

Exploration of SUMO2/3 Expression Levels and Autophagy Process in Fragile X-Associated Tremor/Ataxia Syndrome: Addressing Study Limitations and Insights for Future Research

 , , ,
, , ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Postmortem Human Brain Samples
2.2. Skin Fibroblast Cultures
2.3. SUMO2/3 Immunohistochemistry Examination
2.4. SUMO2/3 Protein Quantification
2.5. Functional Enrichment Analysis
2.6. Autophagic Flux Analysis
2.7. Immunocytochemistry for Autophagosome Evaluation
2.8. Statistical Analysis
3. Results
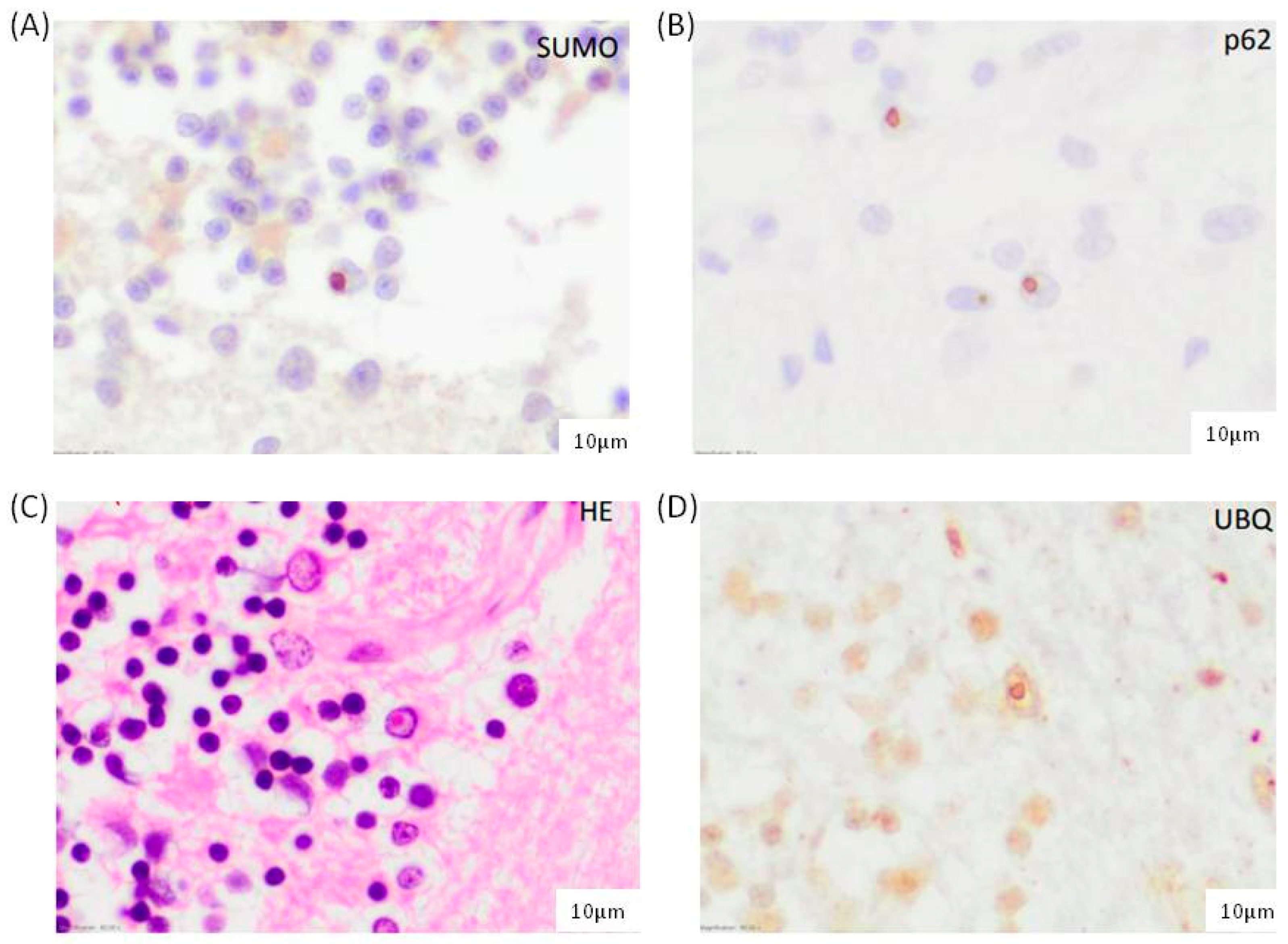
3.1. FXTAS Postmortem Brain Samples Are Positive for SUMO2/3 Conjugates
3.2. Western Blot Analysis of SUMO2/3 Expression in Brain and in Skin Fibroblasts from FXTAS Patients
3.3. SUMOylation Related Pathways Are Significantly Altered in Brain and in Skin Fibroblasts Samples from FXTAS Patients
3.4. Fibroblasts from FXTAS Patient’s Evidence Accumulation of p62 Protein Levels and Increased Autophagy Processes
3.5. Regulation of Autophagy Is Significantly Altered in Brain Samples and in Skin Fibroblasts from FXTAS Patients
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tassone, F.; Iong, K.P.; Tong, T.H.; Lo, J.; Gane, L.W.; Berry-Kravis, E.; Nguyen, D.; Mu, L.Y.; Laffin, J.; Bailey, D.B.; et al. FMR1 CGG allele size and prevalence ascertained through newborn screening in the United States. Genome Med. 2012, 4, 100. [Google Scholar] [CrossRef] [PubMed]
- Hagerman, P.J.; Hagerman, R.J. The fragile-X premutation: A maturing perspective. Am. J. Hum. Genet. 2004, 74, 805–816. [Google Scholar] [CrossRef] [PubMed]
- Jacquemont, S.; Hagerman, R.J.; Leehey, M.; Grigsby, J.; Zhang, L.; Brunberg, J.A.; Greco, C.; Des Portes, V.; Jardini, T.; Levine, R.; et al. Fragile X premutation tremor/ataxia syndrome: Molecular, clinical, and neuroimaging correlates. Am. J. Hum. Genet. 2003, 72, 869–878. [Google Scholar] [CrossRef] [PubMed]
- Hagerman, R.; Hagerman, P. Advances in clinical and molecular understanding of the FMR1 premutation and fragile X-associated tremor/ataxia syndrome. Lancet Neurol. 2013, 12, 786–798. [Google Scholar] [CrossRef] [PubMed]
- Apartis, E.; Blancher, A.; Meissner, W.G.; Guyant-Maréchal, L.; Maltête, D.; De Broucker, T.; Legrand, A.P.; Bouzenada, H.; Thanh, H.T.; Sallansonnet-Froment, M.; et al. FXTAS: New insights and the need for revised diagnostic criteria. Neurology 2012, 79, 1898–1907. [Google Scholar] [CrossRef]
- Rodriguez-Revenga, L.; Madrigal, I.; Pagonabarraga, J.; Xunclà, M.; Badenas, C.; Kulisevsky, J.; Gomez, B.; Milà, M. Penetrance of FMR1 premutation associated pathologies in fragile X syndrome families. Eur. J. Hum. Genet. 2009, 17, 1359–1362. [Google Scholar] [CrossRef]
