Insights into Drought Stress Signaling in Plants and the Molecular Genetic Basis of Cotton Drought Tolerance


 and
and
Abstract
1. Introduction
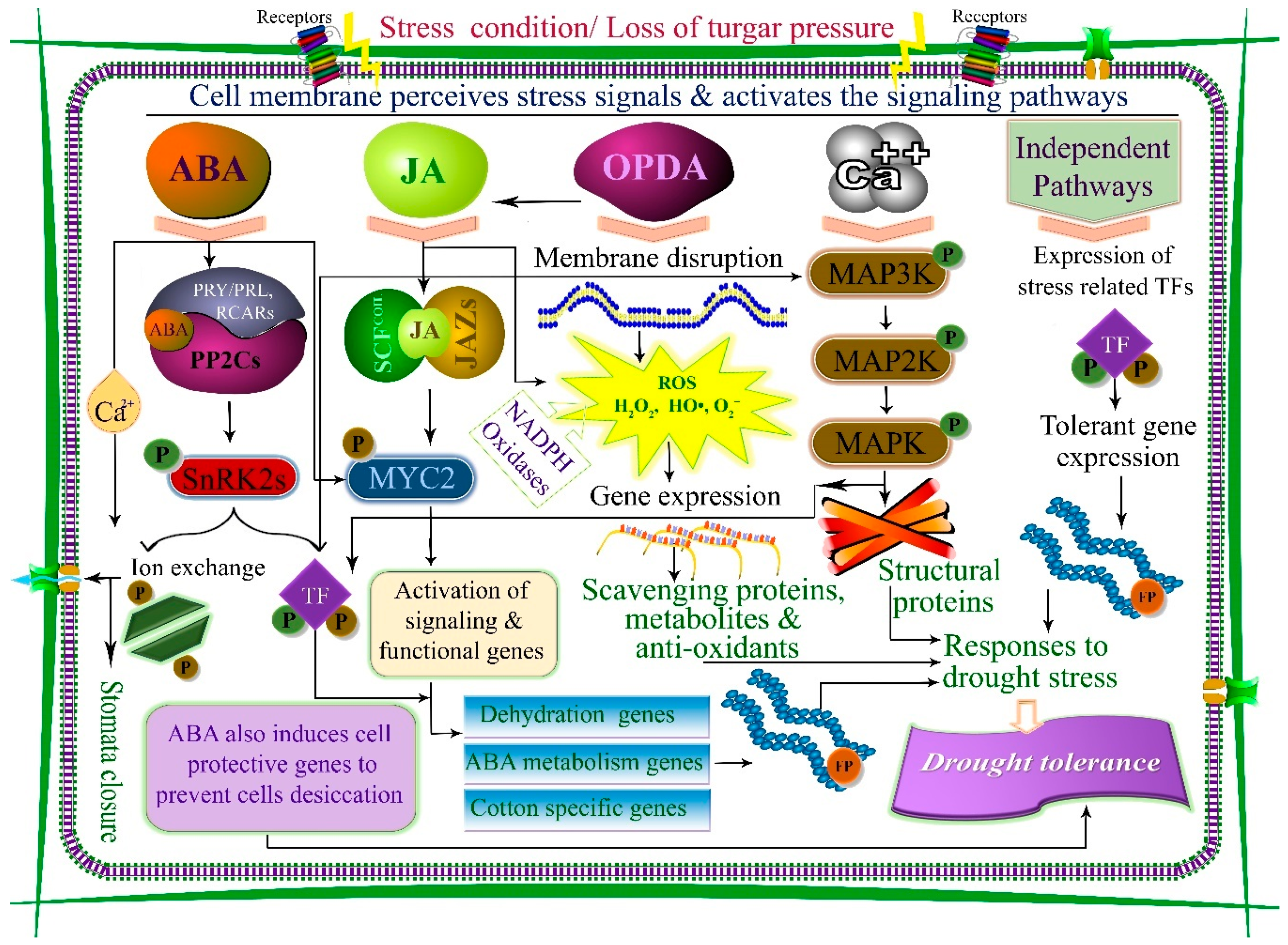
2. Cellular and Molecular Signaling Pathways of Drought Stress Tolerances
2.1. Mitogen Activated Protein Kinase (MAPK) Signaling Pathway
2.2. Ca2+ Signaling Pathway
2.3. Abscisic Acid (Aba)-Mediated Signaling Pathway
3. Role of TFs in Drought Stress Signaling Pathways
4. Cellular and Molecular Responses to Drought Stress in Plants
4.1. Morpho-Physiological Responses
4.2. Biochemical and Cellular Responses
4.3. Antioxidant Defense Against Cellular Reactive Oxygen Species (ROS)
5. Molecular Genetic Basis and QTLs for Drought Tolerance in Cotton
6. Application of Genome-Modification Approaches to Achieve Drought Tolerance in Plants
7. New Functional Genomic Tools to Identify Novel Genes for Stress Tolerance in Plants
8. Functional Genomics for Stress Tolerance
9. Future Perspectives and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Saranga, Y.; Paterson, A.H.; Levi, A. Bridging Classical and Molecular Genetics of Abiotic Stress Resistance in Cotton. Genet. Genom. Cott. 2009, 3, 337–352. [Google Scholar]
- Abdelraheem, A.; Esmaeili, N.; O’Connell, M.; Zhang, J. Progress and perspective on drought and salt stress tolerance in cotton. Ind. Crops Prod. 2019, 130, 118–129. [Google Scholar] [CrossRef]
- Ullah, A.; Sun, H.; Yang, X.; Zhang, X. Drought coping strategies in cotton: Increased crop per drop. Plant Biotechnol. J. 2017, 15, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Kazuya Ichimura, M.G.; Ichimura, K.; Shinozaki, K.; Tena, G.; Sheen, J.; Henry, Y.; Champion, A.; Kreis, M.; Zhang, S.; Hirt, H.; et al. Mitogen-activated protein kinase cascades in plants: A new nomenclature. Trends Plant Sci. 2002, 7, 301–308. [Google Scholar] [CrossRef]
- Li, L.; Yu, D.; Zhao, F.; Pang, C.; Song, M.; Wei, H.; Fan, S.; Yu, S. Genome-wide analysis of the calcium-dependent protein kinase gene family in Gossypium raimondii. J. Integr. Agric. 2015, 14, 29–41. [Google Scholar] [CrossRef]
- Dong, T.; Park, Y.; Hwang, I. Abscisic acid: Biosynthesis, inactivation, homoeostasis and signalling. Essays Biochem. 2015, 58, 29–48. [Google Scholar]
- Yoshida, T.; Mogami, J.; Yamaguchi-Shinozaki, K. ABA-dependent and ABA-independent signaling in response to osmotic stress in plants. Curr. Opin. Plant Biol. 2014, 21, 133–139. [Google Scholar] [CrossRef]
- Danquah, A.; de Zelicourt, A.; Colcombet, J.; Hirt, H. The role of ABA and MAPK signaling pathways in plant abiotic stress responses. Biotechnol. Adv. 2014, 32, 40–52. [Google Scholar] [CrossRef]
- Riemann, M.; Dhakarey, R.; Hazman, M.; Miro, B.; Kohli, A.; Nick, P. Exploring Jasmonates in the Hormonal Network of Drought and Salinity Responses. Front. Plant Sci. 2015, 6, 1–16. [Google Scholar] [CrossRef]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell. Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef]
- Chastain, D.R.; Snider, J.L.; Choinski, J.S.; Collins, G.D.; Perry, C.D.; Whitaker, J.; Grey, T.L.; Sorensen, R.B.; van Iersel, M.; Byrd, S.A.; et al. Leaf ontogeny strongly influences photosynthetic tolerance to drought and high temperature in Gossypium hirsutum. J. Plant Physiol. 2016, 199, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Hejnák, V.; Tatar; Atasoy, G.D.; Martinková, J.; Çelen, A.E.; Hnilička, F.; Skalický, M. Growth and photosynthesis of upland and pima cotton: Response to drought and heat stress. Plant Soil Environ. 2015, 62, 507–514. [Google Scholar]
- Long, L.; Guo, D.-D.; Gao, W.; Yang, W.-W.; Hou, L.-P.; Ma, X.-N.; Miao, Y.-C.; Botella, J.R.; Song, C.-P. Optimization of CRISPR/Cas9 genome editing in cotton by improved sgRNA expression. Plant Methods 2018, 14, 85. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, J.; Zuo, D.; Wang, Q.; Malik, W.; Zhang, Y.; Abid, M.A.; Cheng, H.; Yang, Q.; Song, G. Recent insights into cotton functional genomics: Progress and future perspectives. Plant Biotechnol. J. 2018, 16, 699–713. [Google Scholar] [CrossRef]
- Popescu, S.C.; Popescu, G.V.; Bachan, S.; Zhang, Z.; Gerstein, M.; Snyder, M.; Dinesh-Kumar, S.P. MAPK target networks in Arabidopsis thaliana revealed using functional protein microarrays. Genes Dev. 2009, 23, 80–92. [Google Scholar] [CrossRef]
- Nakagami, H.; Pitzschke, A.; Hirt, H. Emerging MAP kinase pathways in plant stress signalling. Trends Plant Sci. 2005, 10, 339–346. [Google Scholar] [CrossRef]
- Mitula, F.; Tajdel, M.; Cieśla, A.; Kasprowicz-Maluśki, A.; Kulik, A.; Babula-Skowrońska, D.; Michalak, M.; Dobrowolska, G.; Sadowski, J.; Ludwików, A. Arabidopsis ABA-Activated Kinase MAPKKK18 is Regulated by Protein Phosphatase 2C ABI1 and the Ubiquitin-Proteasome Pathway. Plant Cell Physiol. 2015, 56, 2351–2367. [Google Scholar] [CrossRef]
- Li, Y.; Cai, H.; Liu, P.; Wang, C.; Gao, H.; Wu, C.; Yan, K.; Zhang, S.; Huang, J.; Zheng, C. Arabidopsis MAPKKK18 positively regulates drought stress resistance via downstream MAPKK3. Biochem. Biophys. Res. Commun. 2017, 484, 292–297. [Google Scholar] [CrossRef]
- Tajdel, M.; Mituła, F.; Ludwików, A. Regulation of Arabidopsis MAPKKK18 by ABI1 and SnRK2, components of the ABA signaling pathway. Plant Signal. Behav. 2016, 11, e1139277. [Google Scholar] [CrossRef]
- Danquah, A.; de Zélicourt, A.; Boudsocq, M.; Neubauer, J.; Frei Dit Frey, N.; Leonhardt, N.; Pateyron, S.; Gwinner, F.; Tamby, J.-P.; Ortiz-Masia, D.; et al. Identification and characterization of an ABA-activated MAP kinase cascade in Arabidopsis thaliana. Plant J. 2015, 82, 232–244. [Google Scholar] [CrossRef]
- Matsuoka, D.; Yasufuku, T.; Furuya, T.; Nanmori, T. An abscisic acid inducible Arabidopsis MAPKKK, MAPKKK18 regulates leaf senescence via its kinase activity. Plant Mol. Biol. 2015, 87, 565–575. [Google Scholar] [CrossRef]
- Liu, B.; Sun, L.; Ma, L.; Hao, F.S. Both AtrbohD and AtrbohF are essential for mediating responses to oxygen deficiency in Arabidopsis. Plant Cell Rep. 2017, 36, 947–957. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Li, Y.; Miao, W.; Piao, T.; Hao, Y.; Hao, F.S. NADK2 positively modulates abscisic acid-induced stomatal closure by affecting accumulation of H2O2, Ca2+ and nitric oxide in Arabidopsis guard cells. Plant Sci. 2017, 262, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Dongdong, L.; Ming, Z.; Lili, H.; Xiaobo, C.; Yang, G.; Xingqi, G.; Han, L. GhMAPKKK49, a novel cotton (Gossypium hirsutum L.) MAPKKK gene, is involved in diverse stress responses. Acta Physiol. Plant. 2016, 38, 1–12. [Google Scholar] [CrossRef]
- Wang, C.; Lu, W.; He, X.; Wang, F.; Zhou, Y.; Guo, X.X.; Guo, X.X. The Cotton Mitogen-Activated Protein Kinase Kinase 3 Functions in Drought Tolerance by Regulating Stomatal Responses and Root Growth. Plant Cell Physiol. 2016, 57, 1629–1642. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Li, M.; Wang, P.; Cox, K.L.; Duan, L.; Dever, J.K.; Shan, L.; Li, Z.; He, P. Regulation of cotton (Gossypium hirsutum) drought responses by mitogen-activated protein (MAP) kinase cascade-mediated phosphorylation of GhWRKY59. New Phytol. 2017, 215, 1462–1475. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Mi, X.; Chen, C.; Wang, H.; Guo, W. Identification on mitogen-Activated protein kinase signaling cascades by integrating protein interaction with transcriptional profiling analysis in cotton. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Tu, L.; Yang, X.; Tan, J.; Deng, F.; Hao, J.; Guo, K.; Lindsey, K.; Zhang, X. The calcium sensor GhCaM7 promotes cotton fiber elongation by modulating reactive oxygen species (ROS) production. New Phytol. 2014, 202, 509–520. [Google Scholar] [CrossRef]
