Mitotic Chromosomes in Live Cells Characterized Using High-Speed and Label-Free Optical Diffraction Tomography


 , ,
, ,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells
2.2. Osmotic Concentration, Refractive Index (RI), and Culture Media
2.3. Optical Diffraction Tomography (ODT)
2.4. Image Rendering, RI, and Volume Calculation
2.5. Confocal Laser-Scanning Microscopy (CLSM)
2.6. Fluorescence Correlation Spectroscopy (FCS)
2.7. Statistical Analysis
3. Results and Discussion
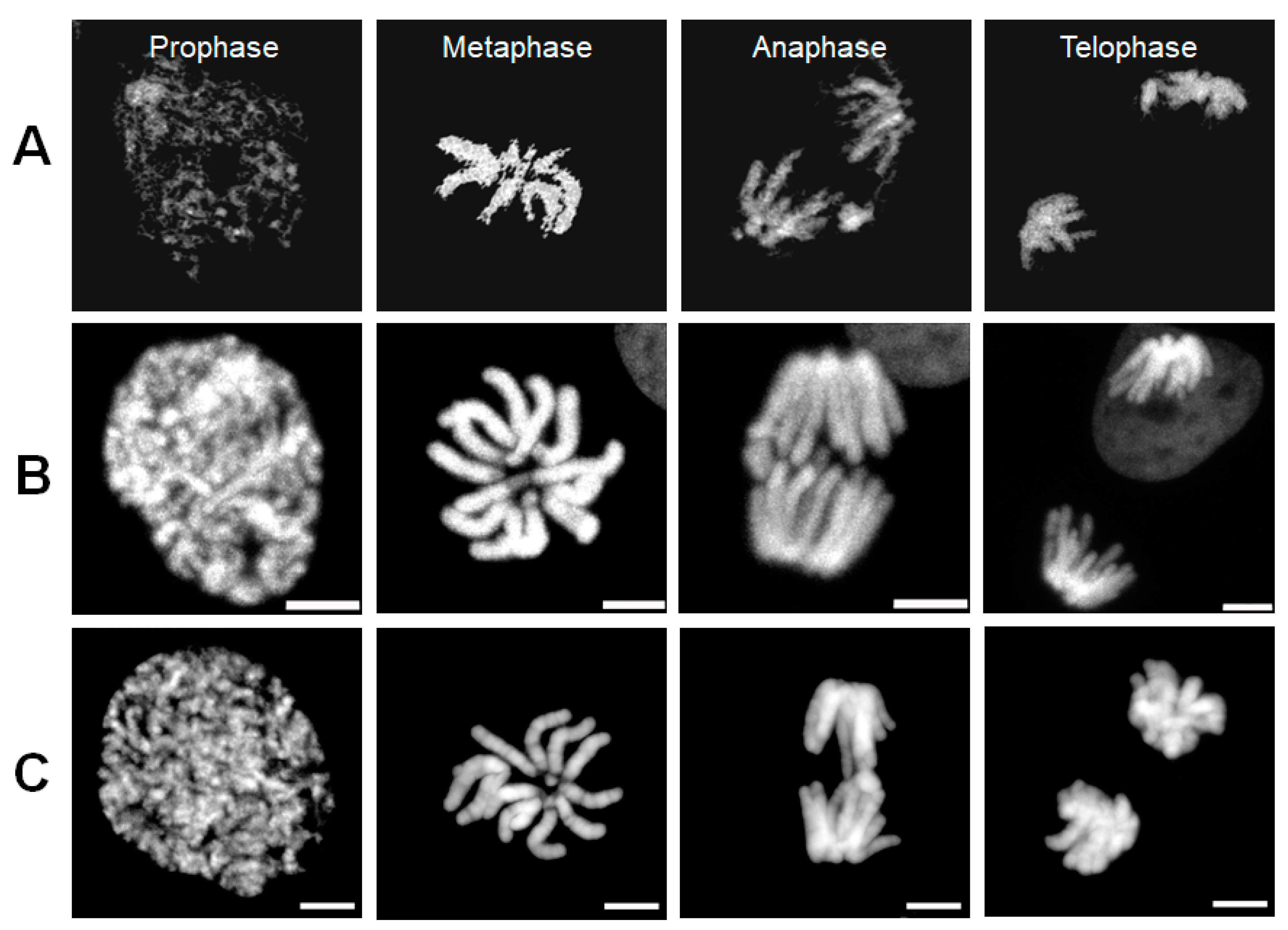
3.1. Comparisons of Images of Chromosomes Acquired Using ODT and 3D CLSM
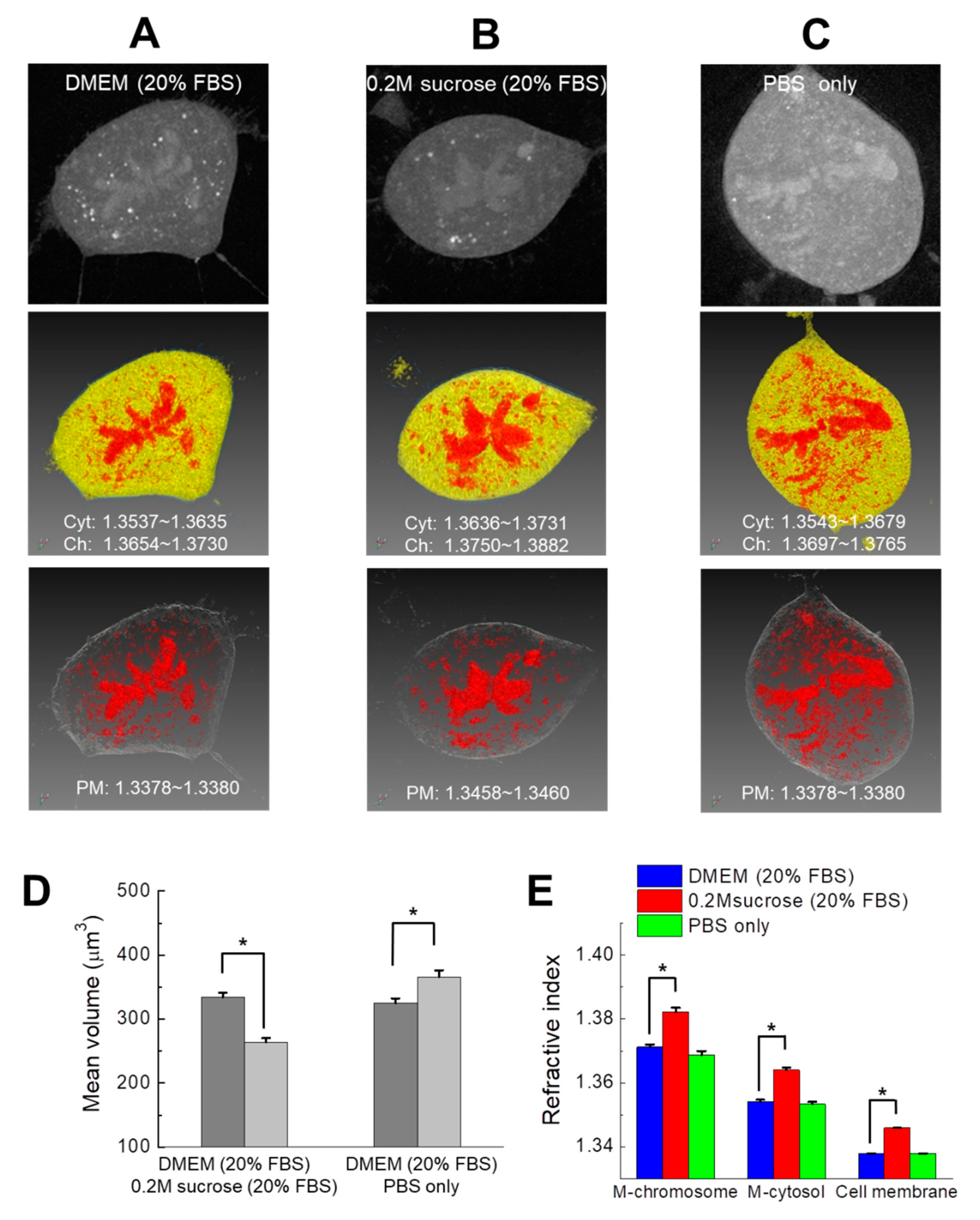
3.2. Quantification of the RI Values of Mitotic Chromosomes as a Function of Osmotic Conditions
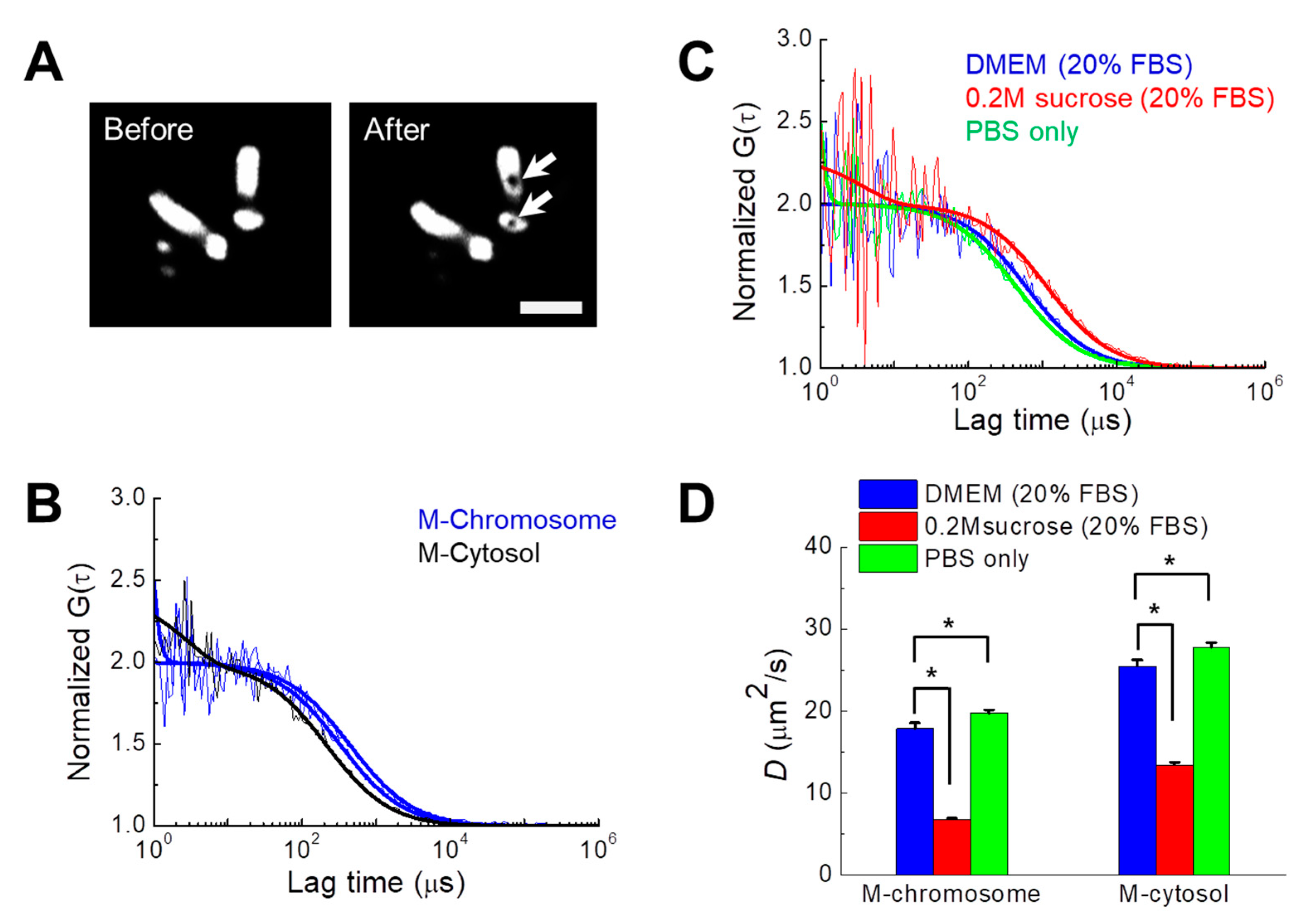
3.3. Quantification of the Diffusional Mobility of Monomeric Green Fluorescent Protein (mGFP) in Mitotic Chromosomes under Different Osmotic Conditions
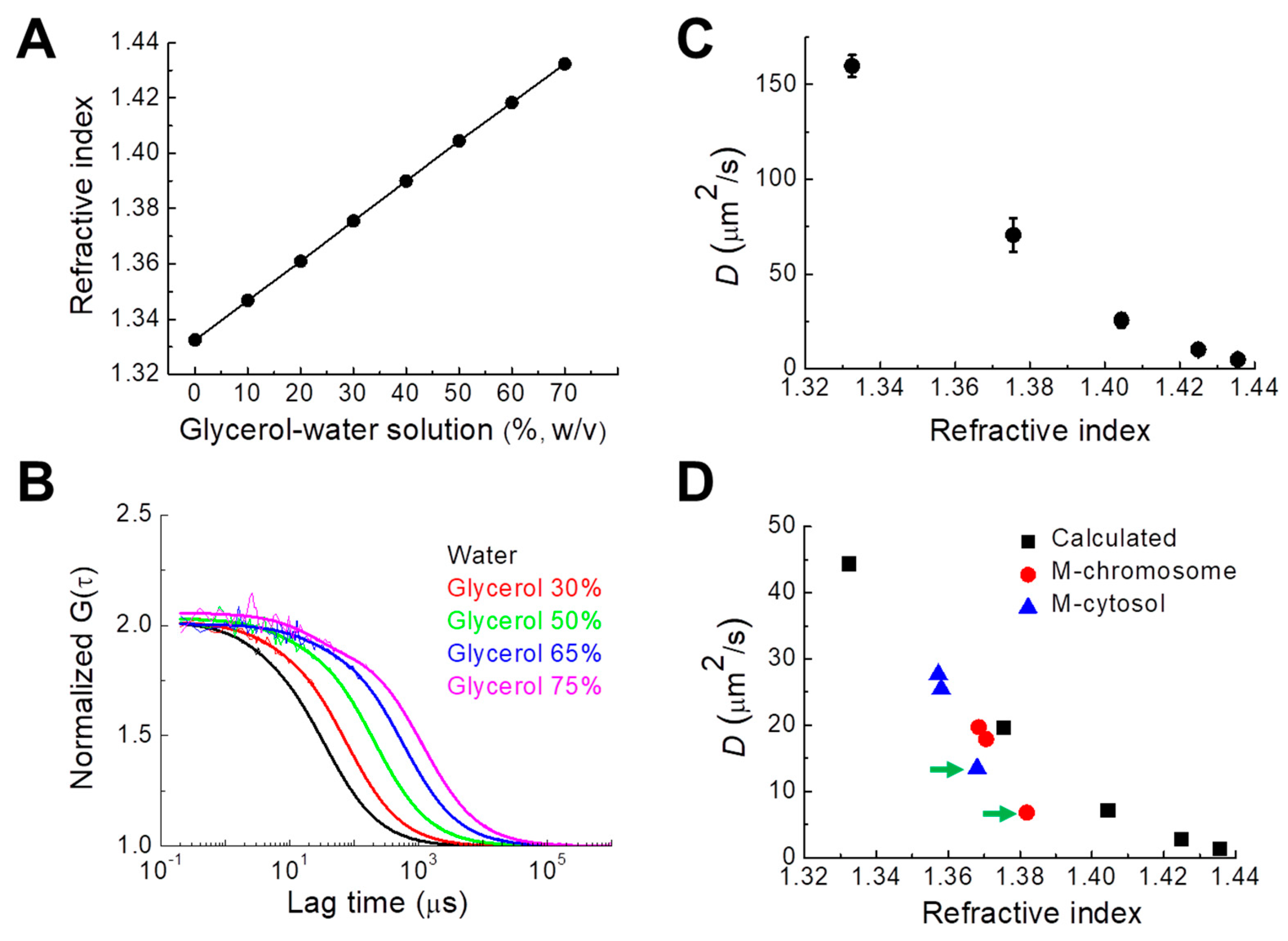
3.4. Relationship between Chromosomal RI and Diffusion of Intrachromosomal mGFP
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pack, C.; Saito, K.; Tamura, M.; Kinjo, M. Microenvironment and effect of energy depletion in the nucleus analyzed by mobility of multiple oligomeric EGFPs. Biophys. J. 2006, 91, 3921–3936. [Google Scholar] [CrossRef] [PubMed]
- Wachsmuth, M.; Caudron-Herger, M.; Rippe, K. Genome organization: Balancing stability and plasticity. Biochim. Biophys. Acta 2008, 1783, 2061–2079. [Google Scholar] [CrossRef] [PubMed]