- Greco, C.M.; Hagerman, R.J.; Tassone, F.; Chudley, A.E.; Del Bigio, M.R.; Jacquemont, S.; Leehey, M.; Hagerman, P.J. Neuronal intranuclear inclusions in a new cerebellar tremor/ataxia syndrome among fragile X carriers. Brain 2002, 125, 1760–1771. [Google Scholar] [CrossRef]
- Greco, C.M.; Berman, R.F.; Martin, R.M.; Tassone, F.; Schwartz, P.H.; Chang, A.; Trapp, B.D.; Iwahashi, C.; Brunberg, J.; Grigsby, J.; et al. Neuropathology of fragile X-associated tremor/ataxia syndrome (FXTAS). Brain 2006, 129, 243–255. [Google Scholar] [CrossRef]
- Tassone, F.; Iwahashi, C.; Hagerman, P.J. FMR1 RNA within the intranuclear inclusions of fragile X-associated tremor/ataxia syndrome (FXTAS). RNA Biol. 2004, 1, 103–105. [Google Scholar] [CrossRef]
- Iwahashi, C.K.; Yasui, D.H.; An, H.J.; Greco, C.M.; Tassone, F.; Nannen, K.; Babineau, B.; Lebrilla, C.B.; Hagerman, R.J.; Hagerman, P.J. Protein composition of the intranuclear inclusions of FXTAS. Brain 2006, 129, 256–271. [Google Scholar] [CrossRef]
- Ma, L.; Herren, A.W.; Espinal, G.; Randol, J.; McLaughlin, B.; Martinez-Cerdeño, V.; Pessah, I.N.; Hagerman, R.J.; Hagerman, P.J. Composition of the Intranuclear Inclusions of Fragile X-associated Tremor/Ataxia Syndrome. Acta Neuropathol. Commun. 2019, 7, 143. [Google Scholar] [CrossRef] [PubMed]
- Monaco, A.; Fraldi, A. Protein Aggregation and Dysfunction of Autophagy-Lysosomal Pathway: A Vicious Cycle in Lysosomal Storage Diseases. Front. Mol. Neurosci. 2020, 13, 37. [Google Scholar] [CrossRef] [PubMed]
- Menzies, F.M.; Fleming, A.; Caricasole, A.; Bento, C.F.; Andrews, S.P.; Ashkenazi, A.; Fullgrabe, J.; Jackson, A.; Sanchez, M.J.; Karabiyik, C.; et al. Autophagy and Neurodegeneration: Pathogenic Mechanisms and Therapeutic Opportunities. Neuron 2017, 93, 1015–1034. [Google Scholar] [CrossRef]
- Tsvetkov, A.S.; Arrasate, M.; Barmada, S.; Ando, D.M.; Sharma, P.; Shaby, B.A.; Finkbeiner, S. Proteostasis of polyglutamine varies among neurons and predicts neurodegeneration. Nat. Chem. Biol. 2013, 9, 586–592. [Google Scholar] [CrossRef]
- Gelpi, E.; Botta-Orfila, T.; Bodi, L.; Marti, S.; Kovacs, G.; Grau-Rivera, O.; Lozano, M.; Sánchez-Valle, R.; Muñoz, E.; Valldeoriola, F.; et al. Neuronal intranuclear (hyaline) inclusion disease and fragile X-associated tremor/ataxia syndrome: A morphological and molecular dilemma. Brain 2017, 140, e51. [Google Scholar] [CrossRef] [PubMed]
