Mouse Models for Food Allergies: Where Do We Stand?


Abstract
1. Introduction
1.1. Relevance of Food Allergies
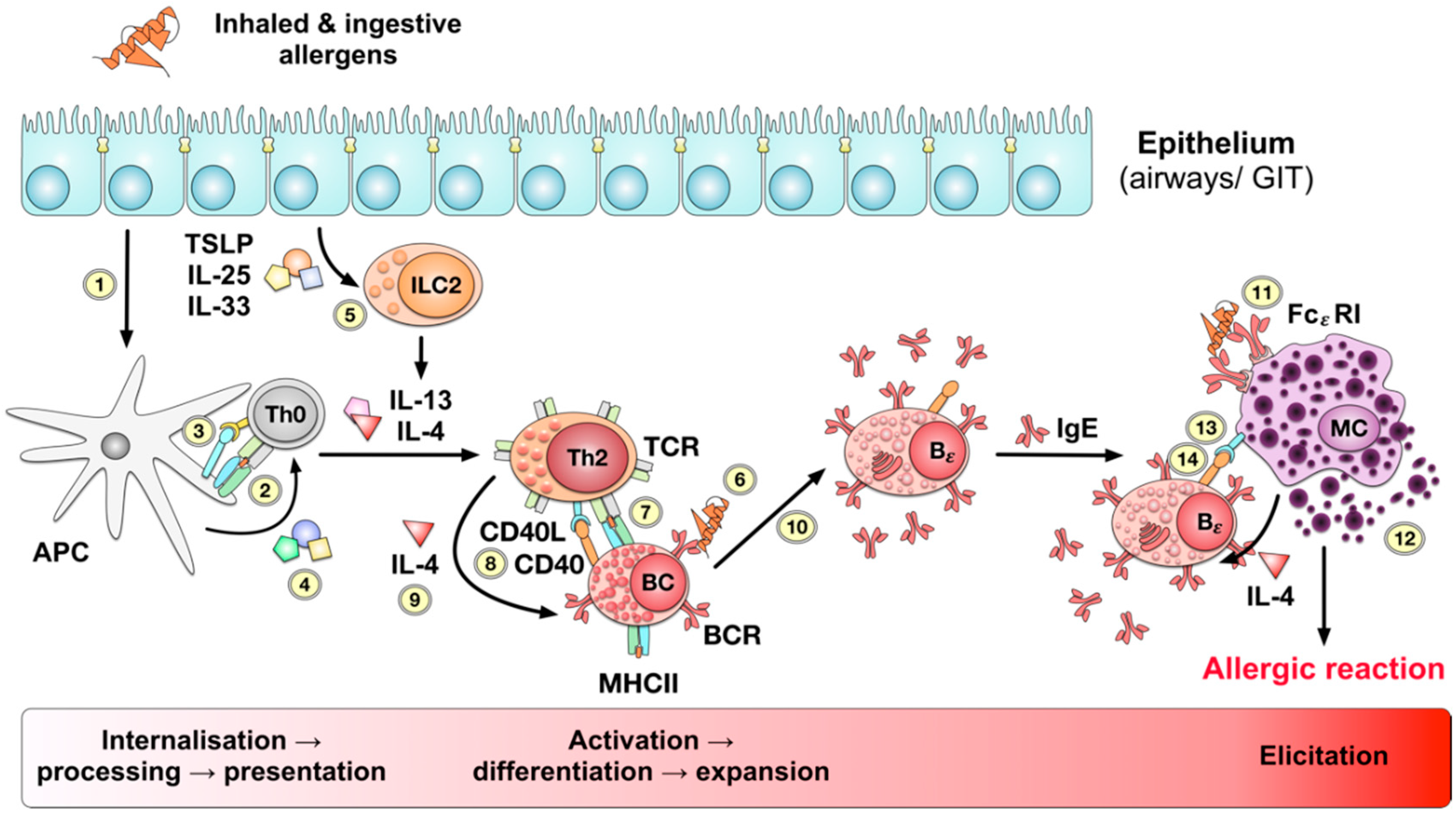
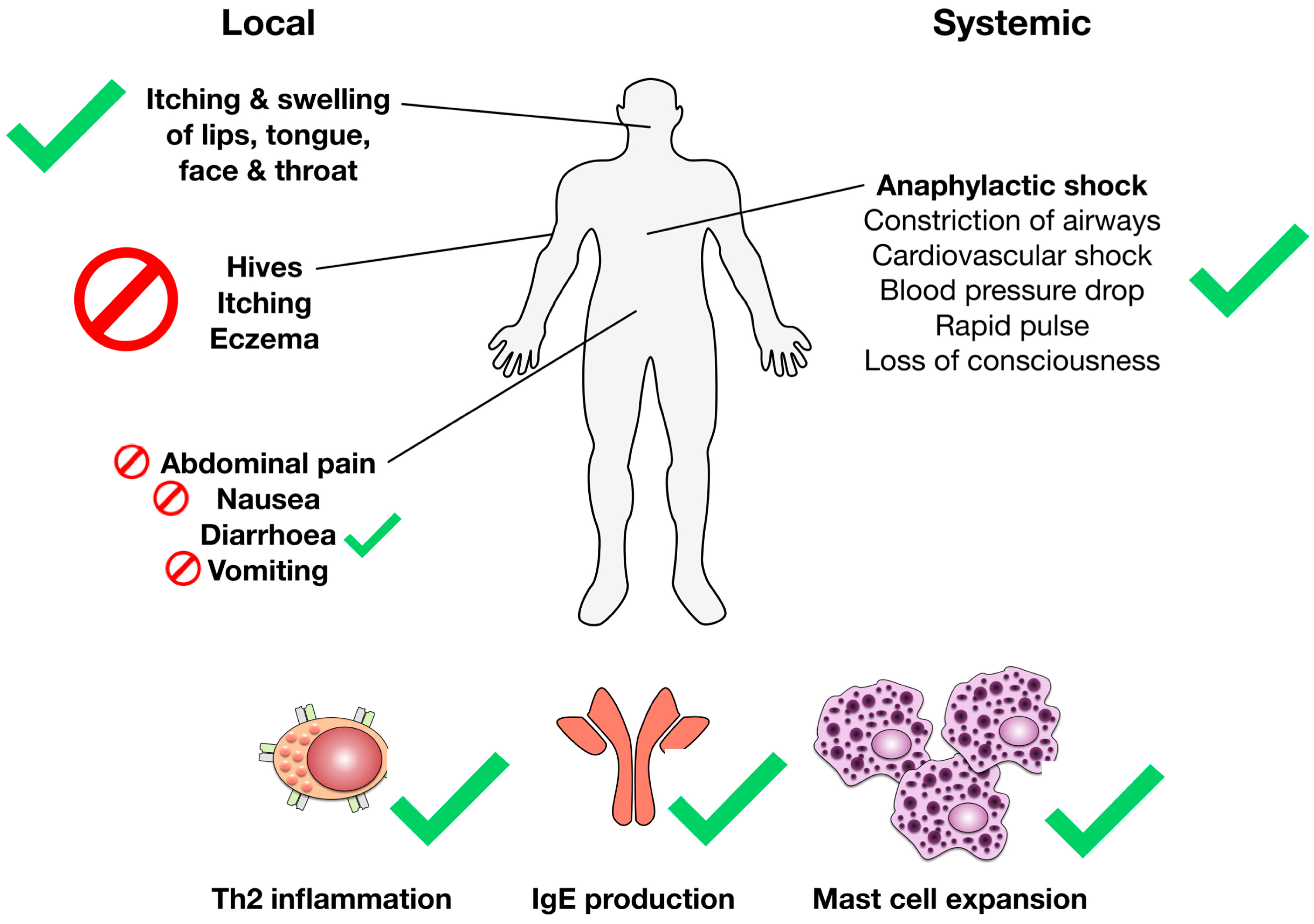
1.2. Pathomechanism of Food Allergy
2. Factors Contributing to the Development of Food Allergy
3. Advantages of Animal Models for Food Allergy Research
- (1)
- to investigate the immune responses underlying the allergic pathology,
- (2)
- to establish and compare the allergenic potency of candidate molecules, and
- (3)
- to evaluate the potency and safety of novel therapeutic options and vaccines derived from these findings in vivo without endangering the lives and health of the patients.
4. General Characteristics of the Models
5. Usage of Adjuvants in Food Allergy Mouse Models
6. Types of Mouse Food Allergy Models
6.1. Adjuvant-Free Models
6.2. Adjuvanted Mouse Food Allergy Models
6.3. Mouse Models Using Genetically-Modified Mouse Strains
6.4. Humanized Mouse Models
7. Summary and Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AAV | Adeno-associated virus |
| AIT | Allergen-specific immunotherapy |
| Alum | Aluminum hydroxide |
| APC | Antigen presenting cell |
| cAMP | Cyclic adenosine monophosphate |
| CT | Cholera toxin |
| CNS1 | Non-coding DNA sequence 1 |
| DC | Dendritic cell |
| DAMP | Damage associated molecular pattern |
| GATA3 | GATA binding protein 3 |
| GM-CSF | Granulocyte-macrophage colony-stimulating factor |
| HSC | Hematopoietic stem cells |
| IL(-25) | Interleukin (25) |
| ILC2s | Innate like lymphocytes type II cells |
| iTregs | Induced Tregs |
| LPS | Lipopolysaccharide |
| MAPK | Mitogen-activated protein kinase |
| mMCP1 | Mouse mast cell protease 1 |
| NFκB | Nuclear factor “kapa-light-enhancer“ of activated B cells |
| NOD | Non-obese diabetic |
| Ova | Ovalbumin |
| PBMC | Peripheral blood mononuclear cells |
| SCID | Severe combined immunodeficiency |
| SCF | Stem cell factor |
| SEB | Staphylococcus enterotoxin B |
| TCR | T cell receptor |
| Th1/2/17 | T helper 1/2/17 cell |
| TLR(4) | “Toll“-like receptor (4) |
| (i)Treg | (Peripherally induced) regulatory T cell |
| TSLP | Thymic stromal lymphopoietin |
| WAS(P) | Wiskott-Aldrich syndrome (protein) |
References
- Sicherer, S.H. Food allergy. Mt. Sinai J. Med. 2011, 78, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.S.; Springston, E.E.; Warrier, M.R.; Smith, B.; Kumar, R.; Pongracic, J.; Holl, J.L. The prevalence, severity, and distribution of childhood food allergy in the United States. Pediatrics 2011, 128, e9–e17. [Google Scholar] [CrossRef] [PubMed]
- Sicherer, S.H.; Sampson, H.A. Food allergy. J. Allergy Clin. Immunol. 2010, 125, S116–S125. [Google Scholar] [CrossRef] [PubMed]
- Decker, W.W.; Campbell, R.L.; Manivannan, V.; Luke, A.; St Sauver, J.L.; Weaver, A.; Bellolio, M.F.; Bergstralh, E.J.; Stead, L.G.; Li, J.T.C. The etiology and incidence of anaphylaxis in Rochester, Minnesota: A report from the Rochester Epidemiology Project. J. Allergy Clin. Immunol. 2008, 122, 1161–1165. [Google Scholar] [CrossRef]
- Patel, D.A.; Holdford, D.A.; Edwards, E.; Carroll, N.V. Estimating the economic burden of food-induced allergic reactions and anaphylaxis in the United States. J. Allergy Clin. Immunol. 2011, 128, 110–115. [Google Scholar] [CrossRef]
- Hefle, S.L.; Nordlee, J.A.; Taylor, S.L. Allergenic foods. Crit. Rev. Food Sci. Nutr. 1996, 36, S69–S89. [Google Scholar] [CrossRef]
- Lehrer, S.B.; Ayuso, R.; Reese, G. Current understanding of food allergens. Ann. N. Y. Acad. Sci. 2002, 964, 69–85. [Google Scholar] [CrossRef]
- Koya, T.; Matsuda, H.; Takeda, K.; Matsubara, S.; Miyahara, N.; Balhorn, A.; Dakhama, A.; Gelfand, E.W. IL-10–treated dendritic cells decrease airway hyperresponsiveness and airway inflammation in mice. J. Allergy Clin. Immunol. 2007, 119, 1241–1250. [Google Scholar] [CrossRef]
