
Cell–Fibronectin Interactions and Actomyosin Contractility Regulate the Segmentation Clock and Spatio-Temporal Somite Cleft Formation during Chick Embryo Somitogenesis

 , , ,
, , ,  and
and 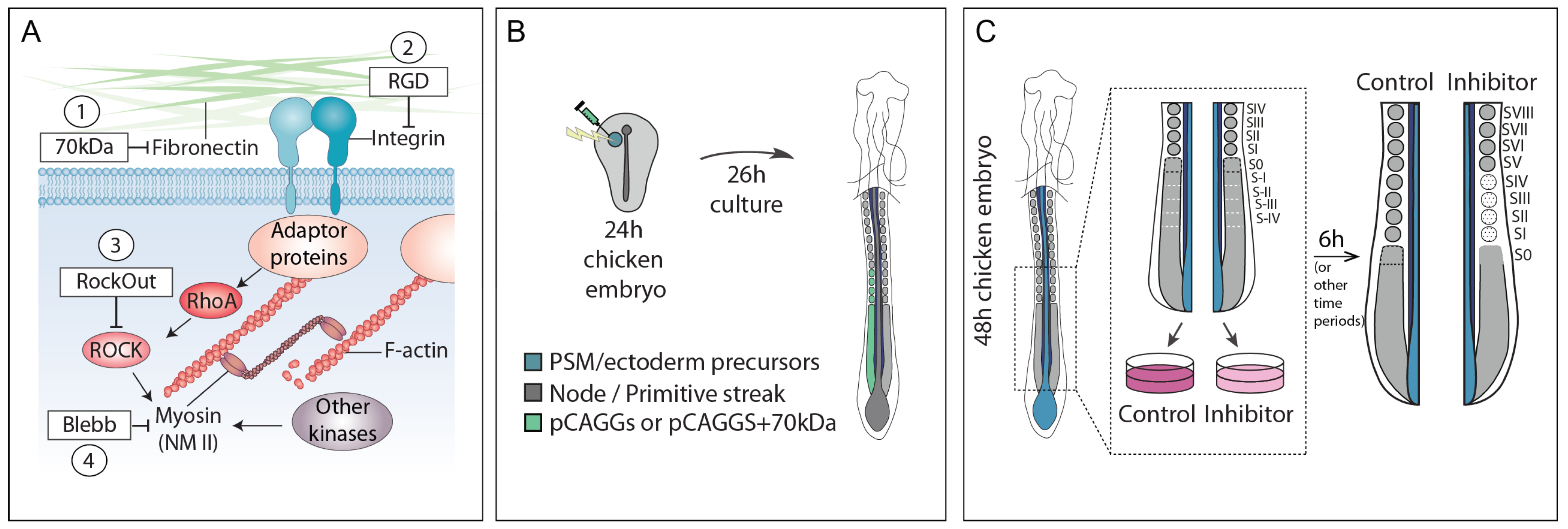{kind=link}
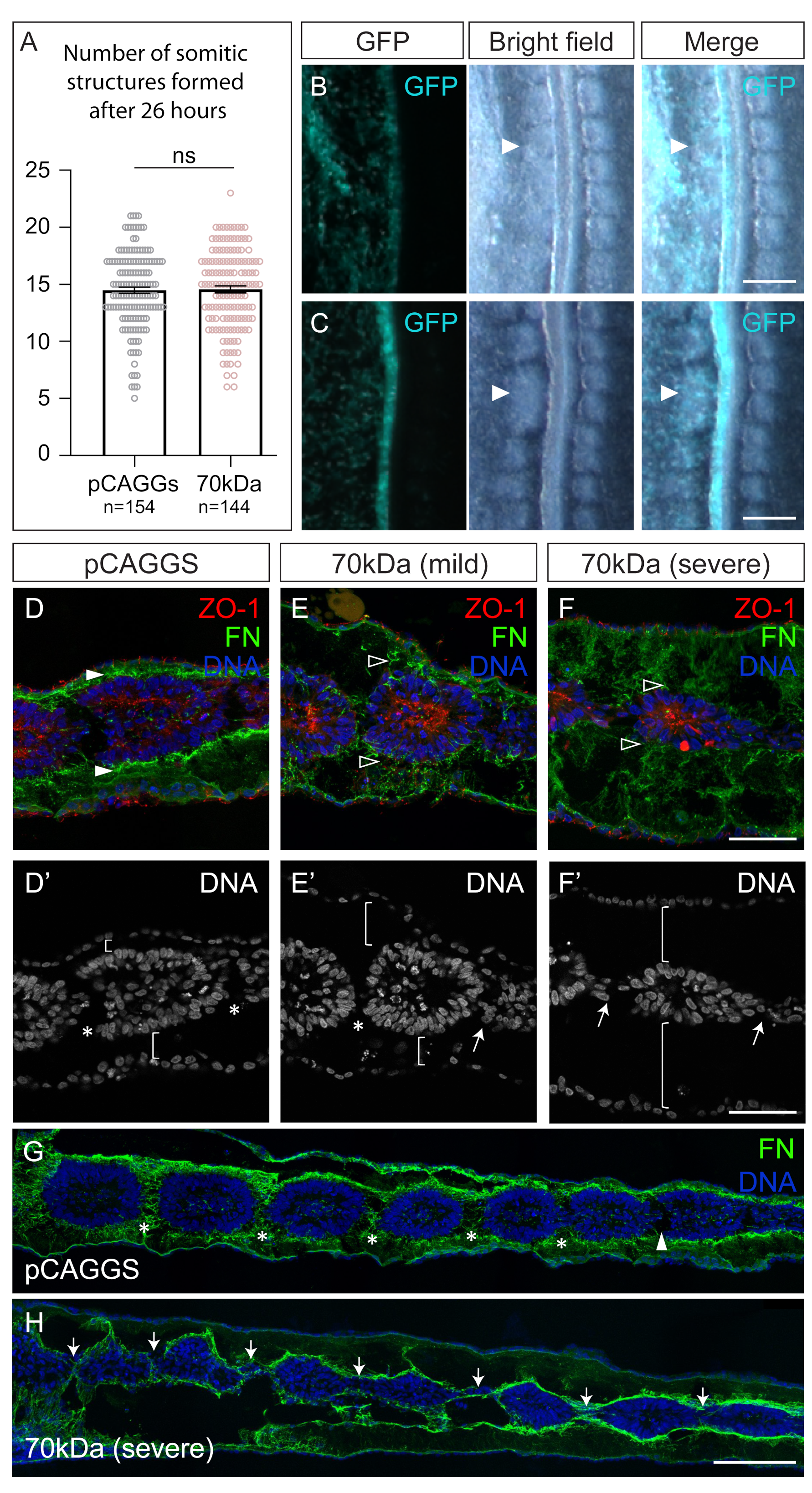{kind=link}
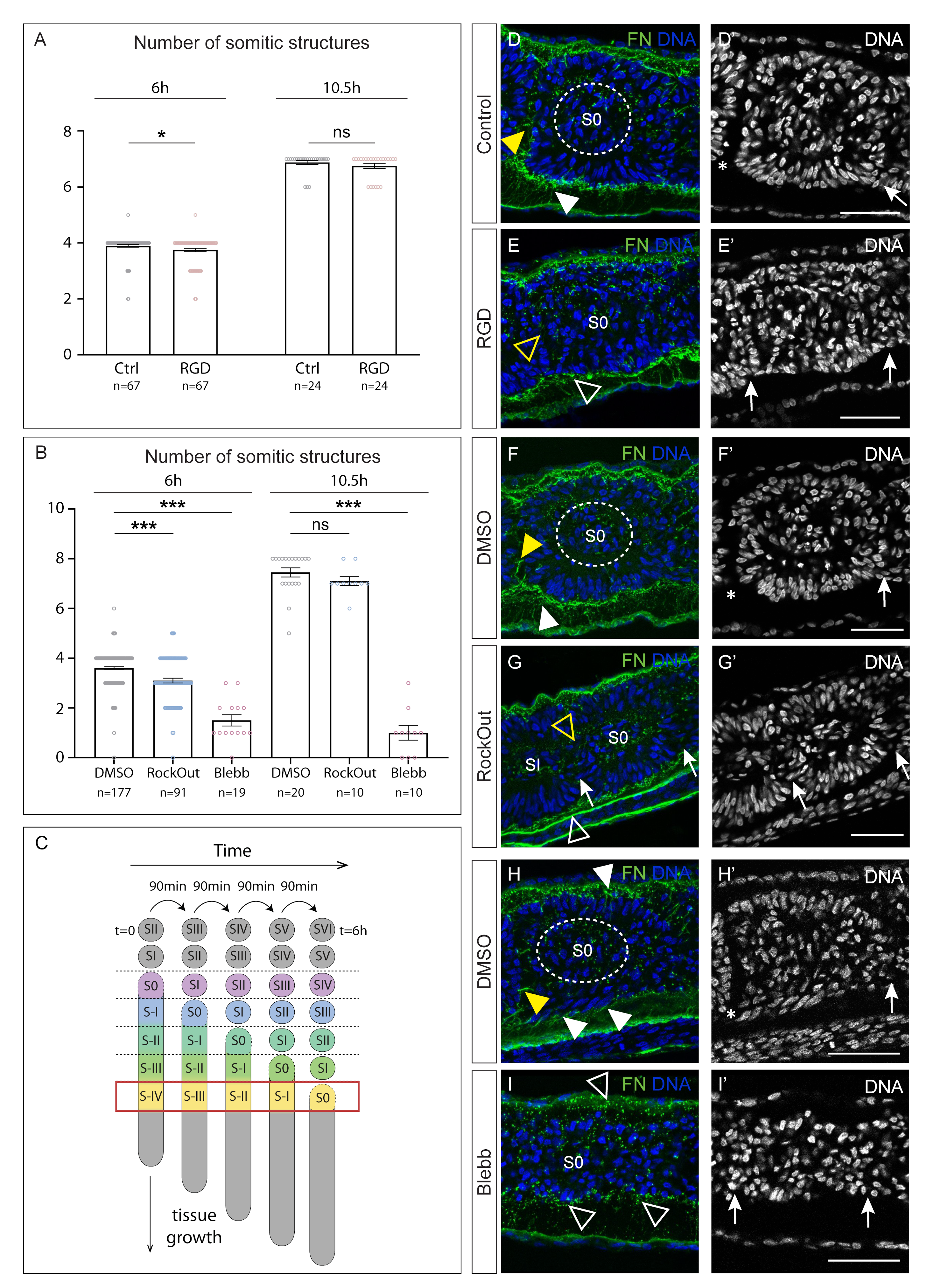{kind=link}
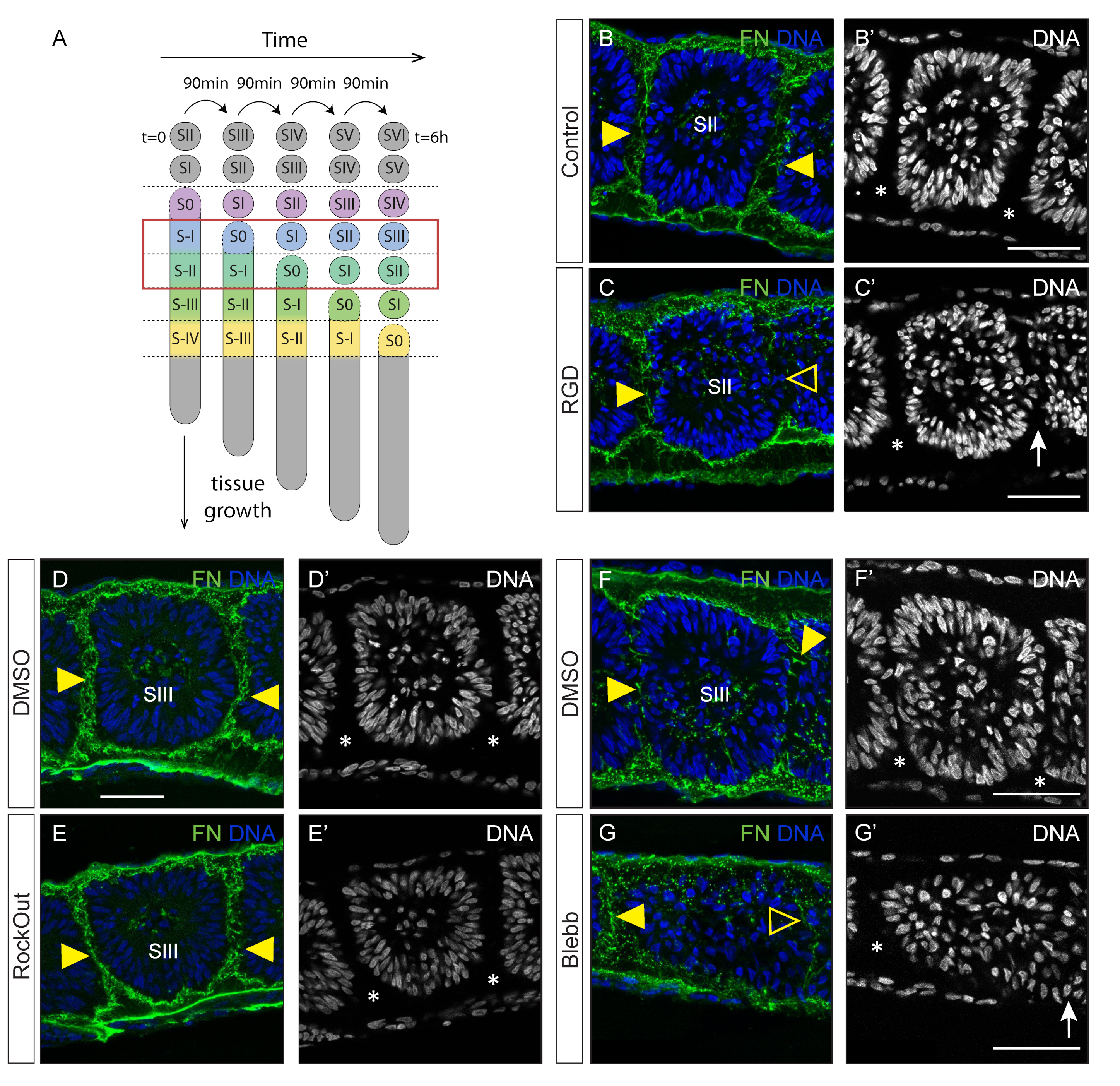{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
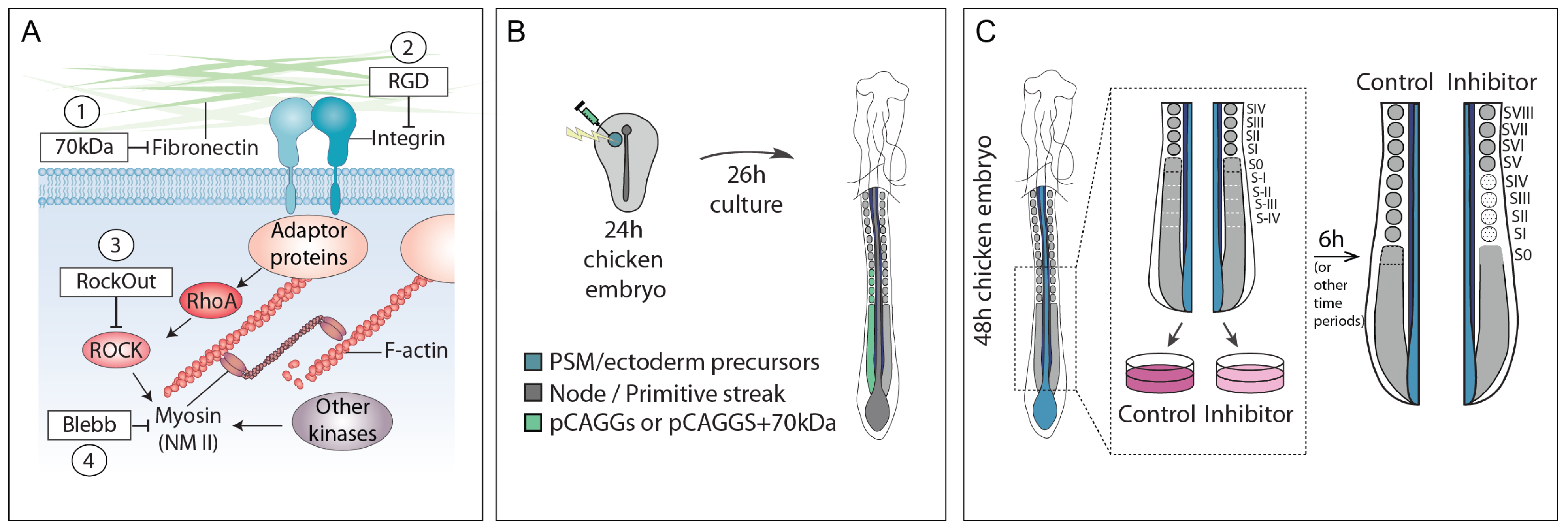
2.1. Embryos and Experimental Design
2.2. Embryo Electroporation and ex ovo Culture
2.3. Embryo Explant Culture and Treatments
2.4. Immunohistochemistry
2.5. In Situ Hybridization
2.6. Sample Preparation and Imaging
2.7. Statistical Analysis
3. Results
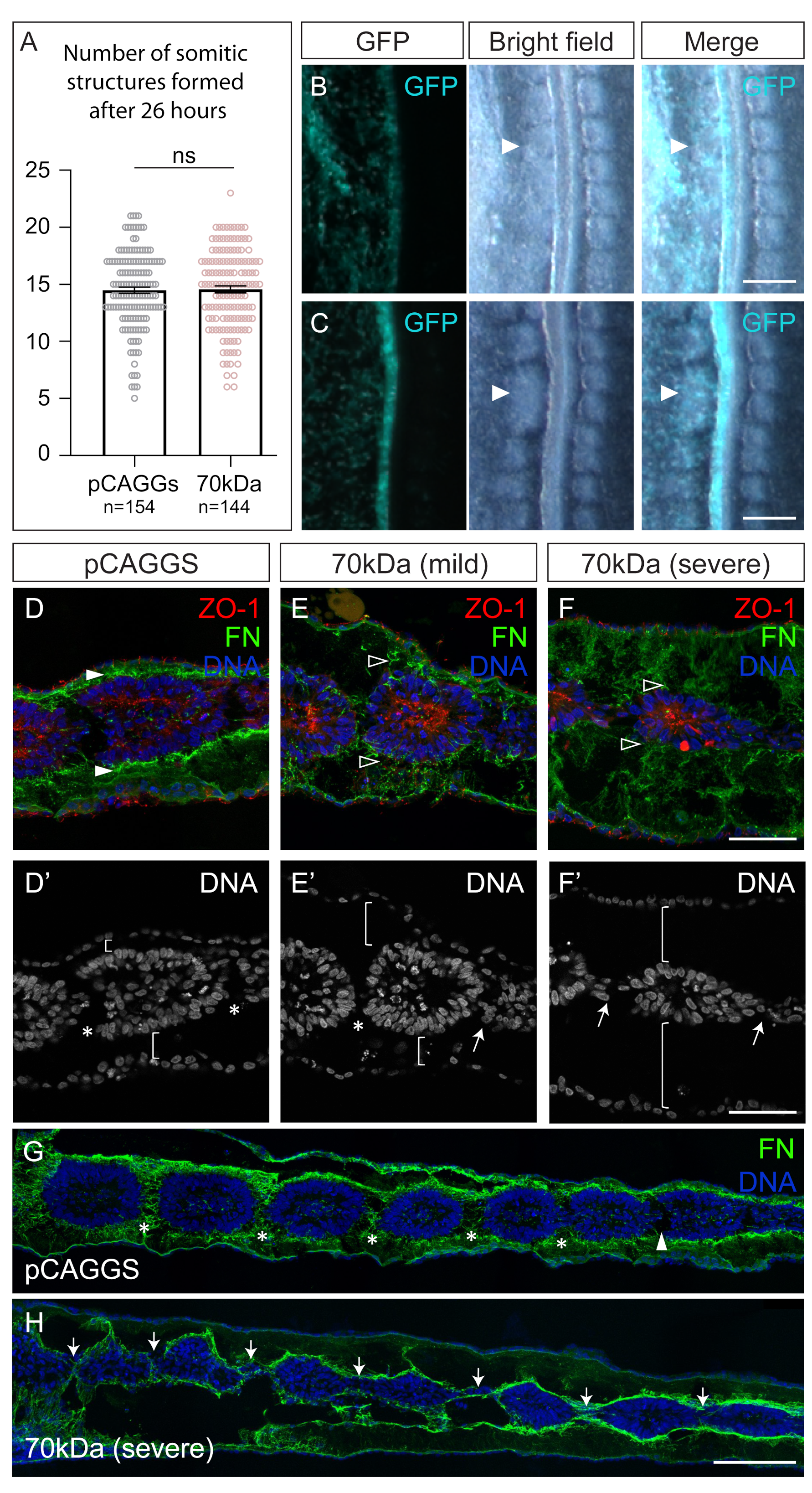
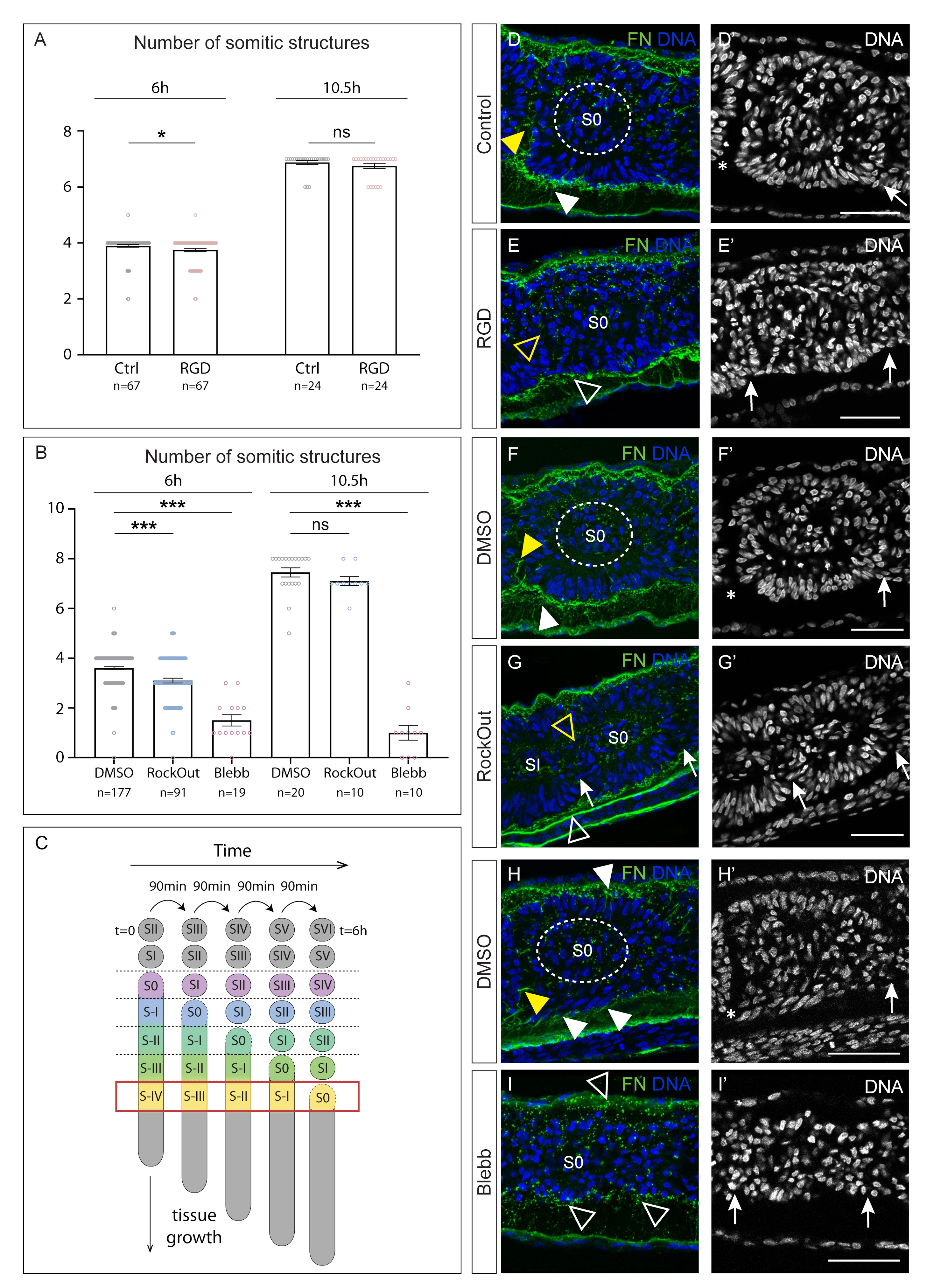
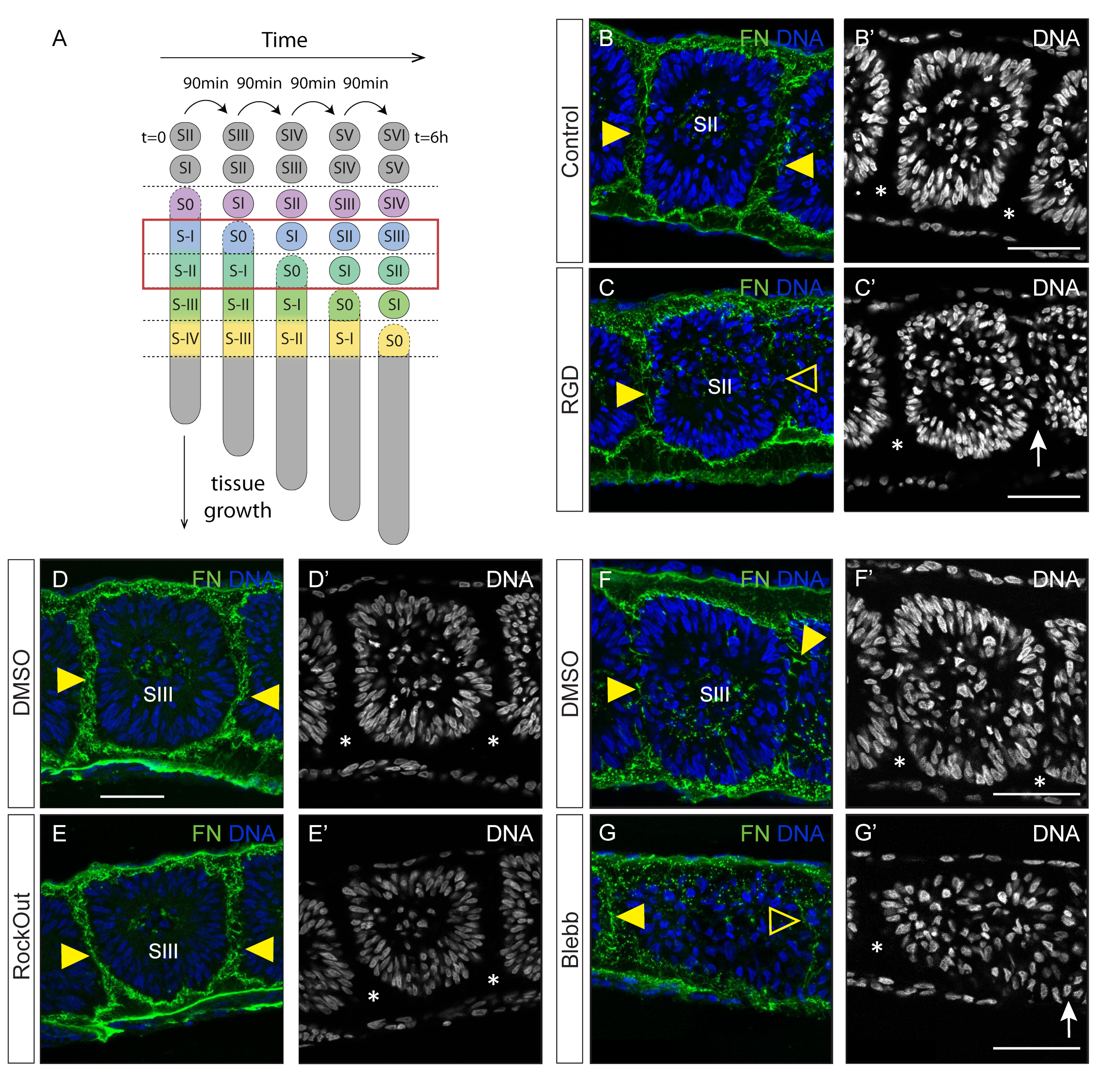
3.1. Interfering with Fibronectin Matrix Assembly, Cell–Fibronectin Binding or Actomyosin Contractility Leads to Defective Somite Cleft Formation
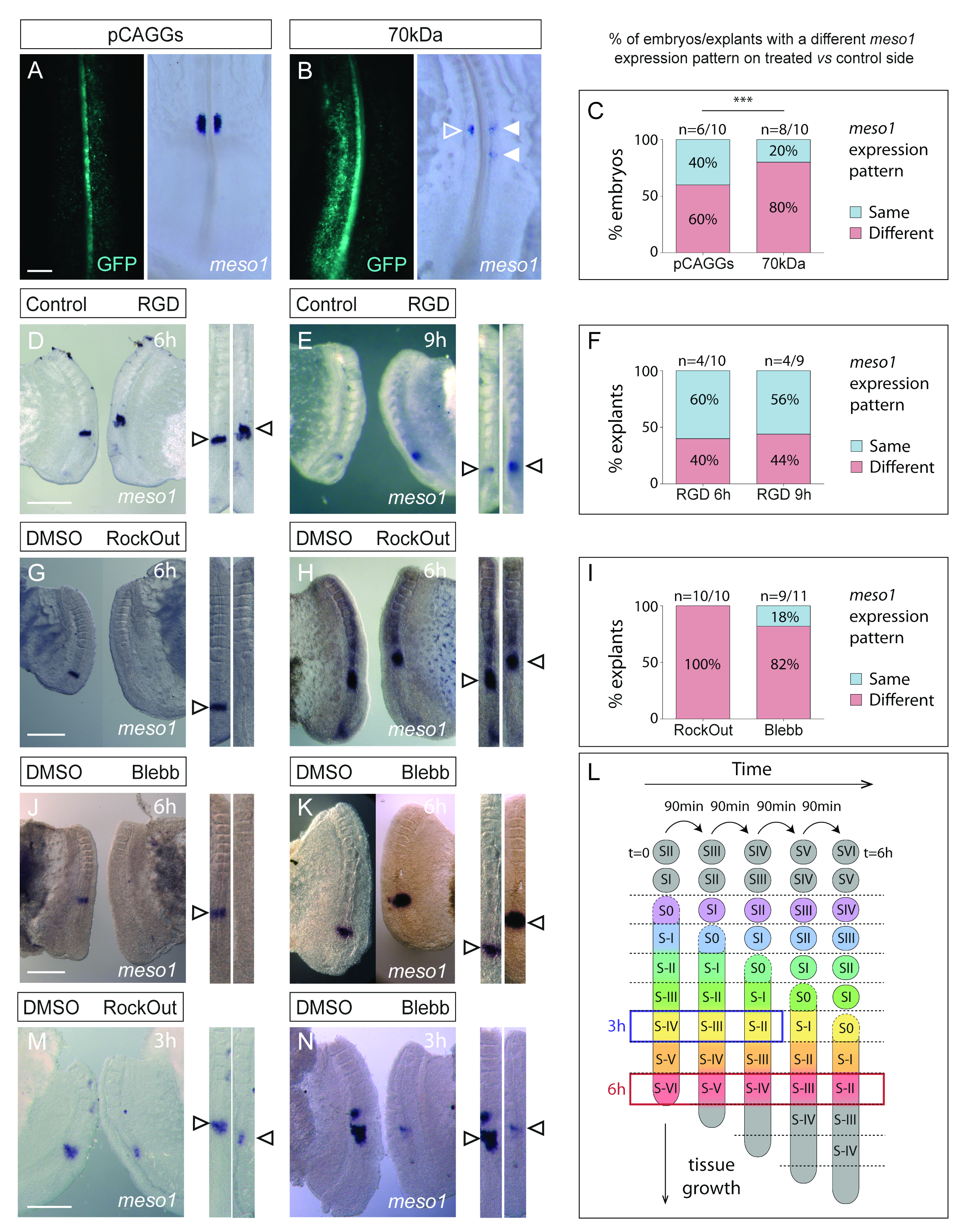
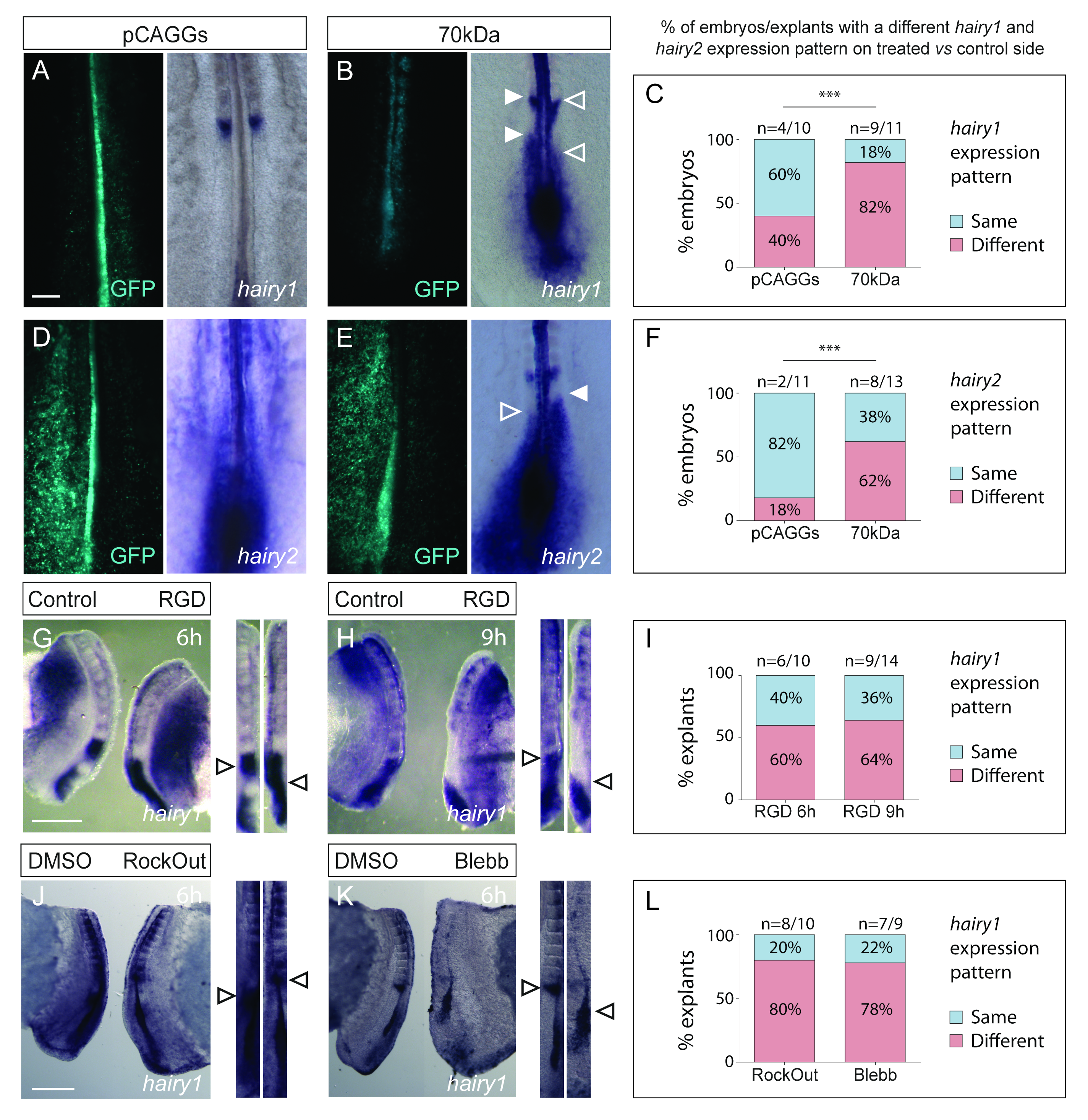
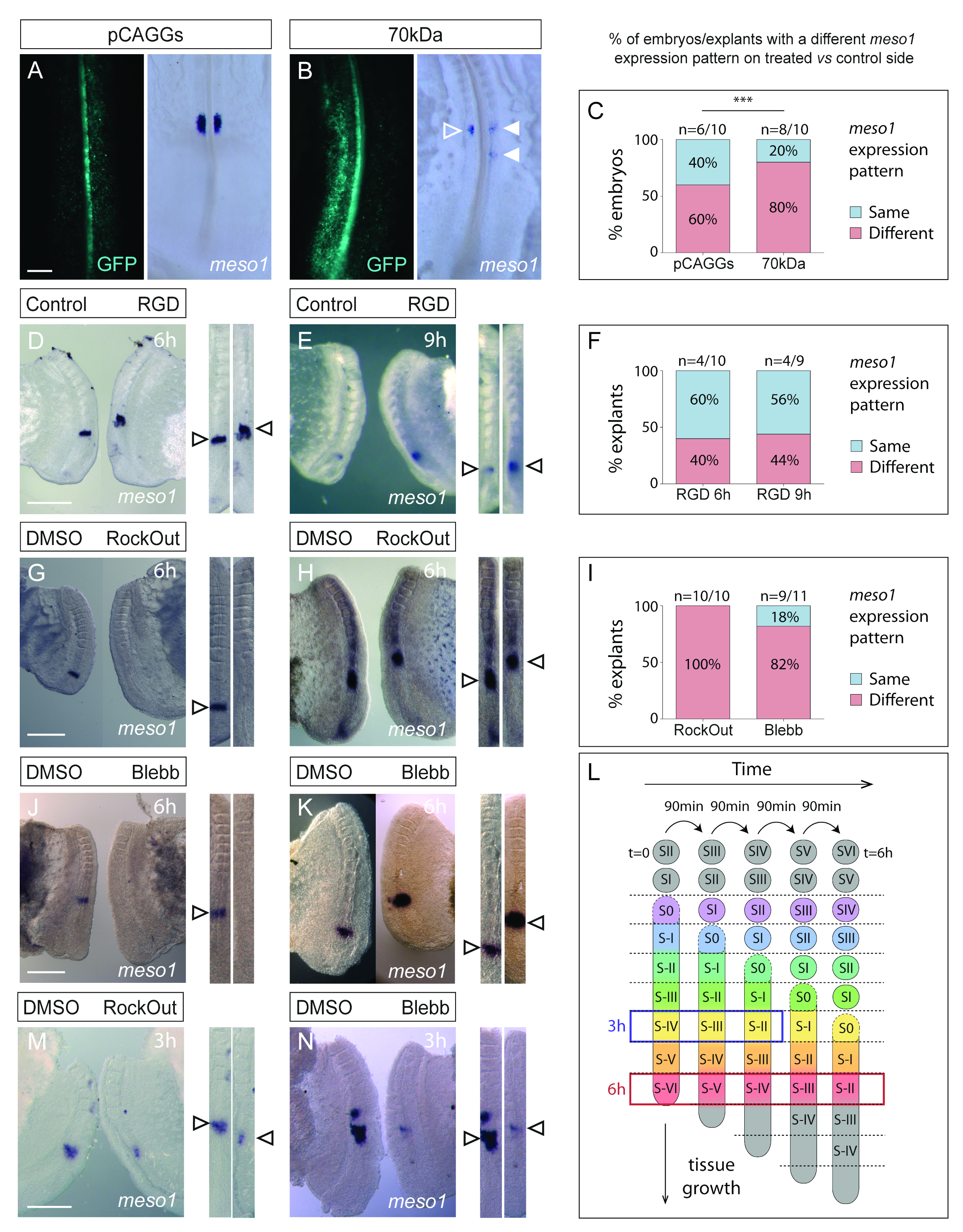
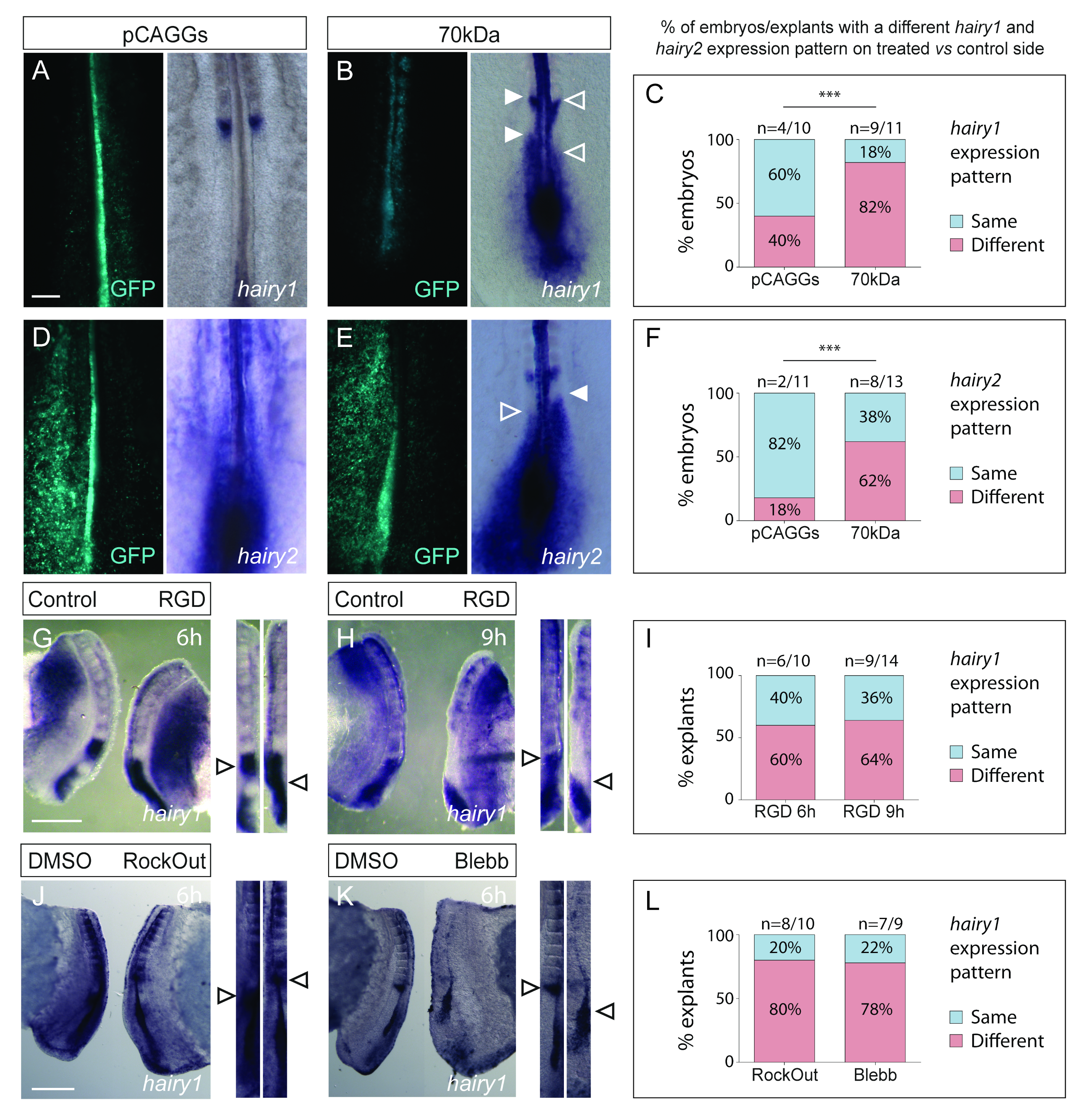
3.2. Interfering with Fibronectin Matrix Assembly, Cell–Fibronectin Binding or Actomyosin Contractility Alters meso1 and Cyclic hairy1 Expression
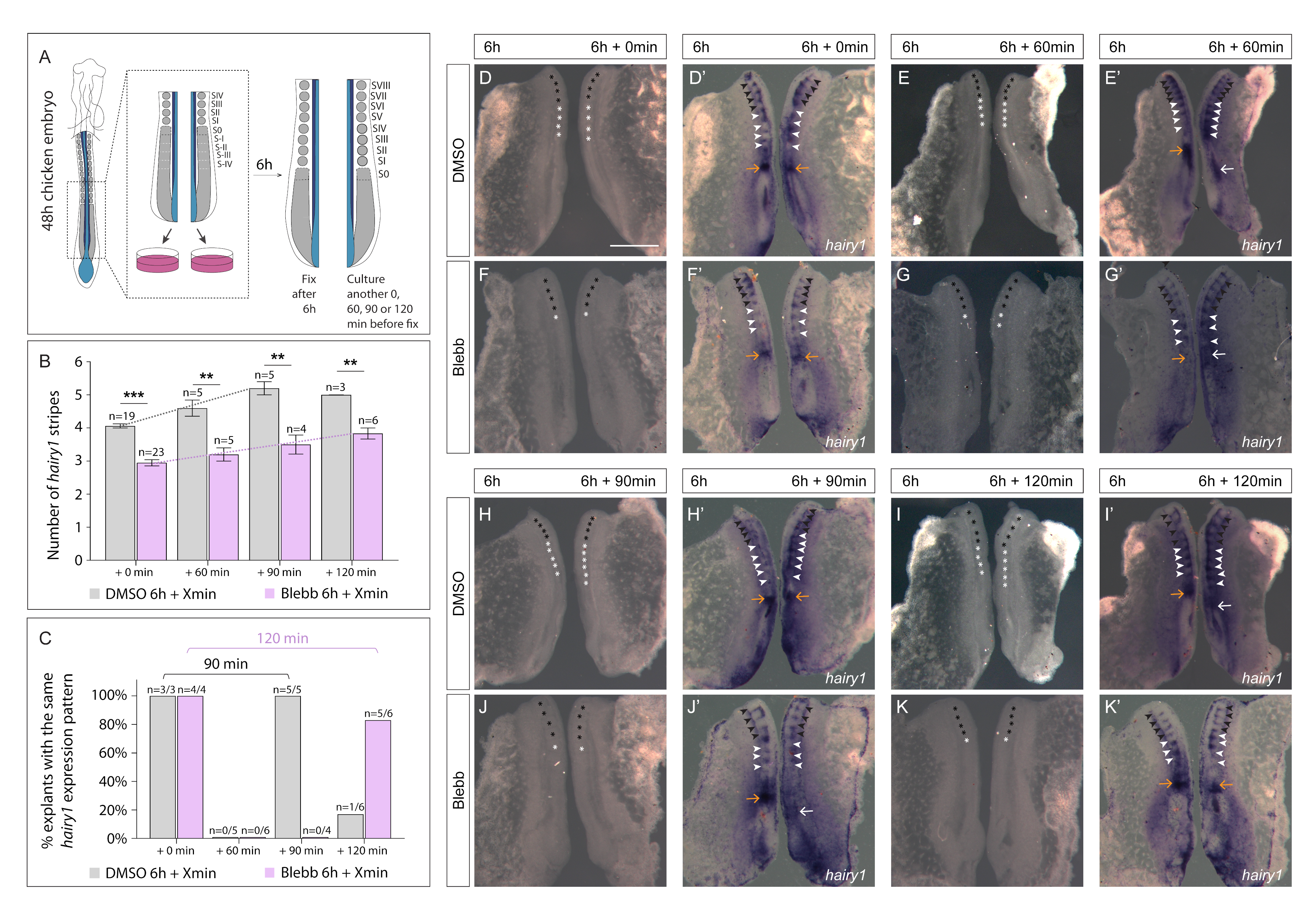
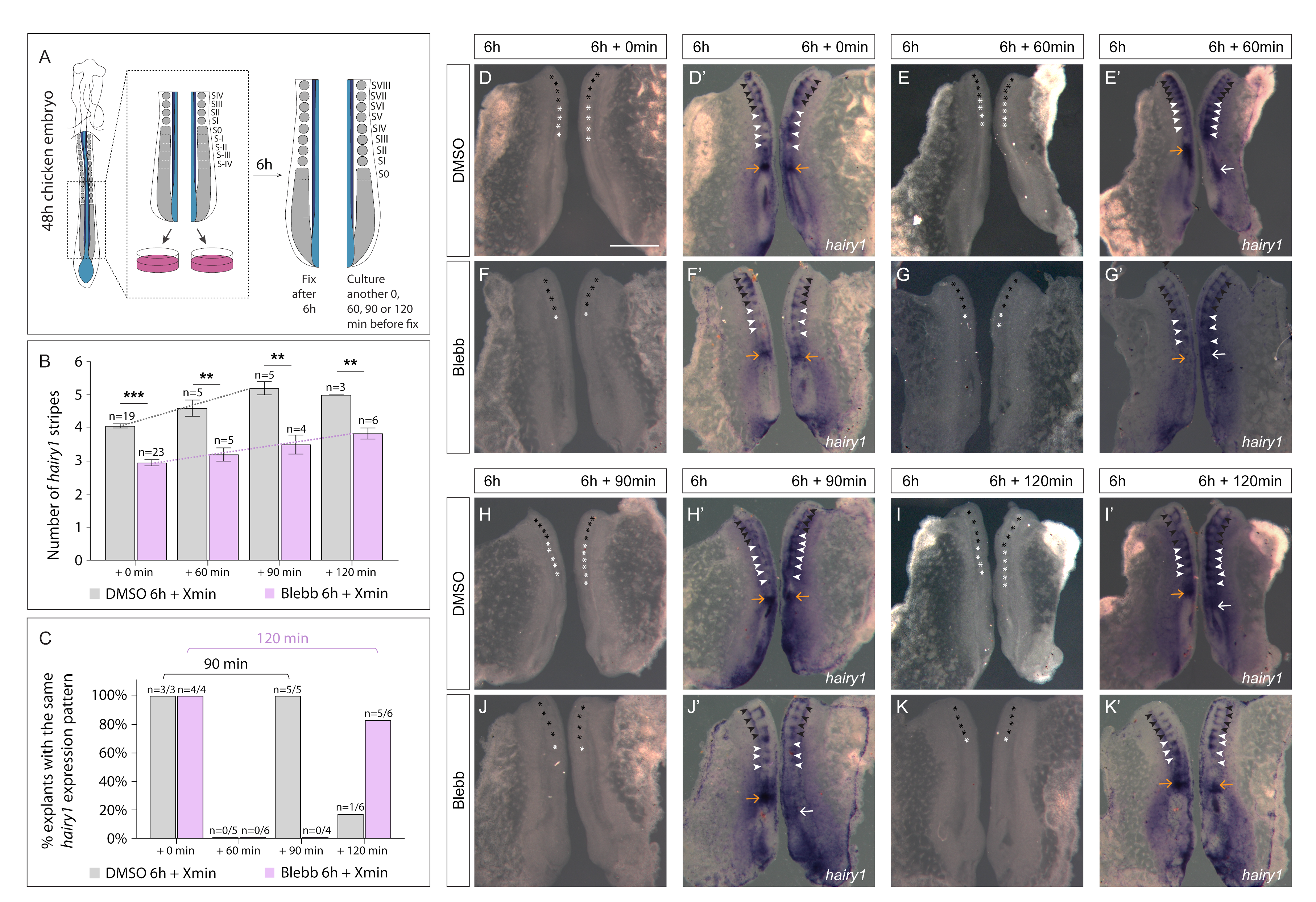
3.3. Delayed Periodicity of hairy1 Oscillations in Explants Treated with Blebbistatin
4. Discussion
4.1. Integrin–Fibronectin Engagement and ROCK-Independent Actomyosin Contractility Are Required for Somite Epithelialization
4.2. A Fibronectin–Integrin–ROCK–NM II Axis Contributes to the Prepatterning of the PSM for Cleft Formation by Regulating the Cycles of hairy1 and meso1 Expression
4.3. The PSM Cells’ Mechanical Landscape as a New Player in Temporal Control of the Segmentation Clock
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Schwarzbauer, J.E.; DeSimone, D.W. Fibronectins, their fibrillogenesis, and in vivo functions. Cold Spring Harb. Perspect. Biol. 2011, 3, a005041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dzamba, B.J.; DeSimone, D.W. Extracellular Matrix (ECM) and the Sculpting of Embryonic Tissues. Curr. Top. Dev. Biol. 2018, 130, 245–274. [Google Scholar] [CrossRef]
- George, E.L.; Georges-Labouesse, E.N.; Patel-King, R.S.; Rayburn, H.; Hynes, R.O. Defects in mesoderm, neural tube and vascular development in mouse embryos lacking fibronectin. Development 1993, 119, 1079–1091. [Google Scholar] [CrossRef] [PubMed]
- Davidson, L.A.; Marsden, M.; Keller, R.; DeSimone, D.W. Integrin α5β1 and Fibronectin Regulate Polarized Cell Protrusions Required for Xenopus Convergence and Extension. Curr. Biol. 2006, 16, 833–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsden, M.; DeSimone, D.W. Regulation of cell polarity, radial intercalation and epiboly in Xenopus: Novel roles for integrin and fibronectin. Development 2001, 128, 3635–3647. [Google Scholar] [CrossRef] [PubMed]