- Montine, T.J.; Phelps, C.H.; Beach, T.G.; Bigio, E.H.; Cairns, N.J.; Dickson, D.W.; Duyckaerts, C.; Frosch, M.P.; Masliah, E.; Mirra, S.S.; et al. National Institute on Aging-Alzheimer’s Association guidelines for the neuropathologic assessment of Alzheimer’s disease: A practical approach. Acta Neuropathol. 2012, 123, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Ruberu, N.N.; Sawabe, M.; Arai, T.; Tanaka, N.; Kakuta, Y.; Yamanouchi, H.; Murayama, S. Staging of argyrophilic grains: An age-associated tauopathy. J. Neuropathol. Exp. Neurol. 2004, 63, 911–918. [Google Scholar] [CrossRef]
- Nelson, P.T.; Dickson, D.W.; Trojanowski, J.Q.; Jack, C.R.; Boyle, P.A.; Arfanakis, K.; Rademakers, R.; Alafuzoff, I.; Attems, J.; Brayne, C.; et al. Limbic-predominant age-related TDP-43 encephalopathy (LATE): Consensus working group report. Brain 2019, 142, 1503–1527. [Google Scholar] [CrossRef]
- McKeith, I.G.; Boeve, B.F.; Dickson, D.W.; Halliday, G.; Taylor, J.P.; Weintraub, D.; Aarsland, D.; Galvin, J.; Attems, J.; Ballard, C.G.; et al. Diagnosis and management of dementia with Lewy bodies: Fourth consensus report of the DLB Consortium. Neurology 2017, 89, 88–100. [Google Scholar] [CrossRef]
- Braak, H.; Del Tredici, K.; Rüb, U.; de Vos, R.A.; Jansen Steur, E.N.; Braak, E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol. Aging 2003, 24, 197–211. [Google Scholar] [CrossRef]
- Crary, J.F.; Trojanowski, J.Q.; Schneider, J.A.; Abisambra, J.F.; Abner, E.L.; Alafuzoff, I.; Arnold, S.E.; Attems, J.; Beach, T.G.; Bigio, E.H.; et al. Primary age-related tauopathy (PART): A common pathology associated with human aging. Acta Neuropathol. 2014, 128, 755–766. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Mora, M.I.; Rodriguez-Revenga, L.; Madrigal, I.; Guitart-Mampel, M.; Garrabou, G.; Mila, M. Impaired Mitochondrial Function and Dynamics in the Pathogenesis of FXTAS. Mol. Neurobiol. 2017, 54, 6896–6902. [Google Scholar] [CrossRef]
- Montaner, D.; Dopazo, J. Multidimensional Gene Set Analysis of Genomic Data. PLoS ONE 2010, 5, e10348. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Rubinsztein, D.C.; Cuervo, A.M.; Ravikumar, B.; Sarkar, S.; Korolchuk, V.; Kaushik, S.; Klionsky, D.J. In search of an “autophagomometer”. Autophagy 2009, 5, 585–589. [Google Scholar] [CrossRef] [PubMed]
- Liebelt, F.; Vertegaal, A.C. Ubiquitin-dependent and independent roles of SUMO in proteostasis. Am. J. Physiol. Cell Physiol. 2016, 311, C284–C296. [Google Scholar] [CrossRef]
- Schimmel, J.; Larsen, K.M.; Matic, I.; van Hagen, M.; Cox, J.; Mann, M.; Andersen, J.S.; Vertegaal, A.C. The ubiquitin-proteasome system is a key component of the SUMO-2/3 cycle. Mol. Cell Proteom. 2008, 7, 2107–2122. [Google Scholar] [CrossRef]
- Tatham, M.H.; Matic, I.; Mann, M.; Hay, R.T. Comparative proteomic analysis identifies a role for SUMO in protein quality control. Sci. Signal. 2011, 4, rs4. [Google Scholar] [CrossRef]