- Huang, S.; Wang, H.-Y.; Gao, P.; Wang, G.-Y.; Xia, G.-X. Cloning and characterization of a calcium dependent protein kinase gene associated with cotton fiber development. Plant Cell Rep. 2008, 27, 1869–1875. [Google Scholar] [CrossRef]
- He, L.; Yang, X.; Wang, L.; Zhu, L.; Zhou, T.; Deng, J.; Zhang, X. Molecular cloning and functional characterization of a novel cotton CBL-interacting protein kinase gene (GhCIPK6) reveals its involvement in multiple abiotic stress tolerance in transgenic plants. Biochem. Biophys. Res. Commun. 2013, 435, 209–215. [Google Scholar] [CrossRef]
- Lee, S.C.; Luan, S. ABA signal transduction at the crossroad of biotic and abiotic stress responses. Plant Cell Environ. 2012, 35, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhao, Y.; Li, Z.; Hsu, C.C.; Liu, X.; Fu, L.; Hou, Y.J.; Du, Y.; Xie, S.; Zhang, C.; et al. Reciprocal Regulation of the TOR Kinase and ABA Receptor Balances Plant Growth and Stress Response. Mol. Cell 2018, 69, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Fujita, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. ABA-mediated transcriptional regulation in response to osmotic stress in plants. J. Plant Res. 2011, 124, 509–525. [Google Scholar] [CrossRef] [PubMed]
- Cutler, S.R.; Rodriguez, P.L.; Finkelstein, R.R.; Abrams, S.R. Abscisic Acid: Emergence of a Core Signaling Network. Annu. Rev. Plant Biol. 2010, 61, 651–679. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, Y.; Wen, F.; Yao, D.; Wang, L.; Guo, J.; Ni, L.; Zhang, A.; Tan, M.; Jiang, M. A novel rice C2H2-type zinc finger protein, ZFP36, is a key player involved in abscisic acid-induced antioxidant defence and oxidative stress tolerance in rice. J. Exp. Bot. 2014, 65, 5795–5809. [Google Scholar] [CrossRef]
- Jagodzik, P.; Tajdel-Zielinska, M.; Ciesla, A.; Marczak, M.; Ludwikow, A. Mitogen-activated protein kinase cascades in plant hormone signaling. Front. Plant Sci. 2018, 9, 1–26. [Google Scholar] [CrossRef]
- Guo, M.; Liu, J.-H.; Ma, X.; Luo, D.-X.; Gong, Z.-H.; Lu, M.-H. The Plant Heat Stress Transcription Factors (HSFs): Structure, Regulation, and Function in Response to Abiotic Stresses. Front. Plant Sci. 2016, 7, 114. [Google Scholar] [CrossRef]
- Lata, C.; Prasad, M. Role of DREBs in regulation of abiotic stress responses in plants. J. Exp. Bot. 2011, 62, 4731–4748. [Google Scholar] [CrossRef]
- Liang, C.; Meng, Z.Z.; Meng, Z.Z.; Malik, W.; Yan, R.; Lwin, K.M.; Lin, F.; Wang, Y.; Sun, G.; Zhou, T.; et al. GhABF2, a bZIP transcription factor, confers drought and salinity tolerance in cotton (Gossypium hirsutum L.). Sci. Rep. 2016, 6, 1–14. [Google Scholar] [CrossRef]
- Chen, T.; Li, W.; Hu, X.; Guo, J.; Liu, A.; Zhang, B. A cotton MYB transcription factor, GbMYB5, is positively involved in plant adaptive response to drought stress. Plant Cell Physiol. 2014, 56, 917–929. [Google Scholar] [CrossRef]
- Chu, X.; Wang, C.; Chen, X.; Lu, W.; Li, H.; Wang, X.; Hao, L.; Guo, X. The Cotton WRKY Gene GhWRKY41 Positively Regulates Salt and Drought Stress Tolerance in Transgenic Nicotiana benthamiana. PLoS ONE 2015, 10, e0143022. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Pang, C.; Jia, X.; Ma, Q.; Dou, L.; Zhao, F.; Gu, L.; Wei, H.; Wang, H.; Fan, S.; et al. An NAM Domain Gene, GhNAC79, Improves Resistance to Drought Stress in Upland Cotton. Front. Plant Sci. 2017, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Feng, H.; Zhang, X.; Zhang, C.; Wang, T. An Arabidopsis E3 ligase HUB2 increases histone H2B monoubiquitination and enhances drought tolerance in transgenic cotton. Plant Biotechnol. J. 2019, 17, 556–568. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, J.; Zhu, M.; Jia, H.; Liu, D.; Hao, L.; Guo, X. A cotton Raf-like MAP3K gene, GhMAP3K40, mediates reduced tolerance to biotic and abiotic stress in Nicotiana benthamiana by negatively regulating growth and development. Plant Sci. 2015, 240, 10–24. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.-N.; Zhao, L.; Lu, R.; Li, Y.; Li, X.-B. Cotton mitogen-activated protein kinase4 (GhMPK4) confers the transgenic Arabidopsis hypersensitivity to salt and osmotic stresses. Plant Cell Tissue Organ Cult. 2015, 123, 619–632. [Google Scholar] [CrossRef]
- Zhang, J.; Zou, D.; Li, Y.; Sun, X.; Wang, N.N.; Gong, S.Y.; Zheng, Y.; Li, X.B. GhMPK17, a cotton mitogen-activated protein kinase, is involved in plant response to high salinity and osmotic stresses and ABA signaling. PLoS ONE 2014, 9, 1–12. [Google Scholar] [CrossRef]
- Long, L.; Gao, W.; Xu, L.; Liu, M.; Luo, X.; He, X.; Yang, X.; Zhang, X.; Zhu, L. GbMPK3, a mitogen-activated protein kinase from cotton, enhances drought and oxidative stress tolerance in tobacco. Plant Cell Tissue Organ Cult. 2014, 116, 153–162. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, L.; Wang, X.; Zhang, W.; Hao, L.; Chu, X.; Guo, X. Cotton GhMPK6a negatively regulates osmotic tolerance and bacterial infection in transgenic Nicotiana benthamiana, and plays a pivotal role in development. FEBS J. 2013, 280, 5128–5144. [Google Scholar] [CrossRef]
- Lu, W.; Chu, X.; Li, Y.; Wang, C.; Guo, X. Cotton GhMKK1 Induces the Tolerance of Salt and Drought Stress, and Mediates Defence Responses to Pathogen Infection in Transgenic Nicotiana benthamiana. PLoS ONE 2013, 8, e68503. [Google Scholar] [CrossRef]
- Zhang, L.; Li, Y.; Lu, W.; Meng, F.; Wu, C.; Guo, X. Cotton GhMKK5 affects disease resistance, induces HR-like cell death, and reduces the tolerance to salt and drought stress in transgenic Nicotiana benthamiana. J. Exp. Bot. 2012, 63, 3935–3951. [Google Scholar] [CrossRef]
- Zhang, L.; Xi, D.; Li, S.; Gao, Z.; Zhao, S.; Shi, J.; Wu, C.; Guo, X. A cotton group C MAP kinase gene, GhMPK2, positively regulates salt and drought tolerance in tobacco. Plant Mol. Biol. 2011, 77, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Gao, Y.; Zhang, Z.; Chen, T.; Guo, W.; Zhang, T. A receptor-like kinase gene (GbRLK) from Gossypium barbadense enhances salinity and drought-stress tolerance in Arabidopsis. BMC Plant Biol. 2013, 13, 110. [Google Scholar] [CrossRef]
- Chen, E.; Zhang, X.X.; Yang, Z.Z.; Wang, X.; Yang, Z.Z.; Zhang, C.; Wu, Z.; Kong, D.; Liu, Z.; Zhao, G.; et al. Genome-wide analysis of the HD-ZIP IV transcription factor family in Gossypium arboreum and GaHDG11 involved in osmotic tolerance in transgenic Arabidopsis. Mol. Genet. Genom. 2017, 292, 593–609. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Hu, L.; Fan, J.; Amombo, E.; Khaldun, A.B.M.; Zheng, Y.; Chen, L. Cotton GhERF38 gene is involved in plant response to salt/drought and ABA. Ecotoxicology 2017, 26, 841–854. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.G.; Li, H.; Liu, J.Y. Molecular Characterization of Three Ethylene Responsive Element Binding Factor Genes from Cotton. J. Integr. Plant Biol. 2010, 52, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Song, Y.; Xing, F.; Wang, N.; Wen, F.; Zhu, C. GhWRKY25, a group I WRKY gene from cotton, confers differential tolerance to abiotic and biotic stresses in transgenic Nicotiana benthamiana. Protoplasma 2016, 253, 1265–1281. [Google Scholar] [CrossRef]
