
Ancestral Sperm Ecotypes Reveal Multiple Invasions of a Non-Native Fish in Northern Europe

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
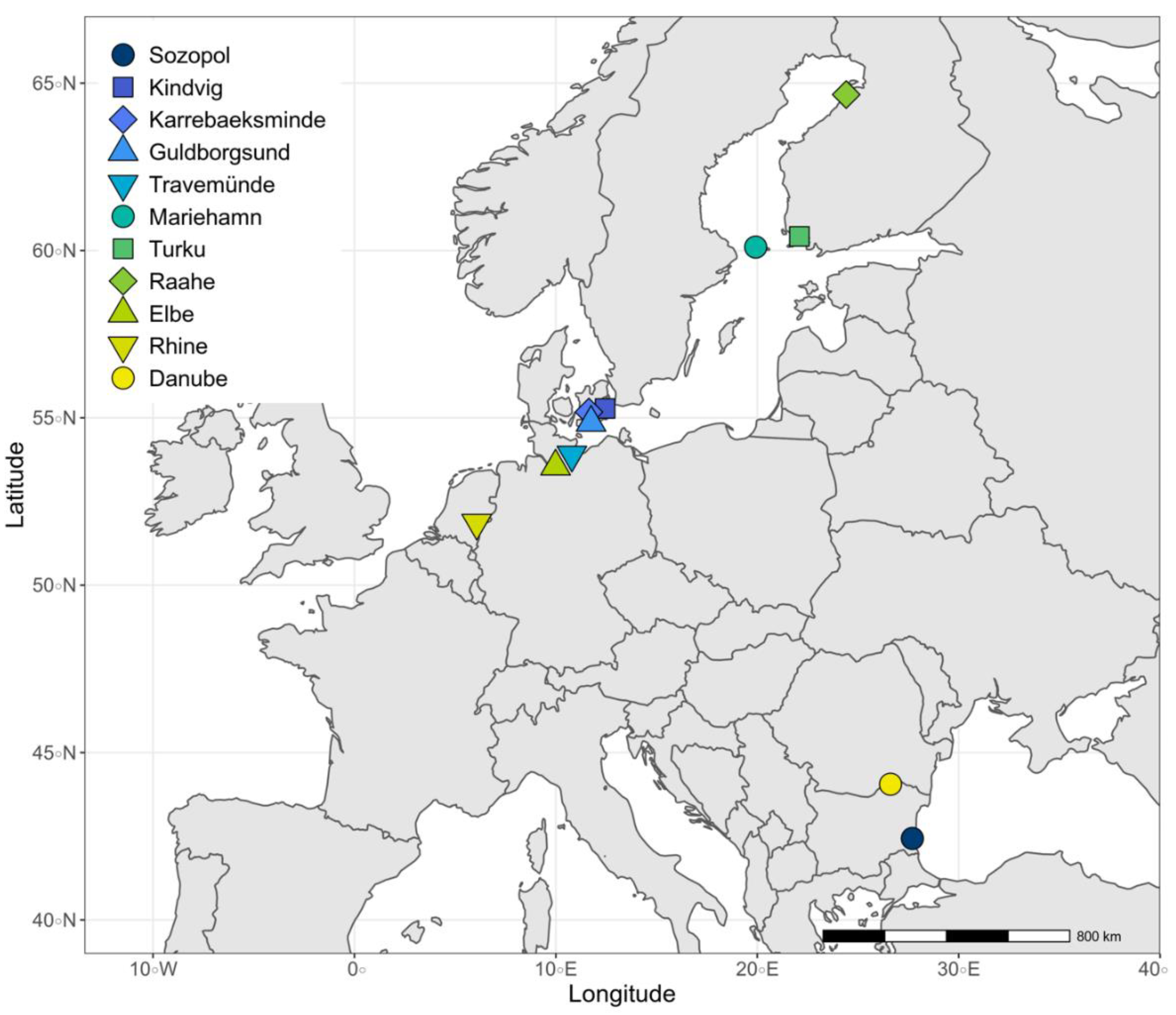
2.1. Sampling Sites in the Ancestral Range
2.2. Sampling of Sperm and Reproductive Tissues from the Ancestral Range
2.3. Compiling Sperm Data from the Introduced Range
2.4. DNA Sampling and GBS Library Preparation
2.5. Sequencing and SNP Calling
2.6. Analysis of Sperm Traits and Reproductive Tissues from the Ancestral Range
2.7. Analysis of Sperm Traits between the Ancestral and Introduced Ranges
2.8. Genotype Analyses
2.9. Shipping Intensity
3. Results
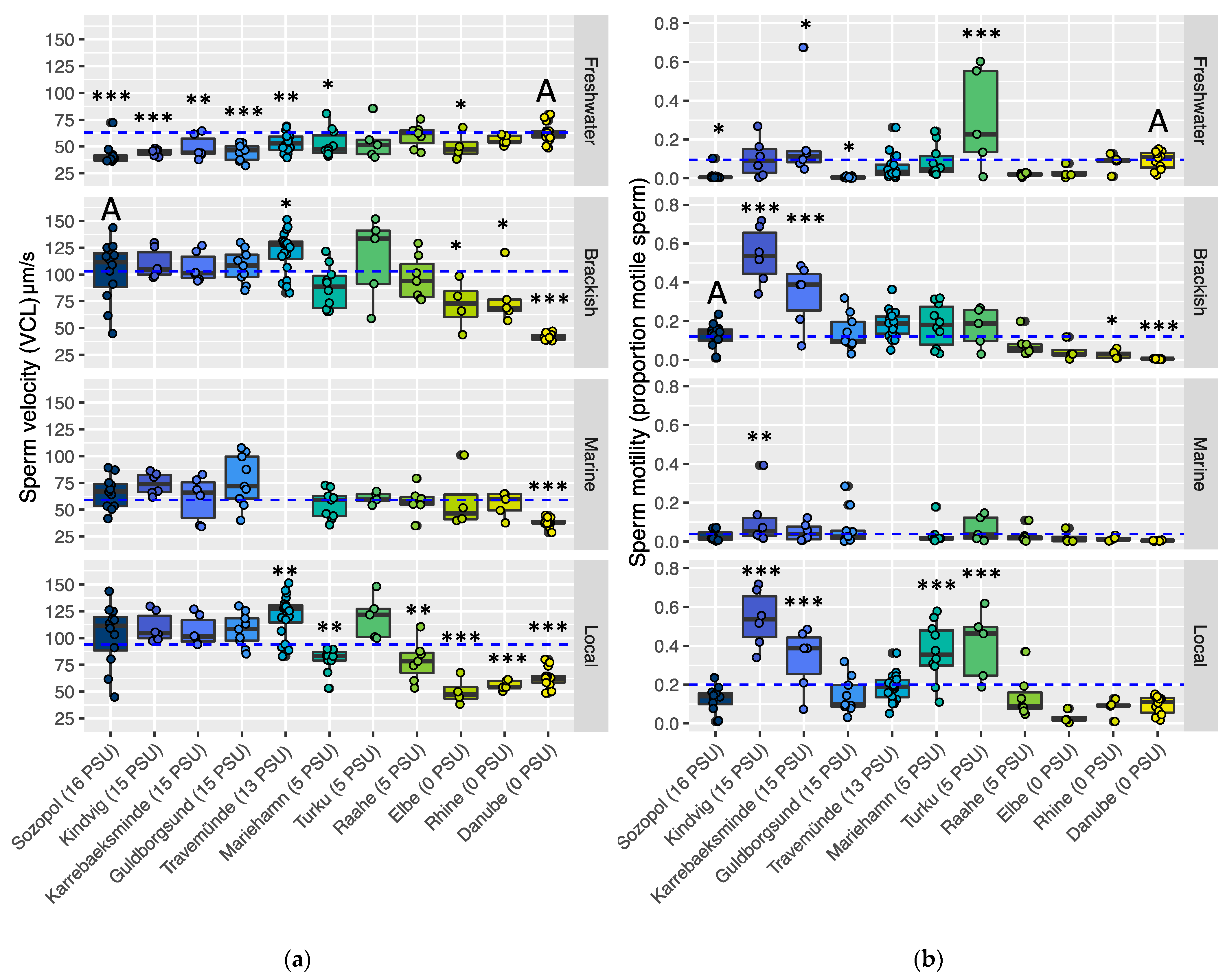
3.1. Sperm Performance in the Ancestral Region
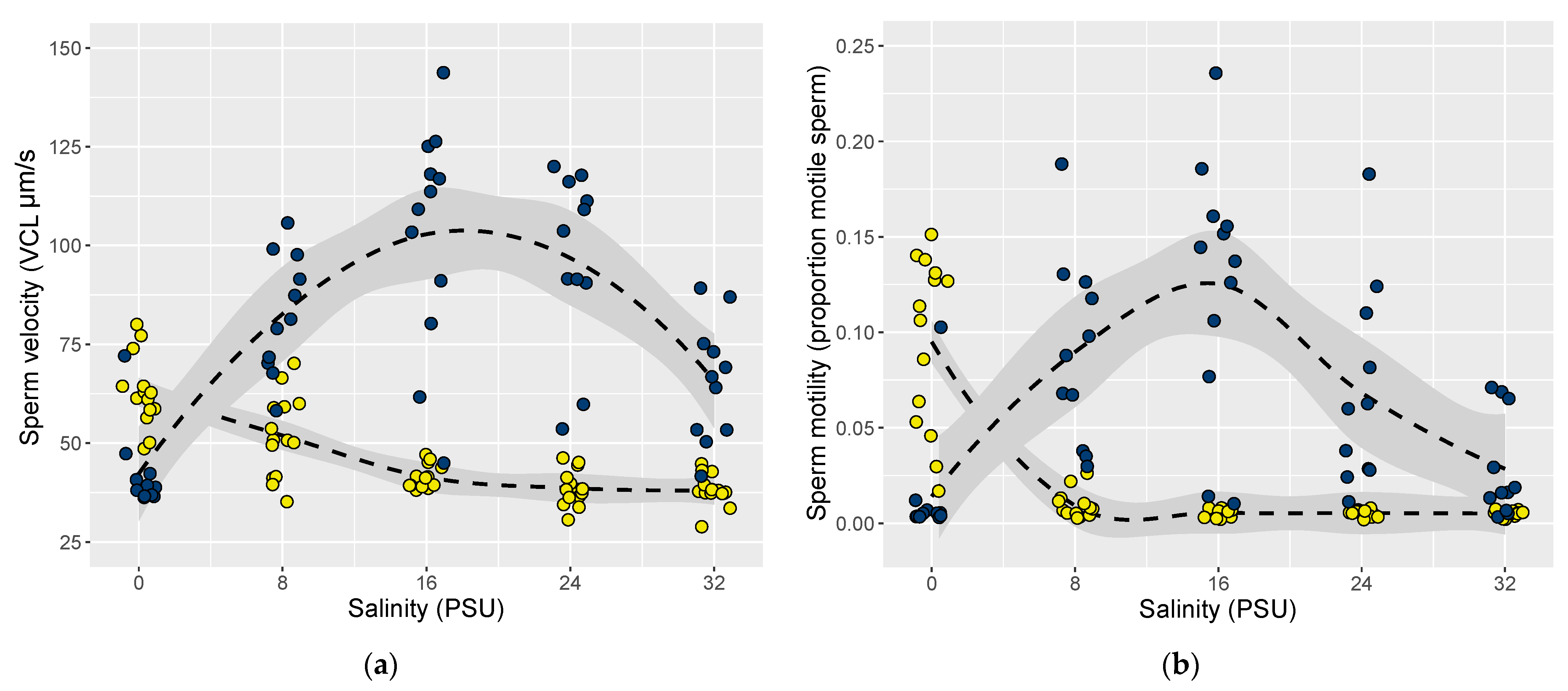
3.2. Comparing Sperm Performance between Ancestral and Introduced Sites
3.2.1. Sperm in Freshwater Conditions (0–1 PSU)
3.2.2. Sperm in Brackish Conditions (15–16 PSU)
3.2.3. Sperm in Marine Conditions (30–32 PSU)
3.2.4. Sperm Performance at Each Site’s Local Salinity
3.3. Differences in Reproductive Tactics in the Ancestral Region
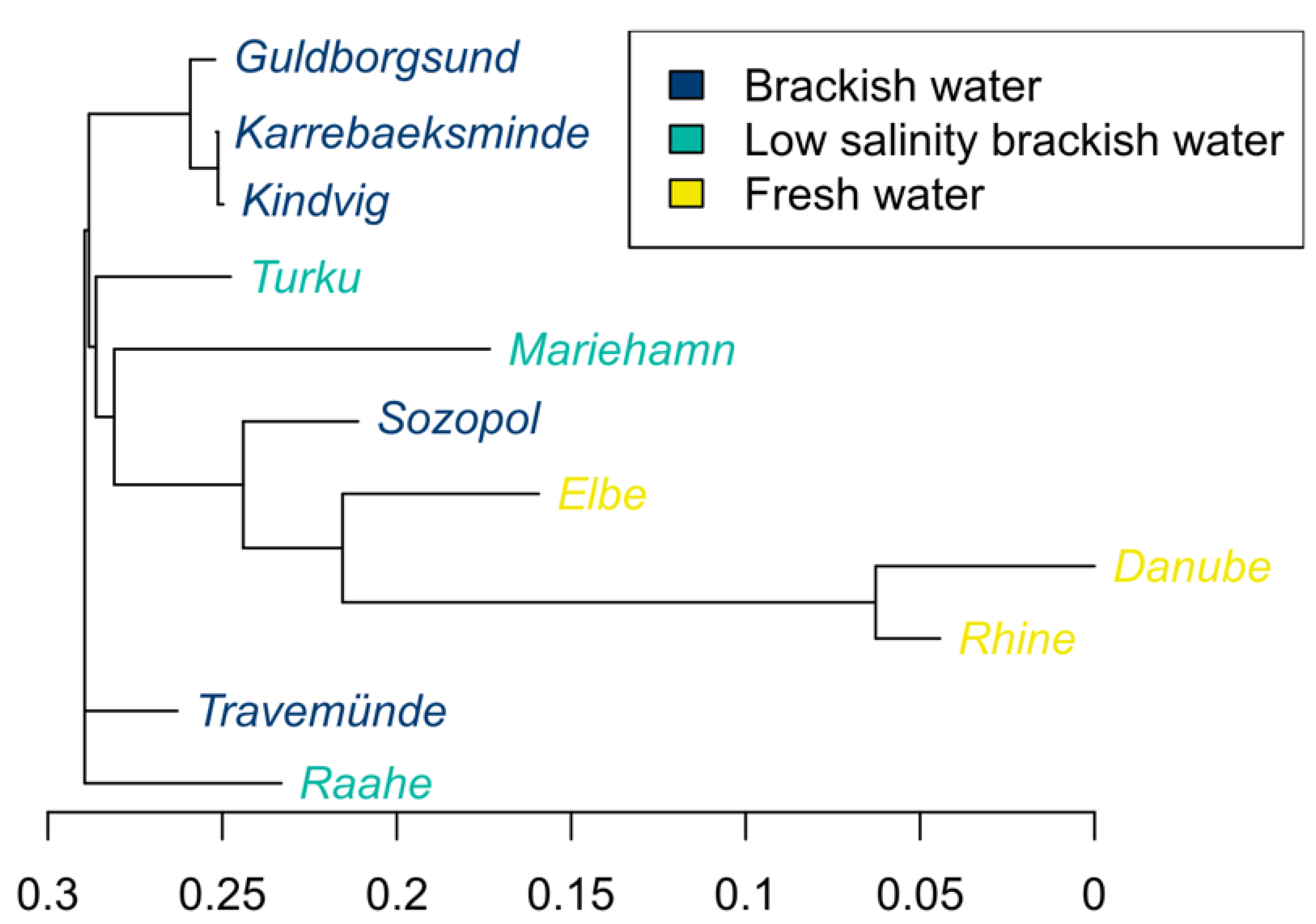
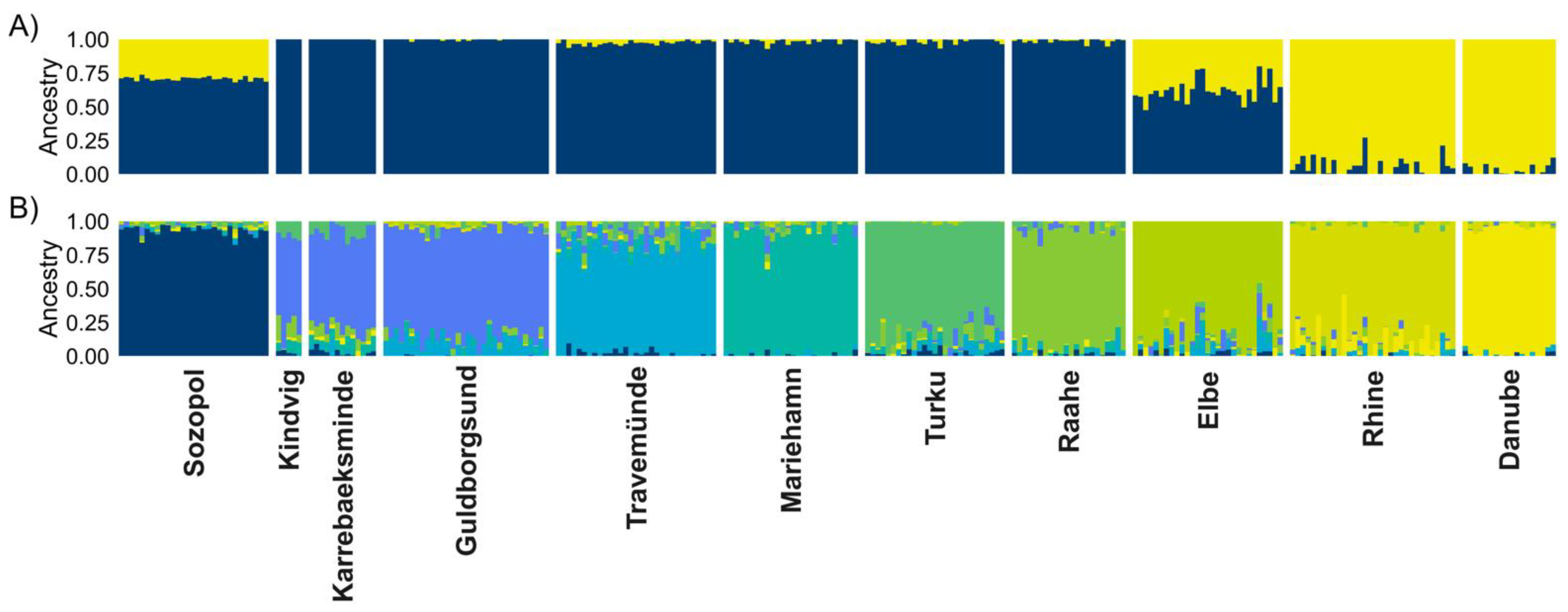
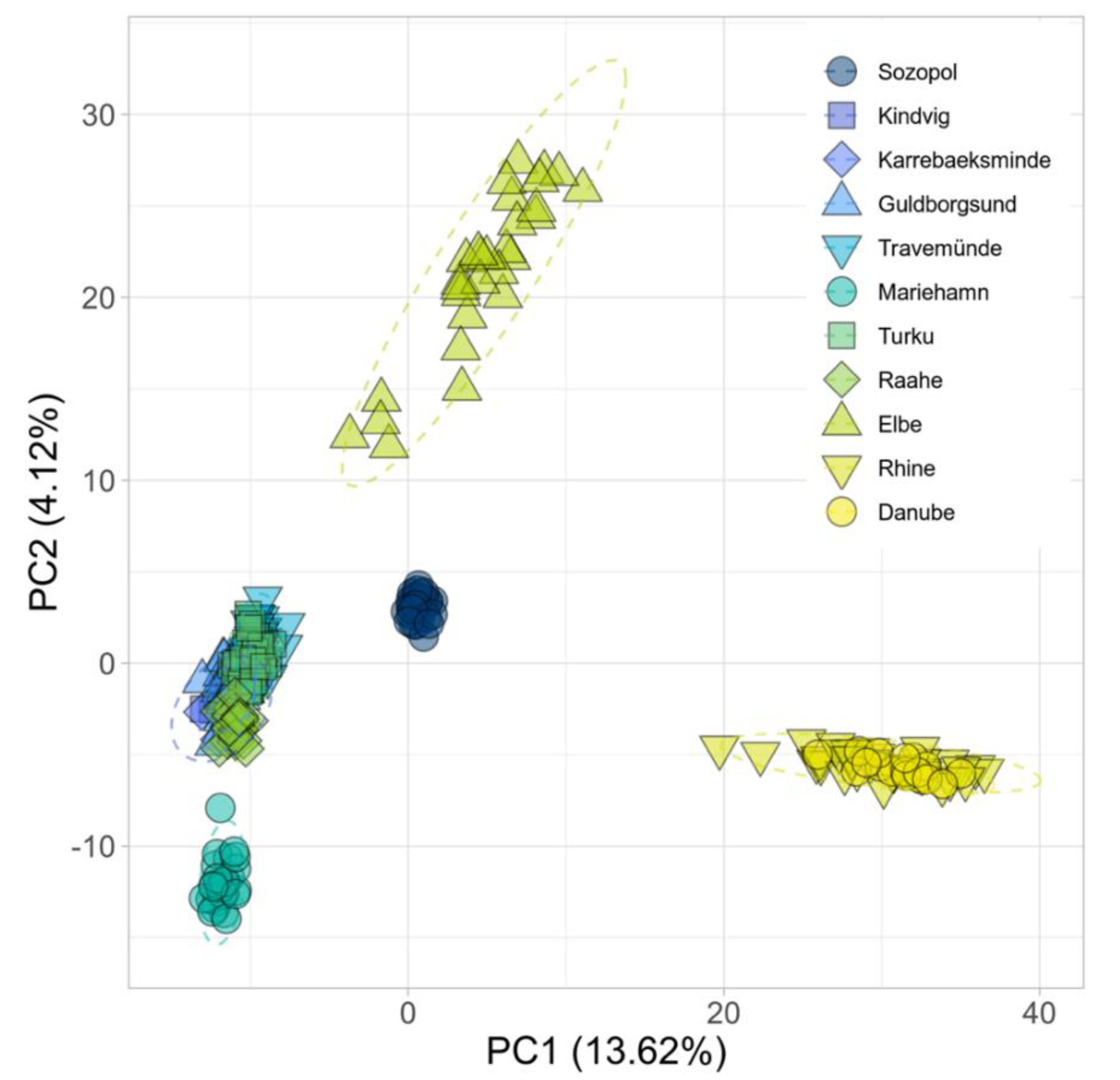
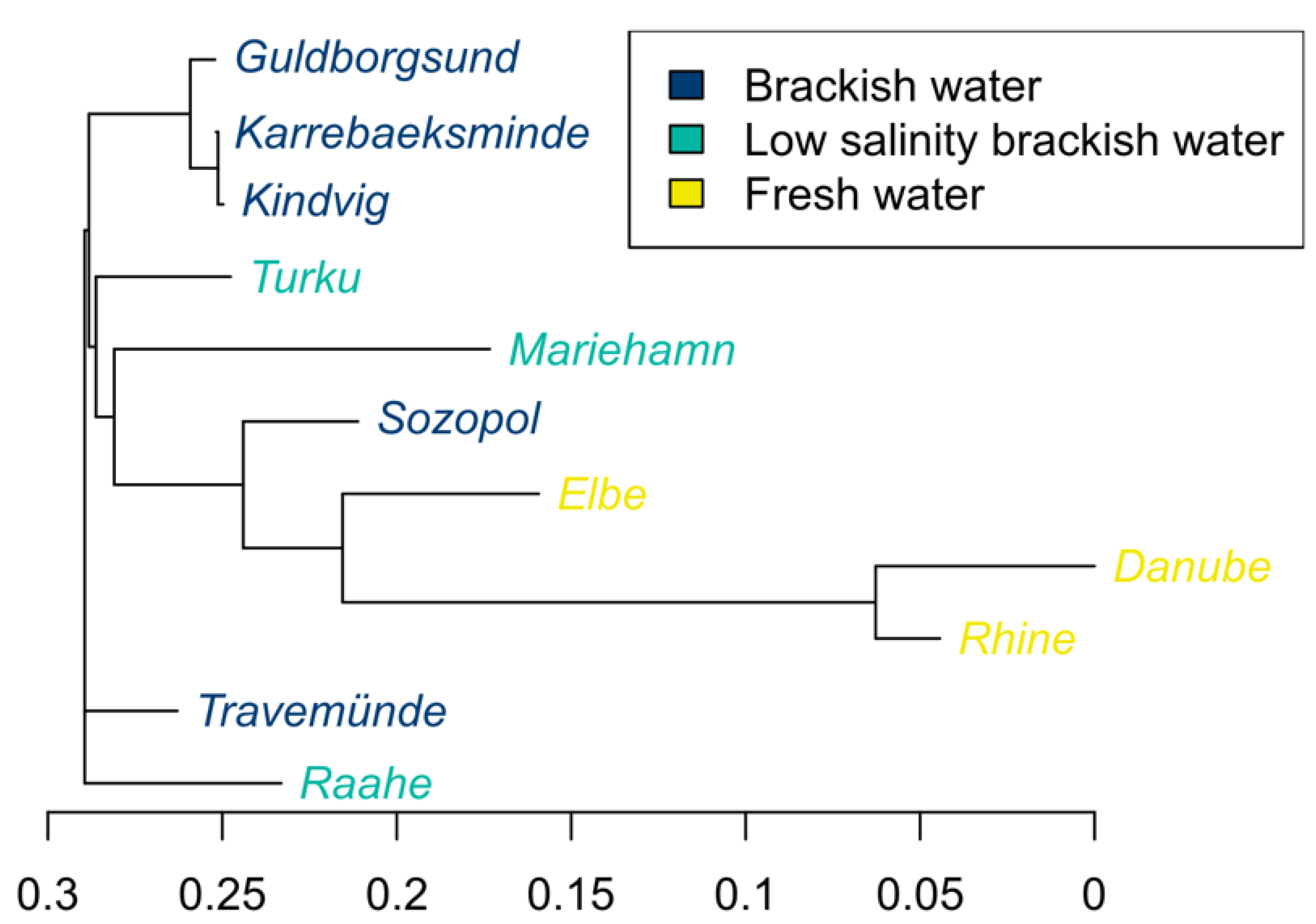
3.4. Population Genetic Structures between Ancestral and Introduced Sites
4. Discussion
4.1. Summary
4.2. Separate Ancestral Sperm Ecotypes Fuel the Round Goby Invasion
4.3. Individual Sites Provide Details of Eco-Evolutionary Dynamics and Ongoing Local Adaptation
4.4. Sperm Motility, Relaxed Selection and Plasticity
4.5. Implications for Management
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parker, G.A. Sperm competition and its evolutionary consequences in the insects. Biol. Rev. 1970, 45, 525–567. [Google Scholar] [CrossRef]