- Hihara, S.; Pack, C.G.; Kaizu, K.; Tani, T.; Hanafusa, T.; Nozaki, T.; Takemoto, S.; Yoshimi, T.; Yokota, H.; Imamoto, N.; et al. Local nucleosome dynamics facilitate chromatin accessibility in living mammalian cells. Cell Rep. 2012, 2, 1645–1656. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Han, S.S.; Sako, Y.; Pack, C.G. Dynamic and unique nucleolar microenvironment revealed by fluorescence correlation spectroscopy. FASEB J. 2015, 29, 837–848. [Google Scholar] [CrossRef]
- Hancock, R. A role for macromolecular crowding effects in the assembly and function of compartments in the nucleus. J. Struct. Biol. 2004, 146, 281–290. [Google Scholar] [CrossRef]
- Richter, K.; Nessling, M.; Lichter, P. Experimental evidence for the influence of molecular crowding on nuclear architecture. J. Cell. Sci. 2007, 120, 1673–1680. [Google Scholar] [CrossRef]
- Richter, K.; Nessling, M.; Lichter, P. Macromolecular crowding and its potential impact on nuclear function. Biochim. Biophys. Acta 2008, 1783, 2100–2107. [Google Scholar] [CrossRef]
- Matera, A.G.; Izaguire-Sierra, M.; Praveen, K.; Rajendra, T.K. Nuclear bodies: Random aggregates of sticky proteins or crucibles of macromolecular assembly? Dev. Cell 2009, 17, 639–647. [Google Scholar] [CrossRef]
- Gnutt, D.; Gao, M.; Brylski, O.; Heyden, M.; Ebbinghaus, S. Excluded-volume effects in living cells. Angew Chem. Int. Ed. Engl. 2015, 54, 2548–2551. [Google Scholar] [CrossRef]
- Verkman, A.S. Solute and macromolecule diffusion in cellular aqueous compartments. Trends Biochem. Sci. 2002, 27, 27–33. [Google Scholar] [CrossRef]