- Balch, W.E.; Morimoto, R.I.; Dillin, A.; Kelly, J.W. Adapting proteostasis for disease intervention. Science 2008, 319, 916–919. [Google Scholar] [CrossRef]
- Yerbury, J.J.; Ooi, L.; Dillin, A.; Saunders, D.N.; Hatters, D.M.; Beart, P.M.; Cashman, N.R.; Wilson, M.R.; Ecroyd, H. Walking the tightrope: Proteostasis and neurodegenerative disease. J. Neurochem. 2016, 137, 489–505. [Google Scholar] [CrossRef]
- Chen, B.; Retzlaff, M.; Roos, T.; Frydman, J. Cellular strategies of protein quality control. Cold Spring Harb. Perspect. Biol. 2011, 3, a004374. [Google Scholar] [CrossRef] [PubMed]
- Nah, J.; Yuan, J.; Jung, Y.K. Autophagy in neurodegenerative diseases: From mechanism to therapeutic approach. Mol. Cells. 2015, 38, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Ross-Inta, C.; Omanska-Klusek, A.; Wong, S.; Barrow, C.; Garcia-Arocena, D.; Iwahashi, C.; Berry-Kravis, E.; Hagerman, R.J.; Hagerman, P.J.; Giulivi, C. Evidence of mitochondrial dysfunction in fragile X-associated tremor/ataxia syndrome. Biochem. J. 2010, 429, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Napoli, E.; Ross-Inta, C.; Wong, S.; Omanska-Klusek, A.; Barrow, C.; Iwahashi, C.; Garcia-Arocena, D.; Sakaguchi, D.; Berry-Kravis, E.; Hagerman, R.; et al. Altered zinc transport disrupts mitochondrial protein processing/import in fragile X-associated tremor/ataxia syndrome. Hum. Mol. Genet. 2011, 20, 3079–3092. [Google Scholar] [CrossRef]
- Lin, Y.; Tang, C.; He, H.; Duan, R. Activation of mTOR ameliorates fragile X premutation rCGG repeat-mediated neurodegeneration. PLoS ONE 2013, 8, e62572. [Google Scholar] [CrossRef]
- Das, G.; Shravage, B.V.; Baehrecke, E.H. Regulation and function of autophagy during cell survival and cell death. Cold Spring Harb. Perspect. Biol. 2012, 4, a008813. [Google Scholar] [CrossRef]
- Saitoh, H.; Hinchey, J. Functional heterogeneity of small ubiquitin-related protein modifiers SUMO-1 versus SUMO-2/3. J. Biol. Chem. 2000, 275, 6252–6258. [Google Scholar] [CrossRef]
- Taylor, J.P.; Tanaka, F.; Robitschek, J.; Sandoval, C.M.; Taye, A.; Markovic-Plese, S.; Fischbeck, K.H. Aggresomes protect cells by enhancing the degradation of toxic polyglutamine-containing protein. Hum. Mol. Genet. 2003, 12, 749–757. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| ID Sample | Sex | Age at Disease Onset (Years) | Age at Death (Years) | Expanded FMR1 (CGG)n | Frontal Cortex SUMO Intranuclear Inclusions | Hippocampal SUMO Intranuclear Inclusions | Cerebellar SUMO Intranuclear Inclusions | Clinical Symptoms | Relevant Neuropathological Findings |
|---|---|---|---|---|---|---|---|---|---|