- Rifes, P.; Carvalho, L.; Lopes, C.; Andrade, R.P.; Rodrigues, G.; Palmeirim, I.; Thorsteinsdóttir, S. Redefining the role of ectoderm in somitogenesis: A player in the formation of the fibronectin matrix of presomitic mesoderm. Development 2007, 134, 3155–3165. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Kim, H.Y.; Davidson, L.A. Actomyosin stiffens the vertebrate embryo during crucial stages of elongation and neural tube closure. Development 2009, 136, 677–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theocharis, A.D.; Manou, D.; Karamanos, N.K. The extracellular matrix as a multitasking player in disease. FEBS J. 2019, 286, 2830–2869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, Y.; Schwarzbauer, J.E. Fibronectin fibrillogenesis, a cell-mediated matrix assembly process. Matrix Biol. 2005, 24, 389–399. [Google Scholar] [CrossRef]
- Singh, P.; Carraher, C.; Schwarzbauer, J.E. Assembly of fibronectin extracellular matrix. Annu. Rev. Cell Dev. Biol. 2010, 26, 397–419. [Google Scholar] [CrossRef] [Green Version]
- Dalton, C.J.; Lemmon, C.A. Fibronectin: Molecular structure, fibrillar structure and mechanochemical signaling. Cells 2021, 10, 2443. [Google Scholar] [CrossRef] [PubMed]
- Campbell, I.D.; Humphries, M.J. Integrin structure, activation, and interactions. Cold Spring Harb. Perspect. Biol. 2011, 3, a004994. [Google Scholar] [CrossRef] [Green Version]
- Wolfenson, H.; Lavelin, I.; Geiger, B. Dynamic regulation of the structure and functions of integrin adhesions. Dev. Cell 2013, 24, 447–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horton, E.R.; Humphries, J.D.; James, J.; Jones, M.C.; Askari, J.A.; Humphries, M.J. The integrin adhesome network at a glance. J. Cell Sci. 2016, 129, 4159–4163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaidel-Bar, R.; Zhenhuan, G.; Luxenburg, C. The contractome—A systems view of actomyosin contractility in non-muscle cells. J. Cell Sci. 2015, 128, 2209–2217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaidel-Bar, R. Cadherin adhesome at a glance. J. Cell Sci. 2013, 126, 373–378. [Google Scholar] [CrossRef] [Green Version]
- Burute, M.; Thery, M. Spatial segregation between cell-cell and cell-matrix adhesions. Curr. Opin. Cell Biol. 2012, 24, 628–636. [Google Scholar] [CrossRef]
- Mui, K.L.; Chen, C.S.; Assoian, R.K. The mechanical regulation of integrin-cadherin crosstalk organizes cells, signaling and forces. J. Cell Sci. 2016, 129, 1093–1100. [Google Scholar] [CrossRef] [Green Version]
- Wolfenson, H.; Yang, B.; Sheetz, M.P. Steps in Mechanotransduction Pathways that Control Cell Morphology. Annu. Rev. Physiol. 2019, 81, 585–605. [Google Scholar] [CrossRef]
- Bailey, C.; Dale, K. Somitogenesis in Vertebrate Development. eLS 2015, 1–15. [Google Scholar] [CrossRef]
- Christ, B.; Huang, R.; Scaal, M. Amniote somite derivatives. Dev. Dyn. 2007, 236, 2382–2396. [Google Scholar] [CrossRef]
- Dequéant, M.L.; Glynn, E.; Gaudenz, K.; Wahl, M.; Chen, J.; Mushegian, A.; Pourquié, O. A complex oscillating network of signaling genes underlies the mouse segmentation clock. Science 2006, 314, 1595–1598. [Google Scholar] [CrossRef] [PubMed]