- Gentile, D.A.; Schreiber, R.; Howe-Adams, J.; Trecki, J.; Patel, A.; Angelini, B.; Skoner, D.P. Diminished dendritic cell interleukin 10 production in atopic children. Ann. Allergy Asthma Immunol. 2004, 92, 538–544. [Google Scholar] [CrossRef]
- Barlow, J.L.; Peel, S.; Fox, J.; Panova, V.; Hardman, C.S.; Camelo, A.; Bucks, C.; Wu, X.; Kane, C.M.; Neill, D.R.; et al. IL-33 is more potent than IL-25 in provoking IL-13–producing nuocytes (type 2 innate lymphoid cells) and airway contraction. J. Allergy Clin. Immunol. 2013, 132, 933–941. [Google Scholar] [CrossRef]
- Wang, C.; Liu, Q.; Chen, F.; Xu, W.; Zhang, C.; Xiao, W. IL-25 Promotes Th2 Immunity Responses in Asthmatic Mice via Nuocytes Activation. PLoS ONE 2016, 11, 0162393. [Google Scholar] [CrossRef] [PubMed]
- Barlow, J.L.; McKenzie, A.N. Type-2 innate lymphoid cells in human allergic disease. Curr. Opin. Allergy Clin. Immunol. 2014, 14, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Kanagaratham, C.; Sallis, B.F.; Fiebiger, E. Experimental Models for Studying Food Allergy. Cell. Mol. Gastroenterol. Hepatol. 2018, 6, 356–369.e1. [Google Scholar] [CrossRef] [PubMed]
- Sutton, B.J.; Davies, A.M. Structure and dynamics of IgE-receptor interactions: FcεRI and CD23/FcεRII. Immunol. Rev. 2015, 268, 222–235. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Sampson, H.A. Food allergy. J. Clin. Investig. 2011, 121, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Schouten, B.; Van Esch, B.C.; Hofman, G.A.; Elsen, L.W.V.D.; Willemsen, L.E.; Garssen, J. Acute Allergic Skin Reactions and Intestinal Contractility Changes in Mice Orally Sensitized against Casein or Whey. Int. Arch. Allergy Immunol. 2008, 147, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Andreassen, M.; Rudi, K.; Angell, I.L.; Dirven, H.; Nygaard, U.C. Allergen Immunization Induces Major Changes in Microbiota Composition and Short-Chain Fatty Acid Production in Different Gut Segments in a Mouse Model of Lupine Food Allergy. Int. Arch. Allergy Immunol. 2018, 177, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Bøgh, K.L.; van Bilsen, J.; Głogowski, R.; López-Expósito, I.; Bouchaud, G.; Blanchard, C.; Bodinier, M.; Smit, J.; Pieters, R.; Bastiaan-Net, S.; et al. Current challenges facing the assessment of the allergenic capacity of food allergens in animal models. Clin. Transl. Allergy 2016, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Lauener, R.P.; Frei, R.; Crameri, R.; O’Mahony, L. Microbiota and dietary interactions–An update to the hygiene hypothesis? Allergy 2012, 67, 451–461. [Google Scholar]
- Konieczna, P.; Groeger, D.; Ziegler, M.; Frei, R.; Ferstl, R.; Shanahan, F.; Quigley, E.M.M.; Kiely, B.; Akdis, C.A.; O’Mahony, L. Bifidobacterium infantis 35624 administration induces Foxp3 T regulatory cells in human peripheral blood: potential role for myeloid and plasmacytoid dendritic cells. Gut 2012, 61, 354–366. [Google Scholar] [CrossRef]
- Ruiter, B.; Shreffler, W.G. Innate immunostimulatory properties of allergens and their relevance to food allergy. Semin. Immunopathol. 2012, 34, 617–632. [Google Scholar] [CrossRef] [PubMed]
- Reithofer, M.; Jahn-Schmid, B. Allergens with Protease Activity from House Dust Mites. Int. J. Mol. Sci. 2017, 18, 1368. [Google Scholar] [CrossRef] [PubMed]
- Nowak-Węgrzyn, A.; Fiocchi, A. Rare, medium, or well done? The effect of heating and food matrix on food protein allergenicity. Curr. Opin. Allergy Clin. Immunol. 2009, 9, 234–237. [Google Scholar] [PubMed]
- Kim, J.S.; Nowak-Wegrzyn, A.; Sicherer, S.H.; Noone, S.; Moshier, E.L.; Sampson, H.A. Dietary baked milk accelerates the resolution of cow’s milk allergy in children. J. Allergy Clin. Immunol. 2011, 128, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Feehley, T.; Plunkett, C.H.; Bao, R.; Choi Hong, S.M.; Culleen, E.; Belda-Ferre, P.; Campbell, E.; Aitoro, R.; Nocerino, R.; Paparo, L.; et al. Healthy infants harbor intestinal bacteria that protect against food allergy. Nat. Med. 2019, 25, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Mastrorilli, C.; Caffarelli, C.; Hoffmann-Sommergruber, K.; Hoffmann-Sommergruber, K. Food allergy and atopic dermatitis: prediction, progression and prevention. Pediatr. Allergy Immunol. 2017, 28, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Renz, H.; Allen, K.J.; Sicherer, S.H.; Sampson, H.A.; Lack, G.; Beyer, K.; Oettgen, H.C. Food allergy. Nat. Rev. Dis. Primers 2018, 4, 17098. [Google Scholar] [CrossRef] [PubMed]