- Konopka, M.C.; Sochacki, K.A.; Bratton, B.P.; Shkel, I.A.; Record, M.T.; Weisshaar, J.C. Cytoplasmic protein mobility in osmotically stressed Escherichia coli. J. Bacteriol. 2009, 191, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Finan, J.D.; Leddy, H.A.; Guilak, F. Osmotic stress alters chromatin condensation and nucleocytoplasmic transport. Biochem. Biophys. Res. Commun. 2011, 408, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Mika, J.T.; Schavemaker, P.E.; Krasnikov, V.; Poolman, B. Impact of osmotic stress on protein diffusion in Lactococcus lactis. Mol. Microbiol. 2014, 94, 857–870. [Google Scholar] [CrossRef] [PubMed]
- Schavemaker, P.E.; Boersma, A.J.; Poolman, B. How Important Is Protein Diffusion in Prokaryotes? Front. Mol. Biosci. 2018, 5, 93. [Google Scholar] [CrossRef] [PubMed]
- Magde, D.; Elson, E.L.; Webb, W.W. Fluorescence correlation spectroscopy. II. An experimental realization. Biopolymers 1974, 13, 29–61. [Google Scholar] [CrossRef]
- Eigen, M.; Rigler, R. Sorting single molecules: Application to diagnostics and evolutionary biotechnology. Proc. Natl. Acad. Sci. USA 1994, 91, 5740–5747. [Google Scholar] [CrossRef]
- Erdel, F.; Muller-Ott, K.; Baum, M.; Wachsmuth, M.; Rippe, K. Dissecting chromatin interactions in living cells from protein mobility maps. Chromosome. Res. 2001, 19, 99–115. [Google Scholar] [CrossRef]
- Brunsting, A.; Mullaney, P.F. Differential light scattering from spherical mammalian cells. Biophys. J. 1974, 14, 439–453. [Google Scholar] [CrossRef]
- Choi, W.; Fang-Yen, C.; Badizadegan, K.; Oh, S.; Lue, N.; Dasari, R.R.; Feld, M.S. Tomographic phase microscopy. Nat. Methods 2007, 4, 717–719. [Google Scholar] [CrossRef]