- Gunapati, S.; Naresh, R.; Ranjan, S.; Nigam, D.; Hans, A.; Verma, P.C.; Gadre, R.; Pathre, U.V.; Sane, A.P.; Sane, V.A. Expression of GhNAC2 from G. herbaceum, improves root growth and imparts tolerance to drought in transgenic cotton and Arabidopsis. Sci. Rep. 2016, 6, 24978. [Google Scholar] [CrossRef]
- Ma, L.-F.; Li, Y.; Chen, Y.; Li, X.-B. Improved drought and salt tolerance of Arabidopsis thaliana by ectopic expression of a cotton (Gossypium hirsutum) CBF gene. Plant Cell Tissue Organ Cult. 2016, 124, 583–598. [Google Scholar] [CrossRef]
- Huang, G.-Q.; Li, W.; Zhou, W.; Zhang, J.-M.; Li, D.-D.; Gong, S.-Y.; Li, X.-B. Seven cotton genes encoding putative NAC domain proteins are preferentially expressed in roots and in responses to abiotic stress during root development. Plant Growth Regul. 2013, 71, 101–112. [Google Scholar] [CrossRef]
- Yan, H.; Jia, H.; Chen, X.; Hao, L.; An, H.; Guo, X. The Cotton WRKY Transcription Factor GhWRKY17 Functions in Drought and Salt Stress in Transgenic Nicotiana benthamiana Through ABA Signaling and the Modulation of Reactive Oxygen Species Production. Plant Cell Physiol. 2014, 55, 2060–2076. [Google Scholar] [CrossRef]
- Shah, S.T.; Pang, C.; Fan, S.; Song, M.; Arain, S.; Yu, S. Isolation and expression profiling of GhNAC transcription factor genes in cotton (Gossypium hirsutum L.) during leaf senescence and in response to stresses. Gene 2013, 531, 220–234. [Google Scholar] [CrossRef] [PubMed]
- Meng, C.; Cai, C.; Zhang, T.; Guo, W. Characterization of six novel NAC genes and their responses to abiotic stresses in Gossypium hirsutum L. Plant Sci. 2009, 176, 352–359. [Google Scholar] [CrossRef]
- Gao, S.-Q.Q.; Chen, M.; Xia, L.-Q.Q.; Xiu, H.-J.J.; Xu, Z.-S.S.; Li, L.-C.C.; Zhao, C.-P.P.; Cheng, X.-G.G.; Ma, Y.-Z.Z. A cotton (Gossypium hirsutum) DRE-binding transcription factor gene, GhDREB, confers enhanced tolerance to drought, high salt, and freezing stresses in transgenic wheat. Plant Cell Rep. 2009, 28, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.-G.; Yang, M.; Liu, P.; Yang, G.-D.; Wu, C.-A.; Zheng, C.-C. GhDREB1 enhances abiotic stress tolerance, delays GA-mediated development and represses cytokinin signalling in transgenic Arabidopsis. Plant Cell Environ. 2009, 32, 1132–1145. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Jin, L.; Liu, J.-Y. Identification and characterization of the novel gene GhDBP2 encoding a DRE-binding protein from cotton (Gossypium hirsutum). J. Plant Physiol. 2008, 165, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Z.; Huang, B.; Liu, J. Molecular cloning and functional analysis of an ERF gene from cotton (Gossypium hirsutum). Biochim. Biophys. Acta Gene Regul. Mech. 2008, 1779, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.G.; Liu, J.Y. Molecular cloning, expression profile and promoter analysis of a novel ethylene responsive transcription factor gene GhERF4 from cotton (Gossypium hirstum). Plant Physiol. Biochem. 2008, 46, 46–53. [Google Scholar] [CrossRef]
- Huang, B.; Jin, L.; Liu, J. Molecular cloning and functional characterization of a DREB1/CBF-like gene (GhDREB1L) from cotton. Sci. China Ser. C Life Sci. 2007, 50, 7–14. [Google Scholar] [CrossRef]
- Liang, C.; Liu, Y.; Li, Y.; Meng, Z.; Yan, R.; Zhu, T.; Wang, Y.; Kang, S.; Ali Abid, M.; Malik, W.; et al. Activation of ABA Receptors Gene GhPYL9-11A Is Positively Correlated with Cotton Drought Tolerance in Transgenic Arabidopsis. Front. Plant Sci. 2017, 8, 1–13. [Google Scholar] [CrossRef]
- Bello, B.; Zhang, X.; Liu, C.; Yang, Z.; Yang, Z.; Wang, Q.; Zhao, G.; Li, F. Cloning of Gossypium hirsutum sucrose non-fermenting 1-related protein kinase 2 gene (GhSnRK2) and its overexpression in transgenic Arabidopsis escalates drought and low temperature tolerance. PLoS ONE 2014, 9, 1–18. [Google Scholar] [CrossRef][Green Version]
- Magwanga, R.O.; Lu, P.; Kirungu, J.N.; Cai, X.; Zhou, Z.; Wang, X.; Diouf, L.; Xu, Y.; Hou, Y.; Hu, Y.; et al. Whole genome analysis of cyclin dependent kinase (CDK) gene family in cotton and functional evaluation of the role of CDKF4 gene in drought and salt stress tolerance in plants. Int. J. Mol. Sci. 2018, 19, 2625. [Google Scholar] [CrossRef] [PubMed]
- Butt, H.I.; Yang, Z.; Chen, E.; Zhao, G.; Gong, Q.; Yang, Z.; Zhang, X.; Li, F. Functional Characterization of Cotton GaMYB62L, a Novel R2R3 TF in Transgenic Arabidopsis. PLoS ONE 2017, 12, e0170578. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, S.; Qi, S.; Zheng, C.; Wu, C. Delayed germination of Arabidopsis seeds under chilling stress by overexpressing an abiotic stress inducible GhTPS11. Gene 2016, 575, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Shen, G.; Kuppu, S.; Gaxiola, R.; Payton, P. Creating drought- and salt-tolerant cotton by overexpressing a vacuolar pyrophosphatase gene. Plant Signal. Behav. 2011, 6, 861–863. [Google Scholar] [CrossRef] [PubMed]
- Zafar, S.A.; Patil, S.B.; Uzair, M.; Fang, J.; Zhao, J.; Guo, T.; Yuan, S.; Uzair, M.; Luo, Q.; Shi, J.; et al. DEGENERATED PANICLE AND PARTIAL STERILITY 1 (DPS 1) encodes a cystathionine β-synthase domain containing protein required for anther cuticle and panicle development in rice. New Phytol. 2019, 1, 356–375. [Google Scholar] [CrossRef] [PubMed]
- De Brito, G.G.; Sofiatti, V.; de Andrade Lima, M.M.; de Carvalho, L.P.; da Silva Filho, J.L. Physiological traits for drought phenotyping in cotton Giovani. Acta Sci. Agron. 2011, 33, 117–125. [Google Scholar] [CrossRef]
- Shavrukov, Y.; Kurishbayev, A.; Jatayev, S.; Shvidchenko, V.; Zotova, L.; Koekemoer, F.; de Groot, S.; Soole, K.; Langridge, P. Early Flowering as a Drought Escape Mechanism in Plants: How Can It Aid Wheat Production? Front. Plant Sci. 2017, 8, 1950. [Google Scholar] [CrossRef]
- Luo, L.J. Breeding for water-saving and drought-resistance rice (WDR) in China. J. Exp. Bot. 2010, 61, 3509–3517. [Google Scholar] [CrossRef]
- Cal Andrew, J.; Henry, A.; McNally, K.L.; Luquet, D.; Sanciangco, M.; Torres, R.O.; Rebolledo, M.C.; Cal, A.J. Leaf morphology, rather than plant water status, underlies genetic variation of rice leaf rolling under drought. Plant Cell Environ. 2019, 42, 1–13. [Google Scholar]
- Hetherington, A.M.; Woodward, F.I. The role of stomata in sensing and driving environmental change. Nature 2003, 424, 901–908. [Google Scholar] [CrossRef]
- Levi, A.; Paterson, A.H.; Chee, P.W.; Saranga, Y.; Chee, P.W. Field evaluation of cotton near-isogenic lines introgressed with QTLs for productivity and drought related traits. Mol. Breed. 2009, 23, 179–195. [Google Scholar] [CrossRef]
- Singh, M.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Roles of osmoprotectants in improving salinity and drought tolerance in plants: A review. Rev. Environ. Sci. Biol. 2015, 14, 407–426. [Google Scholar] [CrossRef]
- Khan, M.S.M.A.; Ahmad, D.; Khan, M.S.M.A. Utilization of genes encoding osmoprotectants in transgenic plants for enhanced abiotic stress tolerance. Electron. J. Biotechnol. 2015, 18, 257–266. [Google Scholar] [CrossRef]
- Divya, K.; Jami, S.K.; Kirti, P.B. Constitutive expression of mustard annexin, AnnBj1 enhances abiotic stress tolerance and fiber quality in cotton under stress. Plant Mol. Biol. 2010, 73, 293–308. [Google Scholar] [CrossRef] [PubMed]
- Lv, S.; Yang, A.; Zhang, K.; Wang, L.; Zhang, J. Increase of glycinebetaine synthesis improves drought tolerance in cotton. Mol. Breed. 2007, 20, 233–248. [Google Scholar] [CrossRef]