- Bush, A.; Saglani, S. Management of severe asthma in children. Lancet 2010, 376, 814–825. [Google Scholar] [CrossRef]
- Johansen, P.; Senti, G.; Martinez Gomez, J.M.; Storni, T.; Beust von, B.R.; Wüthrich, B.; Bot, A.; Kündig, T.M. Toll-like receptor ligands as adjuvants in allergen-specific immunotherapy. Clin. Exp. Allergy 2005, 35, 1591–1598. [Google Scholar] [CrossRef]
- Oppenheimer, J.; Nelson, H.; Bock, S.; Christensen, F.; Leung, D. Treatment of peanut allergy with rush immunotherapy. J. Allergy Clin. Immunol. 1992, 90, 256–262. [Google Scholar] [CrossRef]
- Feuille, E.; Nowak-Węgrzyn, A. Oral Immunotherapy for Food Allergies. Ann. Nutr. Metab. 2016, 68, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Nurmatov, U.; Dhami, S.; Arasi, S.; Pajno, G.B.; Fernandez-Rivas, M.; Muraro, A.; Roberts, G.; Akdis, C.; Alvaro-Lozano, M.; Beyer, K.; et al. Allergen immunotherapy for IgE-mediated food allergy: A systematic review and meta-analysis. Allergy 2017, 72, 1133–1147. [Google Scholar] [CrossRef] [PubMed]
- Knippels, L.M.; Vanwijk, F.; Penninks, A.H. Food allergy: What do we learn from animal models? Curr. Opin. Allergy Clin. Immunol. 2004, 4, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Finkelman, F.D. Anaphylaxis: Lessons from mouse models. J. Allergy Clin. Immunol. 2007, 120, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Beutier, H.; Gillis, C.M.; Iannascoli, B.; Godon, O.; England, P.; Sibilano, R.; Reber, L.L.; Galli, S.J.; Cragg, M.S.; Van Rooijen, N.; et al. IgG subclasses determine pathways of anaphylaxis in mice. J. Allergy Clin. Immunol. 2017, 139, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Mestas, J.; Hughes, C.C.W. Of Mice and Not Men: Differences between Mouse and Human Immunology. J. Immunol. 2004, 172, 2731–2738. [Google Scholar] [CrossRef]
- Graham, M.T.; Nadeau, K.C. Lessons learned from mice and man: Mimicking human allergy through mouse models. Clin. Immunol. 2014, 155, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, S.C. Role of mast cells in allergic and non-allergic immune responses: Comparison of human and murine data. Nat. Rev. Immunol. 2007, 7, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, R.G.; Oppenheimer, J. Serological IgE Analyses in the Diagnostic Algorithm for Allergic Disease. J. Allergy Clin. Immunol. Pr. 2015, 3, 833–840. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-M.; Serebrisky, D.; Lee, S.-Y.; Huang, C.-K.; Bardina, L.; Schofield, B.H.; Stanley, J.; Burks, A.; Bannon, G.A.; Sampson, H.A. A murine model of peanut anaphylaxis: T- and B-cell responses to a major peanut allergen mimic human response. J. Allergy Clin. Immunol. 2000, 106, 150–158. [Google Scholar] [CrossRef]
- Brandt, E.B.; Strait, R.T.; Hershko, D.; Wang, Q.; Muntel, E.E.; Scribner, T.A.; Zimmermann, N.; Finkelman, F.D.; Rothenberg, M.E. Mast cells are required for experimental oral allergen–induced diarrhea. J. Clin. Investig. 2003, 112, 1666–1677. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, M.; Ahrens, R.; Osterfeld, H.; Kottyan, L.C.; Shang, X.; A MacLennan, J.; Zimmermann, N.; Zheng, Y.; Finkelman, F.D.; Hogan, S.P. Increased susceptibility of 129SvEvBrd mice to IgE-Mast cell mediated anaphylaxis. BMC Immunol. 2011, 12, 14. [Google Scholar] [CrossRef] [PubMed]
- Wavrin, S.; Bernard, H.; Meziti, N.; Wal, J.-M.; Adel-Patient, K.; Ah-Leung, S.; Wal, J. Oral tolerance and Treg cells are induced in BALB/c mice after gavage with bovine β-lactoglobulin. Allergy 2011, 66, 1312–1321. [Google Scholar]
