
Commander Complex—A Multifaceted Operator in Intracellular Signaling and Cargo


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
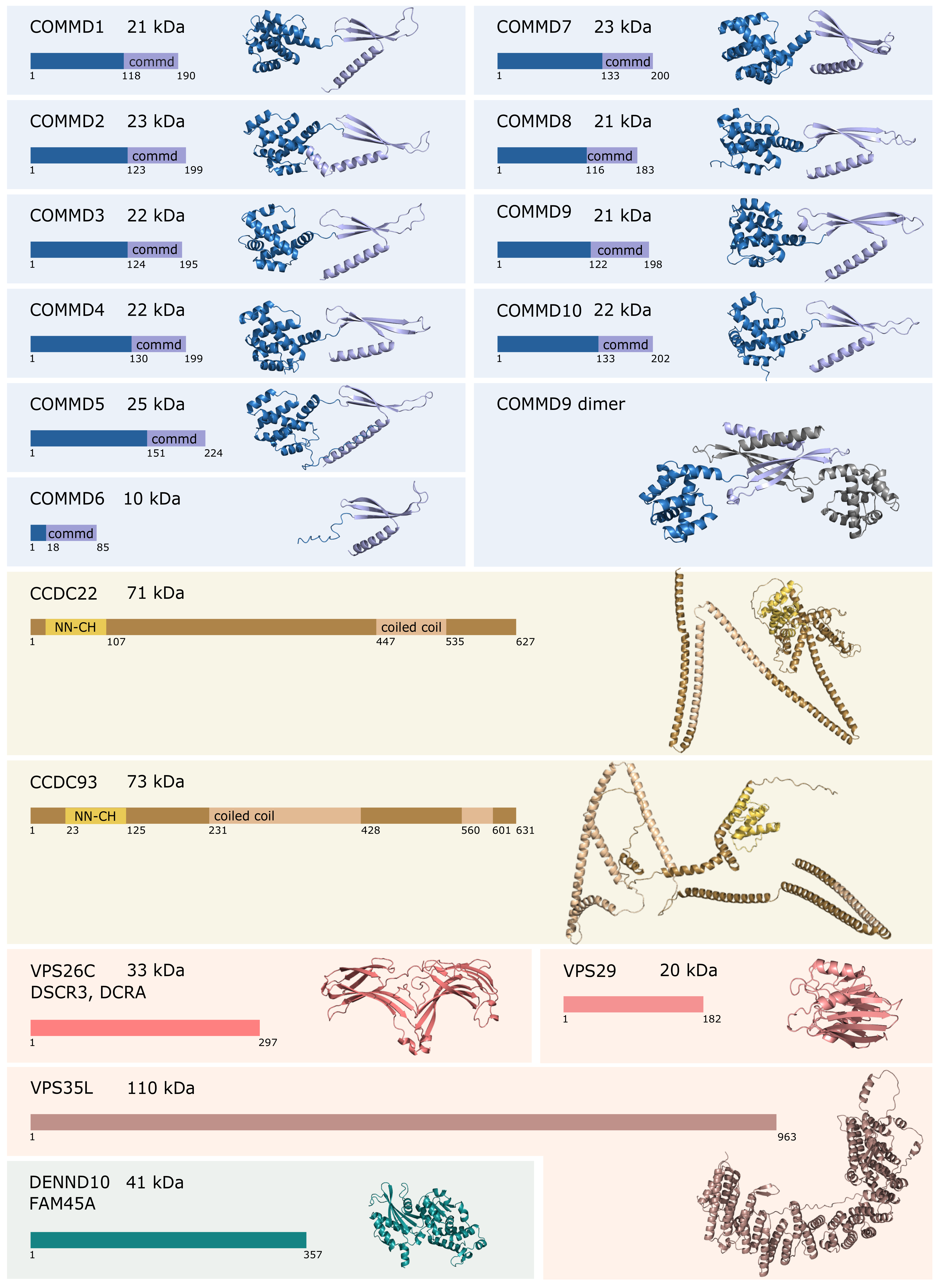
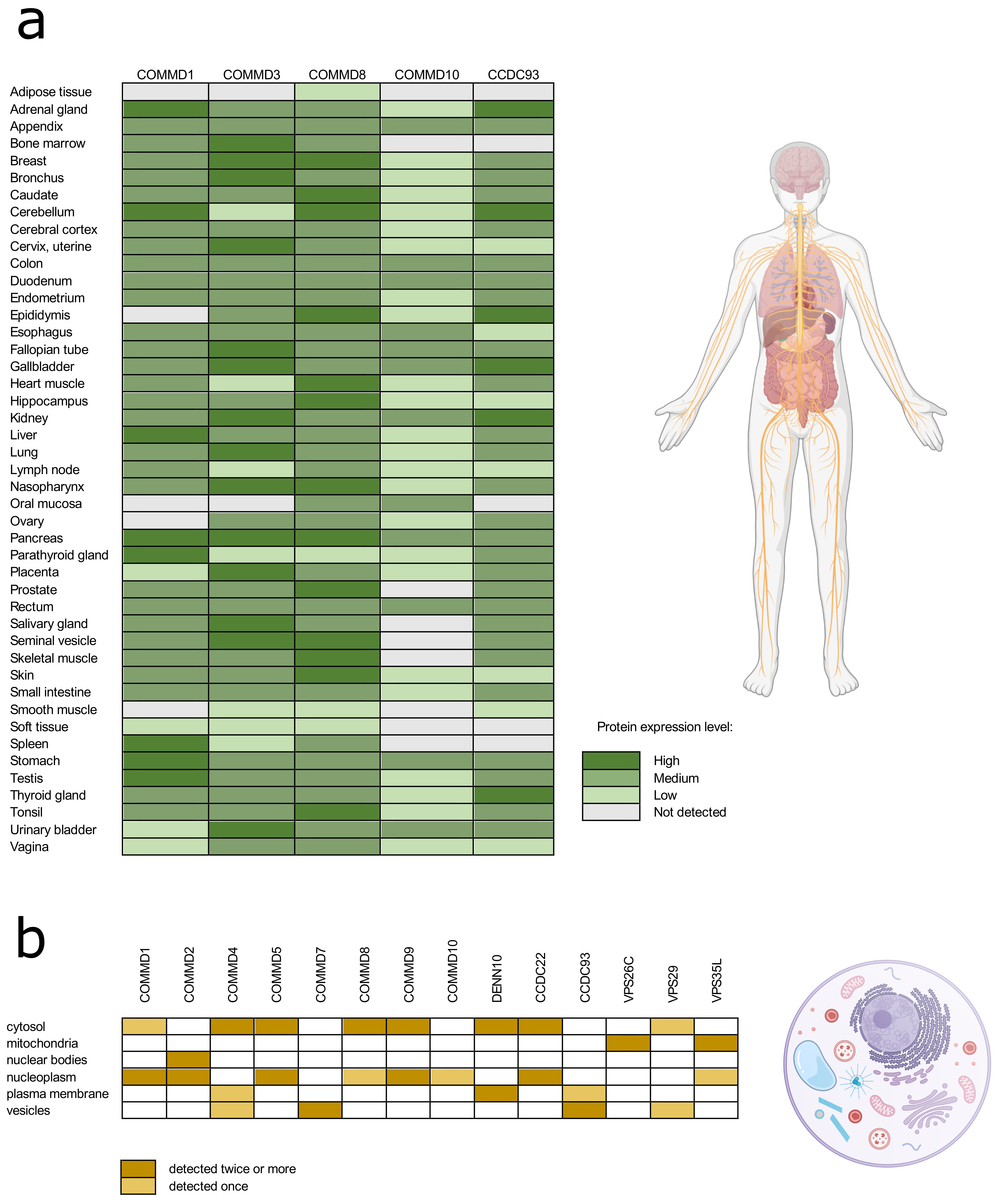
2. Composition and Assembly of the Commander Complex
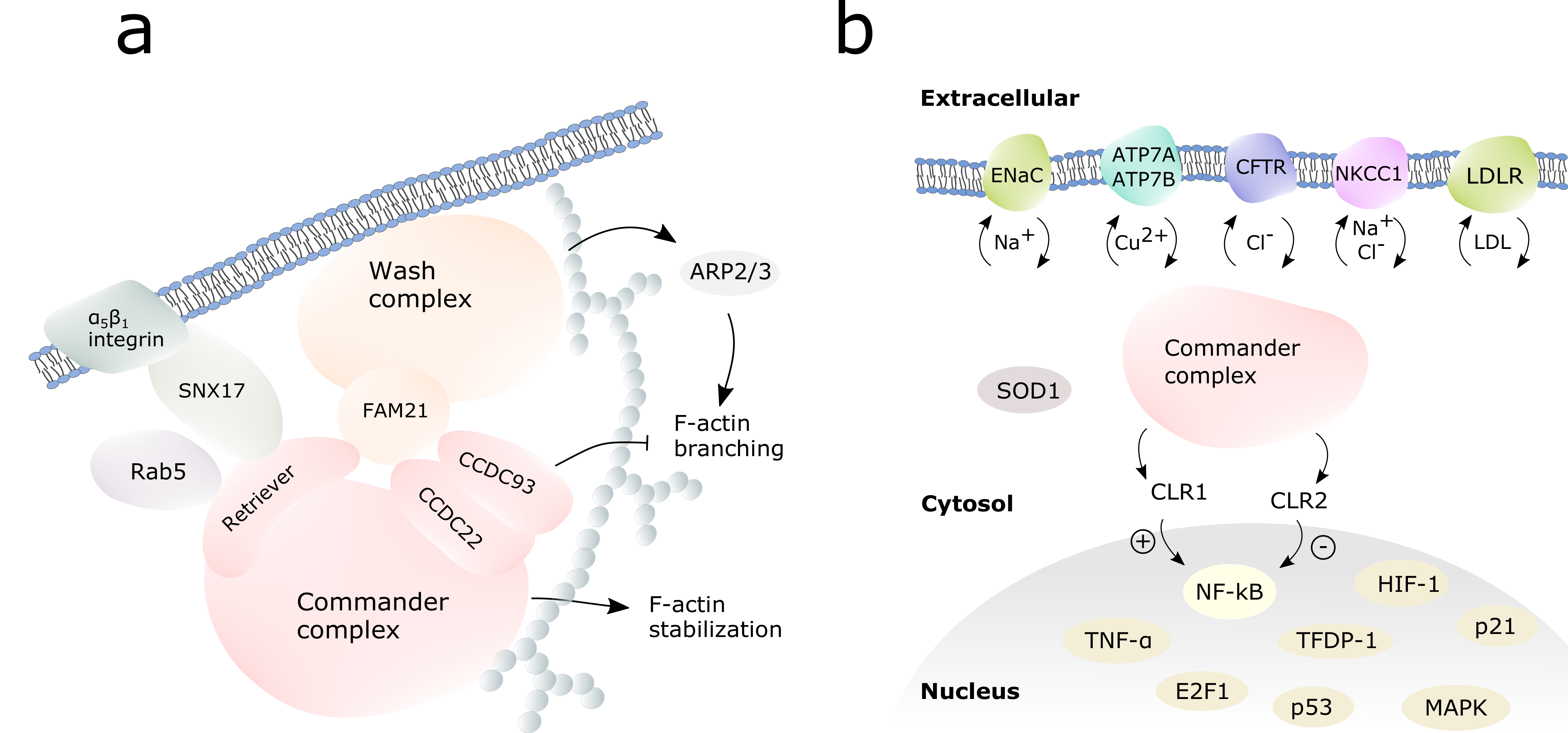
3. Endosomal Trafficking
3.1. Ion Channels and Transporters
3.2. Cholesterol Intake
3.3. Viral-Host Interactions
4. Immune Response Regulation
5. Cell Cycle and Cancer
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guo, Y.; Sirkis, D.W.; Schekman, R. Protein Sorting at the trans-Golgi Network. Annu. Rev. Cell Dev. Biol. 2014, 30, 169–206. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Fedoseienko, A.; Chen, B.; Burstein, E.; Jia, D.; Billadeau, D.D. Endosomal Receptor Trafficking: Retromer and Beyond. Traffic 2018, 19, 578–590. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.E.; Healy, M.D.; Collins, B.M. Towards a molecular understanding of endosomal trafficking by Retromer and Retriever. Traffic 2019, 20, 465–478. [Google Scholar] [CrossRef] [PubMed]
- Wan, C.; Borgeson, B.; Phanse, S.; Tu, F.; Drew, K.; Clark, G.; Xiong, X.; Kagan, O.; Kwan, J.; Bezginov, A.; et al. Panorama of ancient metazoan macromolecular complexes. Nature 2015, 525, 339–344. [Google Scholar] [CrossRef]
- Mallam, A.L.; Marcotte, E.M. Systems-wide studies uncover Commander, a multiprotein complex essential to human development. Cell Syst. 2017, 4, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Dey, G.; Jaimovich, A.; Collins, S.R.; Seki, A.; Meyer, T. Systematic Discovery of Human Gene Function and Principles of Modular Organization through Phylogenetic Profiling. Cell Rep. 2015, 10, 993–1006. [Google Scholar] [CrossRef] [PubMed]