- Zhang, F.; Li, S.; Yang, S.; Wang, L.; Guo, W. Overexpression of a cotton annexin gene, GhAnn1, enhances drought and salt stress tolerance in transgenic cotton. Plant Mol. Biol. 2014, 87, 47–67. [Google Scholar] [CrossRef] [PubMed]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef]
- Luo, H.H.; Zhang, Y.L.; Zhang, W.F. Effects of water stress and rewatering on photosynthesis, root activity, and yield of cotton with drip irrigation under mulch. Photosynthetica 2016, 54, 65–73. [Google Scholar] [CrossRef]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Gene networks involved in drought stress response and tolerance. J. Exp. Bot. 2006, 58, 221–227. [Google Scholar] [CrossRef]
- Kuromori, T.; Miyaji, T.; Yabuuchi, H.; Shimizu, H.; Sugimoto, E.; Kamiya, A.; Moriyama, Y.; Shinozaki, K. ABC transporter AtABCG25 is involved in abscisic acid transport and responses. Proc. Natl. Acad. Sci. USA 2010, 107, 2361–2366. [Google Scholar] [CrossRef]
- Seo, M.; Koshiba, T. Complex regulation of ABA biosynthesis in plants. Trends Plant Sci. 2002, 7, 41–48. [Google Scholar] [CrossRef]
- Ma, L.-F.; Li, Y.; Chen, Y.; Li, X.-B. Plant cell, tissue, and organ culture. Plant Cell Tissue Organ Cult. 2016, 124, 583–598. [Google Scholar] [CrossRef]
- Wasternack, C. Jasmonates: An update on biosynthesis, signal transduction and action in plant stress response, growth and development. Ann. Bot. 2007, 100, 681–697. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, Z.H.; Feng, L.; Zheng, Y.; Li, D.D.; Li, X.B. Genome-wide functional analysis of cotton (Gossypium hirsutum) in response to drought. PLoS ONE 2013, 8, 1–11. [Google Scholar] [CrossRef]
- Chini, A.; Fonseca, S.; Fernández, G.; Adie, B.; Chico, J.M.; Lorenzo, O.; García-Casado, G.; López-Vidriero, I.; Lozano, F.M.; Ponce, M.R.; et al. The JAZ family of repressors is the missing link in jasmonate signalling. Nature 2007, 448, 666–671. [Google Scholar] [CrossRef]
- Hussain, H.A.; Men, S.; Hussain, S.; Chen, Y.; Ali, S.; Zhang, S.; Zhang, K.; Li, Y.; Xu, Q.; Liao, C.; et al. Interactive effects of drought and heat stresses on morpho-physiological attributes, yield, nutrient uptake and oxidative status in maize hybrids. Sci. Rep. 2019, 9, 3890. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Noctor, G.; Veljovic-Jovanovic, S.; Driscoll, S.; Novitskaya, L.; Foyer, C.H. Drought and oxidative load in the leaves of C3plants: A predominant role for photorespiration? Ann. Bot. 2002, 89, 841–850. [Google Scholar] [CrossRef]
- Uzilday, B.; Turkan, I.; Sekmen, A.H.; Ozgur, R.; Karakaya, H.C. Comparison of ROS formation and antioxidant enzymes in Cleome gynandra (C4) and Cleome spinosa (C3) under drought stress. Plant Sci. 2012, 182, 59–70. [Google Scholar] [CrossRef]
- Wu, S.; Hu, C.; Tan, Q.; Li, L.; Shi, K.; Zheng, Y.; Sun, X. Drought stress tolerance mediated by zinc-induced antioxidative defense and osmotic adjustment in cotton (Gossypium Hirsutum). Acta Physiol. Plant. 2015, 37, 167. [Google Scholar] [CrossRef]
- Azevedo Neto, A.; Prisco, J.T.; Enéas-Filho, J.; Eduardo Braga de Abreu, C.; Gomes-Filho, E. Effect of Salt Stress on Antioxidative Enzymes and Lipid Peroxidation in Leaves and Roots of Salt-Tolerant and Salt-Sensitive Maize Genotypes. Environ. Exp. Bot. 2006, 56, 87–94. [Google Scholar] [CrossRef]
- Szalai, G.; Kellős, T.; Galiba, G.; Kocsy, G. Glutathione as an Antioxidant and Regulatory Molecule in Plants Under Abiotic Stress Conditions. J. Plant Growth Regul. 2009, 28, 66–80. [Google Scholar] [CrossRef]
- Chan, K.X.; Wirtz, M.; Phua, S.Y.; Estavillo, G.M.; Pogson, B.J. Balancing metabolites in drought: The sulfur assimilation conundrum. Trends Plant Sci. 2013, 18, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Peng, J.; Chen, T.T.; Zhao, X.H.; Zhang, S.P.; Liu, S.D.; Dong, H.L.; Feng, L.; Yu, S.X. Effect of Drought Stress on Lipid Peroxidation and Proline Content in Cotton Roots. J. Anim. Plant Sci. 2014, 24, 1729–1736. [Google Scholar]
- Sekmen, A.H.; Ozgur, R.; Uzilday, B.; Turkan, I. Reactive oxygen species scavenging capacities of cotton (Gossypium hirsutum) cultivars under combined drought and heat induced oxidative stress. Environ. Exp. Bot. 2014, 99, 141–149. [Google Scholar] [CrossRef]
- Kaur, G.; Asthir, B. Molecular responses to drought stress in plants. Biol. Plant. 2017, 61, 201–209. [Google Scholar] [CrossRef]
- Zhang, J.; Li, X.; Lin, H.; Chong, K. Crop Improvement Through Temperature Resilience. Annu. Rev. Plant Biol. 2019, 70, 753–780. [Google Scholar] [CrossRef]
- Reinisch, A.J.; Dong, J.M.; Brubaker, C.L.; Stelly, D.M.; Wendel, J.F.; Paterson, A.H. A detailed RFLP map of cotton, Gossypium hirsutum x Gossypium barbadense: Chromosome organization and evolution in a disomic polyploid genome. Genetics 1994, 138, 829–847. [Google Scholar]
- Saleem, M.A.; Malik, T.A.; Shakeel, A.; Ashraf, M. QTL mapping for some important drought tolerant traits in upland cotton. J. Anim. Plant Sci. 2015, 25, 502–509. [Google Scholar]
- Saranga, Y.; Jiang, C.; Wright, R.J.; Yakir, D.; Paterson, A.H. Genetic dissection of cotton physiological responses to arid conditions and their inter-relationships with productivity. Plant Cell Environ. 2004, 27, 263–277. [Google Scholar] [CrossRef]
- Pettigrew, W.T.; Heitholt, J.J.; Vaughn, K.C. Gas Exchange Differences and Comparative Anatomy among Cotton Leaf-type Isolines. Crop Sci. 1993, 33, 1295. [Google Scholar] [CrossRef]
- Jiang, C.; Wright, R.J.; Woo, S.S.; DelMonte, T.A.; Paterson, A.H. QTL analysis of leaf morphology in tetraploid Gossypium (cotton). Theor. Appl. Genet. 2000, 100, 409–418. [Google Scholar] [CrossRef]
- Zheng, J.Y.; Oluoch, G.; Khan, M.K.R.; Wang, X.X.; Cai, X.Y. Mapping QTLs for drought tolerance in an F 2: 3 population from an inter-specific cross between Gossypium tomentosum and Gossypium hirsutum. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A Robust, Simple Genotyping-by-Sequencing (GBS) Approach for High Diversity Species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef] [PubMed]
- Asins, M.J. Present and future of quantitative trait locus analysis in plant breeding. Plant Breed. 2002, 121, 281–291. [Google Scholar] [CrossRef]
- Abdelraheem, A.; Mahdy, E. The first linkage map for a recombinant inbred line population in cotton (Gossypium barbadense ) and its use in studies of PEG-induced dehydration tolerance. Euphytica 2015, 205, 941–958. [Google Scholar] [CrossRef]
- Adams, N.; Bajaj, S.; Zhang, J. Identification of cotton germplasm and molecular markers for drought tolerance. In Proceedings of the Beltwide Cotton Conferences, Atlanta, GA, USA, 7 January 2011; p. 708. [Google Scholar]
- Tiwari, R.S.; Picchioni, G.A.; Steiner, R.L.; Jones, D.C.; Hughs, S.E.; Zhang, J. Genetic variation in salt tolerance at the seedling stage in an interspecific backcross inbred line population of cultivated tetraploid cotton. Euphytica 2013, 194, 1–11. [Google Scholar] [CrossRef]
- Jia, Y.H.; Sun, J.L.; Wang, X.W.; Zhou, Z.L.; Pan, Z.E.; He, S.P.; Pang, B.Y.; Wang, L.R.; Du, X.M. Molecular Diversity and Association Analysis of Drought and Salt Tolerance in Gossypium hirsutum L. Germplasm. J. Integr. Agric. 2014, 13, 1845–1853. [Google Scholar] [CrossRef]
- Abdelraheem, A. Joint Genetic Linkage Mapping and Genome-wide Association Study of Drought and Salinity Tolerance and Verticillium Wilt and Thrips Resistance in Cotton. Ph.D. Thesis, New Mexico State University, Las Cruces, NM, USA, 2017. [Google Scholar]