- Van Bergenhenegouwen, J.; Braber, S.; Loonstra, R.; Buurman, N.; Rutten, L.; Knipping, K.; Savelkoul, P.J.; Harthoorn, L.F.; Jahnsen, F.L.; Garssen, J.; et al. Oral exposure to the free amino acid glycine inhibits the acute allergic response in a model of cow’s milk allergy in mice. Nutr. Res. 2018, 58, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Van der Kleij, H.P.M.; Warmenhoven, H.J.M.; van Ree, R.; Versteeg, S.A.; Pieters, R.H.H.; Dreskin, S.C.; Knulst, A.C.; Hoffen, E.V.; Opstelten, D.J.E.; Koppelman, S.J.; et al. Chemically modified peanut extract shows increased safety while maintaining immunogenicity. Allergy 2018, 65, 103. [Google Scholar]
- Wagenaar, L.; Bol-Schoenmakers, M.; Giustarini, G.; Vonk, M.M.; van Esch, B.C.A.M.; Knippels, L.M.J.; Garssen, J.; Smit, J.J.; Pieters, R.H.H. Dietary Supplementation with Nondigestible Oligosaccharides Reduces Allergic Symptoms and Supports Low Dose Oral Immunotherapy in a Peanut Allergy Mouse Model. Mol. Nutr. Food Res. 2018, 62, e1800369. [Google Scholar] [CrossRef] [PubMed]
- Ganeshan, K.; Neilsen, C.V.; Hadsaitong, A.; Schleimer, R.P.; Luo, X.; Bryce, P.J. Impairing oral tolerance promotes allergy and anaphylaxis: A new murine food allergy model. J. Allergy Clin. Immunol. 2009, 123, 231–238. [Google Scholar] [CrossRef]
- Tanaka, M.; Nagano, T.; Yano, H.; Matsuda, T.; Ikeda, T.M.; Haruma, K.; Kato, Y. Impact of ω-5 Gliadin on Wheat-Dependent Exercise-Induced Anaphylaxis in Mice. Biosci. Biotechnol. Biochem. 2011, 75, 313–317. [Google Scholar] [CrossRef]
- Katayama, S.; Yamaguchi, D.; Suzuki, Y.; Athamneh, A.M.A.; Mitani, T.; Satoh, R.; Teshima, R.; Mine, Y.; Nakamura, S. Oral Immunotherapy with a Phosphorylated Hypoallergenic Allergen Ameliorates Allergic Responses More Effectively Than Intact Allergen in a Murine Model of Buckwheat Allergy. Mol. Nutr. Food Res. 2018, 62, e1800303. [Google Scholar] [CrossRef]
- Rodriguez, M.J.; Aranda, A.; Fernandez, T.D.; Cubells-Baeza, N.; Torres, M.J.; Gomez, F.; Palomares, F.; Perkins, J.R.; Rojo, J.; Diaz-Perales, A.; et al. LPS promotes Th2 dependent sensitisation leading to anaphylaxis in a Pru p 3 mouse model. Sci. Rep. 2017, 7, 40449. [Google Scholar] [CrossRef]
- Snider, D.P.; Marshall, J.S.; Perdue, M.H.; Liang, H. Production of IgE antibody and allergic sensitization of intestinal and peripheral tissues after oral immunization with protein Ag and cholera toxin. J. Immunol. 1994, 153, 647–657. [Google Scholar] [PubMed]
- Mattsson, J.; Schön, K.; Ekman, L.; Fahlén-Yrlid, L.; Yrlid, U.; Lycke, N.Y. Cholera toxin adjuvant promotes a balanced Th1/Th2/Th17 response independently of IL-12 and IL-17 by acting on Gsα in CD11b+ DCs. Mucosal Immunol. 2015, 8, 815–827. [Google Scholar] [CrossRef] [PubMed]
- Eisenbarth, S.C.; Colegio, O.R.; O’Connor, W.; Sutterwala, F.S.; Flavell, R.A. Crucial role for the Nalp3 inflammasome in the immunostimulatory properties of aluminium adjuvants. Nature 2008, 453, 1122–1126. [Google Scholar] [CrossRef] [PubMed]
- Nyati, K.K.; Masuda, K.; Zaman, M.M.-U.; Dubey, P.K.; Millrine, D.; Chalise, J.P.; Higa, M.; Li, S.; Standley, D.M.; Saito, K.; et al. TLR4-induced NF-κB and MAPK signaling regulate the IL-6 mRNA stabilizing protein Arid5a. Nucleic Acids Res. 2017, 45, 2687–2703. [Google Scholar] [CrossRef] [PubMed]
- Fischer, R.; McGhee, J.R.; Vu, H.L.; Atkinson, T.P.; Jackson, R.J.; Tomé, D.; Boyaka, P.N. Oral and Nasal Sensitization Promote Distinct Immune Responses and Lung Reactivity in a Mouse Model of Peanut Allergy. Am. J. Pathol. 2005, 167, 1621–1630. [Google Scholar] [CrossRef]
- Bernard, H.; Creminon, C.; Wal, J.-M.; Adel-Patient, K.; Ah-Leung, S.; Adel-Patient, K.; Ah-Leung, S. Peanut- and cow’s milk-specific IgE, Th2 cells and local anaphylactic reaction are induced in Balb/c mice orally sensitized with cholera toxin. Allergy 2005, 60, 658–664. [Google Scholar]
- Sun, J.; Arias, K.; Alvarez, D.; Fattouh, R.; Walker, T.; Goncharova, S.; Kim, B.; Waserman, S.; Reed, J.; Coyle, A.J.; et al. Impact of CD40 Ligand, B Cells, and Mast Cells in Peanut-Induced Anaphylactic Responses. J. Immunol. 2007, 179, 6696–6703. [Google Scholar] [CrossRef] [PubMed]