- Masamizu, Y.; Ohtsuka, T.; Takashima, Y.; Nagahara, H.; Takenaka, Y.; Yoshikawa, K.; Okamura, H.; Kageyama, R. Real-time imaging of the somite segmentation clock: Revelation of unstable oscillators in the individual presomitic mesoderm cells. Proc. Natl. Acad. Sci. USA 2006, 103, 1313–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmeirim, I.; Henrique, D.; Ish-Horowicz, D.; Pourquié, O. Avian hairy gene expression identifies a molecular clock linked to vertebrate segmentation and somitogenesis. Cell 1997, 91, 639–648. [Google Scholar] [CrossRef] [Green Version]
- Morimoto, M.; Takahashi, Y.; Endo, M.; Saga, Y. The Mesp2 transcription factor establishes segmental borders by suppressing Notch activity. Nature 2005, 435, 354–359. [Google Scholar] [CrossRef]
- Takahashi, Y.; Koizumi, K.I.; Takagi, A.; Kitajima, S.; Inoue, T.; Koseki, H.; Saga, Y. Mesp2 initiates somite segmentation through the Notch signalling pathway. Nat. Genet. 2000, 25, 390–396. [Google Scholar] [CrossRef]
- Saga, Y. The synchrony and cyclicity of developmental events. Cold Spring Harb. Perspect. Biol. 2012, 4, a008201. [Google Scholar] [CrossRef]
- Barrios, A.; Poole, R.J.; Durbin, L.; Brennan, C.; Holder, N.; Wilson, S.W. Eph/Ephrin signaling regulates the mesenchymalto-epithelial transition of the paraxial mesoderm during somite morphogenesis. Curr. Biol. 2003, 13, 1571–1582. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, Y.; Morimoto, M.; Takahashi, Y.; Koseki, H.; Saga, Y. Identification of Epha4 enhancer required for segmental expression and the regulation by Mesp2. Development 2006, 133, 2517–2525. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, T.; Sato, Y.; Saito, D.; Tadokoro, R.; Takahashi, Y. EphrinB2 coordinates the formation of a morphological boundary and cell epithelialization during somite segmentation. Proc. Natl. Acad. Sci. USA 2009, 106, 7467–7472. [Google Scholar] [CrossRef] [Green Version]
- Martins, G.G.; Rifes, P.; Amaândio, R.; Rodrigues, G.; Palmeirim, I.; Thorsteinsdóttir, S. Dynamic 3D cell rearrangements guided by a fibronectin matrix underlie somitogenesis. PLoS ONE 2009, 4, e7429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jülich, D.; Cobb, G.; Melo, A.M.; McMillen, P.; Lawton, A.K.; Mochrie, S.G.J.; Rhoades, E.; Holley, S.A. Cross-Scale Integrin Regulation Organizes ECM and Tissue Topology. Dev. Cell 2015, 34, 33–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georges-Labouesse, E.N.; George, E.L.; Rayburn, H.; Hynes, R.O. Mesodermal development in mouse embryos mutant for fibronectin. Dev. Dyn. 1996, 207, 145–156. [Google Scholar] [CrossRef]
- Goh, K.L.; Yang, J.T.; Hynes, R.O. Mesodermal defects and cranial neural crest apoptosis in α5 integrin-null embryos. Development 1997, 124, 4309–4319. [Google Scholar] [CrossRef] [PubMed]
- Koshida, S.; Kishimoto, Y.; Ustumi, H.; Shimizu, T.; Furutani-Seiki, M.; Kondoh, H.; Takada, S. Integrinα5-dependent fibronectin accumulation for maintenance of somite boundaries in zebrafish embryos. Dev. Cell 2005, 8, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Kragtorp, K.A.; Miller, J.R. Integrin α5 is required for somite rotation and boundary formation in Xenopus. Dev. Dyn. 2007, 236, 2713–2720. [Google Scholar] [CrossRef]