- Birkhead, T.R.; Hosken, D.J.; Pitnick, S. Sperm Biology: An Evolutionary Perspective; Elsevier: Amsterdam, The Netherlands; London, UK, 2009. [Google Scholar]
- Parker, G.A. Conceptual developments in sperm competition: A very brief synopsis. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 120–124. [Google Scholar] [CrossRef]
- Simmons, L.W.; Wedell, N. Fifty years of sperm competition: The structure of a scientific revolution. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375. [Google Scholar] [CrossRef]
- Parker, G.A. The origin and evolution of gamete dimorphism and the male-female phenomenon. J. Theor. Biol. 1972, 36, 529–553. [Google Scholar] [CrossRef]
- Parker, G.A. The sexual cascade and the rise of pre-ejaculatory (darwinian) sexual selection, sex roles, and sexual conflict. Cold Spring Harb. Perspect. Biol. 2014, 6, a017509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stockley, P.; Gage, M.J.G.; Parker, G.A.; Møller, A.P. Sperm competition in fishes: The evolution of testis size and ejaculate characteristics. Am. Nat. 1997, 149, 933–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitnick, S.; Hosken, D.J.; Birkhead, T.R. Sperm morphological diversity. In Sperm Biology: An Evolutionary Perspective; Birkhead, T.R., Hosken, D.J., Pitnick, S., Eds.; Academic Press: Oxford, UK, 2009; pp. 69–149. [Google Scholar]
- Pizzari, T.; Parker, G.A. Sperm competition and sperm phenotype. In Sperm Biology: An Evolutionary Perspective; Birkhead, T.R., Hosken, D.J., Pitnick, S., Eds.; Academic Press: Oxford, UK, 2009; pp. 207–245. ISBN 9780123725684. [Google Scholar]
- Birkhead, T.R.; Møller, A.P. Sperm Competition and Sexual Selection, 1st ed.; Birkhead, T.R., Møller, A.P., Eds.; Academic Press: London, UK, 1998. [Google Scholar]