- Hein, M.Y.; Hubner, N.C.; Poser, I.; Cox, J.; Nagaraj, N.; Toyoda, Y.; Gak, I.A.; Weisswange, I.; Mansfeld, J.; Buchholz, F.; et al. A Human Interactome in Three Quantitative Dimensions Organized by Stoichiometries and Abundances. Cell 2015, 163, 712–723. [Google Scholar] [CrossRef] [PubMed]
- Huttlin, E.L.; Ting, L.; Bruckner, R.J.; Gebreab, F.; Gygi, M.P.; Szpyt, J.; Tam, S.; Zarraga, G.; Colby, G.; Baltier, K.; et al. The BioPlex Network: A Systematic Exploration of the Human Interactome. Cell 2015, 162, 425–440. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Calvo, S.E.; Gutman, R.; Liu, J.S.; Mootha, V.K. Expansion of biological pathways based on evolutionary inference. Cell 2014, 158, 213–225. [Google Scholar] [CrossRef]
- McNally, K.E.; Faulkner, R.; Steinberg, F.; Gallon, M.; Ghai, R.; Pim, D.; Langton, P.; Pearson, N.; Danson, C.M.; Nägele, H.; et al. Retriever is a multiprotein complex for retromer-independent endosomal cargo recycling. Nat. Cell Biol. 2017, 19, 1214–1225. [Google Scholar] [CrossRef]
- Singla, A.; Fedoseienko, A.; Giridharan, S.S.P.; Overlee, B.L.; Lopez, A.; Jia, D.; Song, J.; Huff-Hardy, K.; Weisman, L.; Burstein, E.; et al. Endosomal PI(3)P regulation by the COMMD/CCDC22/CCDC93 (CCC) complex controls membrane protein recycling. Nat. Commun. 2019, 10, 4271. [Google Scholar] [CrossRef]
