
From 2D Myotube Cultures to 3D Engineered Skeletal Muscle Constructs: A Comprehensive Review of In Vitro Skeletal Muscle Models and Disease Modeling Applications


Abstract
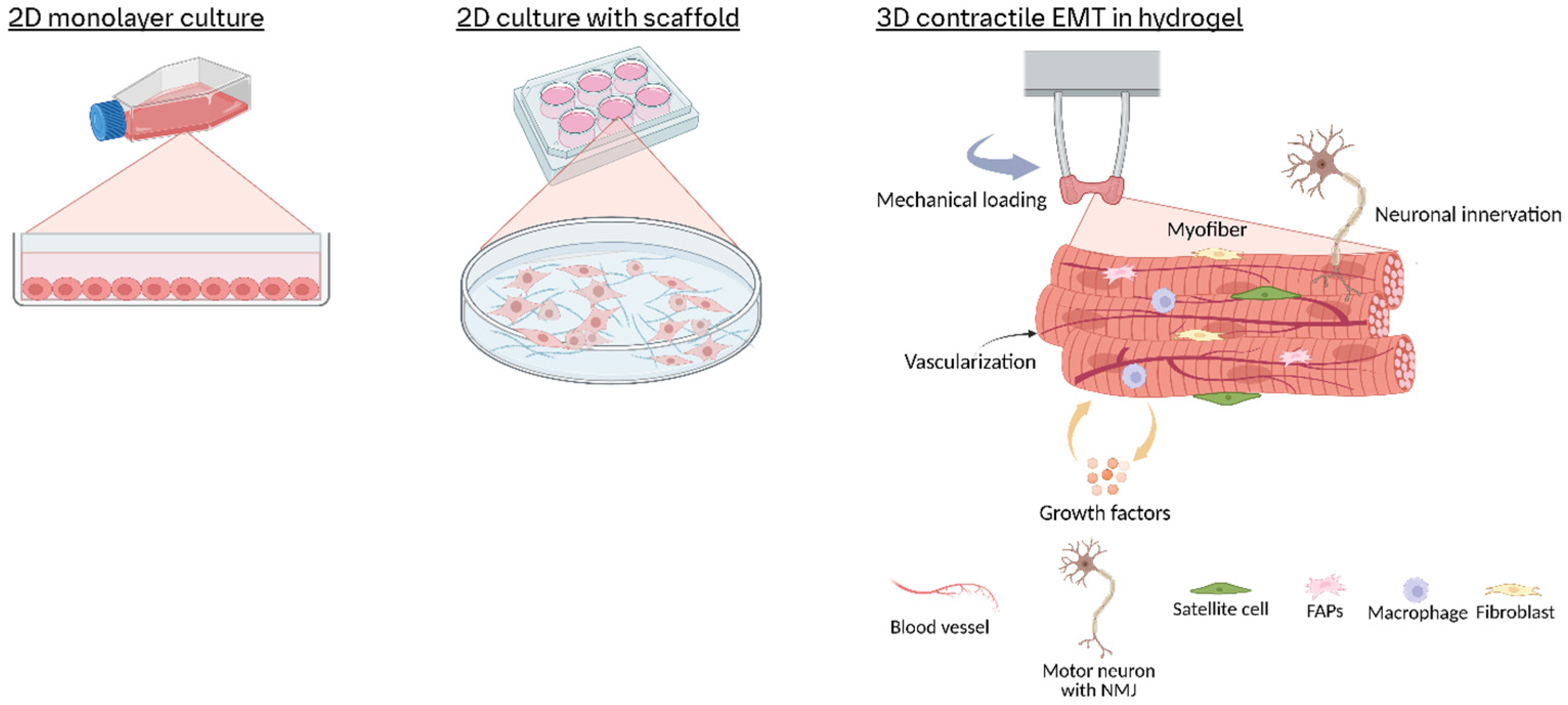
1. Introduction
2. Overview of In Vitro Skeletal Muscle Models
2.1. Conventional 2D Cell Culture
2.2. Transition to 3D Models: Bioengineering Strategies for 3D Skeletal Muscle Models
2.2.1. Scaffold Selection and Microarchitectural Design

{kind=link}
| 2D or 3D | Scaffold Materials | Cell Type/Model Used | Contractility Measurement | Other Key Findings | Advantages | Disadvantages | References |
|---|---|---|---|---|---|---|---|
| 2D; Thin fibrous meshes (~0.1–0.3 mm thick) | Poly(lactide-co-glycolide) PLGA | C2C12 | N/A | • Aligned PLGA fibers induced myoblast elongation, alignment, and differentiation into multinucleated myotubes. • Enhanced differentiation on aligned fibers (higher fusion index vs. random fibers/glass controls). • No biologic components (e.g., collagen/Matrigel) required for adhesion or differentiation | Synthetic, biodegradable scaffold with tunable degradation (PLGA); Compatible with long-term culture (no pH shifts or detachment issues) | No direct contractility measurement; Lack of cell infiltration due to dense fibrous structure | [43] |
| 2D; Electrospun fibrous mats (quasi-3D environment) | Poly(butylene 1,4-cyclohexanedicarboxylate) (PBCE) and copolymers with triethylene cyclohexanedicarboxylate [P(BCE-co-TECE)] | C2C12 | N/A | • The copolymer with higher “TECE” content (P73) promoted better C2C12 proliferation, alignment, and differentiation. • In vivo, the P73 scaffold became vascularized and integrated into both healthy and injured/dystrophic mouse muscle with minimal inflammation | Chemical tunability; Suitable for muscle repair | No direct contractility data | [52] |
| 2D with 3D-like environment | Poly-IL-lactic acid (PLLA) nanofibers (~500 nm diameter; ~150 µm thick) | C2C12 | N/A | Aligned nanofibers promote global myotube alignment, greater myotube length, and enhanced differentiation | Mimic native ECM and drive robust, aligned myotube formation. | Limited to thin-film culture; no direct functional (contractility) data. | [46] |
| 2D with nanofibrous mat | Nanofibers made from a blend of polycaprolactone (PCL) and polyaniline (PANi) | C2C12 | N/A | Aligned nanofibers and increased PANi content synergistically promote enhanced myotube alignment, longer myotubes, and upregulated myogenic genes | Mimics native ECM topography; incorporates electrical stimulation to boost differentiation; tunable via PANi concentration | Lacks direct contractile function | [47] |
| 2D with thin nanofibrous scaffold (~10–15 μm thick) | PCL blended with 4% gelatin | C2C12 | Assessed by calcium imaging (luminescence) under high-K+ and electrical stimulation (0.2–2 Hz). Mechanically stimulated aligned scaffolds showed enhanced contractility | Mechanical stimulation increases myotube width, actin density, and contractile function | Biomimetic nanofibrous architecture promotes adhesion and differentiation; mechanical stimulation further enhances regeneration | Low force production at high frequency stimulation in 2D | [55] |
| 3D tubular construct | Chitosan/polycaprolactone (CS/PCL) blend | C2C12 | N/A | Direct electrospinning of aligned 3D nanofibrous tubes; The inner surface supports formation of densely aligned myotubes mimicking native muscle architecture | 3D fabrication with tunable scaffold dimensions | Limited function assessment; Long-term degradation | [50] |
| 3D-bioprinted muscle tissue | GelMA-based bioink | C2C12 | Under electrical stimulation (1 Hz), the engineered tissues generated a twitch contraction force of 443.085 μN for pennate muscle versus 239.662 μN for parallel muscle | • Pennate design (15° fiber orientation) significantly improved myotube alignment (51.93%) and enhanced contractile performance. • High cell viability (~79.89% by day 7) and uniform cell distribution were achieved. | 3D bioprinting allows precise, customizable fabrication with complex microstructures; Direct contractility measurement | Use of murine C2C12 cells may not fully represent human muscle functionality. | [51] |
| 3D engineered muscle | Fibrin-based gel | Primary rat myoblasts | Twitch force is 329 ± 26.3 μN and tetanic force is 805.8 ± 55 μN under electric stimulation | • Normal length-tension and force-frequency relationships; 50% force increase in response to IGF-I. | • Rapid, scaffold-free self-organization with minimal extracellular material. • Functional properties that mimic native muscle physiology. • Culture up to 6 weeks | Weaker contractile force due to the soft fibrin-based hydrogel | [56] |
| 3D | Fibrin hydrogels enriched with Laminin-111 | C2C12 | N/A | • LM-111 enrichment produces a highly fibrous 3D architecture with thinner, more interconnected fibers • Lower Young’s modulus (2–6 kPa) favorable for myoblast proliferation • Enhanced VEGF, MyoD, and desmin expression, reduced IL-6 and myogenin at high LM-111 levels | Biomimetic ECM: mimics basal lamina composition and structure, and promotes a pro-regenerative secretory profile to supports cell adhesion, migration, and proliferation | High LM-111 (>500 µg/mL) impairs gel stability | [57] |
| 3D | Fibrin + 10% Matrigel | Human primary myoblasts + HUVECs | N/A | Aligned myofibers and endothelial networks with 2 × 106 total cells (50–70% muscle cells) Matrigel addition leads to uneven myofiber distribution. | Fibrin is proangiogenic and supports simultaneous myofiber alignment and vascular network formation | No direct contractility; HUVECs culture medium results in thinner myotubes. | [58] |
| 3D | Collagen-1 + fibrin gel | Primary rat myoblasts | N/A | Hybrid gel prevents contraction; Higher cell proliferation in low-fibrin groups. | Hydrogel is biocompatible and mimicking natural ECM for myoblast differentiation. | Gel instability over time. Protease inhibitor, aprotinin is required. | [59] |