- Rifes, P.; Thorsteinsdóttir, S. Extracellular matrix assembly and 3D organization during paraxial mesoderm development in the chick embryo. Dev. Biol. 2012, 368, 370–381. [Google Scholar] [CrossRef]
- Bénazéraf, B.; Francois, P.; Baker, R.E.; Denans, N.; Little, C.D.; Pourquié, O. A random cell motility gradient downstream of FGF controls elongation of an amniote embryo. Nature 2010, 466, 248–252. [Google Scholar] [CrossRef] [Green Version]
- Lawton, A.K.; Nandi, A.; Stulberg, M.J.; Dray, N.; Sneddon, M.W.; Pontius, W.; Emonet, T.; Holley, S.A. Regulated tissue fluidity steers zebrafish body elongation. Development 2013, 140, 573–582. [Google Scholar] [CrossRef] [Green Version]
- Mongera, A.; Rowghanian, P.; Gustafson, H.J.; Shelton, E.; Kealhofer, D.A.; Carn, E.K.; Serwane, F.; Lucio, A.A.; Giammona, J.; Campàs, O. A fluid-to-solid jamming transition underlies vertebrate body axis elongation. Nature 2018, 561, 401–405. [Google Scholar] [CrossRef]
- Marrese, M.; Antonovaite, N.; Nelemans, B.K.A.; Smit, T.H.; Iannuzzi, D. Micro-indentation and optical coherence tomography for the mechanical characterization of embryos: Experimental setup and measurements on chicken embryos. Acta Biomater. 2019, 97, 524–534. [Google Scholar] [CrossRef] [PubMed]
- Marrese, M.; Antonovaité, N.; Nelemans, B.K.A.; Ahmadzada, A.; Iannuzzi, D.; Smit, T.H. In vivo characterization of chick embryo mesoderm by optical coherence tomography-assisted microindentation. FASEB J. 2020, 34, 12269–12277. [Google Scholar] [CrossRef] [PubMed]
- Jülich, D.; Mould, A.P.; Koper, E.; Holley, S.A. Control of extracellular matrix assembly along tissue boundaries via integrin and Eph/Ephrin signaling. Development 2009, 136, 2913–2921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naganathan, S.R.; Popović, M.; Oates, A.C. Left–right symmetry of zebrafish embryos requires somite surface tension. Nature 2022, 605, 516–521. [Google Scholar] [CrossRef] [PubMed]
- Hamburger, V.; Hamilton, H.L. A series of normal stages in the development of the chick embryo. Dev. Dyn. 1992, 195, 231–272. [Google Scholar] [CrossRef] [PubMed]
- Pourquié, O.; Tam, P.P.L. A nomenclature for prospective somites and phases of cyclic gene expression in the presomitic mesoderm. Dev. Cell 2001, 1, 619–620. [Google Scholar] [CrossRef] [Green Version]
- McKeown-Longo, P.J.; Mosher, D.F. Interaction of the 70,000-mol-wt amino-terminal fragment of fibronectin with the matrix-assembly receptor of fibroblasts. J. Cell Biol. 1985, 100, 364–374. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Nagatoshi, K.; Hamano, A.; Imamura, Y.; Huss, D.; Uchida, S.; Lansford, R. Basal filopodia and vascular mechanical stress organize fibronectin into pillars bridging the mesoderm-endoderm gap. Development 2017, 144, 281–291. [Google Scholar] [CrossRef] [Green Version]