- van de Sluis, B.; Rothuizen, J.; Pearson, P.L.; van Oost, B.A.; Wijmenga, C. Identification of a new copper metabolism gene by positional cloning in a purebred dog population. Hum. Mol. Genet. 2002, 11, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Biasio, W.; Chang, T.; McIntosh, C.J.; McDonald, F.J. Identification of Murr1 as a Regulator of the Human Epithelial Sodium Channel. J. Biol. Chem. 2004, 279, 5429–5434. [Google Scholar] [CrossRef]
- Drevillon, L.; Tanguy, G.; Hinzpeter, A.; Arous, N.; de Becdelievre, A.; Aissat, A.; Tarze, A.; Goossens, M.; Fanen, P. COMMD1-Mediated Ubiquitination Regulates CFTR Trafficking. PLoS ONE 2011, 6, e18334. [Google Scholar] [CrossRef]
- Smith, L.; Litman, P.; Liedtke, C.M. COMMD1 interacts with the COOH terminus of NKCC1 in Calu-3 airway epithelial cells to modulate NKCC1 ubiquitination. Am. J. Physiol. Cell Physiol. 2013, 305, C133–C146. [Google Scholar] [CrossRef] [PubMed]
- Burkhead, J.L.; Morgan, C.T.; Shinde, U.; Haddock, G.; Lutsenko, S. COMMD1 Forms Oligomeric Complexes Targeted to the Endocytic Membranes via Specific Interactions with Phosphatidylinositol 4,5-Bisphosphate. J. Biol. Chem. 2009, 284, 696–707. [Google Scholar] [CrossRef]
- Fedoseienko, A.; Wijers, M.; Wolters, J.C.; Dekker, D.; Smit, M.; Huijkman, N.; Kloosterhuis, N.; Klug, H.; Schepers, A.; van Dijk, K.W.; et al. The COMMD Family Regulates Plasma LDL Levels and Attenuates Atherosclerosis through Stabilizing the CCC Complex in Endosomal LDLR Trafficking. Circ. Res. 2018, 122, 1648–1660. [Google Scholar] [CrossRef]