| 3D | Collagen sponges/OPLA (open-cell poly-lactic-acid) scaffolds | Primary rat myoblasts | N/A | High apoptosis (45–66% in sponges); OPLA lacks elasticity for muscle. | Sponges is high stability and synthetic OPLA is biodegradable. | Poor cell viability. • Unsuitable pore structure (sponges). • Rigid (OPLA). | [59] |
| 3D | Decellularized skeletal muscle ECM (mdECM) bioink + PCL constraints | C2C12 | Observed spontaneous contractions upon eletrical stimulation without quantitative measurement | Elastic modulus ≈ 12 kPa (similar to native muscle); Striated myotubes formed. | High cell viability (>90%); Customizable architecture via 3D printing. | Requires specialized 3D printing setup. | [60] |
2.2.2. Perfusion System Integration
2.2.3. In Vitro Neuromuscular Junction (NMJ) Formation and Integration into Engineered Models
2.2.4. Co-Culture Strategies with Supporting Cell Types
2.3. Modeling Muscular Disease in 3D Constructs
2.3.1. Mendelian Muscular Disorders
2.3.2. Polygenic (Common) Muscle Diseases
2.4. Current Challenges and Future Directions
2.4.1. Technical Challenges
2.4.2. Scalability
2.4.3. Future Directions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sanger, J.W.; Wang, J.; Fan, Y.; White, J.; Mi-Mi, L.; Dube, D.K.; Sanger, J.M.; Pruyne, D. Assembly and Maintenance of Myofibrils in Striated Muscle. Handb. Exp. Pharmacol. 2017, 235, 39–75. [Google Scholar] [CrossRef] [PubMed]
- Rassier, D.E. Sarcomere Mechanics in Striated Muscles: From Molecules to Sarcomeres to Cells. Am. J. Physiol. Cell Physiol. 2017, 313, C134–C145. [Google Scholar] [CrossRef] [PubMed]
- Yanagida, T.; Arata, T.; Oosawa, F. Sliding Distance of Actin Filament Induced by a Myosin Crossbridge during One ATP Hydrolysis Cycle. Nature 1985, 316, 366–369. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, M.; Spriet, L.L. Skeletal Muscle Energy Metabolism during Exercise. Nat. Metab. 2020, 2, 817–828. [Google Scholar] [CrossRef]
- Merz, K.E.; Thurmond, D.C. Role of Skeletal Muscle in Insulin Resistance and Glucose Uptake. Compr. Physiol. 2020, 10, 785–809. [Google Scholar] [CrossRef]
- Sanes, J.R.; Lichtman, J.W. Development of the Vertebrate Neuromuscular Junction. Annu. Rev. Neurosci. 1999, 22, 389–442. [Google Scholar] [CrossRef]
- Tidball, J.G. Inflammatory Processes in Muscle Injury and Repair. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R345–R353. [Google Scholar] [CrossRef]
- Burden, S.J. Building the Vertebrate Neuromuscular Synapse. J. Neurobiol. 2002, 53, 501–511. [Google Scholar] [CrossRef]
- Schiaffino, S.; Reggiani, C. Fiber Types in Mammalian Skeletal Muscles. Physiol. Rev. 2011, 91, 1447–1531. [Google Scholar] [CrossRef]
- Bodine, S.C.; Stitt, T.N.; Gonzalez, M.; Kline, W.O.; Stover, G.L.; Bauerlein, R.; Zlotchenko, E.; Scrimgeour, A.; Lawrence, J.C.; Glass, D.J.; et al. Akt/mTOR Pathway Is a Crucial Regulator of Skeletal Muscle Hypertrophy and Can Prevent Muscle Atrophy in Vivo. Nat. Cell Biol. 2001, 3, 1014–1019. [Google Scholar] [CrossRef]
- Aguilar-Agon, K.W.; Capel, A.J.; Martin, N.R.W.; Player, D.J.; Lewis, M.P. Mechanical Loading Stimulates Hypertrophy in Tissue-engineered Skeletal Muscle: Molecular and Phenotypic Responses. J. Cell Physiol. 2019, 234, 23547–23558. [Google Scholar] [CrossRef] [PubMed]
- Schiaffino, S.; Dyar, K.A.; Ciciliot, S.; Blaauw, B.; Sandri, M. Mechanisms Regulating Skeletal Muscle Growth and Atrophy. FEBS J. 2013, 280, 4294–4314. [Google Scholar] [CrossRef] [PubMed]
- Chargé, S.B.P.; Rudnicki, M.A. Cellular and Molecular Regulation of Muscle Regeneration. Physiol. Rev. 2004, 84, 209–238. [Google Scholar] [CrossRef]
- Hudlicka, O.; Brown, M.; Egginton, S. Angiogenesis in Skeletal and Cardiac Muscle. Physiol. Rev. 1992, 72, 369–417. [Google Scholar] [CrossRef]
- Yaffe, D.; Saxel, O. Serial Passaging and Differentiation of Myogenic Cells Isolated from Dystrophic Mouse Muscle. Nature 1977, 270, 725–727. [Google Scholar] [CrossRef]
- Smith, A.S.; Luttrell, S.M.; Dupont, J.-B.; Gray, K.; Lih, D.; Fleming, J.W.; Cunningham, N.J.; Jepson, S.; Hesson, J.; Mathieu, J.; et al. High-Throughput, Real-Time Monitoring of Engineered Skeletal Muscle Function Using Magnetic Sensing. J. Tissue Eng. 2022, 13, 20417314221122127. [Google Scholar] [CrossRef]
- Nicolas, R.; Bonnin, M.-A.; Blavet, C.; de Lima, J.E.; Legallais, C.; Duprez, D. 3D-Environment and Muscle Contraction Regulate the Heterogeneity of Myonuclei. Skelet. Muscle 2024, 14, 27. [Google Scholar] [CrossRef]
- Afshar Bakooshli, M.; Lippmann, E.S.; Mulcahy, B.; Iyer, N.; Nguyen, C.T.; Tung, K.; Stewart, B.A.; van den Dorpel, H.; Fuehrmann, T.; Shoichet, M.; et al. A 3D Culture Model of Innervated Human Skeletal Muscle Enables Studies of the Adult Neuromuscular Junction. eLife 2019, 8, e44530. [Google Scholar] [CrossRef]
- Bersini, S.; Francescato, R.; Moretti, M. Biofabrication of 3D Human Muscle Model with Vascularization and Endomysium. Methods Mol. Biol. 2022, 2373, 213–230. [Google Scholar] [CrossRef]
- Dennis, R.G.; Kosnik, P.E.; Gilbert, M.E.; Faulkner, J.A. Excitability and Contractility of Skeletal Muscle Engineered from Primary Cultures and Cell Lines. Am. J. Physiol. Cell Physiol. 2001, 280, C288–C295. [Google Scholar] [CrossRef]
- Juhas, M.; Engelmayr, G.C.; Fontanella, A.N.; Palmer, G.M.; Bursac, N. Biomimetic Engineered Muscle with Capacity for Vascular Integration and Functional Maturation in Vivo. Proc. Natl. Acad. Sci. USA 2014, 111, 5508–5513. [Google Scholar] [CrossRef] [PubMed]
- Rao, L.; Qian, Y.; Khodabukus, A.; Ribar, T.; Bursac, N. Engineering Human Pluripotent Stem Cells into a Functional Skeletal Muscle Tissue. Nat. Commun. 2018, 9, 126. [Google Scholar] [CrossRef] [PubMed]
- Blau, H.M.; Pavlath, G.K.; Hardeman, E.C.; Chiu, C.P.; Silberstein, L.; Webster, S.G.; Miller, S.C.; Webster, C. Plasticity of the Differentiated State. Science 1985, 230, 758–766. [Google Scholar] [CrossRef]