- Abdelraheem, A.; Thyssen, G.; Jenkins, J.; McCarty, J.; Fang, D.D.; Zhang, J.F. A genome-wide association study of tolerance to biotic and abiotic stresses in a MAGIC population of upland cotton. In Proceedings of the 2018 Beltwild Cotton Conferences, San Antonio, TX, USA, 3–5 January 2018; p. 565. [Google Scholar]
- Hou, S.; Zhu, G.; Li, Y.; Li, W.; Fu, J.; Niu, E.; Li, L.; Zhang, D. Genome-Wide Association Studies Reveal Genetic Variation and Candidate Genes of Drought Stress Related Traits in Cotton (Gossypium hirsutum L.). Front. Plant Sci. 2018, 9, 1–15. [Google Scholar] [CrossRef]
- Said, J.I.; Lin, Z.; Zhang, X.; Song, M.; Zhang, J. A comprehensive meta QTL analysis for fiber quality, yield, yield related and morphological traits, drought tolerance, and disease resistance in tetraploid cotton. BMC Genom. 2013, 14, 776. [Google Scholar] [CrossRef]
- Abdelraheem, A.; Liu, F.; Song, M.; Zhang, J.F. A meta analysis of quantitative trait loci for abiotic and biotic stress resistance in tetraploid cotton. Mol. Genet. Genom. 2017, 292, 1221–1235. [Google Scholar] [CrossRef] [PubMed]
- Abdelraheem, A.; Fang, D.D.; Zhang, J. Quantitative trait locus mapping of drought and salt tolerance in an introgressed recombinant inbred line population of Upland cotton under the greenhouse and field conditions. Euphytica 2018, 214, 8. [Google Scholar] [CrossRef]
- Pauli, D.; Andrade-Sanchez, P.; Carmo-Silva, A.E.; Gazave, E.; French, A.N.; Heun, J.; Hunsaker, D.J.; Lipka, A.E.; Setter, T.L.; Strand, R.J.; et al. Field-Based High-Throughput Plant Phenotyping Reveals the Temporal Patterns of Quantitative Trait Loci Associated with Stress-Responsive Traits in Cotton. Genes Genomes Genet. 2016, 6, 865–879. [Google Scholar] [CrossRef] [PubMed]
- Abdelraheem, A.; Hughs, S.E.; Jones, D.C.; Zhang, J. Genetic analysis and quantitative trait locus mapping of PEG-induced osmotic stress tolerance in cotton. Plant Breed. 2015, 134, 111–120. [Google Scholar] [CrossRef]
- Amjid, M.W.; Malik, T.A.; Shakeel, A.; Wahid, A. QTL Mapping for Relative Leaf Water Contents, Cell Membrane Stability and Excised Leaf Water Loss under Drought by using EST-SSR Markers in Gossypium hirsutum. Int. J. Agric. Biol. 2015, 17, 4. [Google Scholar]
- Saeed, M.; Guo, W.; Ullah, I.; Tabbasam, N.; Zafar, Y. QTL mapping for physiology, yield and plant architecture traits in cotton (Gossypium hirsutum L.) grown under well-watered versus water-stress conditions. Electron. J. Biotechnol. 1983, 14, 3. [Google Scholar]
- Kushanov, F.N.; Pepper, A.E.; Yu, J.Z.; Buriev, Z.T.; Shermatov, S.E.; Saha, S.; Ulloa, M.; Jenkins, J.N.; Abdukarimov, A.; Abdurakhmonov, I.Y. Development, genetic mapping and QTL association of cotton PHYA, PHYB, and HY5-specific CAPS and dCAPS markers. BMC Genet. 2016, 17, 1–11. [Google Scholar] [CrossRef]
- Da Silva, C.R.C.; Jose, J.V.C.; de Albuquerque Melo Filho, P.; Jean, P.C.R.; Roseane, C.S.; Jessica, D.R.; Rennan, F.P. Characterization of water-stress tolerant cotton cultivars based on plant growth and in activity of antioxidant enzymes. Afr. J. Agric. Res. 2016, 11, 3763–3770. [Google Scholar]
- Kader, A.; Esmail, A.M.; El Shouny, K.A.; Ahmed, M.F. Screening cotton genotypes for drought tolerance by using non-enzymatic and enzymatic antioxidant systems. Egypt. J. Plant Breed 2015, 19, 169–184. [Google Scholar]
- Hamoud, H.; Soliman, Y.; Eldemery, S.; Abdellatif, K. Field Performance and Gene Expression of Drought Stress Tolerance in Cotton (Gossypium barbadense L.). Br. Biotechnol. J. 2016, 14, 1–9. [Google Scholar] [CrossRef]
- Mvula, J.; Bokosi, J.M.; Kabambe, V.; Banda, M.H.P. Screening cotton (Gossypium hirsutum L.) genotypes for drought tolerance under screen house conditions in Malawi. J. Plant Breed. Crop Sci. 2018, 10, 48–57. [Google Scholar]
- Paloti, M.; Patil, R.; Mummigatti, U. Breeding for Drought Tolerance in Cotton (Gossypium hirsutum L.) with an Emphasis on Physiological Parameters. Vegetos-Int. J. Plant Res. 2017, 1, 1–5. [Google Scholar] [CrossRef]
- Megha, B.R.; Mummigatti, U.V. Screening of Hirsutum Cotton Genotypes for Drought Tolerance under Different Osmotic Potential and Field Capacities. Int. J. Bio-Resour. Stress Manag. 2017, 8, 299–308. [Google Scholar] [CrossRef]
- Singh, C.; Kumar, V.; Prasad, I.; Patil, V.R.; Rajkumar, B.K. Response of upland cotton (G.Hirsutum L.) genotypes to drought stress using drought tolerance indices. J. Crop Sci. Biotechnol. 2016, 19, 53–59. [Google Scholar] [CrossRef]
- Sezener, V.; Basal, H.; Peynircioglu, C.; Gurbuz, T.; Kizilkaya, K. Screening of cotton cultivars for drought tolerance under field conditions. Turk. J. Field Crops 2015, 20, 223–232. [Google Scholar] [CrossRef]
- Memon, S.; Jatoi, W.A.; Chandio, G.M.; Section, C. Screening of Cotton Genotypes for Yield Traits. Pak. J. Agric. 2014, 30, 24–31. [Google Scholar]
- Riaz, M.; Farooq, J.; Sakhawat, G.; Mahmood, A.; Sadiq, M.A.; Yaseen, M. Genotypic variability for root/shoot parameters under water stress in some advanced lines of cotton (Gossypium hirsutum L.). Genet. Mol. Res. 2013, 12, 552–561. [Google Scholar] [CrossRef]
- Soomro, M.H.; Markhand, G.S.; Soomro, B.A. Screening Pakistani cotton for drought tolerance. Pak. J. Bot. 2012, 44, 383–388. [Google Scholar]
- Shah, A.R.; Khan, T.M.; Sadaqat, H.A.; Chatha, A.A. Alterations in leaf pigments in cotton (Gossypium hirsutum) genotypes subjected to drought stress conditions. Int. J. Agric. Biol. 2011, 13, 902–908. [Google Scholar]
- Iqbal, K.; Azhar, F.M.; Khan, I.A. Ehsan-Ullah Variability for drought tolerance in cotton (Gossypium hirsutum) and its genetic basis. Int. J. Agric. Biol. 2011, 13, 61–66. [Google Scholar]
- Longenberger, P.S.; Smith, C.W.; Thaxton, P.S.; Mcmichael, B.L. Development of a Screening Method for Drought Tolerance in Cotton Seedlings. Crop Sci. 2006, 46, 2104–2110. [Google Scholar] [CrossRef]
- Yu, L.-H.L.; Wu, S.-J.; Peng, Y.-S.; Liu, R.-N.; Chen, X.; Zhao, P.; Xu, P.; Zhu, J.-B.; Jiao, G.-L.; Pei, Y.; et al. Arabidopsis EDT1/HDG11 improves drought and salt tolerance in cotton and poplar and increases cotton yield in the field. Plant Biotechnol. J. 2015, 14, 72–84. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Sun, L.; Kuppu, S.; Hu, R.; Mishra, N.; Smith, J.; Esmaeili, N.; Herath, M.; Gore, M.A.; Payton, P.; et al. The yield difference between wild-type cotton and transgenic cotton that expresses IPT depends on when water-deficit stress is applied. Sci. Rep. 2018, 8, 2538. [Google Scholar] [CrossRef] [PubMed]
- Kerr, T.C.C.; Abdel-Mageed, H.; Aleman, L.; Lee, J.; Payton, P.; Cryer, D.; Allen, R.D. Ectopic expression of two AREB/ABF orthologs increases drought tolerance in cotton (Gossypium hirsutum). Plant Cell Environ. 2018, 41, 898–907. [Google Scholar] [CrossRef]
- Mishra, N.; Sun, L.; Zhu, X.; Smith, J.; Prakash Srivastava, A.; Yang, X.; Pehlivan, N.; Esmaeili, N.; Luo, H.; Shen, G.; et al. Overexpression of the Rice SUMO E3 Ligase Gene OsSIZ1 in Cotton Enhances Drought and Heat Tolerance, and Substantially Improves Fiber Yields in the Field under Reduced Irrigation and Rainfed Conditions. Plant Cell Physiol. 2017, 58, 735–746. [Google Scholar] [CrossRef]
- Honglan, Y.; Zhang, D.; Li, X.; Li, H.; Zhang, D.; Lan, H.; Wood, A.J.; Wang, J. Overexpression of ScALDH21 gene in cotton improves drought tolerance and growth in greenhouse and field conditions. Mol. Breed. 2016, 36, 34. [Google Scholar]