- Birkhead, T.R.; Pizzari, T. Postcopulatory sexual selection. Nat. Rev. Genet. 2002, 3, 262–273. [Google Scholar] [CrossRef] [PubMed]
- Lüpold, S.; De Boer, R.A.; Evans, J.P.; Tomkins, J.L.; Fitzpatrick, J.L. How sperm competition shapes the evolution of testes and sperm: A meta-analysis. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20200064. [Google Scholar] [CrossRef] [PubMed]
- Vicens, A.; Lüke, L.; Roldan, E.R.S. Proteins involved in motility and sperm-egg interaction evolve more rapidly in mouse spermatozoa. PLoS ONE 2014, 9, e91302. [Google Scholar] [CrossRef] [Green Version]
- Beirão, J.; Lewis, J.A.; Wringe, B.F.; Purchase, C.F. A novel sperm adaptation to evolutionary constraints on reproduction: Pre-ejaculatory sperm activation in the beach spawning capelin (Osmeridae). Ecol. Evol. 2018, 8, 2343–2349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purchase, C.F. Low tolerance of salt water in a marine fish: New and historical evidence for surprising local adaption in the well-studied commercially exploited capelin. Can. J. Fish. Aquat. Sci. 2017, 75, 673–681. [Google Scholar] [CrossRef]
- Svensson, O.; Gräns, J.; Celander, M.C.; Havenhand, J.; Leder, E.H.; Lindström, K.; Schöld, S.; van Oosterhout, C.; Kvarnemo, C. Immigrant reproductive dysfunction facilitates ecological speciation. Evolution 2017, 71, 2510–2521. [Google Scholar] [CrossRef] [PubMed]
- Momigliano, P.; Jokinen, H.; Fraimout, A.; Florin, A.-B.; Norkko, A.; Merilä, J. Extraordinarily rapid speciation in a marine fish. Proc. Natl. Acad. Sci. USA 2017, 114, 201615109. [Google Scholar] [CrossRef] [Green Version]
- Dahlke, F.T.; Wohlrab, S.; Butzin, M.; Pörtner, H.O. Thermal bottlenecks in the life cycle define climate vulnerability of fish. Science 2020, 369, 65–70. [Google Scholar] [CrossRef]