- Richler, C.; Yaffe, D. The in Vitro Cultivation and Differentiation Capacities of Myogenic Cell Lines. Dev. Biol. 1970, 23, 1–22. [Google Scholar] [CrossRef]
- McMahon, D.K.; Anderson, P.A.; Nassar, R.; Bunting, J.B.; Saba, Z.; Oakeley, A.E.; Malouf, N.N. C2C12 Cells: Biophysical, Biochemical, and Immunocytochemical Properties. Am. J. Physiol. 1994, 266, C1795–C1802. [Google Scholar] [CrossRef]
- Abdelmoez, A.M.; Sardón Puig, L.; Smith, J.A.B.; Gabriel, B.M.; Savikj, M.; Dollet, L.; Chibalin, A.V.; Krook, A.; Zierath, J.R.; Pillon, N.J. Comparative Profiling of Skeletal Muscle Models Reveals Heterogeneity of Transcriptome and Metabolism. Am. J. Physiol. Cell Physiol. 2020, 318, C615–C626. [Google Scholar] [CrossRef]
- Rossi, S.; Poliani, P.L.; Cominelli, M.; Bozzato, A.; Vescovi, R.; Monti, E.; Fanzani, A. Caveolin 1 Is a Marker of Poor Differentiation in Rhabdomyosarcoma. Eur. J. Cancer 2011, 47, 761–772. [Google Scholar] [CrossRef]
- Wang, S.; Guo, L.; Dong, L.; Guo, L.; Li, S.; Zhang, J.; Sun, M. TGF-β1 Signal Pathway May Contribute to Rhabdomyosarcoma Development by Inhibiting Differentiation. Cancer Sci. 2010, 101, 1108–1116. [Google Scholar] [CrossRef]
- Keller, C.; Guttridge, D.C. Mechanisms of Impaired Differentiation in Rhabdomyosarcoma. FEBS J. 2013, 280, 4323–4334. [Google Scholar] [CrossRef]
- Zhu, C.-H.; Mouly, V.; Cooper, R.N.; Mamchaoui, K.; Bigot, A.; Shay, J.W.; Di Santo, J.P.; Butler-Browne, G.S.; Wright, W.E. Cellular Senescence in Human Myoblasts Is Overcome by Human Telomerase Reverse Transcriptase and Cyclin-Dependent Kinase 4: Consequences in Aging Muscle and Therapeutic Strategies for Muscular Dystrophies. Aging Cell 2007, 6, 515–523. [Google Scholar] [CrossRef]
- Mamchaoui, K.; Trollet, C.; Bigot, A.; Negroni, E.; Chaouch, S.; Wolff, A.; Kandalla, P.K.; Marie, S.; Di Santo, J.; St Guily, J.L.; et al. Immortalized Pathological Human Myoblasts: Towards a Universal Tool for the Study of Neuromuscular Disorders. Skelet. Muscle 2011, 1, 34. [Google Scholar] [CrossRef] [PubMed]
- Blau, H.M.; Webster, C. Isolation and Characterization of Human Muscle Cells. Proc. Natl. Acad. Sci. USA 1981, 78, 5623–5627. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Wilschut, K.J.; Kouklis, G.; Tian, H.; Hesse, R.; Garland, C.; Sbitany, H.; Hansen, S.; Seth, R.; Knott, P.D.; et al. Human Satellite Cell Transplantation and Regeneration from Diverse Skeletal Muscles. Stem Cell Rep. 2015, 5, 419–434. [Google Scholar] [CrossRef]
- Bentzinger, C.F.; Wang, Y.X.; Dumont, N.A.; Rudnicki, M.A. Cellular Dynamics in the Muscle Satellite Cell Niche. EMBO Rep. 2013, 14, 1062–1072. [Google Scholar] [CrossRef] [PubMed]
- Uchimura, T.; Otomo, J.; Sato, M.; Sakurai, H. A Human iPS Cell Myogenic Differentiation System Permitting High-Throughput Drug Screening. Stem Cell Res. 2017, 25, 98–106. [Google Scholar] [CrossRef]
- Chal, J.; Al Tanoury, Z.; Hestin, M.; Gobert, B.; Aivio, S.; Hick, A.; Cherrier, T.; Nesmith, A.P.; Parker, K.K.; Pourquié, O. Generation of Human Muscle Fibers and Satellite-like Cells from Human Pluripotent Stem Cells in Vitro. Nat. Protoc. 2016, 11, 1833–1850. [Google Scholar] [CrossRef]
- Hinds, S.; Bian, W.; Dennis, R.G.; Bursac, N. The Role of Extracellular Matrix Composition in Structure and Function of Bioengineered Skeletal Muscle. Biomaterials 2011, 32, 3575–3583. [Google Scholar] [CrossRef]
- Dessauge, F.; Schleder, C.; Perruchot, M.-H.; Rouger, K. 3D in Vitro Models of Skeletal Muscle: Myopshere, Myobundle and Bioprinted Muscle Construct. Vet. Res. 2021, 52, 72. [Google Scholar] [CrossRef]
- Vandenburgh, H.; Del Tatto, M.; Shansky, J.; Lemaire, J.; Chang, A.; Payumo, F.; Lee, P.; Goodyear, A.; Raven, L. Tissue-Engineered Skeletal Muscle Organoids for Reversible Gene Therapy. Hum. Gene Ther. 1996, 7, 2195–2200. [Google Scholar] [CrossRef]
- Fleming, J.W.; McCloskey, M.C.; Gray, K.; Nash, D.R.; Leung, V.; Michas, C.; Luttrell, S.M.; Cavanaugh, C.; Mathieu, J.; Mcquire, S.; et al. An Automated Platform for Simultaneous, Longitudinal Analysis of Engineered Neuromuscular Tissues for Applications in Neurotoxin Potency Testing. Curr. Res. Toxicol. 2025, 8, 100218. [Google Scholar] [CrossRef]
- Osaki, T.; Uzel, S.G.M.; Kamm, R.D. Microphysiological 3D Model of Amyotrophic Lateral Sclerosis (ALS) from Human iPS-Derived Muscle Cells and Optogenetic Motor Neurons. Sci. Adv. 2018, 4, eaat5847. [Google Scholar] [CrossRef] [PubMed]
- Murugan, R.; Ramakrishna, S. Design Strategies of Tissue Engineering Scaffolds with Controlled Fiber Orientation. Tissue Eng. 2007, 13, 1845–1866. [Google Scholar] [CrossRef]
- Aviss, K.J.; Gough, J.E.; Downes, S. Aligned Electrospun Polymer Fibres for Skeletal Muscle Regeneration. Eur. Cell Mater. 2010, 19, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Huber, A.; Pickett, A.; Shakesheff, K.M. Reconstruction of Spatially Orientated Myotubes in Vitro Using Electrospun, Parallel Microfibre Arrays. Eur. Cell Mater. 2007, 14, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Mulbauer, G.D.; Matthew, H.W.T. Biomimetic Scaffolds in Skeletal Muscle Regeneration. Discoveries 2019, 7, e90. [Google Scholar] [CrossRef]
- Huang, N.F.; Patel, S.; Thakar, R.G.; Wu, J.; Hsiao, B.S.; Chu, B.; Lee, R.J.; Li, S. Myotube Assembly on Nanofibrous and Micropatterned Polymers. Nano Lett. 2006, 6, 537–542. [Google Scholar] [CrossRef]
- Ku, S.H.; Lee, S.H.; Park, C.B. Synergic Effects of Nanofiber Alignment and Electroactivity on Myoblast Differentiation. Biomaterials 2012, 33, 6098–6104. [Google Scholar] [CrossRef]
- Pham, Q.P.; Sharma, U.; Mikos, A.G. Electrospinning of Polymeric Nanofibers for Tissue Engineering Applications: A Review. Tissue Eng. 2006, 12, 1197–1211. [Google Scholar] [CrossRef]
- Huang, C.; Thomas, N.L. Fabrication of Porous Fibers via Electrospinning: Strategies and Applications. Polym. Rev. 2020, 60, 595–647. [Google Scholar] [CrossRef]