- Shen, G.; Wei, J.; Qiu, X.; Hu, R.; Kuppu, S.; Auld, D.; Blumwald, E.; Gaxiola, R.; Payton, P.; Zhang, H. Co-overexpression of AVP1 and AtNHX1 in Cotton Further Improves Drought and Salt Tolerance in Transgenic Cotton Plants. Plant Mol. Biol. Rep. 2015, 33, 167–177. [Google Scholar] [CrossRef]
- Liu, G.; Li, X.; Jin, S.; Liu, X.; Zhu, L.; Nie, Y.; Zhang, X. Overexpression of Rice NAC Gene SNAC1 Improves Drought and Salt Tolerance by Enhancing Root Development and Reducing Transpiration Rate in Transgenic Cotton. PLoS ONE 2014, 9, e86895. [Google Scholar] [CrossRef]
- Mittal, A.; Gampala, S.S.L.; Ritchie, G.L.; Payton, P.; Burke, J.J.; Rock, C.D. Related to ABA-Insensitive3(ABI3)/Viviparous1 and AtABI5 transcription factor coexpression in cotton enhances drought stress adaptation. Plant Biotechnol. J. 2014, 12, 578–589. [Google Scholar] [CrossRef]
- Yue, Y.; Zhang, M.; Zhang, J.; Tian, X.; Duan, L.; Li, Z. Overexpression of the AtLOS5 gene increased abscisic acid level and drought tolerance in transgenic cotton. J. Exp. Bot. 2012, 63, 3741–3748. [Google Scholar] [CrossRef]
- Pasapula, V.; Shen, G.; Kuppu, S.; Paez-Valencia, J.; Mendoza, M.; Hou, P.; Chen, J.; Qiu, X.; Zhu, L.; Zhang, X.; et al. Expression of an Arabidopsis vacuolar H+-pyrophosphatase gene (AVP1) in cotton improves drought- and salt tolerance and increases fibre yield in the field conditions. Plant Biotechnol. J. 2011, 9, 88–99. [Google Scholar] [CrossRef] [PubMed]
- Lv, S.-L.; Lian, L.-J.; Tao, P.-L.; Li, Z.-X.; Zhang, K.-W.; Zhang, J.-R. Overexpression of Thellungiella halophila H+-PPase (TsVP) in cotton enhances drought stress resistance of plants. Planta 2009, 229, 899–910. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; He, C.; Wang, J.; Mao, Z.; Holaday, S.A.; Allen, R.D.; Zhang, H. Overexpression of the Arabidopsis 14-3-3 Protein GF14λ in Cotton Leads to a “Stay-Green” Phenotype and Improves Stress Tolerance under Moderate Drought Conditions. Plant Cell Physiol. 2004, 45, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Belhaj, K.; Chaparro-Garcia, A.; Kamoun, S.; Patron, N.J.; Nekrasov, V. Editing plant genomes with CRISPR/Cas9. Curr. Opin. Biotechnol. 2015, 32, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Shan, Q.; Wang, Y.; Li, J.; Zhang, Y.; Chen, K.; Liang, Z.; Zhang, K.; Liu, J.; Xi, J.J.; Qiu, J.-L.; et al. Targeted genome modification of crop plants using a CRISPR-Cas system. Nat. Biotechnol. 2013, 31, 686–688. [Google Scholar] [CrossRef]
- Wang, Y.; Meng, Z.; Liang, C.; Meng, Z.; Wang, Y.; Sun, G.; Zhu, T.; Cai, Y.; Guo, S.; Zhang, R.; et al. Increased lateral root formation by CRISPR/Cas9-mediated editing of arginase genes in cotton. Sci. China Life Sci. 2017, 60, 524–527. [Google Scholar] [CrossRef]
- Shi, J.; Gao, H.; Wang, H.; Lafitte, H.R.; Archibald, R.L.; Yang, M.; Hakimi, S.M.; Mo, H.; Habben, J.E. ARGOS8 variants generated by CRISPR-Cas9 improve maize grain yield under field drought stress conditions. Plant Biotechnol. J. 2017, 15, 207–216. [Google Scholar] [CrossRef]
- Liu, J.H.; Peng, T.; Dai, W. Critical cis-Acting Elements and Interacting Transcription Factors: Key Players Associated with Abiotic Stress Responses in Plants. Plant Mol. Biol. Rep. 2014, 32, 303–317. [Google Scholar] [CrossRef]
- Zafar, S.A.; Zaidi, S.S.-A.; Gaba, Y.; Singla-Pareek, S.L.; Dhankher, O.P.; Xueyong Li1, S.M.; Pareek, A. Engineering Abiotic Stress Tolerance via CRISPR-Cas mediated genome editing. J. Exp. Bot. 2019. [Google Scholar] [CrossRef]
- Zhang, T.; Hu, Y.; Jiang, W.; Fang, L.; Guan, X.; Chen, J.; Zhang, J.; Saski, C.A.; Scheffler, B.E.; Stelly, D.M.; et al. Sequencing of allotetraploid cotton (Gossypium hirsutum L. acc. TM-1) provides a resource for fiber improvement. Nat. Biotechnol. 2015, 33, 531. [Google Scholar] [CrossRef]
- Wang, K.; Wang, Z.; Li, F.; Ye, W.; Wang, J.; Song, G.; Yue, Z.; Cong, L.; Shang, H.; Zhu, S.; et al. The draft genome of a diploid cotton Gossypium raimondii. Nat. Genet. 2012, 44, 1098–1103. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhao, B.; Zheng, H.-J.; Hu, Y.; Lu, G.; Yang, C.-Q.; Chen, J.-D.; Chen, J.-J.; Chen, D.-Y.; Zhang, L.; et al. Gossypium barbadense genome sequence provides insight into the evolution of extra-long staple fiber and specialized metabolites. Sci. Rep. 2015, 5, 14139. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Fan, G.; Wang, K.; Sun, F.; Yuan, Y.; Song, G.; Li, Q.; Ma, Z.; Lu, C.; Zou, C.; et al. Genome sequence of the cultivated cotton Gossypium arboreum. Nat. Genet. 2014, 46, 567–572. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Tu, L.; Lin, M.; Lin, Z.; Wang, P.; Yang, Q.; Ye, Z.; Shen, C.; Li, J.; Zhang, L.; et al. Asymmetric subgenome selection and cis-regulatory divergence during cotton domestication. Nat. Genet. 2017, 49, 579–587. [Google Scholar] [CrossRef]
- Du, X.; Huang, G.; He, S.; Yang, Z.; Sun, G.; Ma, X.; Li, N.; Zhang, X.; Sun, J.; Liu, M.; et al. Resequencing of 243 diploid cotton accessions based on an updated A genome identifies the genetic basis of key agronomic traits. Nat. Genet. 2018, 50, 796–802. [Google Scholar] [CrossRef]
- Ma, Z.; He, S.; Wang, X.; Sun, J.; Zhang, Y.; Zhang, G.; Wu, L.; Li, Z.; Liu, Z.; Sun, G.; et al. Resequencing a core collection of upland cotton identifies genomic variation and loci influencing fiber quality and yield. Nat. Genet. 2018, 50, 803–813. [Google Scholar] [CrossRef]
- Ashrafi, H.; Stelly, D.M.; Guan, X.; Chen, Z.J.; Wang, F.; Jones, D.C.; Van Deynze, A.; Matvienko, M.; Yang, S.S.; Mockaitis, K.; et al. A Long-Read Transcriptome Assembly of Cotton (L.) and Intraspecific Single Nucleotide Polymorphism Discovery. Plant Genome 2015, 8, 2. [Google Scholar] [CrossRef]
- Mills, R.E.; Walter, K.; Stewart, C.; Handsaker, R.E.; Chen, K.; Alkan, C.; Abyzov, A.; Yoon, S.C.; Ye, K.; Cheetham, R.K.; et al. Mapping copy number variation by population-scale genome sequencing. Nature 2011, 470, 59–65. [Google Scholar] [CrossRef]
- Zhou, B.; Zhang, L.; Ullah, A.; Jin, X.; Yang, X.; Zhang, X. Identification of multiple stress responsive genes by sequencing a normalized cDNA library from sea-land cotton (Gossypium barbadense L.). PLoS ONE 2016, 11, 1–22. [Google Scholar] [CrossRef]
- Zhang, B.; Chen, X.; Lu, X.; Shu, N.; Wang, X.; Yang, X.; Wang, S.; Wang, J.; Guo, L.; Wang, D.; et al. Transcriptome Analysis of Gossypium hirsutum L. Reveals Different Mechanisms among NaCl, NaOH and Na2CO3 Stress Tolerance. Sci. Rep. 2018, 8, 13527. [Google Scholar] [CrossRef]
- Jin, F.; Hu, L.; Yuan, D.; Xu, J.; Gao, W.; He, L.; Yang, X. Comparative transcriptome analysis between somatic embryos (SEs) and zygotic embryos in cotton: Evidence for stress response functions in SE development. Plant Biotechnol. J. 2014, 12, 161–173. [Google Scholar] [CrossRef]
- Osabe, K.; Clement, J.D.; Bedon, F.; Pettolino, F.A.; Ziolkowski, L.; Llewellyn, D.J.; Finnegan, E.J.; Wilson, I.W. Genetic and DNA Methylation Changes in Cotton (Gossypium) Genotypes and Tissues. PLoS ONE 2014, 9, e86049. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Pang, Y.; Jia, F.; Xiao, G.; Li, Q.; Zhu, Y. A Potential Role for CHH DNA Methylation in Cotton Fiber Growth Patterns. PLoS ONE 2013, 8, e60547. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.M.; Bird, A. DNA methylation landscapes: Provocative insights from epigenomics. Nat. Rev. Genet. 2008, 9, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Phillips, T. The role of methylation in gene expression. Nat. Educ. 2008, 1, 116. [Google Scholar]