| FXTAS_brain_1 | Female | 73 | 78 | 77 | Isolated GII in white matter | Isolated GII inclusion in dentate gyrus | 0 | Dementia, tremor, ataxia | Low AD-NC (A1B1C0), AGD (stage I), e, VBI |
| FXTAS_brain_2 | Female | 70 | 95 | 60 | mild GII and NII in gray matter | Moderate NII and GII in CA4/CA3 | Moderate GII and NII in Grey matter | Hypoacusia, tremor, ataxia, dementia | High AD-NC (A3B3C3), LBD (Braak 1), VBI |
| Control_brain_1 | Female | - | 81 | - | none | none | none | Control | Low AD-NC (A3B1C2), LATE-NC (stage 2) |
| Control_brain_2 | Female | - | 90 | - | none | none | none | Control, stroke | PART (definitive, Braak II), VBI |
| Sex | Age (Years) at Skin Biopsy | CGG Repeat Size | Clinical Diagnosis | |
|---|---|---|---|---|
| FXTAS_1 | Male | 70 | 65 | Ataxia and tremor |
| FXTAS_2 | Male | 62 | 95 | Ataxia and tremor |
| FXTAS_3 | Female | 53 | 30/149 | Intention tremor and memory defects |
| FXTAS_4 | Male | 66 | 98 | Ataxia, tremor and dementia |
| Control_1 | Female | 82 | 21/30 | control |
| Control_2 | Male | 57 | 30 | control |
| Control_3 | Male | 86 | 30 | control |
| Control_4 | Female | 68 | 30/32 | control |
| Brain Sample | Fibroblast Sample | ||||||
|---|---|---|---|---|---|---|---|
| PATH_ID | PATH_NAME | Size | padj | LOR | Size | padj | LOR |
| R-HSA-3108232 | SUMO E3 ligases SUMOylate target proteins | 146 | 0.003515 | 0.3012 | 151 | 0.0004707 | 0.3446 |
| R-HSA-2990846 | SUMOylation | 152 | 0.005241 | 0.2856 | 157 | 0.0002597 | 0.349 |
| R-HSA-3108214 | SUMOylation of DNA damage response and repair proteins | 71 | 0.0000425 | 0.5717 | 70 | 0.009913 | 0.3955 |
| R-HSA-4570464 | SUMOylation of RNA binding proteins | 42 | 0.04191 | 0.4147 | 41 | 0.0321 | 0.4362 |
| R-HSA-4085377 | SUMOylation of SUMOylation proteins | 31 | 0.03038 | 0.5127 | NS | ||
| R-HSA-3232142 | SUMOylation of ubiquitinylation proteins | 34 | 0.04552 | 0.4545 | NS | ||
| GO:0016241 | Regulation of macroautophagy | 48 | 0.00108 | −0.6426 | 47 | 0.01945 | −0.4926 |
| Genes Related to SUMOylation Processes | Genes Related to Autophagy Processes | ||||
|---|---|---|---|---|---|
| Gene | Fold Change | padj | Gene | Fold Change | padj |
| THRB | −2.231 | 0.001 | ATP6V1H | −1.486 | 0.004 |
| SATB1 | −1.555 | 0 | UCHL1 | −2.233 | 0 |
| CHD3 | −1.401 | 0.013 | ATP6V1G2 | −2.104 | 0 |
| PPARGC1A | −1.374 | 0.005 | CDK5R1 | −2.059 | 0.002 |
| SATB2 | −1.168 | 0.008 | ATP6V1A | −2.03 | 0 |
| SUMO3 | −0.876 | 0.027 | ATP6V1B2 | −2.004 | 0 |
| SAE1 | −0.728 | 0.016 | CDK5 | −1.872 | 0 |
| NUP93 | −0.669 | 0.048 | PRKAA2 | −1.774 | 0 |
| SUMO1 | −0.659 | 0.029 | ATP6V1C1 | −1.739 | 0 |
| CETN2 | −0.593 | 0.039 | ATP13A2 | −1.096 | 0.012 |
| NRIP1 | −0.57 | 0.027 | ATP6V1D | −1.008 | 0.01 |