- Jana, S.; Zhang, M. Fabrication of 3D Aligned Nanofibrous Tubes by Direct Electrospinning. J. Mater. Chem. B 2013, 1, 2575–2581. [Google Scholar] [CrossRef]
- Gao, L.; Li, L.; Wu, W.; Feng, J.; Liu, Z.; He, J.; Li, D. 3D-Bioprinted in Vitro Skeletal Muscle with Pennate Fiber Architecture to Enhance Contractile Function. Int. J. Bioprinting 2024, 10, 4371. [Google Scholar] [CrossRef]
- Bloise, N.; Berardi, E.; Gualandi, C.; Zaghi, E.; Gigli, M.; Duelen, R.; Ceccarelli, G.; Cortesi, E.E.; Costamagna, D.; Bruni, G.; et al. Ether-Oxygen Containing Electrospun Microfibrous and Sub-Microfibrous Scaffolds Based on Poly(butylene 1,4-cyclohexanedicarboxylate) for Skeletal Muscle Tissue Engineering. Int. J. Mol. Sci. 2018, 19, 3212. [Google Scholar] [CrossRef] [PubMed]
- Namjoo, A.R.; Abrbekoh, F.N.; Saghati, S.; Amini, H.; Saadatlou, M.A.E.; Rahbarghazi, R. Tissue Engineering Modalities in Skeletal Muscles: Focus on Angiogenesis and Immunomodulation Properties. Stem Cell Res. Ther. 2023, 14, 90. [Google Scholar] [CrossRef] [PubMed]
- Drury, J.L.; Mooney, D.J. Hydrogels for Tissue Engineering: Scaffold Design Variables and Applications. Biomaterials 2003, 24, 4337–4351. [Google Scholar] [CrossRef] [PubMed]
- Calero-Castro, F.J.; Perez-Puyana, V.M.; Laga, I.; Padillo Ruiz, J.; Romero, A.; De La Portilla De Juan, F. Mechanical Stimulation and Aligned Poly(ε-Caprolactone)–Gelatin Electrospun Scaffolds Promote Skeletal Muscle Regeneration. ACS Appl. Biol. Mater. 2024, 7, 6430–6440. [Google Scholar] [CrossRef]
- Huang, Y.-C.; Dennis, R.G.; Larkin, L.; Baar, K. Rapid Formation of Functional Muscle in Vitro Using Fibrin Gels. J. Appl. Physiol. 2005, 98, 706–713. [Google Scholar] [CrossRef]
- Marcinczyk, M.; Elmashhady, H.; Talovic, M.; Dunn, A.; Bugis, F.; Garg, K. Laminin-111 Enriched Fibrin Hydrogels for Skeletal Muscle Regeneration. Biomaterials 2017, 141, 233–242. [Google Scholar] [CrossRef]
- Gholobova, D.; Decroix, L.; Van Muylder, V.; Desender, L.; Gerard, M.; Carpentier, G.; Vandenburgh, H.; Thorrez, L. Endothelial Network Formation Within Human Tissue-Engineered Skeletal Muscle. Tissue Eng. Part A 2015, 21, 2548–2558. [Google Scholar] [CrossRef]
- Beier, J.P.; Klumpp, D.; Rudisile, M.; Dersch, R.; Wendorff, J.H.; Bleiziffer, O.; Arkudas, A.; Polykandriotis, E.; Horch, R.E.; Kneser, U. Collagen matrices from sponge to nano: New perspectives for tissue engineering of skeletal muscle. BMC Biotechnol. 2009, 9, 34. [Google Scholar] [CrossRef]
- Urciuolo, A.; De Coppi, P. Decellularized Tissue for Muscle Regeneration. Int. J. Mol. Sci. 2018, 19, 2392. [Google Scholar] [CrossRef]
- Ross, J.J.; Tranquillo, R.T. ECM Gene Expression Correlates with in Vitro Tissue Growth and Development in Fibrin Gel Remodeled by Neonatal Smooth Muscle Cells. Matrix Biol. 2003, 22, 477–490. [Google Scholar] [CrossRef] [PubMed]
- Antoine, E.E.; Vlachos, P.P.; Rylander, M.N. Review of Collagen I Hydrogels for Bioengineered Tissue Microenvironments: Characterization of Mechanics, Structure, and Transport. Tissue Eng. Part B Rev. 2014, 20, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Abraham, L.C.; Zuena, E.; Perez-Ramirez, B.; Kaplan, D.L. Guide to Collagen Characterization for Biomaterial Studies. J. Biomed. Mater. Res. Part B Appl. Biomater. 2008, 87B, 264–285. [Google Scholar] [CrossRef]
- Sarrafian, T.L.; Bodine, S.C.; Murphy, B.; Grayson, J.K.; Stover, S.M. Extracellular Matrix Scaffolds for Treatment of Large Volume Muscle Injuries: A Review. Vet. Surg. 2018, 47, 524–535. [Google Scholar] [CrossRef]
- Perniconi, B.; Costa, A.; Aulino, P.; Teodori, L.; Adamo, S.; Coletti, D. The Pro-Myogenic Environment Provided by Whole Organ Scale Acellular Scaffolds from Skeletal Muscle. Biomaterials 2011, 32, 7870–7882. [Google Scholar] [CrossRef]
- Mase, V.J.; Hsu, J.R.; Wolf, S.E.; Wenke, J.C.; Baer, D.G.; Owens, J.; Badylak, S.F.; Walters, T.J. Clinical Application of an Acellular Biologic Scaffold for Surgical Repair of a Large, Traumatic Quadriceps Femoris Muscle Defect. Orthopedics 2010, 33, 7. [Google Scholar] [CrossRef]
- Sicari, B.M.; Agrawal, V.; Siu, B.F.; Medberry, C.J.; Dearth, C.L.; Turner, N.J.; Badylak, S.F. A Murine Model of Volumetric Muscle Loss and a Regenerative Medicine Approach for Tissue Replacement. Tissue Eng. Part A 2012, 18, 1941–1948. [Google Scholar] [CrossRef]
- Dziki, J.L.; Huleihel, L.; Scarritt, M.E.; Badylak, S.F. Extracellular Matrix Bioscaffolds as Immunomodulatory Biomaterials. Tissue Eng. Part A 2017, 23, 1152–1159. [Google Scholar] [CrossRef]
- Sicari, B.M.; Rubin, J.P.; Dearth, C.L.; Wolf, M.T.; Ambrosio, F.; Boninger, M.; Turner, N.J.; Weber, D.J.; Simpson, T.W.; Wyse, A.; et al. An Acellular Biologic Scaffold Promotes Skeletal Muscle Formation in Mice and Humans with Volumetric Muscle Loss. Sci. Transl. Med. 2014, 6, 234ra58. [Google Scholar] [CrossRef]
- Dziki, J.; Badylak, S.; Yabroudi, M.; Sicari, B.; Ambrosio, F.; Stearns, K.; Turner, N.; Wyse, A.; Boninger, M.L.; Brown, E.H.P.; et al. An Acellular Biologic Scaffold Treatment for Volumetric Muscle Loss: Results of a 13-Patient Cohort Study. NPJ Regen. Med. 2016, 1, 16008. [Google Scholar] [CrossRef]
- Kozan, N.G.; Joshi, M.; Sicherer, S.T.; Grasman, J.M. Porous Biomaterial Scaffolds for Skeletal Muscle Tissue Engineering. Front. Bioeng. Biotechnol. 2023, 11, 1245897. [Google Scholar] [CrossRef] [PubMed]
- Fuoco, C.; Rizzi, R.; Biondo, A.; Longa, E.; Mascaro, A.; Shapira-Schweitzer, K.; Kossovar, O.; Benedetti, S.; Salvatori, M.L.; Santoleri, S.; et al. In Vivo Generation of a Mature and Functional Artificial Skeletal Muscle. EMBO Mol. Med. 2015, 7, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Engler, A.J.; Griffin, M.A.; Sen, S.; Bönnemann, C.G.; Sweeney, H.L.; Discher, D.E. Myotubes Differentiate Optimally on Substrates with Tissue-like Stiffness: Pathological Implications for Soft or Stiff Microenvironments. J. Cell Biol. 2004, 166, 877–887. [Google Scholar] [CrossRef]