- Song, Q.; Guan, X.; Chen, Z.J. Dynamic Roles for Small RNAs and DNA Methylation during Ovule and Fiber Development in Allotetraploid Cotton. PLOS Genet. 2015, 11, e1005724. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; Zhang, T.; Stelly, D.M.; Chen, Z.J. Epigenomic and functional analyses reveal roles of epialleles in the loss of photoperiod sensitivity during domestication of allotetraploid cottons. Genome Biol. 2017, 18, 99. [Google Scholar] [CrossRef]
- Dixon, J.E.; Dowen, R.H.; Nery, J.R.; Pelizzola, M.; Ecker, J.R.; Lister, R.; Schmitz, R.J.; Dowen, J.M. Widespread dynamic DNA methylation in response to biotic stress. Proc. Natl. Acad. Sci. USA 2012, 109, E2183–E2191. [Google Scholar]
- Xie, F.; Zhang, B. microRNA evolution and expression analysis in polyploidized cotton genome. Plant Biotechnol. J. 2015, 13, 421–434. [Google Scholar] [CrossRef]
- Guo, J.; Shi, G.; Guo, X.; Zhang, L.; Xu, W.; Wang, Y.; Su, Z.; Hua, J. Transcriptome analysis reveals that distinct metabolic pathways operate in salt-tolerant and salt-sensitive upland cotton varieties subjected to salinity stress. Plant Sci. 2015, 238, 33–45. [Google Scholar] [CrossRef]
- Paterson, A.H.; Wendel, J.F. Unraveling the fabric of polyploidy. Nat. Biotechnol. 2015, 33, 491–493. [Google Scholar] [CrossRef] [PubMed]
- Paterson, A.H.; Wendel, J.F.; Gundlach, H.; Guo, H.; Jenkins, J.; Jin, D.; Llewellyn, D.; Showmaker, K.C.; Shu, S.; Udall, J.; et al. Repeated polyploidization of Gossypium genomes and the evolution of spinnable cotton fibres. Nature 2012, 492, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Giovannoni, J.J.; Katzir, N.; Jiao, C.; Wu, S.; Burger, J.; Ando, K.; Wechter, W.P.; Blanca, J.; Zheng, Y.; et al. Cucurbit Genomics Database (CuGenDB): A central portal for comparative and functional genomics of cucurbit crops. Nucleic Acids Res. 2018, 47, D1128–D1136. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-M.; Kim, S.; Koo, N.; Shin, A.-Y.; Yeom, S.-I.; Seo, E.; Park, S.-J.; Kang, W.-H.; Kim, M.-S.; Park, J.; et al. Genome analysis of Hibiscus syriacus provides insights of polyploidization and indeterminate flowering in woody plants. DNA Res. 2017, 24, 71–80. [Google Scholar] [PubMed]
- Argout, X.; Salse, J.; Aury, J.M.; Guiltinan, M.J.; Droc, G.; Gouzy, J.; Allegre, M.; Chaparro, C.; Legavre, T.; Maximova, S.N.; et al. The genome of Theobroma cacao. Nat. Genet. 2011, 43, 101–108. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Type | Phenotypic Effect/Function | Reference |
|---|---|---|---|
| GhHUB2 | Histone H2B monoubiquitinatin E3 ligase encoding gene | Drought tolerance through increased soluble sugar, proline, and leaf relative water contents | [43] |
| GrMAPKKK and GhMAPKKK | MAPK gene family | Drought and salt responsive | [20] |
| GhMAP3K1, GhMKK4, and GhMPK6 | MAPK signaling gene | Regulates the drought stress response by interacting with GhWRKY59–GhDREB2 | [26] |
| GhMKK3 | MAPK signaling gene | Enhanced drought tolerance | [25] |
| GhMAP3K40 | MAPK signaling gene | Salt and drought stress tolerance at the germination stage | [44] |
| GhMPK4 | MAPK signaling gene | Increased sensitivity to ABA, salt, and drought | [45] |
| GhMPK17 | MAPK signaling gene | Osmotic and salt stress tolerance | [46] |
| GbMPK3 | MAPK signaling gene | Enhanced oxidative and drought stress tolerance | [47] |
| GhMPK6a | MAPK signaling gene | Drought and salinity | [48] |
| GhMKK1 | MAPK signaling gene | Drought and salinity | [49] |
| GhMKK5 | MAPK signaling gene | Drought and salinity | [50] |
| GhMPK2 | MAPK signaling gene | Drought and salinity | [51] |
| GbRLK | Receptor-like kinase | Drought and salinity | [52] |
| GaHDG11 (HD-ZIP) | Transcription factor | Drought and heat stress | [53] |
| GhNAC79 | Transcription factor | Improves resistance to drought stress | [42] |
| GhERF38 | Transcription factor | Drought, abscisic acid, and salinity | [54] |
| GhERF2, GhERF3, GhERF6 | Transcription factor | Drought, salt, ethylene, and abscisic acid | [55] |
| GhWRKY59 | Transcription factor | Activates MAPK signaling gene under drought | [26] |
| GhWRKY25 | Transcription factor | Drought and salinity | [56] |
| GhABF2 (bZIP) | Transcription factor | Enhances the activities of CAT and SOD, regulates gene expression related to ABA | [39] |
| GhNAC2 | Transcription factor | Longer roots, and enhanced salt and drought tolerance | [57] |
| GhCBF3, GhAREB1, and GhAREB2 | ABA-induced gene | Small stomatal aperture, enhanced drought- and high salinity-tolerance via the ABA signaling pathway | [58] |
| GhNAC7-GhNAC13 | Transcription factor | Cold, abscisic acid, drought, and salinity | [59] |
| GbMYB5 | Transcription factor | Reduced water loss trough stomatal conductance, and increased proline content and antioxidant enzymes | [27] |
| GhWRKY41 | Transcription factor | Lower malondialdehyde content, higher antioxidant activity, and induced stomatal conductance | [41] |
| GhWRKY17 | Transcription factor | Increases sensitivity to ABA and drought stress | [60] |
| GhNAC8-GhNAC17 | Transcription factor | Drought, salinity, cold, and ABA | [61] |
| GhNAC1-GhNAC6 | Transcription factor | Drought, cold, salinity, and ABA | [62] |
| GhDREB | Transcription factor | Drought, cold, and salinity | [63] |
| GhDREB1 | Transcription factor | Drought, cold, and salinity | [64] |
| GhDBP2 | Transcription factor | Drought, cold, and ABA | [65] |
| GhERF1 | Transcription factor | ABA production and drought stress signaling regulation | [66] |
| GhERF4 | Transcription factor | ABA production and drought stress signaling regulation | [67] |
| GhDREB1L | Transcription factor | Drought, cold, and salinity | [68] |
| GhPYL9–11A | ABA receptor gene | ABA receptor that mediates the response to drought stress | [69] |
| GhSnRK2 | Involved in ABA signaling | Drought, salinity, cold, and ABA | [70] |
| GhCDPK35, GhCDPK28, GhCDPK16, GhCDPK14, GhCDPK11 and GhCDPK3 | Ca2+-activated gene | Drought and salinity stress responsive | [7] |
| GhCIPK6 | Ca2+-activated gene | Increased drought, salinity, and ABA stress tolerance | [30] |
| GhD12G207 | CDK gene family | Increased concentration of antioxidant enzymes (POD, SOD, and CAT), cell membrane stability, and chlorophyll content under drought and salt stress | [71] |
| GaMYB62L | Transcription factor | Increased chlorophyll and proline contents, higher germination rate under drought salt stress | [72] |
| GhTPS11 | Functional gene | Drought, heat, salinity, ABA, and gibberellin acid | [73] |
| GhAVP1 | Functional gene | Drought and salinity tolerance | [74] |
| QTL | Traits | Size and Type of Population | Number and Types of Markers Used | Reference |
|---|---|---|---|---|
| 49 | Lint yield, seed cotton yield, fiber length, fiber elongation, boll weight, leaf area, fresh shoot weight, and plant height | 97 F5:9 RILs (TM-1 × NM24016) | RGA-AFLP, SSR and GBS-SNP (1004) | [125] |
| 59 | Canopy temperature, normalized difference vegetation index, canopy height, and leaf area index | 95 RIL (TM-1 × NM24016) | SSR (429) | [126] |
| 67 | Plant height, chlorophyll content, leaf number, leaf area, leaf dry and fresh weights, number of fruiting branches, number of bolls, and boll weight | 188 F2:3 (CRI-12 × AD3-00) | SSR (1295) | [113] |
| 6 | Plant height, and fresh shoot and root weight. | 142 BILs (Pima S-7 × Sure-Grow747) | AFLP, RGA and RGA-AFLP (34) | [127] |