- Patzner, R.A. Reproductive strategies of fish. In Fish Reproduction; Rocha, M.J., Ed.; CRC Press: Boca Raton, FL, USA, 2008; p. 40. ISBN 9780429094163. [Google Scholar]
- Liao, W.B.; Huang, Y.; Zeng, Y.; Zhong, M.J.; Luo, Y.; Lüpold, S. Ejaculate evolution in external fertilizers: Influenced by sperm competition or sperm limitation? Evolution 2018, 72, 4–17. [Google Scholar] [CrossRef] [Green Version]
- Fitzpatrick, J.L. Sperm competition and fertilization mode in fishes. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20200074. [Google Scholar] [CrossRef] [PubMed]
- Alavi, S.M.H.; Cosson, J. Sperm motility in fishes. (II) Effects of ions and osmolality: A review. Cell Biol. Int. 2006, 30, 1–14. [Google Scholar] [CrossRef]
- Browne, R.K.; Kaurova, S.A.; Uteshev, V.K.; Shishova, N.V.; McGinnity, D.; Figiel, C.R.; Mansour, N.; Agnew, D.; Wu, M.; Gakhova, E.N.; et al. Sperm motility of externally fertilizing fish and amphibians. Theriogenology 2015, 83, 1–13. [Google Scholar] [CrossRef]
- Taugbøl, A.; Mazzarella, A.B.; Cramer, E.R.A.; Laskemoen, T. Salinity-induced phenotypic plasticity in threespine stickleback sperm activation. Biol. Lett. 2017, 13, 20170516. [Google Scholar] [CrossRef] [PubMed]
- Harvey, B.; Kelley, R.N. Control of spermatozoan motility in a euryhaline teleost acclimated to different salinities. Can. J. Zool. 1984, 62, 2674–2677. [Google Scholar] [CrossRef]
- Palmer, R.E.; Able, K.W. Effect of acclimation salinity on fertilization success in the mummichog, Fundulus heteroclitus. Physiol. Biochem. Zool. 1987, 60, 614–621. [Google Scholar] [CrossRef]
- Ascunce, M.S.; Yang, C.C.; Oakey, J.; Calcaterra, L.; Wu, W.J.; Shih, C.J.; Goudet, J.; Ross, K.G.; Shoemaker, D.W. Global invasion history of the fire ant Solenopsis invicta. Science 2011, 331, 1066–1068. [Google Scholar] [CrossRef] [PubMed]