- Gonipeta, B.; Kim, E.; Gangur, V. Mouse models of food allergy: how well do they simulate the human disorder? Crit. Rev. Food Sci. Nutr. 2015, 55, 437–452. [Google Scholar] [CrossRef] [PubMed]
- Proust, B.; Astier, C.; Jacquenet, S.; Ogier, V.; Magueur, E.; Roitel, O.; Belcourt, C.; Morisset, M.; Moneret-Vautrin, D.A.; Bihain, B.E.; et al. A Single Oral Sensitization to Peanut without Adjuvant Leads to Anaphylaxis in Mice. Int. Arch. Allergy Immunol. 2008, 146, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Oyoshi, M.K.; Oettgen, H.C.; Chatila, T.A.; Geha, R.S.; Bryce, P.J. Food allergy: Insights into etiology, prevention and treatment provided by murine models. J. Allergy Clin. Immunol. 2014, 133, 309–317. [Google Scholar] [CrossRef]
- Lack, G.; Fox, D.; Northstone, K.; Golding, J. Avon Longitudinal Study of Parents and Children Study Team Factors associated with the development of peanut allergy in childhood. N. Engl. J. Med. 2003, 348, 977–985. [Google Scholar] [CrossRef] [PubMed]
- Fox, D.E.; Lack, G. Peanut allergy. Lancet 1998, 352, 741. [Google Scholar] [CrossRef]
- Brown, S.J.; Asai, Y.; Cordell, H.J.; Campbell, L.E.; Zhao, Y.; Liao, H.; Northstone, K.; Henderson, J.; Alizadehfar, R.; Ben-Shoshan, M.; et al. Loss-of-function variants in the filaggrin gene are a significant risk factor for peanut allergy. J. Allergy Clin. Immunol. 2011, 127, 661–667. [Google Scholar] [CrossRef] [PubMed]
- A Palmer, C.N.; Irvine, A.D.; Terron-Kwiatkowski, A.; Zhao, Y.; Liao, H.; Lee, S.P.; Goudie, D.R.; Sandilands, A.; E Campbell, L.; Smith, F.J.D.; et al. Common loss-of-function variants of the epidermal barrier protein filaggrin are a major predisposing factor for atopic dermatitis. Nat. Genet. 2006, 38, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Bartnikas, L.M.; Gurish, M.F.; Burton, O.T.; Leisten, S.; Janssen, E.; Oettgen, H.C.; Beaupré, J.; Lewis, C.N.; Austen, K.F.; Schulte, S.; et al. Epicutaneous sensitization results in IgE-dependent intestinal mast cell expansion and food-induced anaphylaxis. J. Allergy Clin. Immunol. 2013, 131, 451–460. [Google Scholar] [CrossRef]
- Birmingham, N.P.; Parvataneni, S.; Hassan, H.M.A.; Harkema, J.; Samineni, S.; Navuluri, L.; Kelly, C.J.; Gangur, V. An Adjuvant-Free Mouse Model of Tree Nut Allergy Using Hazelnut as a Model Tree Nut. Int. Arch. Allergy Immunol. 2007, 144, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Strid, J.; Hourihane, J.; Kimber, I.; Callard, R.; Strobel, S. Epicutaneous exposure to peanut protein prevents oral tolerance and enhances allergic sensitization. Clin. Exp. Allergy 2005, 35, 757–766. [Google Scholar] [CrossRef]
- Galand, C.; Leyva-Castillo, J.-M.; Geha, R.S.; Yoon, J.; Oyoshi, M.K.; Han, A.; McKenzie, A.; Stassen, M. IL-33 Promotes Food Anaphylaxis in Epicutaneously-Sensitized Mice by Targeting Mast Cells. J. Allergy Clin. Immunol. 2016, 137, AB150. [Google Scholar] [CrossRef][Green Version]
- Wang, M.; Han, J.; Domenico, J.; Shin, Y.S.; Jia, Y.; Gelfand, E.W. Combined Blockade of the Histamine H1 and H4 Receptor Suppresses Peanut-Induced Intestinal Anaphylaxis by Regulating Dendritic Cell Function. Allergy 2016, 71, 1561–1574. [Google Scholar] [CrossRef]
- Song, Y.; Qu, C.; Srivastava, K.; Yang, N.; Busse, P.; Zhao, W.; Li, X.-M. Food Allergy Herbal Formula -2 protection against peanut anaphylactic reaction is via inhibition of mast cells and basophils. J. Allergy Clin. Immunol. 2010, 126, 1208–1217. [Google Scholar] [CrossRef]
- Fu, G.; Zhao, K.; Chen, H.; Wang, Y.; Nie, L.; Wei, H.; Wan, C. Effect of 3 lactobacilli on immunoregulation and intestinal microbiota in a β-lactoglobulin–induced allergic mouse model. J. Dairy Sci. 2019, 102, 1943–1958. [Google Scholar] [CrossRef] [PubMed]
