. Ecol. Lett. 2010, 13, 829–837. [Google Scholar] [CrossRef] [PubMed]
- Lihoreau, M.; Charleston, M.A.; Senior, A.M.; Clissold, F.J.; Raubenheimer, D.; Simpson, S.J.; Buhl, J. Collective foraging in spatially complex nutritional environments. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160238. [Google Scholar] [CrossRef] [PubMed]
- Storelli, G.; Defaye, A.; Erkosar, B.; Hols, P.; Royet, J.; Leulier, F. Lactobacillus plantarum promotes Drosophila systemic growth by modulating hormonal signals through TOR-dependent nutrient sensing. Cell Metab. 2011, 14, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Callens, M.; Macke, E.; Muylaert, K.; Bossier, P.; Lievens, B.; Waud, M.; Decaestecker, E. Food availability affects the strength of mutualistic host-microbiota interactions in Daphnia magna. ISME J. 2016, 10, 911. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.A.; Raubenheimer, D.; Rothman, J.M.; Clarke, D.; Swedell, L. 30 Days in the life: Daily nutrient balancing in a wild chacma baboon. PLoS ONE 2013, 8, e70383. [Google Scholar] [CrossRef] [PubMed]
- Raubenheimer, D.; Machovsky-Capuska, G.E.; Chapman, C.A.; Rothman, J.M. Geometry of nutrition in field studies: An illustration using wild primates. Oecologia 2015, 177, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Vitousek, M.N.; Zonana, D.M.; Safran, R.J. An integrative view of the signaling phenotype: Dynamic links between signals, physiology, behavior and social context. Curr. Zool. 2014, 60, 739–754. [Google Scholar] [CrossRef]
- Senior, A.M.; Lihoreau, M.; Buhl, J.; Raubenheimer, D.; Simpson, S.J. Social network analysis and nutritional behavior: An integrated modeling approach. Front. Psychol. 2016, 7, 18. [Google Scholar] [CrossRef] [PubMed]
- Pruitt, J.N.; Berdahl, A.; Riehl, C.; Pinter-Wollman, N.; Moeller, H.V.; Pringle, E.G.; Aplin, L.M.; Robinson, E.J.; Grilli, J.; Yeh, P. Social tipping points in animal societies. Proc. R. Soc. B 2018, 285, 20181282. [Google Scholar] [CrossRef] [PubMed]
- Sokolowski, M.B. Foraging strategies of Drosophila melanogaster: A chromosomal analysis. Behav. Genet. 1980, 10, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Pasquaretta, C.; Battesti, M.; Klenschi, E.; Bousquet, C.A.; Sueur, C.; Mery, F. How social network structure affects decision-making in Drosophila melanogaster. Proc. Biol. Sci. 2016, 283. [Google Scholar] [CrossRef] [PubMed]
- Kort, R.; Caspers, M.; van de Graaf, A.; van Egmond, W.; Keijser, B.; Roeselers, G. Shaping the oral microbiota through intimate kissing. Microbiome 2014, 2, 41. [Google Scholar] [CrossRef] [PubMed]
- Alberdi, A.; Aizpurua, O.; Bohmann, K.; Zepeda-Mendoza, M.L.; Gilbert, M.T.P. Do vertebrate gut metagenomes confer rapid ecological adaptation? Trends Ecol. Evol. 2016, 31, 689–699. [Google Scholar] [CrossRef] [PubMed]
- Cho, I.; Yamanishi, S.; Cox, L.; Methé, B.A.; Zavadil, J.; Li, K.; Gao, Z.; Mahana, D.; Raju, K.; Teitler, I. Antibiotics in early life alter the murine colonic microbiome and adiposity. Nature 2012, 488, 621. [Google Scholar] [CrossRef] [PubMed]
- Delsuc, F.; Metcalf, J.L.; Wegener Parfrey, L.; Song, S.J.; González, A.; Knight, R. Convergence of gut microbiomes in myrmecophagous mammals. Mol. Ecol. 2014, 23, 1301–1317. [Google Scholar] [CrossRef] [PubMed]
- Roggenbuck, M.; Schnell, I.B.; Blom, N.; Bælum, J.; Bertelsen, M.F.; Sicheritz-Pontén, T.; Sørensen, S.J.; Gilbert, M.T.P.; Graves, G.R.; Hansen, L.H. The microbiome of New World vultures. Nat. Commun. 2014, 5, 5498. [Google Scholar] [CrossRef] [PubMed]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pasquaretta, C.; Gómez-Moracho, T.; Heeb, P.; Lihoreau, M. Exploring Interactions between the Gut Microbiota and Social Behavior through Nutrition. Genes 2018, 9, 534. https://doi.org/10.3390/genes9110534
Pasquaretta C, Gómez-Moracho T, Heeb P, Lihoreau M. Exploring Interactions between the Gut Microbiota and Social Behavior through Nutrition. Genes. 2018; 9(11):534. https://doi.org/10.3390/genes9110534
Chicago/Turabian StylePasquaretta, Cristian, Tamara Gómez-Moracho, Philipp Heeb, and Mathieu Lihoreau. 2018. "Exploring Interactions between the Gut Microbiota and Social Behavior through Nutrition" Genes 9, no. 11: 534. https://doi.org/10.3390/genes9110534
APA StylePasquaretta, C., Gómez-Moracho, T., Heeb, P., & Lihoreau, M. (2018). Exploring Interactions between the Gut Microbiota and Social Behavior through Nutrition. Genes, 9(11), 534. https://doi.org/10.3390/genes9110534