- Haska, C.L.; Yarish, C.; Kraemer, G.; Blaschik, N.; Whitlatch, R.; Zhang, H.; Lin, S. Bait worm packaging as a potential vector of invasive species. Biol. Invasions 2012, 14, 481–493. [Google Scholar] [CrossRef]
- Kelly, D.W.; Paterson, R.A.; Townsend, C.R.; Poulin, R.; Tompkins, D.M. Parasite spillback: A neglected concept in invasion ecology? Ecology 2009, 90, 2047–2056. [Google Scholar] [CrossRef] [PubMed]
- Pallas, P.S. Zoographia Rosso-Asiatica: Sistens Omnium Animalium in Extenso Imperio Rossico, et Adjacentibus Maribus Observatorum Recensionem, Domicilia, Mores et Descriptiones, Anatomen Atque Icones Plurimorum; Acadamiae Scientiarum Impress: Sankt-Petersburg, Russia, 1831. [Google Scholar]
- Azour, F.; van Deurs, M.; Behrens, J.; Carl, H.; Hüssy, K.; Greisen, K.; Ebert, R.; Møller, P. Invasion rate and population characteristics of the round goby Neogobius melanostomus: Effects of density and invasion history. Aquat. Biol. 2015, 24, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, P.E.; N’Guyen, A.; Adrian-Kalchhauser, I.; Burkhardt-Holm, P. What do we really know about the impacts of one of the 100 worst invaders in Europe? A reality check. Ambio 2016, 45, 267–279. [Google Scholar] [CrossRef] [Green Version]
- Puntila, R.; Strake, S.; Florin, A.B.; Naddafi, R.; Lehtiniemi, M.; Behrens, J.W.; Kotta, J.; Oesterwind, D.; Putnis, I.; Ojaveer, H. Abundance and Distribution of Round Goby (Neogobius melanostromus): HELCOM Baltic Sea Environment Fact. Sheet 2018; HELCOM. 2018. Available online: http://www.helcom.fi/baltic-sea-trends/environment-fact-sheets/biodiversity/abundance-and-distribution-of-round-goby/ (accessed on 6 July 2021).