- Kemper, B.; Vollmer, A.; Rommel, C.E.; Schnekenburger, J.; Von Bally, G. Simplified approach for quantitative digital holographic phase contrast imaging of living cells. J. Biomed. Opt. 2001, 16, 026014. [Google Scholar] [CrossRef]
- Kim, K.; Kim, K.S.; Park, H.; Ye, J.C.; Park, Y. Real-time visualization of 3-D dynamic microscopic objects using optical diffraction tomography. Opt. Express 2013, 21, 32269–32278. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Yoon, H.; Diez-Silva, M.; Dao, M.; Dasari, R.R.; Park, Y. High-resolution three-dimensional imaging of red blood cells parasitized by Plasmodium falciparum and in situ hemozoin crystals using optical diffraction tomography. J. Biomed. Opt. 2014, 19, 011005. [Google Scholar] [CrossRef] [PubMed]
- Wolf, E. Three-dimensional structure determination of semi-transparent objects from holographic data. Opt. Commun. 1969, 1, 153–156. [Google Scholar] [CrossRef]
- Liu, P.Y.; Chin, L.K.; Ser, W.; Chen, H.F.; Hsieh, C.M.; Lee, C.H.; Sung, K.B.; Ayi, T.C.; Yap, P.H.; Liedberg, B.; et al. Cell refractive index for cell biology and disease diagnosis: Past, present and future. Lab. Chip 2016, 16, 634–644. [Google Scholar] [CrossRef]
- Sung, Y.; Choi, W.; Lue, N.; Dasari, R.R.; Yaqoob, Z. Stain-free quantification of chromosomes in live cells using regularized tomographic phase microscopy. PLoS ONE 2012, 7, e49502. [Google Scholar] [CrossRef]
- Ma, L.; Rajshekhar, G.; Wang, R.; Bhaduri, B.; Sridharan, S.; Mir, M.; Chakraborty, A.; Iyer, R.; Prasanth, S.; Millet, L.; et al. Phase correlation imaging of unlabeled cell dynamics. Sci. Rep. 2016, 6, 32702. [Google Scholar] [CrossRef]