| 14 | Chlorophyll content, leaf temperature, fresh shoot and root weight, evapotranspiration, and plant height | 140 RILs (Dan-dara × Giza-70) | SSCP (165) | [116] |
| 3 | Excised leaf water loss and relative water content | 100 F2 (B-557 × FH1000) | SSR and EST-SSR (524) | [109] |
| 6 | Relative water content, excised leaf water loss, cell membrane stability, stomatal frequency, and stomatal size | 100 F2 (FH-207 × FH901) | EST-SSR (2365) | [128] |
| 7 | Chlorophyll content, osmotic potential, carbon isotope ratio, and seed cotton yield | 28 NILs (GH ‘Sivon’ × GB cv.F-177) | RFLP (279) | [81] |
| 79 | Chlorophyll a and b, carbon isotope ratio, osmotic potential, canopy temperature, dry matter, harvest index, boll weight and boll number, and seed cotton yield | 208 F3 (GH ‘Sivon’ × GB cv.F-177) | RFLP (253) | [110] |
| 3 | Osmotic potential, osmotic adjustment, and plant height | 136 F2 and F2:4 (FH-901 × RH-510) | SSR (6500) | [129] |
| Genotypes | Origin | Traits/Method | Reference |
|---|---|---|---|
| 06K485, 06K486, SPAN 837, FQMA (05)5bcp, Chureza, and RASAM 17 | DARS, Malawi | Fresh and dry root weight, lateral roots number, tap root length, root volume, fresh and dry shoot weight, stem diameter, shoot length, and number of leaves per plant | [134] |
| GhAM-46, GhAM-9, EC560413, and GhAM-78 | India | SPAD chlorophyll contents, excised leaf water loss, root volume, root and shoot length, root and shoot weight, and final yielding | [135] |
| LRA-5166, BS-37, CCH-12-3, BS39, GBHV-177, GBHV-182, and ARBH-1352 | India | Root and shoot length, percent seed germination, and seedling vigor (shoot vigor index, seedling vigor index, and root vigor index) | [136] |
| BRS 286, CNPA 7MH, and CNPA 5M | Brazil | Antioxidant enzymes activities (APX, CAT, and SOD) | [131] |
| H1353/10 × G.Cot.16 and G.Cot.16 × H-1353/10 | India | Yield index, yield stability index, yield reduction ratio, mean productivity, geometric mean productivity, stress susceptibility index, tolerance index, and stress tolerance index | [137] |
| Giza75 | Egypt | Drought stress index (DSI) and expression of drought-related genes (Gossypium heat shock protein 1 [GhSP1] and flowering locus T-like protein 1 [FTL1]) | [133] |
| Suvin | India | ||
| 10229 | Australia | ||
| Giza80, Giza90, Giza80 × Tamco C.E., Giza90 × (Giza9 × Giza Australian) and Giza90 × TamcoC | Egypt | Enzymatic (ascorbate peroxidase, catalase, peroxidase, and superoxide dismutase) and non-enzymatic (phenolic content, lipid peroxidation, and proline) antioxidant activities | [132] |
| Acala-1517-99, DAK-66/3, and GC-555 | USA | Seed germination, seedling growth, yield, yield components, and genotypes characterized with low drought susceptibility index, and high geometric mean productivity | [138] |
| Nieves | Australia | ||
| MS-30/1 and Nazilli M-503 | Turkey | ||
| Eva and Zeta 2, | Greece | ||
| NIAB-999 | Pakistan | ||
| Delta Diamond | Spain | ||
| Sindh-1 and Shahbaz-95 | Pakistan | Lint yield per plant, boll weight, bolls per plant, sympodial branches per plant, and plant height | [139] |
| FH-942 and FH-113, | Pakistan | Excised leaf water loss, shoot and root lengths, number of lateral roots, fresh root and shoot weights (g), dry shoot and root weight (g), and total plant fresh weight (g) | [140] |
| MARVI, CRIS-9, CRIS-, CRIS-337, CRIS-126, CRIS-355, and 377CRIS-134 | Pakistan | Chlorophyll content, RWC, transpiration rate, excised leaf water loss, yield components, and yield | [141] |
| FH-113, MNH-789, and PB-899 | Pakistan | Chlorophyll, carotenoids, and polyphenols | [142] |
| 149F, BOU 1724-3, B-557, and DPL-26 | Pakistan | Drought tolerance indices, relative shoot and root length | [143] |
| Acala-1517–99 and CRS-M-9044–0165 | USA | Seedling traits | [144] |
| Gene | Effects on Cotton Drought Tolerance | Effect on Yield | Stress Type | Donor Specie | References |
|---|---|---|---|---|---|
| AtABF3 | Improved stomatal regulation, less transpiration, and photosynthetic productivity | Yield increased | Drought | Arabidopsis thaliana | [147] |
| IPT | More number of bolls and larger root systems | Yield increased | Drought and heat | Agrobacterium tumefaciens | [146] |
| OsSIZ1 | Higher net photosynthesis, better growth, and cotton fiber yield | Yield increased | Drought and heat | Rice | [148] |
| ScALDH21 | Soluble sugar and proline content increased, higher peroxidase activity, reduced loss of net photosynthesis, reduced lipid peroxidation, greater plant height, and larger bolls | Yield increased | Drought | Syntrichia caninervis | [149] |
| AtEDT1/HDG11 | Soluble sugar and proline content increased, well-developed roots, low stomatal density, increased ROS scavenging enzymes | 43% more seeds | Drought and salt | Arabidopsis thaliana | [145] |
| AtNHX1 and AVP1 | Plant height, boll number, and fiber yield | 24–35% more fiber | Drought and Salt | Arabidopsis thaliana | [150] |
| SNAC1 | Enhanced proline content and root development, decreased transpiration rate | 31% more bolls | Drought and salt | Rice | [151] |
| SNAC1 | Reduced transpiration rate and more vigorous root system | Salt and drought | Rice | [151] | |
| AtABI5 & AtRAV1/2 | ROS scavenging, osmotic adjustment, improved photo-assimilation, root and shoot sink strengths, enhanced expression of GhRAV and genes for antioxidant and osmolyte biosynthesis | Yield affected | Drought | Arabidopsis thaliana | [152] |
| AtLOS5 | Enhanced ABA levels to improve drought tolerance with 13% more fresh biomass | 13% more fresh biomass | Drought and heat | Arabidopsis thaliana | [153] |
| AVP1 | Enhanced sequestration of ions and sugars into the vacuole, reduced water potential, and enhanced root biomass | Increased 20% | Drought and salt | Arabidopsis thaliana | [154] |
| TsVP | Improved root and shoot growth, higher rate of photosynthesis and relative water content, less cell membrane damaged | 27–53% higher in Lumianyan2142–61% in Lumianyan19 | Drought | Thellungiella halophila | [155] |
| betA | Increased photosynthesis, higher relative water content, better osmotic adjustment, less ion leakage, and lipid membrane peroxidation | 3–12% higher | Drought | Escherichia coli | [85] |
| GF14λ | Higher photosynthesis rate, enhanced senescence, and chlorophyll content | Enhanced | Drought tolerance | Arabidopsis thaliana | [156] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahmood, T.; Khalid, S.; Abdullah, M.; Ahmed, Z.; Shah, M.K.N.; Ghafoor, A.; Du, X. Insights into Drought Stress Signaling in Plants and the Molecular Genetic Basis of Cotton Drought Tolerance. Cells 2020, 9, 105. https://doi.org/10.3390/cells9010105
Mahmood T, Khalid S, Abdullah M, Ahmed Z, Shah MKN, Ghafoor A, Du X. Insights into Drought Stress Signaling in Plants and the Molecular Genetic Basis of Cotton Drought Tolerance. Cells. 2020; 9(1):105. https://doi.org/10.3390/cells9010105
Chicago/Turabian StyleMahmood, Tahir, Shiguftah Khalid, Muhammad Abdullah, Zubair Ahmed, Muhammad Kausar Nawaz Shah, Abdul Ghafoor, and Xiongming Du. 2020. "Insights into Drought Stress Signaling in Plants and the Molecular Genetic Basis of Cotton Drought Tolerance" Cells 9, no. 1: 105. https://doi.org/10.3390/cells9010105
APA StyleMahmood, T., Khalid, S., Abdullah, M., Ahmed, Z., Shah, M. K. N., Ghafoor, A., & Du, X. (2020). Insights into Drought Stress Signaling in Plants and the Molecular Genetic Basis of Cotton Drought Tolerance. Cells, 9(1), 105. https://doi.org/10.3390/cells9010105