| NUP50 | −0.557 | 0.027 | ATP6V0A1 | −0.939 | 0 |
| ING2 | −0.548 | 0.019 | ATP6V0C | −0.936 | 0.008 |
| TP53BP1 | −0.523 | 0.008 | PRKACA | −0.732 | 0.016 |
| NSMCE3 | −0.519 | 0.023 | ATP6V0D1 | −0.717 | 0.033 |
| INCENP | −0.409 | 0.047 | VPS26B | −0.712 | 0.001 |
| RAE1 | −0.344 | 0.048 | MAPK8 | −0.,708 | 0.007 |
| TRIM28 | 0.292 | 0.035 | VPS35 | −0.669 | 0.047 |
| SP3 | 0.329 | 0.041 | UBQLN2 | −0.608 | 0.001 |
| SENP1 | 0.343 | 0.036 | UBQLN1 | −0.451 | 0.003 |
| SEC13 | 0.367 | 0.024 | SNX5 | 0.319 | 0.008 |
| ZBED1 | 0.435 | 0.009 | HDAC6 | 0.436 | 0.032 |
| PARP1 | 0.462 | 0.027 | SH3GLB1 | 0.542 | 0.016 |
| NUP43 | 0.473 | 0.015 | ATP6V0E1 | 0.991 | 0.003 |
| AAAS | 0.515 | 0.013 | |||
| STAG1 | 0.555 | 0.002 | |||
| SMC3 | 0.569 | 0.007 | |||
| NUP160 | 0.606 | 0.032 | |||
| NSMCE1 | 0.638 | 0.002 | |||
| PCNA | 0.638 | 0.033 | |||
| NUP37 | 0.645 | 0.036 | |||
| NSMCE4A | 0.74 | 0.033 | |||
| STAG2 | 0.809 | 0.018 | |||
| TP53 | 0.911 | 0.02 | |||
| NFKBIA | 1.03 | 0.046 | |||
| SP100 | 1.165 | 0.005 | |||
| CDKN2A | 1.522 | 0.028 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvarez-Mora, M.I.; Garrabou, G.; Molina-Porcel, L.; Grillo-Risco, R.; Garcia-Garcia, F.; Barcos, T.; Cantó-Santos, J.; Rodriguez-Revenga, L. Exploration of SUMO2/3 Expression Levels and Autophagy Process in Fragile X-Associated Tremor/Ataxia Syndrome: Addressing Study Limitations and Insights for Future Research. Cells 2023, 12, 2364. https://doi.org/10.3390/cells12192364
Alvarez-Mora MI, Garrabou G, Molina-Porcel L, Grillo-Risco R, Garcia-Garcia F, Barcos T, Cantó-Santos J, Rodriguez-Revenga L. Exploration of SUMO2/3 Expression Levels and Autophagy Process in Fragile X-Associated Tremor/Ataxia Syndrome: Addressing Study Limitations and Insights for Future Research. Cells. 2023; 12(19):2364. https://doi.org/10.3390/cells12192364
Chicago/Turabian StyleAlvarez-Mora, Maria Isabel, Glòria Garrabou, Laura Molina-Porcel, Ruben Grillo-Risco, Francisco Garcia-Garcia, Tamara Barcos, Judith Cantó-Santos, and Laia Rodriguez-Revenga. 2023. "Exploration of SUMO2/3 Expression Levels and Autophagy Process in Fragile X-Associated Tremor/Ataxia Syndrome: Addressing Study Limitations and Insights for Future Research" Cells 12, no. 19: 2364. https://doi.org/10.3390/cells12192364
APA StyleAlvarez-Mora, M. I., Garrabou, G., Molina-Porcel, L., Grillo-Risco, R., Garcia-Garcia, F., Barcos, T., Cantó-Santos, J., & Rodriguez-Revenga, L. (2023). Exploration of SUMO2/3 Expression Levels and Autophagy Process in Fragile X-Associated Tremor/Ataxia Syndrome: Addressing Study Limitations and Insights for Future Research. Cells, 12(19), 2364. https://doi.org/10.3390/cells12192364