- Park, Y.; Diez-Silva, M.; Fu, D.; Popescu, G.; Choi, W.; Barman, I.; Suresh, S.; Feld, M.S. Static and dynamic light scattering of healthy and malaria-parasite invaded red blood cells. J. Biomed. Opt. 2010, 15, 020506. [Google Scholar] [CrossRef]
- Kim, T.K.; Lee, B.W.; Fujii, F.; Kim, J.K.; Pack, C.G. Physicochemical Properties of Nucleoli in Live Cells Analyzed by Label-Free Optical Diffraction Tomography. Cells 2019, 8, 699. [Google Scholar] [CrossRef]
- Kim, G.; Lee, M.; Youn, S.; Lee, E.; Kwon, D.; Shin, J.; Lee, S.; Lee, Y.S.; Park, Y. Measurements of three-dimensional refractive index tomography and membrane deformability of live erythrocytes from Pelophylax nigromaculatus. Sci. Rep. 2018, 8, 9192. [Google Scholar] [CrossRef]
- Jung, J.; Kim, K.; Yoon, J.; Park, Y. Hyperspectral optical diffraction tomography. Opt. Express 2016, 24, 2006–2012. [Google Scholar] [CrossRef]
- Kang, M.H.; Choi, H.; Oshima, M.; Cheong, J.H.; Kim, S.; Lee, J.H.; Park, Y.S.; Choi, H.S.; Kweon, M.N.; Pack, C.G.; et al. Estrogen-related receptor gamma functions as a tumor suppressor in gastric cancer. Nat. Commun. 2018, 9, 1920. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Park, W.S.; Na, S.; Kim, S.; Kim, T.; Do Heo, W.; Park, Y. Correlative three-dimensional fluorescence and refractive index tomography: Bridging the gap between molecular specificity and quantitative bioimaging. Biomed. Opt. Express 2017, 8, 5688–5697. [Google Scholar] [CrossRef] [PubMed]
- Robbins, E.; Pederson, T.; Klein, P. Comparison of mitotic phenomena and effects induced by hypertonic solutions in HeLa cells. J. Cell. Biol. 1970, 44, 400–416. [Google Scholar] [CrossRef] [PubMed]