- Burggraf, M.; Nakajima-Adachi, H.; Hachimura, S.; Ilchmann, A.; Pemberton, A.D.; Kiyono, H.; Vieths, S.; Toda, M. Oral tolerance induction does not resolve gastrointestinal inflammation in a mouse model of food allergy. Mol. Nutr. Food Res. 2011, 55, 1475–1483. [Google Scholar] [CrossRef] [PubMed]
- Mathias, C.B.; Hobson, S.A.; Garcia-Lloret, M.; Lawson, G.; Poddighe, D.; Freyschmidt, E.-J.; Xing, W.; Gurish, M.F.; Chatila, T.A.; Oettgen, H.C. IgE-mediated systemic anaphylaxis and impaired tolerance to food antigens in mice with enhanced IL-4 receptor signaling. J. Allergy Clin. Immunol. 2011, 127, 795–805. [Google Scholar] [CrossRef] [PubMed]
- Josefowicz, S.Z.; Niec, R.E.; Kim, H.Y.; Treuting, P.; Chinen, T.; Zheng, Y.; Umetsu, D.T.; Rudensky, A.Y. Extrathymically generated regulatory T cells control mucosal TH2 inflammation. Nature 2012, 482, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.D.; Maillard, M.H.; Cotta–De–Almeida, V.; Mizoguchi, E.; Klein, C.; Fuss, I.; Nagler, C.; Mizoguchi, A.; Bhan, A.K.; Snapper, S.B. Lymphocyte-Dependent and Th2 Cytokine-Associated Colitis in Mice Deficient in Wiskott-Aldrich Syndrome Protein. Gastroenterology 2007, 133, 1188–1197. [Google Scholar] [CrossRef] [PubMed]
- Lexmond, W.S.; Goettel, J.A.; Lyons, J.J.; Jacobse, J.; Deken, M.M.; Lawrence, M.G.; DiMaggio, T.H.; Kotlarz, D.; Garabedian, E.; Sackstein, P.; et al. FOXP3+ Tregs require WASP to restrain Th2-mediated food allergy. J. Clin. Investig. 2016, 126, 4030–4044. [Google Scholar] [CrossRef] [PubMed]
- Lexmond, W.S.; Goettel, J.A.; Sallis, B.F.; McCann, K.; Rings, E.H.H.M.; Jensen-Jarolim, E.; Nurko, S.; Snapper, S.B.; Fiebiger, E.; Jensen-Jarolim, E.; et al. Spontaneous food allergy in Was−/− mice occurs independent of FcεRI-mediated mast cell activation. Allergy 2017, 72, 1916–1924. [Google Scholar] [CrossRef]
- Platzer, B.; Baker, K.; Vera, M.P.; Singer, K.; Panduro, M.; Lexmond, W.S.; Turner, D.; Vargas, S.O.; Kinet, J.-P.; Maurer, D.; et al. Dendritic cell-bound IgE functions to restrain allergic inflammation at mucosal sites. Mucosal Immunol. 2015, 8, 516–532. [Google Scholar] [CrossRef]
- Weigmann, B.; Schughart, N.; Wiebe, C.; Sudowe, S.; Lehr, H.A.; Jonuleit, H.; Vogel, L.; Becker, C.; Neurath, M.F.; Grabbe, S.; et al. Allergen-induced IgE-dependent gut inflammation in a human PBMC–engrafted murine model of allergy. J. Allergy Clin. Immunol. 2012, 129, 1126–1135. [Google Scholar] [CrossRef]
- Ito, M.; Hiramatsu, H.; Kobayashi, K.; Suzue, K.; Kawahata, M.; Hioki, K.; Ueyama, Y.; Koyanagi, Y.; Sugamura, K.; Tsuji, K.; et al. NOD/SCID/gamma(c)(null) mouse: an excellent recipient mouse model for engraftment of human cells. Blood 2002, 100, 3175–3182. [Google Scholar] [CrossRef]
- Burton, O.T.; Stranks, A.J.; Tamayo, J.M.; Koleoglou, K.J.; Schwartz, L.B.; Oettgen, H.C. A humanized mouse model of anaphylactic peanut allergy. J. Allergy Clin. Immunol. 2017, 139, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Pagovich, O.E.; Wang, B.; Chiuchiolo, M.J.; Kaminsky, S.M.; Sondhi, D.; Jose, C.L.; Price, C.C.; Brooks, S.F.; Mezey, J.G.; Crystal, R.G. Anti-hIgE gene therapy of peanut-induced anaphylaxis in a humanized murine model of peanut allergy. J. Allergy Clin. Immunol. 2016, 138, 1652–1662. [Google Scholar] [CrossRef] [PubMed]
- Tachdjian, R.; Al Khatib, S.; Schwinglshackl, A.; Kim, H.S.; Chen, A.; Blasioli, J.; Mathias, C.; Kim, H.Y.; Umetsu, D.T.; Oettgen, H.C.; et al. In vivo regulation of the allergic response by the IL-4 receptor alpha chain immunoreceptor tyrosine-based inhibitory motif. J. Allergy Clin. Immunol. 2010, 125, 1128–1136. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Josefowicz, S.; Chaudhry, A.; Peng, X.P.; Forbush, K.; Rudensky, A.Y. Role of conserved non-coding DNA elements in the Foxp3 gene in regulatory T-cell fate. Nature 2010, 463, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Harrison, O.J.; Powrie, F.M. Regulatory T Cells and Immune Tolerance in the Intestine. Cold Spring Harb. Perspect. Boil. 2013, 5, a018341. [Google Scholar] [CrossRef]