- Maji, S.; Lee, H. Engineering Hydrogels for the Development of Three-Dimensional In Vitro Models. Int. J. Mol. Sci. 2022, 23, 2662. [Google Scholar] [CrossRef]
- Kiran, S.; Dwivedi, P.; Kumar, V.; Price, R.L.; Singh, U.P. Immunomodulation and Biomaterials: Key Players to Repair Volumetric Muscle Loss. Cells 2021, 10, 2016. [Google Scholar] [CrossRef]
- Rodriguez Ayala, A.; Christ, G.; Griffin, D. Cell-Scale Porosity Minimizes Foreign Body Reaction and Promotes Innervated Myofiber Formation after Volumetric Muscle Loss. NPJ Regen. Med. 2025, 10, 12. [Google Scholar] [CrossRef]
- Kim, J.H.; Seol, Y.-J.; Ko, I.K.; Kang, H.-W.; Lee, Y.K.; Yoo, J.J.; Atala, A.; Lee, S.J. 3D Bioprinted Human Skeletal Muscle Constructs for Muscle Function Restoration. Sci. Rep. 2018, 8, 12307. [Google Scholar] [CrossRef]
- Jain, R.K.; Au, P.; Tam, J.; Duda, D.G.; Fukumura, D. Engineering Vascularized Tissue. Nat. Biotechnol. 2005, 23, 821–823. [Google Scholar] [CrossRef]
- Malda, J.; Visser, J.; Melchels, F.P.; Jüngst, T.; Hennink, W.E.; Dhert, W.J.A.; Groll, J.; Hutmacher, D.W. 25th Anniversary Article: Engineering Hydrogels for Biofabrication. Adv. Mater. 2013, 25, 5011–5028. [Google Scholar] [CrossRef]
- Chromiak, J.A.; Shansky, J.; Perrone, C.; Vandenburgh, H.H. Bioreactor Perfusion System for the Long-Term Maintenance of Tissue-Engineered Skeletal Muscle Organoids. In Vitro Cell Dev. Biol. Anim. 1998, 34, 694–703. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Ito, A.; Jitsunobu, H.; Yamaguchi, K.; Kawabe, Y.; Mizumoto, H.; Kamihira, M. Hollow Fiber Bioreactor Perfusion Culture System for Magnetic Force-Based Skeletal Muscle Tissue Engineering. J. Chem. Eng. Jpn. 2012, 45, 348–354. [Google Scholar] [CrossRef]
- Agrawal, G.; Aung, A.; Varghese, S. Skeletal Muscle-on-a-Chip: An in Vitro Model to Evaluate Tissue Formation and Injury. Lab. Chip 2017, 17, 3447–3461. [Google Scholar] [CrossRef] [PubMed]
- Davis, L.A.; Fogarty, M.J.; Brown, A.; Sieck, G.C. Structure and Function of the Mammalian Neuromuscular Junction. In Comprehensive Physiology; John Wiley & Sons: Hoboken, NJ, USA, 2022; pp. 3731–3766. ISBN 978-0-470-65071-4. [Google Scholar]
- Massih, B.; Veh, A.; Schenke, M.; Mungwa, S.; Seeger, B.; Selvaraj, B.T.; Chandran, S.; Reinhardt, P.; Sterneckert, J.; Hermann, A.; et al. A 3D Cell Culture System for Bioengineering Human Neuromuscular Junctions to Model ALS. Front. Cell Dev. Biol. 2023, 11, 996952. [Google Scholar] [CrossRef] [PubMed]
- Buckingham, M.; Rigby, P.W.J. Gene Regulatory Networks and Transcriptional Mechanisms That Control Myogenesis. Dev. Cell 2014, 28, 225–238. [Google Scholar] [CrossRef] [PubMed]
- Tidball, J.G. Regulation of Muscle Growth and Regeneration by the Immune System. Nat. Rev. Immunol. 2017, 17, 165–178. [Google Scholar] [CrossRef]
- Rhee, S. Fibroblasts in Three Dimensional Matrices: Cell Migration and Matrix Remodeling. Exp. Mol. Med. 2009, 41, 858–865. [Google Scholar] [CrossRef]
- Mofarrahi, M.; McClung, J.M.; Kontos, C.D.; Davis, E.C.; Tappuni, B.; Moroz, N.; Pickett, A.E.; Huck, L.; Harel, S.; Danialou, G.; et al. Angiopoietin-1 Enhances Skeletal Muscle Regeneration in Mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 308, R576–R589. [Google Scholar] [CrossRef]
- Verma, M.; Asakura, Y.; Wang, X.; Zhou, K.; Ünverdi, M.; Kann, A.P.; Krauss, R.S.; Asakura, A. Endothelial Cell Signature in Muscle Stem Cells Validated by VEGFA-FLT1-AKT1 Axis Promoting Survival of Muscle Stem Cell. eLife 2024, 13, e73592. [Google Scholar] [CrossRef]
- Koffler, J.; Kaufman-Francis, K.; Shandalov, Y.; Egozi, D.; Amiad Pavlov, D.; Landesberg, A.; Levenberg, S. Improved Vascular Organization Enhances Functional Integration of Engineered Skeletal Muscle Grafts. Proc. Natl. Acad. Sci. USA 2011, 108, 14789–14794. [Google Scholar] [CrossRef]
- Sassoli, C.; Zecchi-Orlandini, S.; Formigli, L. Trophic Actions of Bone Marrow-Derived Mesenchymal Stromal Cells for Muscle Repair/Regeneration. Cells 2012, 1, 832–850. [Google Scholar] [CrossRef]
- Klimczak, A.; Kozlowska, U.; Kurpisz, M. Muscle Stem/Progenitor Cells and Mesenchymal Stem Cells of Bone Marrow Origin for Skeletal Muscle Regeneration in Muscular Dystrophies. Arch. Immunol. Ther. Exp. 2018, 66, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Shahin-Shamsabadi, A.; Selvaganapathy, P.R. A 3D Self-Assembled In Vitro Model to Simulate Direct and Indirect Interactions between Adipocytes and Skeletal Muscle Cells. Adv. Biosyst. 2020, 4, e2000034. [Google Scholar] [CrossRef]
- Bushby, K.; Finkel, R.; Birnkrant, D.J.; Case, L.E.; Clemens, P.R.; Cripe, L.; Kaul, A.; Kinnett, K.; McDonald, C.; Pandya, S.; et al. Diagnosis and Management of Duchenne Muscular Dystrophy, Part 1: Diagnosis, and Pharmacological and Psychosocial Management. Lancet Neurol. 2010, 9, 77–93. [Google Scholar] [CrossRef]
- Petrof, B.J.; Shrager, J.B.; Stedman, H.H.; Kelly, A.M.; Sweeney, H.L. Dystrophin Protects the Sarcolemma from Stresses Developed during Muscle Contraction. Proc. Natl. Acad. Sci. USA 1993, 90, 3710–3714. [Google Scholar] [CrossRef]
- Tejedera-Villafranca, A.; Montolio, M.; Ramón-Azcón, J.; Fernández-Costa, J.M. Mimicking Sarcolemmal Damagein Vitro: A Contractile 3D Model of Skeletal Muscle for Drug Testing in Duchenne Muscular Dystrophy. Biofabrication 2023, 15, 4. [Google Scholar] [CrossRef]
- Tinsley, J.M.; Potter, A.C.; Phelps, S.R.; Fisher, R.; Trickett, J.I.; Davies, K.E. Amelioration of the Dystrophic Phenotype of Mdx Mice Using a Truncated Utrophin Transgene. Nature 1996, 384, 349–353. [Google Scholar] [CrossRef]
- Ebrahimi, M.; Lad, H.; Fusto, A.; Tiper, Y.; Datye, A.; Nguyen, C.T.; Jacques, E.; Moyle, L.A.; Nguyen, T.; Musgrave, B.; et al. De Novo Revertant Fiber Formation and Therapy Testing in a 3D Culture Model of Duchenne Muscular Dystrophy Skeletal Muscle. Acta Biomater. 2021, 132, 227–244. [Google Scholar] [CrossRef]