- Bartuzi, P.; Billadeau, D.D.; Favier, R.; Rong, S.; Dekker, D.; Fedoseienko, A.; Fieten, H.; Wijers, M.; Levels, J.H.; Huijkman, N.; et al. CCC- and WASH-mediated endosomal sorting of LDLR is required for normal clearance of circulating LDL. Nat. Commun. 2017, 7, 10961. [Google Scholar] [CrossRef]
- Maine, G.N.; Mao, X.; Komarck, C.M.; Burstein, E. COMMD1 promotes the ubiquitination of NF-jB subunits through a cullin-containing ubiquitin ligase. EMBO J. 2007, 26, 436–447. [Google Scholar] [CrossRef]
- Mao, X.; Gluck, N.; Chen, B.; Starokadomskyy, P.; Li, H.; Maine, G.N.; Burstein, E. COMMD1 (Copper Metabolism MURR1 Domain-containing Protein 1) Regulates Cullin RING Ligases by Preventing CAND1 (Cullin-associated Nedd8-dissociated Protein 1) Binding. J. Biol. Chem. 2011, 286, 32355–32365. [Google Scholar] [CrossRef]
- Starokadomskyy, P.; Gluck, N.; Li, H.; Chen, B.; Wallis, M.; Maine, G.N.; Mao, X.; Zaidi, I.W.; Hein, M.Y.; McDonald, F.J.; et al. CCDC22 deficiency in humans blunts activation of proinflammatory NF-κB signaling. J. Clin. Investig. 2013, 123, 2244–2256. [Google Scholar] [CrossRef] [PubMed]
- Mouhadeb, O.; Shlomo, S.B.; Cohen, K.; Farkash, I.; Gruber, S.; Maharshak, N.; Halpern, Z.; Burstein, E.; Gluck, N.; Varol, C. Impaired COMMD10-Mediated Regulation of Ly6C(hi) Monocyte-Driven Inflammation Disrupts Gut Barrier Function. Front. Immunol. 2018, 9, 2623. [Google Scholar] [CrossRef]
- Nakai, A.; Fujimoto, J.; Miyata, H.; Stumm, R.; Narazaki, M.; Schulz, S.; Baba, Y.; Kumanogoh, A.; Suzuki, K. The COMMD3/8 complex determines GRK6 specificity for chemoattractant receptors. J. Exp. Med. 2019, 216, 1630–1647. [Google Scholar] [CrossRef] [PubMed]