- Klein, C.; Nguyen, D.; Liu, C.-H.; Mizoguchi, A.; Bhan, A.K.; Miki, H.; Takenawa, T.; Rosen, F.S.; Alt, F.W.; Mulligan, R.C.; et al. Gene therapy for Wiskott-Aldrich syndrome: Rescue of T-cell signaling and amelioration of colitis upon transplantation of retrovirally transduced hematopoietic stem cells in mice. Blood 2003, 101, 2159–2166. [Google Scholar] [CrossRef] [PubMed]
- Snapper, S.B.; Rosen, F.S. The Wiskott-Aldrich Syndrome Protein (WASP): Roles in Signaling and Cytoskeletal Organization. Annu. Rev. Immunol. 1999, 17, 905–929. [Google Scholar] [CrossRef] [PubMed]
- Sallmann, E.; Reininger, B.; Brandt, S.; Duschek, N.; Hoflehner, E.; Garner-Spitzer, E.; Platzer, B.; Dehlink, E.; Hammer, M.; Holcmann, M.; et al. High-Affinity IgE Receptors on Dendritic Cells Exacerbate Th2-Dependent Inflammation. J. Immunol. 2011, 187, 164–171. [Google Scholar] [CrossRef]
- Shultz, L.D.; Lyons, B.L.; Burzenski, L.M.; Gott, B.; Chen, X.; Chaleff, S.; Kotb, M.; Gillies, S.D.; King, M.; Mangada, J.; et al. Human lymphoid and myeloid cell development in NOD/LtSz-scid IL2R gamma null mice engrafted with mobilized human hemopoietic stem cells. J. Immunol. 2005, 174, 6477–6489. [Google Scholar]
- Ito, R.; Takahashi, T.; Katano, I.; Kawai, K.; Kamisako, T.; Ogura, T.; Ida-Tanaka, M.; Suemizu, H.; Nunomura, S.; Ra, C.; et al. Establishment of a Human Allergy Model Using Human IL-3/GM-CSF-Transgenic NOG Mice. J. Immunol. 2013, 191, 2890–2899. [Google Scholar] [CrossRef]
- Shultz, L.D.; Ishikawa, F.; Greiner, D.L. Humanized mice in translational biomedical research. Nat. Rev. Immunol. 2007, 7, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Piacentini, G.L.; Vicentini, L.; Bodini, A.; Mazzi, P.; Peroni, D.G.; Maffeis, C.; Boner, A.L. Allergenicity of a hydrolyzed rice infant formula in a guinea pig model. Ann. Allergy Asthma Immunol. 2003, 91, 61–64. [Google Scholar] [CrossRef]
- Abril-Gil, M.; Garcia-Just, A.; Pérez-Cano, F.J.; Franch, À.; Castell, M. Development and Characterization of an Effective Food Allergy Model in Brown Norway Rats. PLoS ONE 2015, 10, e0125314. [Google Scholar] [CrossRef] [PubMed]
- Helm, R.M.; Furuta, G.T.; Stanley, J.; Ye, J.; Cockrell, G.; Connaughton, C.; Simpson, P.; Bannon, G.A.; Burks, A. A neonatal swine model for peanut allergy. J. Allergy Clin. Immunol. 2002, 109, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Rupa, P.; Hamilton, K.; Cirinna, M.; Wilkie, B.N. A neonatal swine model of allergy induced by the major food allergen chicken ovomucoid (Gal d 1). Int. Arch. Allergy Immunol. 2008, 146, 11–18. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Adjuvant-Free Models | |||||
| Reference | Mouse Strain | Sensitization | Challenge | Read-Out | Comments |
| [59] | C3H/HeJ | 1 × 80 mg peanut extract, i.g. | 1 × 30 mg peanut extract, i.p. |
| |
| [68] | BALB/c | Three one-week cycles of tape stripping each followed by application of 100 µg Ova onto the skin | 1 × 100 mg of Ova, i.g. |
|
|
| Adjuvanted Mouse Food Allergy Models | |||||
| Reference | Mouse Strain | Sensitization | Challenge | Read-Out | Comments |
| [25] | Gnotobiotic C3H/HeN | 20 mg β-lactoglobulin + 10 µg cholera toxin, i.g. | 2 × 100 mg β-lactoglobulin, i.g. 30 min apart |
|
|
| [41] | BALB/c vs. C57BL/6 | 2 × 50 µg Ova + 1 mg Alum, i.p. | 1 × 50 mg Ova i.g. |
|
|
| [42] | BALB/c vs. 129ScEvBrd | 1 × 50 µg Ova + 2 mg Alum, s.c. | 1 × 50 mg Ova, i.g. |
| |
| [57] | C57BL/6 | 4 × 1 mg peanut protein + 10 µg cholera toxin | 1 × 5 mg peanut protein, i.p. |
| |
| [40] | C3H/HeJ | 2 × 5 or 25 mg peanut protein + 10 µg cholera toxin, i.g. | 1 × 10 mg peanut extract, i.g. |
| In this publication Li et al. defined a symptom score for the evaluation of the induced allergic reactions (see Table 2) which has proven very useful for many other mouse models. |
| [72] | BALB/c | 2 × 50 µg Ova plus 1 mg Alum, i.p. | Continuous challenge by feeding of Ova-containing food pellets for a period of seven days |
| |