- Arechavala-Gomeza, V.; Kinali, M.; Feng, L.; Guglieri, M.; Edge, G.; Main, M.; Hunt, D.; Lehovsky, J.; Straub, V.; Bushby, K.; et al. Revertant Fibres and Dystrophin Traces in Duchenne Muscular Dystrophy: Implication for Clinical Trials. Neuromuscul. Disord. 2010, 20, 295–301. [Google Scholar] [CrossRef]
- In ’t Groen, S.L.M.; Franken, M.; Bock, T.; Krüger, M.; de Greef, J.C.; Pijnappel, W.W.M.P. A Knock down Strategy for Rapid, Generic, and Versatile Modelling of Muscular Dystrophies in 3D-Tissue-Engineered-Skeletal Muscle. Skelet. Muscle 2024, 14, 3. [Google Scholar] [CrossRef]
- Bouchard, C.; Tremblay, J.P. Limb–Girdle Muscular Dystrophies Classification and Therapies. J. Clin. Med. 2023, 12, 4769. [Google Scholar] [CrossRef]
- Khodabukus, A.; Prabhu, N.K.; Roberts, T.; Buldo, M.; Detwiler, A.; Fralish, Z.D.; Kondash, M.E.; Truskey, G.A.; Koves, T.R.; Bursac, N. Bioengineered Model of Human LGMD2B Skeletal Muscle Reveals Roles of Intracellular Calcium Overload in Contractile and Metabolic Dysfunction in Dysferlinopathy. Adv. Sci. 2024, 11, e2400188. [Google Scholar] [CrossRef] [PubMed]
- Hartman, J.; Patki, T.; Johnson, N.E. Diagnosis and Management of Myotonic Dystrophy Type 1. JAMA 2024, 331, 1227–1228. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Garibay, X.; Ortega, M.A.; Cerro-Herreros, E.; Comelles, J.; Martínez, E.; Artero, R.; Fernández-Costa, J.M.; Ramón-Azcón, J. Bioengineeredin vitro3D Model of Myotonic Dystrophy Type 1 Human Skeletal Muscle. Biofabrication 2021, 13, 3. [Google Scholar] [CrossRef] [PubMed]
- Cerro-Herreros, E.; Sabater-Arcis, M.; Fernandez-Costa, J.M.; Moreno, N.; Perez-Alonso, M.; Llamusi, B.; Artero, R. miR-23b and miR-218 Silencing Increase Muscleblind-like Expression and Alleviate Myotonic Dystrophy Phenotypes in Mammalian Models. Nat. Commun. 2018, 9, 2482. [Google Scholar] [CrossRef]
- Lemmers, R.J.L.F.; van der Vliet, P.J.; Klooster, R.; Sacconi, S.; Camaño, P.; Dauwerse, J.G.; Snider, L.; Straasheijm, K.R.; van Ommen, G.J.; Padberg, G.W.; et al. A Unifying Genetic Model for Facioscapulohumeral Muscular Dystrophy. Science 2010, 329, 1650–1653. [Google Scholar] [CrossRef]
- Franken, M.; van der Wal, E.; Zheng, D.; den Hamer, B.; van der Vliet, P.J.; Lemmers, R.J.L.F.; van den Heuvel, A.; Dorn, A.L.; Duivenvoorden, C.G.A.; in’t Groen, S.L.M.; et al. Three-Dimensional Tissue Engineered Skeletal Muscle Modelling Facioscapulohumeral Muscular Dystrophy. Brain 2024, 148, awae379. [Google Scholar] [CrossRef]
- Marion, R.W.; Paljevic, E. The Glycogen Storage Disorders. Pediatr. Rev. 2020, 41, 41–44. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, C.J.; Khodabukus, A.; Tran, S.; Han, S.-O.; Carlson, A.L.; Madden, L.; Kishnani, P.S.; Koeberl, D.D.; Bursac, N. Three-Dimensional Tissue-Engineered Human Skeletal Muscle Model of Pompe Disease. Commun. Biol. 2021, 4, 524. [Google Scholar] [CrossRef]
- Muchir, A.; Worman, H.J. Emery–Dreifuss Muscular Dystrophy: Focal Point Nuclear Envelope. Curr. Opin. Neurol. 2019, 32, 728–734. [Google Scholar] [CrossRef]
- Maffioletti, S.M.; Sarcar, S.; Henderson, A.B.H.; Mannhardt, I.; Pinton, L.; Moyle, L.A.; Steele-Stallard, H.; Cappellari, O.; Wells, K.E.; Ferrari, G.; et al. Three-Dimensional Human iPSC-Derived Artificial Skeletal Muscles Model Muscular Dystrophies and Enable Multilineage Tissue Engineering. Cell Rep. 2018, 23, 899–908. [Google Scholar] [CrossRef]
- Fernández-Costa, J.M.; Fernández-Garibay, X.; Velasco-Mallorquí, F.; Ramón-Azcón, J. Bioengineered in Vitro Skeletal Muscles as New Tools for Muscular Dystrophies Preclinical Studies. J. Tissue Eng. 2021, 12, 2041731420981339. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Yang, Z.; Ying, H.; Zhao, J.; Wang, X.; Gong, J.; Li, L.; Liu, X.; Gong, T.; Ke, Q.; et al. iPSC-Derived Cardiomyocytes and Engineered Heart Tissues Reveal Suppressed JAK2/STAT3 Signaling in LMNA-Related Emery-Dreifuss Muscular Dystrophy. Redox Biol. 2025, 83, 103638. [Google Scholar] [CrossRef] [PubMed]
- Giza, S.; Mojica-Santiago, J.A.; Parafati, M.; Malany, L.K.; Platt, D.; Schmidt, C.E.; Coen, P.M.; Malany, S. Microphysiological System for Studying Contractile Differences in Young, Active, and Old, Sedentary Adult Derived Skeletal Muscle Cells. Aging Cell 2022, 21, e13650. [Google Scholar] [CrossRef]
- Wang, K.; Smith, S.H.; Iijima, H.; Hettinger, Z.R.; Mallepally, A.; Shroff, S.G.; Ambrosio, F. Bioengineered 3D Skeletal Muscle Model Reveals Complement 4b as a Cell-Autonomous Mechanism of Impaired Regeneration with Aging. Adv. Mater. 2023, 35, 2207443. [Google Scholar] [CrossRef]
- Chen, Z.; Li, B.; Zhan, R.-Z.; Rao, L.; Bursac, N. Exercise Mimetics and JAK Inhibition Attenuate IFN-γ-Induced Wasting in Engineered Human Skeletal Muscle. Sci. Adv. 2021, 7, eabd9502. [Google Scholar] [CrossRef]
- Acosta, F.M.; Howland, K.K.; Stojkova, K.; Hernandez, E.; Brey, E.M.; Rathbone, C.R. Adipogenic Differentiation Alters Properties of Vascularized Tissue-Engineered Skeletal Muscle. Tissue Eng. Part A 2022, 28, 54–68. [Google Scholar] [CrossRef]
- Na, Y.-J.; Choi, K.J.; Jung, W.H.; Park, S.B.; Koh, B.; Hoe, K.-L.; Kim, K.Y. Development of 3D Muscle Cell Culture-Based Screening System for Metabolic Syndrome Drug Research. Tissue Eng. Part C Methods 2025, 31, 2. [Google Scholar] [CrossRef]
- Turner, M.C.; Rimington, R.P.; Martin, N.R.W.; Fleming, J.W.; Capel, A.J.; Hodson, L.; Lewis, M.P. Physiological and Pathophysiological Concentrations of Fatty Acids Induce Lipid Droplet Accumulation and Impair Functional Performance of Tissue Engineered Skeletal Muscle. J. Cell Physiol. 2021, 236, 7033–7044. [Google Scholar] [CrossRef]
- Franceschi, C.; Garagnani, P.; Parini, P.; Giuliani, C.; Santoro, A. Inflammaging: A New Immune–Metabolic Viewpoint for Age-Related Diseases. Nat. Rev. Endocrinol. 2018, 14, 576–590. [Google Scholar] [CrossRef]
- Dalle, S.; Rossmeislova, L.; Koppo, K. The Role of Inflammation in Age-Related Sarcopenia. Front. Physiol. 2017, 8, 1045. [Google Scholar] [CrossRef]