- Delpire, E.; Duchene, C.; Goessens, G.; Gilles, R. Effects of osmotic shocks on the ultrastructure of different tissues and cell types. Exp. Cell. Res. 1985, 160, 106–116. [Google Scholar] [CrossRef]
- Albiez, H.; Cremer, M.; Tiberi, C.; Vecchio, L.; Schermelleh, L.; Dittrich, S.; Kupper, K.; Joffe, B.; Thormeyer, T.; Von Hase, J.; et al. Chromatin domains and the interchromatin compartment form structurally defined and functionally interacting nuclear networks. Chromosome. Res. 2006, 14, 707–733. [Google Scholar] [CrossRef]
- Hancock, R. Structure of metaphase chromosomes: A role for effects of macromolecular crowding. PLoS ONE 2012, 7, e36045. [Google Scholar] [CrossRef]
- Weast, R.C. CRC Handbook of Chemistry and Physics; CRC Press: Boca Raton, FL, USA, 1988. [Google Scholar]
- Dix, J.A.; Verkman, A.S. Crowding effects on diffusion in solutions and cells. Annu Rev. Biophys 2008, 37, 247–263. [Google Scholar] [CrossRef]
- Golkaram, M.; Hellander, S.; Drawert, B.; Petzold, L.R. Macromolecular Crowding Regulates the Gene Expression Profile by Limiting Diffusion. PLoS Comput. Biol. 2016, 12, e1005122. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DMEM a (20% FBS) | 0.2 M Sucrose b (20% FBS) | DMEM c (10% FBS) | 0.2M Sucrose (10% FBS) | PBS only | Distilled Water (Reference) | |
|---|---|---|---|---|---|---|
| Osmolarity (mOsm/kg) | ~340 | ~500 | ~330 | ~500 | ~290 | - |
| RI (25 °C) | 1.3365 | 1.3456 | 1.3358 | 1.3447 | 1.3349 | 1.3325 |
| RI (37 °C) | 1.3350 | 1.3441 | 1.3342 | 1.3431 | 1.3336 | 1.3310 |
| Media Condition | M-Chromosomes | M-Cytosol | Cell Membrane (Control) d |