- Behrens, J.W.; Van Deurs, M.; Christensen, E.A.F. Evaluating dispersal potential of an invasive fish by the use of aerobic scope and osmoregulation capacity. PLoS ONE 2017, 12, e0176038. [Google Scholar] [CrossRef] [Green Version]
- Green, L.; Niemax, J.; Herrmann, J.P.; Temming, A.; Kvarnemo, C. Alternative reproductive tactics are associated with sperm performance in invasive round goby from two different salinity environments. Ecol. Evol. 2020, 10, 9981–9999. [Google Scholar] [CrossRef]
- Parker, G.A. Sperm competition games: Sneaks and extra-pair copulations. Proc. R. Soc. B Biol. Sci. 1990, 242, 127–133. [Google Scholar] [CrossRef]
- Parker, G.A. The evolution of expenditure on testes. J. Zool. 2016, 298, 3–19. [Google Scholar] [CrossRef]
- Green, L.; Niemax, J.; Herrmann, J.P.; Temming, A.; Behrens, J.W.; Havenhand, J.N.; Leder, E.; Kvarnemo, C. Sperm performance limits the reproduction of an invasive fish in novel salinities. Divers. Distrib. 2021, 27, 1091–1105. [Google Scholar] [CrossRef]
- Green, L.; Havenhand, J.N.; Kvarnemo, C. Evidence of rapid adaptive trait change to local salinity in the sperm of an invasive fish. Evol. Appl. 2020, 13, 533–544. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.E.; Stepien, C.A. Invasion genetics of the Eurasian round goby in North America: Tracing sources and spread patterns. Mol. Ecol. 2009, 18, 64–79. [Google Scholar] [CrossRef]
- Brown, J.E.; Stepien, C.A. Ancient divisions, recent expansions: Phylogeography and population genetics of the round goby Apollonia melanostoma. Mol. Ecol. 2008, 17, 2598–2615. [Google Scholar] [CrossRef]
- Marentette, J.R.; Fitzpatrick, J.L.; Berger, R.G.; Balshine, S. Multiple male reproductive morphs in the invasive round goby (Apollonia melanostoma). J. Great Lakes Res. 2009, 35, 302–308. [Google Scholar] [CrossRef]
- Vassilev, M.; Apostolou, A.; Velkov, B.; Dobrev, D.; Zarev, V. Atlas of the Gobies (Gobiidae) in Bulgaria, 1st ed.; Institute of Biodiversity and Ecosystem Research, Bulgarian Academy of Sciences: Sofia, Bulgaria, 2012; Volume 1, ISBN 9789549746297. [Google Scholar]
- Skora, K.E.; Stolarski, J. New fish species in the Gulf of Gdansk, Neogobius sp. [cf. Neogobius melanostomus (Pallas 1811)]. Bull. Sea Fish. Inst. 1993, 1, 83. [Google Scholar]
- Karila, P.; Jensen, J.; Holmgren, S. Galanin-like immunoreactivity in extrinsic and intrinsic nerves to the gut of the Atlantic cod, Gadus morhua, and the effect of galanin on the smooth muscle of the gut. Cell Tissue Res. 1993, 271, 537–544. [Google Scholar] [CrossRef]
- Havenhand, J.N.; Schlegel, P. Near-future levels of ocean acidification do not affect sperm motility and fertilization kinetics in the oyster Crassostrea gigas. Biogeosci. Discuss. 2009, 6, 4573–4586. [Google Scholar] [CrossRef]
- Wilson-Leedy, J.G.; Ingermann, R.L. Development of a novel CASA system based on open source software for characterization of zebrafish sperm motility parameters. Theriogenology 2007, 67, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A Robust, Simple Genotyping-by-Sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leder, E.H.; André, C.; Le Moan, A.; Töpel, M.; Blomberg, A.; Havenhand, J.N.; Lindström, K.; Volckaert, F.A.M.; Kvarnemo, C.; Johannesson, K.; et al. Post-glacial establishment of locally adapted fish populations over a steep salinity gradient. J. Evol. Biol. 2021, 34, 138–156. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high throughput sequencing reads. EBMnet. J. 2011, 17, 10–11. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011, 27, 2987–2993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; De Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.; Friendly, M.; Weisberg, S. Hypothesis tests for multivariate linear models using the car package. R J. 2013, 5, 39–52. [Google Scholar] [CrossRef] [Green Version]
- Keenan, K.; Mcginnity, P.; Cross, T.F.; Crozier, W.W.; Prodöhl, P.A. DiveRsity: An R package for the estimation and exploration of population genetics parameters and their associated errors. Methods Ecol. Evol. 2013, 4, 782–788. [Google Scholar] [CrossRef] [Green Version]
- Weir, B.S.; Cockerham, C.C. Estimating F-statistics for the analysis of population structure. Evolution 1984, 38, 1358–1370. [Google Scholar]
- Louis, E.J.; Dempster, E.R. An Exact Test for Hardy-Weinberg and multiple alleles. Biometrics 1987, 43, 805–811. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Frichot, E.; François, O. LEA: An R package for landscape and ecological association studies. Methods Ecol. Evol. 2015, 6, 925–929. [Google Scholar] [CrossRef]
- Chessel, D.; Dufour, A.B.; Thioulouse, J. The ade4 package—I: One-table methods. R News 2004, 4, 5–10. [Google Scholar]
- Dray, S.; Dufour, A.-B.; Chessel, D. The ade4 package—II: Two-table and K-table methods. R News 2007, 7, 47–52. [Google Scholar]
- Jombart, T. Adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jombart, T.; Ahmed, I. adegenet 1.3-1: New tools for the analysis of genome-wide SNP data. Bioinformatics 2011, 27, 3070–3071. [Google Scholar] [CrossRef] [Green Version]
- Kornis, M.S.; Sharma, S.; Jake Vander Zanden, M. Invasion success and impact of an invasive fish, round goby, in Great Lakes tributaries. Divers. Distrib. 2013, 19, 184–198. [Google Scholar] [CrossRef]