- Huveneers, S.; Truong, H.; Fässler, R.; Sonnenberg, A.; Danen, E.H.J. Binding of soluble fibronectin to integrin α5β1—Link to focal adhesion redistribution and contractile shape. J. Cell Sci. 2008, 121, 2452–2462. [Google Scholar] [CrossRef] [Green Version]
- Pierschbacher, M.D.; Ruoslahti, E. Cell attachment activity of fibronectin can be duplicated by small synthetic fragments of the molecule. Nature 1984, 309, 30–33. [Google Scholar] [CrossRef]
- Yarrow, J.C.; Totsukawa, G.; Charras, G.T.; Mitchison, T.J. Screening for cell migration inhibitors via automated microscopy reveals a Rho-kinase inhibitor. Chem. Biol. 2005, 12, 385–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Straight, A.F.; Cheung, A.; Limouze, J.; Chen, I.; Westwood, N.J.; Sellers, J.R.; Mitchison, T.J. Dissecting temporal and spatial control of cytokinesis with a myosin II inhibitor. Science 2003, 299, 1743–1747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, S.C.; Collignon, J.; Schoenwolf, G.C.; Lumsden, A. Improved method for chick whole-embryo culture using a filter paper carrier. Dev. Dyn. 2001, 220, 284–289. [Google Scholar] [CrossRef]
- Momose, T.; Tonegawa, A.; Takeuchi, J.; Ogawa, H.; Umesono, K.; Yasuda, K. Efficient targeting of gene expression in chick embryos by microelectroporation. Dev. Growth Differ. 1999, 41, 335–344. [Google Scholar] [CrossRef]
- Danen, E.H.J.; Sonneveld, P.; Brakebusch, C.; Fässler, R.; Sonnenberg, A. The fibronectin-binding integrins α5β1 and αvβ3 differentially modulate RhoA-GTP loading, organization of cell matrix adhesions, and fibronectin fibrillogenesis. J. Cell Biol. 2002, 159, 1071–1086. [Google Scholar] [CrossRef] [Green Version]
- de Almeida, P.G.; Pinheiro, G.G.; Nunes, A.M.; Gonçalves, A.B.; Thorsteinsdóttir, S. Fibronectin assembly during early embryo development: A versatile communication system between cells and tissues. Dev. Dyn. 2016, 245, 520–535. [Google Scholar] [CrossRef] [Green Version]
- Prieto, D.; Aparicio, G.; Machado, M.; Zolessi, F.R. Application of the DNA-specific stain methyl green in the fluorescent labeling of embryos. J. Vis. Exp. 2015, 2015, e52769. [Google Scholar] [CrossRef] [Green Version]
- Henrique, D.; Adam, J.; Myat, A.; Chitnis, A.; Lewis, J.; Ish-Horowicz, D. Expression of a Delta homologue in prospective neurons in the chick. Nature 1995, 375, 787–790. [Google Scholar] [CrossRef]
- Buchberger, A.; Seidl, K.; Klein, C.; Eberhardt, H.; Arnold, H.H. cMeso-1, a novel bHLH transcription factor, is involved in somite formation in chicken embryos. Dev. Biol. 1998, 199, 201–215. [Google Scholar] [CrossRef] [Green Version]
- Jouve, C.; Palmeirim, I.; Henrique, D.; Beckers, J.; Gossler, A.; Ish-Horowicz, D.; Pourquié, O. Notch signalling is required for cyclic expression of the hairy-like gene HES1 in the presomitic mesoderm. Development 2000, 127, 1421–1429. [Google Scholar] [CrossRef]
- Preibisch, S.; Saalfeld, S.; Tomancak, P. Globally optimal stitching of tiled 3D microscopic image acquisitions. Bioinformatics 2009, 25, 1463–1465. [Google Scholar] [CrossRef] [PubMed]
- Kocsis, E.; Trus, B.L.; Steer, C.J.; Bisher, M.E.; Steven, A.C. Image averaging of flexible fibrous macromolecules: The clathrin triskelion has an elastic proximal segment. J. Struct. Biol. 1991, 107, 6–14. [Google Scholar] [CrossRef]