|---|---|---|---|
| DMEM (20% FBS) a | 1.3712 ± 0.0009 | 1.3541 ± 0.0006 | 1.3379 |
| 0.2 M sucrose b (20% FBS) | 1.3822 ± 0.0013 | 1.3640 ± 0.0008 | 1.3459 |
| PBS c only | 1.3688 ± 0.0012 | 1.3534 ± 0.0007 | 1.3379 |
| Media Condition | M-Chromosome (μm2/s) | M-Cytosol (μm2/s) |
|---|---|---|
| DMEM (20% FBS) | 17.9 ± 0.7 | 25.5 ± 0.8 |
| 0.2 M sucrose (20% FBS) | 6.8 ± 0.2 | 13.5 ± 0.3 |
| PBS only | 19.7 ± 0.4 | 27.7 ± 0.6 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, T.-K.; Lee, B.-W.; Fujii, F.; Lee, K.-H.; Lee, S.; Park, Y.; Kim, J.K.; Lee, S.-W.; Pack, C.-G. Mitotic Chromosomes in Live Cells Characterized Using High-Speed and Label-Free Optical Diffraction Tomography. Cells 2019, 8, 1368. https://doi.org/10.3390/cells8111368
Kim T-K, Lee B-W, Fujii F, Lee K-H, Lee S, Park Y, Kim JK, Lee S-W, Pack C-G. Mitotic Chromosomes in Live Cells Characterized Using High-Speed and Label-Free Optical Diffraction Tomography. Cells. 2019; 8(11):1368. https://doi.org/10.3390/cells8111368
Chicago/Turabian StyleKim, Tae-Keun, Byong-Wook Lee, Fumihiko Fujii, Kee-Hang Lee, Sanghwa Lee, YongKeun Park, Jun Ki Kim, Sang-Wook Lee, and Chan-Gi Pack. 2019. "Mitotic Chromosomes in Live Cells Characterized Using High-Speed and Label-Free Optical Diffraction Tomography" Cells 8, no. 11: 1368. https://doi.org/10.3390/cells8111368
APA StyleKim, T.-K., Lee, B.-W., Fujii, F., Lee, K.-H., Lee, S., Park, Y., Kim, J. K., Lee, S.-W., & Pack, C.-G. (2019). Mitotic Chromosomes in Live Cells Characterized Using High-Speed and Label-Free Optical Diffraction Tomography. Cells, 8(11), 1368. https://doi.org/10.3390/cells8111368