| [50] | BALB/c | 6 × 20 µg Pru p 3 + 20 ng LPS, i.n. | 1 × 100 µg Pru p 3, i.p. |
| Only available Pru p 3 allergy model |
| [48] | B10.A | 1 × 100 µg + 5x 50 µg wheat gliadin fraction + 4 mg Alum, i.p. | 1 × 10 mg gliadin, i.g. + 30 min of strenuous physical treadmill exercise (15 m/min, 20 % gradient) |
| Mouse model of exercise induced anaphylaxis. |
| Mouse Models using Genetically-modified Mouse Strains | |||||
| Reference | Mouse Strain | Sensitization | Challenge | Read-Out | Comments |
| [73] | IL-4RaF709 | 12 × 5 mg Ova + 20 µg cholera toxin, i.g. OR 12 × 5 mg Ova without adjuvant, i.g. | 1 × 150 mg Ova, i.g. |
| Due to the Th2-promoting milieu IL-4raF709 can be sensitized to allergen without adjuvant |
| [74] | Foxp3 CNS1−/− | Not performed | Not performed |
|
|
| [75] | WAS−/− | Not performed | Not performed |
| |
| [76] | WAS−/− |
| Mechanistically, WASP-deficiency drives Th2 responses by interfering with normal Treg function | ||
| [77] | WAS−/− | 7 × 5 mg Ova without adjuvant, i.g., v.s. 3 × 50 µg Ova + 100 µL Alum, i.p. | 1 × 50 mg Ova |
| WAS−/− mice mimic both polysensitization and highly bioactive, but low-level IgE production frequently observed in human food allergic patients |
| Humanized Mouse Models | |||||
| Reference | Mouse Strain | Humanization & Sensitization | Challenge | Read-Out | Comments |
| [78] | Mice expressing the α-chain of human FcεRI in a CD11c-restricted manner | 2 × 100 µg Ova mixed 1:1 with Alum, i.p. | 3–6 × 50 mg Ova, i.g. |
| IgE-signaling in human DCs is involved in down-regulating allergic mucosal inflammation |
| [79] | NOD-SCID-γc−/− | I.p. injection of 2 × 107 PBMC from allergic donors with high allergen-specific IgE levels in combination with the respective allergen (20 µg) followed by i.p. boost with 20 µg allergen 8 days later | 1 × 20 µg allergen rectally or 50 µg allergen orally (d21) |
| Blocking experiments demonstrated that gut inflammation in this model was mediated by human IgE |
| [80] | NOG IL-3/GM-Tg mice | 1 × 105 or 4 × 104 human umbilical cord blood-derived CD34+ hematopoietic stem cells i.v. 24 h after irradiation | Not performed |
| |
| [81] | NOD-SCID-γc−/− stem cell factor (SCF)-Tg mice | 5 × 104–105 human CD34+ cord blood stem cells, i.v., 6 weeks later sensitization: 8 × 22.5 mg peanut butter without adjuvant, i.g. | 1 × 350 mg peanut butter, i.g. |
| Humanized mouse models can be used to recreate the multistep and highly complex human allergic responses |
| [82] | NSG | 3 × 107 PBMC from peanut allergic individuals mixed with 100 µg peanut extract (i.p. split in 2 injections sites), followed by weekly i.p. injections (4×) with 100 µg peanut extract | 4 × weekly 300 µg peanut extract, i.g. |
| Humanized mouse model was used to compare the effectiveness of an experimental adeno-associated virus (AAV)-based expression of anti-human IgE to the already established anti-human IgE treatment with Omalizumab |
| Score | Symptoms |
|---|---|
| 0 | No symptoms |
| 1 | Scratching and rubbing around nose and head |
| 2 | Puffiness around eyes and mouth, diarrhea, pillar erect, reduced activity, and/or decreased activity with increased respiratory rate |
| 3 | Wheezing, labored respiration, and cyanosis around mouth and tail |
| 4 | No activity after prodding or tremor and convulsion |
| 5 | Death |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schülke, S.; Albrecht, M. Mouse Models for Food Allergies: Where Do We Stand? Cells 2019, 8, 546. https://doi.org/10.3390/cells8060546
Schülke S, Albrecht M. Mouse Models for Food Allergies: Where Do We Stand? Cells. 2019; 8(6):546. https://doi.org/10.3390/cells8060546
Chicago/Turabian StyleSchülke, Stefan, and Melanie Albrecht. 2019. "Mouse Models for Food Allergies: Where Do We Stand?" Cells 8, no. 6: 546. https://doi.org/10.3390/cells8060546
APA StyleSchülke, S., & Albrecht, M. (2019). Mouse Models for Food Allergies: Where Do We Stand? Cells, 8(6), 546. https://doi.org/10.3390/cells8060546