- Drake, C.J.; Little, C.D. Integrins play an essential role in somite adhesion to the embryonic axis. Dev. Biol. 1991, 143, 418–421. [Google Scholar] [CrossRef]
- Drake, C.J.; Davis, L.A.; Hungerford, J.E.; Little, C.D. Perturbation of beta 1 integrin-mediated adhesions results in altered somite cell shape and behavior. Dev. Biol. 1992, 149, 327–338. [Google Scholar] [CrossRef]
- Girós, A.; Grgur, K.; Gossler, A.; Costell, M. α5β1 integrin-mediated adhesion to fibronectin is required for axis elongation and somitogenesis in mice. PLoS ONE 2011, 6, e22002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, S.; Leiss, M.; Moser, M.; Ohashi, T.; Kitao, T.; Heckmann, D.; Pfeifer, A.; Kessler, H.; Takagi, J.; Erickson, H.P.; et al. The RGD motif in fibronectin is essential for development but dispensable for fibril assembly. J. Cell Biol. 2007, 178, 167–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubrulle, J.; McGrew, M.J.; Pourquié, O. FGF signaling controls somite boundary position and regulates segmentation clock control of spatiotemporal Hox gene activation. Cell 2001, 106, 219–232. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Yasuda, K.; Takahashi, Y. Morphological boundary forms by a novel inductive event mediated by Lunatic fringe and Notch during somitic segmentation. Development 2002, 129, 3633–3644. [Google Scholar] [CrossRef]
- Palmeirim, I.; Dubrulle, J.; Henrique, D.; Ish-Horowicz, D.; Pourquié, O. Uncoupling segmentation and somitogenesis in the chick presomitic mesoderm. Dev. Genet. 1998, 23, 77–85. [Google Scholar] [CrossRef]
- Furuta, Y.; Ilic, D.; Kanazawa, S.; Takeda, N.; Yamamoto, T.; Aizawa, S. Mesodermal defect in late phase of gastrulation by a targeted mutation of focal adhesion kinase, FAK. Oncogene 1995, 11, 1989–1995. [Google Scholar]
- Kragtorp, K.A.; Miller, J.R. Regulation of somitogenesis by Ena/VASP proteins and FAK during Xenopus development. Development 2006, 133, 685–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, C.A.; Crawford, B.D.; Yan, Y.L.; Postlethwait, J.; Cooper, M.S.; Hille, M.B. Roles for zebrafish focal adhesion kinase in notochord and somite morphogenesis. Dev. Biol. 2001, 240, 474–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newell-Litwa, K.A.; Horwitz, R.; Lamers, M.L. Non-Muscle myosin II in disease: Mechanisms and therapeutic opportunities. DMM Dis. Model. Mech. 2015, 8, 1495–1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, P.; Zaidel-Bar, R. Principles of Actomyosin Regulation In Vivo. Trends Cell Biol. 2019, 29, 150–163. [Google Scholar] [CrossRef]
- Chernoff, E.A.G.; Hilfer, S.R. Calcium dependence and contraction in somite formation. Tissue Cell 1982, 14, 435–449. [Google Scholar] [CrossRef]
- Pasapera, A.M.; Plotnikov, S.V.; Fischer, R.S.; Case, L.B.; Egelhoff, T.T.; Waterman, C.M. Rac1-dependent phosphorylation and focal adhesion recruitment of myosin IIA regulates migration and mechanosensing. Curr. Biol. 2015, 25, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Nakaya, Y.; Kuroda, S.; Katagiri, Y.T.; Kaibuchi, K.; Takahashi, Y. Mesenchymal-epithelial transition during somitic segmentation is regulated by differential roles of Cdc42 and Rac1. Dev. Cell 2004, 7, 425–438. [Google Scholar] [CrossRef] [Green Version]