- Ohayon, J.L.; Stepien, C.A. Genetic and biogeographic relationships of the racer goby Neogobius gymnotrachelus (Gobiidae: Teleostei) from introduced and native Eurasian locations. J. Fish. Biol. 2007, 71, 360–370. [Google Scholar] [CrossRef]
- Hughes, A.R.; Inouye, B.D.; Johnson, M.T.J.; Underwood, N.; Vellend, M. Ecological consequences of genetic diversity. Ecol. Lett. 2008, 11, 609–623. [Google Scholar] [CrossRef]
- Gingerich, P.D. Quantification and comparison of evolutionary rates. Am. J. Sci. 1993, 293, 453–478. [Google Scholar] [CrossRef]
- Rius, M.; Darling, J.A. How important is intraspecific genetic admixture to the success of colonising populations? Trends Ecol. Evol. 2014, 29, 233–242. [Google Scholar] [CrossRef]
- Allendorf, F.W.; Luikart, G.H.; Aitken, S.N. Conservation and the Genetics of Populations, 2nd ed.; Wiley-Blackwell: Oxford, UK, 2012. [Google Scholar]
- Kotta, J.; Nurkse, K.; Puntila, R.; Ojaveer, H. Shipping and natural environmental conditions determine the distribution of the invasive non-indigenous round goby Neogobius melanostomus in a regional sea. Estuar. Coast. Shelf Sci. 2016, 169, 15–24. [Google Scholar] [CrossRef]
- Gage, M.J.G.; Macfarlane, C.P.; Yeates, S.; Ward, R.G.; Searle, J.B.; Parker, G.A. Spermatozoal traits and sperm competition in Atlantic salmon: Relative sperm velocity is the primary determinant of fertilization success. Curr. Biol. 2004, 14, 44–47. [Google Scholar] [CrossRef]
- Gasparini, C.; Simmons, L.W.; Beveridge, M.; Evans, J.P. Sperm swimming velocity predicts competitive fertilization success in the green swordtail Xiphophorus helleri. PLoS ONE 2010, 5, e12146. [Google Scholar] [CrossRef] [PubMed]
- Rudolfsen, G.; Figenschou, L.; Folstad, I.; Kleven, O. Sperm velocity influence paternity in the Atlantic cod (Gadus morhua L.). Aquac. Res. 2008, 39, 212–216. [Google Scholar] [CrossRef]
- Dewsbury, D.A. Ejaculate cost and male choice. Am. Nat. 1982, 119, 601–610. [Google Scholar] [CrossRef]
- Losdat, S.; Richner, H.; Blount, J.D.; Helfenstein, F. Immune activation reduces sperm quality in the great tit. PLoS ONE 2011, 6, e22221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kekäläinen, J.; Pirhonen, J.; Taskinen, J. Do highly ornamented and less parasitized males have high quality sperm?—An experimental test for parasite-induced reproductive trade-offs in European minnow (Phoxinus phoxinus). Ecol. Evol. 2014, 4, 4237–4246. [Google Scholar] [CrossRef] [PubMed]
- Dowling, D.K.; Simmons, L.W. Ejaculate economics: Testing the effects of male sexual history on the trade-off between sperm and immune function in Australian crickets. PLoS ONE 2012, 7, e30172. [Google Scholar] [CrossRef] [PubMed]
- Levitan, D.R. Sperm Limitation, Gamete Competition and Sexual Selection in External Fertilizers. In Sperm Competition and Sexual Selection; Birkhead, T.R., Møller, A.P., Eds.; Academic Press: San Diego, CA, USA, 1998; pp. 175–217. [Google Scholar]
- Yoneda, M.; Wright, P.J. Effect of temperature and food availability on reproductive investment of first-time spawning male Atlantic cod, Gadus morhua. ICES J. Mar. Sci. 2005, 62, 1387–1393. [Google Scholar] [CrossRef]
- Kaiser, S.A.; Scott Sillett, T.; Risk, B.B.; Webster, M.S. Experimental food supplementation reveals habitat-dependent male reproductive investment in a migratory bird. Proc. R. Soc. B Biol. Sci. 2015, 282, 20142523. [Google Scholar] [CrossRef] [PubMed]
- Borg, Å.A.; Forsgren, E.; Magnhagen, C. Plastic sex-roles in the common goby—The effect of nest availability. Oikos 2002, 98, 105–115. [Google Scholar] [CrossRef]
- Levitan, D.R. Influence of body size and population density on fertilization success and reproductive output in a free-spawning invertebrate. Biol. Bull. 1991, 181, 261–268. [Google Scholar] [CrossRef]
- Rejmánek, M.; Richardson, D.M. What Attributes Make Some Plant Species More Invasive? Ecology 1996, 77, 1655–1661. [Google Scholar] [CrossRef]
- Jeschke, J.M. General hypotheses in invasion ecology. Divers. Distrib. 2014, 20, 1229–1234. [Google Scholar] [CrossRef]
- Colautti, R.I.; Ricciardi, A.; Grigorovich, I.A.; MacIsaac, H.J. Is invasion success explained by the enemy release hypothesis? Ecol. Lett. 2004, 7, 721–733. [Google Scholar] [CrossRef]
- Kornis, M.S.; Mercado-Silva, N.; van der Zanden, M.J. Twenty years of invasion: A review of round goby Neogobius melanostomus biology, spread and ecological implications. J. Fish. Biol. 2012, 80, 235–285. [Google Scholar] [CrossRef]
- Snoeijs-Leijonmalm, P.; Schubert, H.; Radziejewska, T. Biological Oceanography of the Baltic Sea; Springer International Publishing: Dordrecht, The Netherlands, 2016; ISBN 9789400706675. [Google Scholar]
- Yankova, M.; Pavlov, D.; Ivanova, P.; Karpova, E.; Boltachev, A.; Öztürk, B.; Bat, L.; Oral, M.; Mgeladze, M. Marine fishes in the Black Sea: Recent conservation status. Mediterr. Mar. Sci. 2014, 15, 366–379. [Google Scholar] [CrossRef] [Green Version]
- Puntila-Dodd, R.; Bekkevold, D.; Behrens, J.W. Estimating salinity stress via hsp70 expression in the invasive round goby (Neogobius melanostomus): Implications for further range expansion. Hydrobiologia 2021, 848, 421–429. [Google Scholar] [CrossRef]