- Antuña, E.; Cachán-Vega, C.; Bermejo-Millo, J.C.; Potes, Y.; Caballero, B.; Vega-Naredo, I.; Coto-Montes, A.; Garcia-Gonzalez, C. Inflammaging: Implications in Sarcopenia. Int. J. Mol. Sci. 2022, 23, 15039. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, D.J.; Erskine, R.M.; Morse, C.I.; Winwood, K.; Onambélé-Pearson, G. The Impact of Obesity on Skeletal Muscle Strength and Structure through Adolescence to Old Age. Biogerontology 2016, 17, 467–483. [Google Scholar] [CrossRef] [PubMed]
- Khodabukus, A. Tissue-Engineered Skeletal Muscle Models to Study Muscle Function, Plasticity, and Disease. Front. Physiol. 2021, 12, 619710. [Google Scholar] [CrossRef]
- Zhuang, P.; An, J.; Chua, C.K.; Tan, L.P. Bioprinting of 3D in Vitro Skeletal Muscle Models: A Review. Mater. Des. 2020, 193, 108794. [Google Scholar] [CrossRef]
- Carraro, E.; Rossi, L.; Maghin, E.; Canton, M.; Piccoli, M. 3D in Vitro Models of Pathological Skeletal Muscle: Which Cells and Scaffolds to Elect? Front. Bioeng. Biotechnol. 2022, 10, 941623. [Google Scholar] [CrossRef]
- Madden, L.; Juhas, M.; Kraus, W.E.; Truskey, G.A.; Bursac, N. Bioengineered Human Myobundles Mimic Clinical Responses of Skeletal Muscle to Drugs. eLife 2015, 4, e04885. [Google Scholar] [CrossRef]
- Sabetkish, S.; Currie, P.; Meagher, L. Recent Trends in 3D Bioprinting Technology for Skeletal Muscle Regeneration. Acta Biomater. 2024, 181, 46–66. [Google Scholar] [CrossRef]
- Borisov, V.; Gili Sole, L.; Reid, G.; Milan, G.; Hutter, G.; Grapow, M.; Eckstein, F.S.; Isu, G.; Marsano, A. Upscaled Skeletal Muscle Engineered Tissue with In Vivo Vascularization and Innervation Potential. Bioengineering 2023, 10, 800. [Google Scholar] [CrossRef]
- Maschmeyer, I.; Lorenz, A.K.; Schimek, K.; Hasenberg, T.; Ramme, A.P.; Hübner, J.; Lindner, M.; Drewell, C.; Bauer, S.; Thomas, A.; et al. A Four-Organ-Chip for Interconnected Long-Term Co-Culture of Human Intestine, Liver, Skin and Kidney Equivalents. Lab. Chip 2015, 15, 2688–2699. [Google Scholar] [CrossRef]
- Skardal, A.; Murphy, S.V.; Devarasetty, M.; Mead, I.; Kang, H.-W.; Seol, Y.-J.; Shrike Zhang, Y.; Shin, S.-R.; Zhao, L.; Aleman, J.; et al. Multi-Tissue Interactions in an Integrated Three-Tissue Organ-on-a-Chip Platform. Sci. Rep. 2017, 7, 8837. [Google Scholar] [CrossRef]
- Vernetti, L.; Gough, A.; Baetz, N.; Blutt, S.; Broughman, J.R.; Brown, J.A.; Foulke-Abel, J.; Hasan, N.; In, J.; Kelly, E.; et al. Functional Coupling of Human Microphysiology Systems: Intestine, Liver, Kidney Proximal Tubule, Blood-Brain Barrier and Skeletal Muscle. Sci. Rep. 2017, 7, 42296. [Google Scholar] [CrossRef] [PubMed]
- Reggio, A.; Fuoco, C.; De Paolis, F.; Deodati, R.; Testa, S.; Celikkin, N.; Volpi, M.; Bernardini, S.; Fornetti, E.; Baldi, J.; et al. 3D Rotary Wet-Spinning (RoWS) Biofabrication Directly Affects Proteomic Signature and Myogenic Maturation in Muscle Pericyte–Derived Human Myo-Substitute. Aggregate 2025, 6, e727. [Google Scholar] [CrossRef]
| Disease Model | Cell Types | Approach of 3D Muscle | Recapitulated Disease Features | Therapeutic Interventions Tested | Limitations | References |
|---|---|---|---|---|---|---|
| Duchenne Muscular Dystrophy (DMD) | Immortalized human muscle precursor cells (from both healthy controls and DMD patients) | Fibrin–Matrigel composite hydrogel cast in PDMS molds with flexible posts | The 3D model mimics key DMD features: sarcolemmal damage and reduced tetanic contraction and less resistance to fatigue. | Tested utrophin up-regulators (Ezutromid, Halofuginone, and a novel SOMutrophin candidate); only SOMutrophin increased utrophin nearly three-fold and improved contractile forces | High inter-individual variability, incomplete recapitulation of all DMD phenotypes (especially sarcolemmal stability). | [96] |
| Immortalized human myoblasts (isolated from DMD patient biopsies and from healthy donors) | Fibrin-based hydrogel with Geltrex, cast in the MyoTACTIC micro-mold device | • Absence of dystrophin in most DMD fibers. • Rare “revertant” dystrophin-positive fibers in one DMD line • Altered calcium handling vs. healthy lines | β1-integrin activating antibody (TS2/16) • Improved sarcomere organization • Some protection from contraction-induced structural damage | 3D muscles were not fully mature, and culture time was relatively short. | [98] | |
| Human iPSC-derived myogenic progenitor cells | Fibrin + 20% Matrigel composite hydrogel cast in PDMS molds with flexible posts | • DMD knockdown was generated via shRNA-mediated knock down delivered via lentiviral transduction • Near-zero twitch force in DMD KD at day 9 | Co-express micro-dystrophin via lentiviral transduction. Partial rescue of contracile force in DMD KD 3D muscle. | Not complete knock down or restore of DMD; Lack of other resident cell types in native muscle. | [100] | |
| Limb-Girdle Muscular Dystrophies type 2A (LGMD2A) | Human iPSC-derived myogenic progenitor cells | Fibrin + 20% Matrigel composite hydrogel cast in PDMS molds with flexible posts | • Lentiviral shRNA knockdown targeting CAPN3 in hiPSC-derived MPCs before 3D tissue formation. • Severe reduction in contractile force, disrupted tissue architecture, and proteomic changes consistent with CAPN3 deficiency | N/A | Knockdown rather than complete KO; short culture period. | [100] |
| LGMD2B | hiPSC-derived myogenic progenitor cells | Fibrin + Matrigel hydrogel | • Severely reduced force generation (50%) Diminished Ca2+ transient amplitude • Deficient membrane repair; • Impaired mitochondrial function under fatty acid treatment and lipid droplet accumulation. | RyR Ca2+ channel inhibitor (dantrolene), novel glucocorticoid (vamorolone); Improved structure and contractile functions | Disease severity in iPSC model can differ from native tissues; does not model all late-stage or systemic features of LGMD2B. | [102] |