- Rallis, C.; Pinchin, S.M.; Ish-Horowicz, D. Cell-autonomous integrin control of Wnt and Notch signalling during somitogenesis. Development 2010, 137, 3591–3601. [Google Scholar] [CrossRef] [Green Version]
- Gordon, W.R.; Zimmerman, B.; He, L.; Miles, L.J.; Huang, J.; Tiyanont, K.; McArthur, D.G.; Aster, J.C.; Perrimon, N.; Loparo, J.J.; et al. Mechanical Allostery: Evidence for a Force Requirement in the Proteolytic Activation of Notch. Dev. Cell 2015, 33, 729–736. [Google Scholar] [CrossRef] [Green Version]
- Luca, V.C.; Kim, B.C.; Ge, C.; Kakuda, S.; Wu, D.; Roein-Peikar, M.; Haltiwanger, R.S.; Zhu, C.; Ha, T.; Garcia, K.C. Notch-Jagged complex structure implicates a catch bond in tuning ligand sensitivity. Science 2017, 355, 1320–1324. [Google Scholar] [CrossRef] [Green Version]
- Meloty-Kapella, L.; Shergill, B.; Kuon, J.; Botvinick, E.; Weinmaster, G. Notch Ligand Endocytosis Generates Mechanical Pulling Force Dependent on Dynamin, Epsins, and Actin. Dev. Cell 2012, 22, 1299–1312. [Google Scholar] [CrossRef] [Green Version]
- Moore, R.E.; Clarke, J.; Alexandre, P. Protrusion-Mediated Signaling Regulates Patterning of the Developing Nervous System. Front. Cell Dev. Biol. 2020, 8, 579073. [Google Scholar] [CrossRef] [PubMed]
- Stassen, O.M.J.A.; Ristori, T.; Sahlgren, C.M. Notch in mechanotransduction—From molecular mechanosensitivity to tissue mechanostasis. J. Cell Sci. 2021, 133, jcs250738. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.; Georgiou, M.; Stevenson, N.L.; Miodownik, M.; Baum, B. Dynamic Filopodia Transmit Intermittent Delta-Notch Signaling to Drive Pattern Refinement during Lateral Inhibition. Dev. Cell 2010, 19, 78–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, G.L.; He, L.; Perrimon, N.; Charras, G.; Giniger, E.; Baum, B. A role for actomyosin contractility in Notch signaling. BMC Biol. 2019, 17, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardy, K.M.; Yatskievych, T.A.; Konieczka, J.; Bobbs, A.S.; Antin, P.B. FGF signalling through RAS/MAPK and PI3K pathways regulates cell movement and gene expression in the chicken primitive streak without affecting E-cadherin expression. BMC Dev. Biol. 2011, 11, 20. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Chrisman, H.; Weijer, C.J. PDGF signalling controls the migration of mesoderm cells during chick gastrulation by regulating N-cadherin expression. Development 2008, 135, 3521–3530. [Google Scholar] [CrossRef] [Green Version]
- Horikawa, K.; Ishimatsu, K.; Yoshimoto, E.; Kondo, S.; Takeda, H. Noise-resistant and synchronized oscillation of the segmentation clock. Nature 2006, 441, 719–723. [Google Scholar] [CrossRef]
- Liao, B.K.; Jörg, D.J.; Oates, A.C. Faster embryonic segmentation through elevated Delta-Notch signalling. Nat. Commun. 2016, 7, 11861. [Google Scholar] [CrossRef] [Green Version]