- Ellis, S.; MacIsaac, H.J. Salinity tolerance of Great Lakes invaders. Freshw. Biol. 2009, 54, 77–89. [Google Scholar] [CrossRef]
- Crawford, K.M.; Whitney, K.D. Population genetic diversity influences colonization success. Mol. Ecol. 2010, 19, 1253–1263. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.C.; Ruiz-Navarro, A.; Britton, J.R. Population density modifies the ecological impacts of invasive species. Oikos 2015, 124, 880–887. [Google Scholar] [CrossRef] [Green Version]
- Laikre, L.; Hoban, S.; Bruford, M.W.; Segelbacher, G.; Allendorf, F.W.; Gajardo, G.; González Rodríguez, A.; Hedrick, P.W.; Heuertz, M.; Hohenlohe, P.A.; et al. Post-2020 goals overlook genetic diversity. Science 2020, 367, 1083–1085. [Google Scholar] [CrossRef] [Green Version]
- Sandström, A.; Lundmark, C.; Andersson, K.; Johannesson, K.; Laikre, L. Understanding and bridging the conservation-genetics gap in marine conservation. Conserv. Biol. 2019, 33, 725–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wennerström, L.; Laikre, L.; Ryman, N.; Utter, F.M.; Ab Ghani, N.I.; André, C.; DeFaveri, J.; Johansson, D.; Kautsky, L.; Merilä, J.; et al. Genetic biodiversity in the Baltic Sea: Species-specific patterns challenge management. Biodivers. Conserv. 2013, 22, 3045–3065. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Region | Local Salinity (PSU) | Salinities Where Sperm Were Tested (PSU) | Genotypes Kept after Filtering | Data from |
|---|---|---|---|---|---|
| Sozopol | ancestral Black Sea | ~16 | 0; 8; 16; 24; 32 | N = 29 | This study |
| Kindvig | Southern Baltic | ~15 | 1; 5; 10; 15; 20; 25; 30 | N = 5 | [39] Green, Havenhand et al., 2020 |
| Karrebaeksminde | Southern Baltic | ~15 | 1; 5; 10; 15; 20; 25; 30 | N = 13 | [39] Green, Havenhand et al., 2020 |
| Guldborgsund | Southern Baltic | ~15 | 0; 16; 32 | N = 32 | [38] Green, Niemax et al., 2021 |
| Travemünde | Southern Baltic | ~13 | 0; 16 | N = 31 | [35] Green, Niemax et al., 2020 |
| Mariehamn | Northern Baltic | ~5 | 1; 5; 10; 15; 20; 25; 30 | N = 26 | [39] Green, Havenhand et al., 2020 |
| Turku | Northern Baltic | ~5 | 1; 5; 10; 15; 20; 25; 30 | N = 27 | [39] Green, Havenhand et al., 2020 |
| Raahe | Northern Baltic | ~2 | 1; 5; 10; 15; 20; 25; 30 | N = 22 | [39] Green, Havenhand et al., 2020 |
| Elbe | western European river | 0 | 0; 16; 32 | N = 29 | [38] Green, Niemax et al., 2021 |
| Rhine | western European river | 0 | 0; 16; 32 | N = 32 | [38] Green, Niemax et al., 2021 |
| Danube | ancestral river | 0 | 0; 8; 16; 24; 32 | N = 18 | This study |
| Site | N | Ar | P(%) | Pa | Ho | He | FIS |
|---|---|---|---|---|---|---|---|
| Sozopol | 29 | 1.6 | 80 | 521 | 0.19 | 0.21 | 0.077 |
| Kindvig | 5 | 1.5 | 50 | 0 | 0.19 | 0.20 | −0.065 |
| Karrebaeksminde | 13 | 1.5 | 62 | 0 | 0.18 | 0.20 | 0.021 |
| Guldborgsund | 32 | 1.5 | 68 | 2 | 0.20 | 0.20 | 0.019 |
| Travemünde | 31 | 1.6 | 74 | 7 | 0.20 | 0.21 | 0.020 |
| Mariehamn | 26 | 1.4 | 49 | 1 | 0.16 | 0.17 | 0.038 |
| Turku | 27 | 1.5 | 68 | 9 | 0.19 | 0.20 | 0.031 |
| Raahe | 22 | 1.5 | 62 | 0 | 0.17 | 0.19 | 0.041 |
| Elbe | 29 | 1.5 | 68 | 182 | 0.19 | 0.20 | 0.018 |
| Rhine | 32 | 1.4 | 66 | 56 | 0.16 | 0.16 | 0.015 |
| Danube | 18 | 1.3 | 44 | 4 | 0.13 | 0.13 | 0.018 |
| Sozopol | Kindvig | Karrebaeks- Minde | Guldborg-Sund | Trave- Münde | Mariehamn | Turku | Raahe | Elbe | Rhine | |
|---|---|---|---|---|---|---|---|---|---|---|
| Kindvig | 0.113 | |||||||||
| Karrebaeksminde | 0.115 | 0.001 | ||||||||
| Guldborgsund | 0.121 | 0.019 | 0.012 | |||||||
| Travemünde | 0.106 | 0.068 | 0.070 | 0.062 | ||||||
| Mariehamn | 0.179 | 0.154 | 0.147 | 0.147 | 0.144 | |||||
| Turku | 0.113 | 0.072 | 0.074 | 0.082 | 0.070 | 0.154 | ||||
| Raahe | 0.138 | 0.092 | 0.093 | 0.095 | 0.083 | 0.173 | 0.097 | |||
| Elbe | 0.139 | 0.169 | 0.168 | 0.158 | 0.142 | 0.232 | 0.160 | 0.194 | ||
| Rhine | 0.218 | 0.308 | 0.294 | 0.275 | 0.261 | 0.339 | 0.282 | 0.304 | 0.230 | |
| Danube | 0.249 | 0.369 | 0.342 | 0.320 | 0.307 | 0.383 | 0.321 | 0.345 | 0.269 | 0.081 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Green, L.; Apostolou, A.; Faust, E.; Palmqvist, K.; Behrens, J.W.; Havenhand, J.N.; Leder, E.H.; Kvarnemo, C. Ancestral Sperm Ecotypes Reveal Multiple Invasions of a Non-Native Fish in Northern Europe. Cells 2021, 10, 1743. https://doi.org/10.3390/cells10071743
Green L, Apostolou A, Faust E, Palmqvist K, Behrens JW, Havenhand JN, Leder EH, Kvarnemo C. Ancestral Sperm Ecotypes Reveal Multiple Invasions of a Non-Native Fish in Northern Europe. Cells. 2021; 10(7):1743. https://doi.org/10.3390/cells10071743
Chicago/Turabian StyleGreen, Leon, Apostolos Apostolou, Ellika Faust, Kajsa Palmqvist, Jane W. Behrens, Jonathan N. Havenhand, Erica H. Leder, and Charlotta Kvarnemo. 2021. "Ancestral Sperm Ecotypes Reveal Multiple Invasions of a Non-Native Fish in Northern Europe" Cells 10, no. 7: 1743. https://doi.org/10.3390/cells10071743
APA StyleGreen, L., Apostolou, A., Faust, E., Palmqvist, K., Behrens, J. W., Havenhand, J. N., Leder, E. H., & Kvarnemo, C. (2021). Ancestral Sperm Ecotypes Reveal Multiple Invasions of a Non-Native Fish in Northern Europe. Cells, 10(7), 1743. https://doi.org/10.3390/cells10071743