| Myotonic Dystrophy Type 1 (DM1) | Immortalized transdifferentiated human myoblast-like cells from DM1 fibroblasts | Gelatin methacryloyl + carboxymethyl cellulose methacrylate (GelMA-CMCMA) | • No direct contracile force • 3D micropatterning improves myogenic fusion. • Thinner myotubes in DM1 | Antisense oligonucleotide (antagomiR-23b) | No functional contractility readout | [102,103] |
| Facioscapulohumeral Dystrophy (FSMD) | hiPSC-derived myogenic progenitors from mosaic FSHD1 patients | Fibrin + Matrigel hydrogel in custom PDMS “T-bone” molds | FSHD3D muscle: upregulated DUX4 & target genes; Reduced absolute force with myofiber atrophy and smaller sarcomeres. | p38 inhibitor (pamapimod), CK1 inhibitor, & rebastinib tested daily; showed no function and fiber size improvement in 3D | No co-cultured immune or supporting cells; immature 3D muscle | [107] |
| Pompe disease | Primary myoblasts from infantile-onset Pompe (IOPD) patients + healthy donors | Fibrin + Matrigel hydrogel in custom PDMS molds | GAA-deficiency, lysosome enlargement, glycogen accumulation. | rhGAA enzyme or AAV-mediated GAA partially reduced glycogen; incomplete rescue of functional deficits. | Partial disease features, no additional cell types. | [109] |
| Sarcopenia/aging | Primary human myoblasts from young and aged donors | Collagen I–Matrigel hydrogel incorporated in a PDMS-based microfluidic chip | Lower fusion index (36% vs. 69% to young muscle); Impaired and non-sychronous contraction | N/A | lack of additional cell types to mimic full muscle complexity | [114] |
| Primary mouse myoblasts from young (3–5 mo) and aged (21–23 mo) mice | Fibrin/Matrigel hydrogel cast in a PDMS chamber | Recapitulated atrophy, reduced contractile function, and delayed regeneration | Supplementation with recombinant α-klotho, extracellular vesicles (EVs), and C4b inhibition via CFI; improved myotube regeneration and enhanced force. | Mouse cells; lacks multicellular complexity | [115] | |
| Primary human myoblasts from skeletal muscle biopsies | Bovine fibrinogen and Matrigel cast in PDMS molds | • Disease phenotype induced by chronic exposure of differentiated EMTs to IFN-γ (20 ng/mL for 7 days) • Key features: myofiber atrophy, reduced contractile strength, slower kinetics, impaired calcium transients, and disrupted sarcomere organization | Both electric stimulation (exercise) and JAK inhibitors (tofacitinib or baricitinib) prevented IFN-γ–induced muscle deficits; "exercise" partially restored force and structure while JAK inhibitors fully normalized contractile and calcium-handling properties | The chronic IFN-γ model may not capture the full complexity of in vivo inflammation; limited donor diversity | [116] | |
| Obesity related metabolic syndrome | C2C12 + 3T3-L1 adipocytes | Bovine collagen I | • Direct or indirect co-culture C2C12 based 3D engineered muscle with 3T3-L1 adipocytes • Recapitulated metabolic crosstalk (higher lipolysis, lipid accumulation, insulin resistance insights) | No rescue; tested Isoproterenol (β-adrenergic agonist) response | Non-human cell lines; No contractility assessment; lack vascular/immune cell types in 3D muscle | [93] |
| Primary rat myoblasts + Microvascular fragments (MVFs) from adipose tissue | Fibrin hydrogel (formed by mixing fibrinogen with thrombin) cast in PDMS molds | • Diabetic-like muscle pathology characterized by increased intramuscular adipose tissue (IMAT) • Co-culture showed disorganized myotube and vessel alignment, impaired mechanical integrity, and altered insulin responsiveness | N/A | Lack of active contractile (twitch/tetanus) measurements; in vitro adipogenic induction may not fully recapitulate human pathology; use of rat cells may limit direct translational relevance | [117] | |
| C2C12 | Composite hydrogel of 1% alginate and 1 mg/mL collagen type I | Insulin resistance induced by treating differentiated 3D myotubes with palmitate (PA; optimal at 0.1 mM) | Pharmacological treatment using known insulin-sensitizers (rosiglitazone and metformin) plus screening of 165 candidate compounds | Focuses solely on metabolic endpoints (glucose uptake) without contractile assessment; uses a murine cell line (C2C12) that may not fully recapitulate human skeletal muscle physiology | [118] | |
| C2C12 | 65% Type I rat tail collagen, 20% Matrigel, and 10% 10X MEM, cast into 3D printed inserts | • 3D constructs exposed to exogenous fatty acids (OPLA mixture: oleic, palmitic, linoleic, α-linoleic acids at 45:30:24:1% ratio) at 200 µM and 800 µM for 4 days • Exhibits lipid droplet accumulation, altered lipid metabolism gene expression, reduced myosin heavy chain coverage, and decreased force generation | N/A | Short-term FA exposure may not mimic chronic conditions; lacks additional cell types to mimic the full in vivo microenvironment | [119] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, T.; Warren, C.R. From 2D Myotube Cultures to 3D Engineered Skeletal Muscle Constructs: A Comprehensive Review of In Vitro Skeletal Muscle Models and Disease Modeling Applications. Cells 2025, 14, 882. https://doi.org/10.3390/cells14120882
Cao T, Warren CR. From 2D Myotube Cultures to 3D Engineered Skeletal Muscle Constructs: A Comprehensive Review of In Vitro Skeletal Muscle Models and Disease Modeling Applications. Cells. 2025; 14(12):882. https://doi.org/10.3390/cells14120882
Chicago/Turabian StyleCao, Tianxin, and Curtis R. Warren. 2025. "From 2D Myotube Cultures to 3D Engineered Skeletal Muscle Constructs: A Comprehensive Review of In Vitro Skeletal Muscle Models and Disease Modeling Applications" Cells 14, no. 12: 882. https://doi.org/10.3390/cells14120882
APA StyleCao, T., & Warren, C. R. (2025). From 2D Myotube Cultures to 3D Engineered Skeletal Muscle Constructs: A Comprehensive Review of In Vitro Skeletal Muscle Models and Disease Modeling Applications. Cells, 14(12), 882. https://doi.org/10.3390/cells14120882