- Herrgen, L.; Ares, S.; Morelli, L.G.; Schröter, C.; Jülicher, F.; Oates, A.C. Intercellular coupling regulates the period of the segmentation clock. Curr. Biol. 2010, 20, 1244–1253. [Google Scholar] [CrossRef] [Green Version]
- Tenin, G.; Wright, D.; Ferjentsik, Z.; Bone, R.; McGrew, M.J.; Maroto, M. The chick somitogenesis oscillator is arrested before all paraxial mesoderm is segmented into somites. BMC Dev. Biol. 2010, 10, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascoal, S.; Carvalho, C.R.; Rodriguez-León, J.; Delfini, M.C.; Duprez, D.; Thorsteinsdóttir, S.; Palmeirim, I. A Molecular Clock Operates During Chick Autopod Proximal-distal Outgrowth. J. Mol. Biol. 2007, 368, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, M.; Yamanaka, Y.; Uemura, M.; Osawa, M.; Saito, M.K.; Nagahashi, A.; Nishio, M.; Guo, L.; Ikegawa, S.; Sakurai, S.; et al. Recapitulating the human segmentation clock with pluripotent stem cells. Nature 2020, 580, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Rayon, T.; Stamataki, D.; Perez-Carrasco, R.; Garcia-Perez, L.; Barrington, C.; Melchionda, M.; Exelby, K.; Lazaro, J.; Tybulewicz, V.L.J.; Fisher, E.M.C.; et al. Species-specific pace of development is associated with differences in protein stability. Science 2020, 369, eaba7667. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, P.; Zaidel-Bar, R. Mechanosensing in embryogenesis. Curr. Opin. Cell Biol. 2021, 68, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Weterings, S.D.C.; van Oostrom, M.J.; Sonnen, K.F. Building bridges between fields: Bringing together development and homeostasis. Development 2021, 148, dev193268. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes de Almeida, P.; Rifes, P.; Martins-Jesus, A.P.; Pinheiro, G.G.; Andrade, R.P.; Thorsteinsdóttir, S. Cell–Fibronectin Interactions and Actomyosin Contractility Regulate the Segmentation Clock and Spatio-Temporal Somite Cleft Formation during Chick Embryo Somitogenesis. Cells 2022, 11, 2003. https://doi.org/10.3390/cells11132003
Gomes de Almeida P, Rifes P, Martins-Jesus AP, Pinheiro GG, Andrade RP, Thorsteinsdóttir S. Cell–Fibronectin Interactions and Actomyosin Contractility Regulate the Segmentation Clock and Spatio-Temporal Somite Cleft Formation during Chick Embryo Somitogenesis. Cells. 2022; 11(13):2003. https://doi.org/10.3390/cells11132003
Chicago/Turabian StyleGomes de Almeida, Patrícia, Pedro Rifes, Ana P. Martins-Jesus, Gonçalo G. Pinheiro, Raquel P. Andrade, and Sólveig Thorsteinsdóttir. 2022. "Cell–Fibronectin Interactions and Actomyosin Contractility Regulate the Segmentation Clock and Spatio-Temporal Somite Cleft Formation during Chick Embryo Somitogenesis" Cells 11, no. 13: 2003. https://doi.org/10.3390/cells11132003
APA StyleGomes de Almeida, P., Rifes, P., Martins-Jesus, A. P., Pinheiro, G. G., Andrade, R. P., & Thorsteinsdóttir, S. (2022). Cell–Fibronectin Interactions and Actomyosin Contractility Regulate the Segmentation Clock and Spatio-Temporal Somite Cleft Formation during Chick Embryo Somitogenesis. Cells, 11(13), 2003. https://doi.org/10.3390/cells11132003