- Devlin, A.M.; Solban, N.; Tremblay, S.; Gutkowska, J.; Schurch, W.; Orlov, S.N.; Lewanczuk, R.; Hamet, P.; Tremblay, J. HCaRG is a novel regulator of renal epithelial cell growth and differentiation causing G2M arrest. Am. J. Physiol. Renal Physiol. 2003, 284, F753–F762. [Google Scholar] [CrossRef] [PubMed]
- Van de Sluis, B.; Mao, X.; Zhai, Y.; Groot, A.J.; Vermeulen, J.F.; van der Wall, E.; van Diest, P.J.; Hofker, M.H.; Wijmenga, C.; Klomp, L.W.; et al. COMMD1 disrupts HIF-1 dimerization and inhibits human tumor cell invasion. J. Clin. Investig. 2010, 120, 2119–2130. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Koo, Y.; Mao, X.; Sifuentes-Dominguez, L.; Morris, L.L.; Jia, D.; Miyata, N.; Faulkner, R.A.; van Deursen, J.M.; Vooijs, M.; et al. Endosomal sorting of Notch receptors through COMMD9- dependent pathways modulates Notch signaling. J. Cell Biol. 2015, 211, 605–617. [Google Scholar] [CrossRef]
- Zhan, W.; Wang, W.; Han, T.; Xie, C.; Zhang, T.; Gan, M.; Wang, J.B. COMMD9 promotes TFDP1/E2F1 transcriptional activity via interaction with TFDP1 in non-small cell lung cancer. Cell Signal 2017, 30, 59–66. [Google Scholar] [CrossRef]
- Bandmann, O.; Heinz Weiss, K.; Kaler, S.G. Wilson’s disease and other neurological copper disorders. Lancet Neurol. 2015, 14, 103–113. [Google Scholar] [CrossRef]
- Vonk, W.I.M.; Kakkar, V.; Bartuzi, P.; Jaarsma, D.; Berger, R.; Hofker, M.H.; Klomp, L.W.; Wijmenga, C.; Kampinga, H.H.; van de Sluis, B. The Copper Metabolism MURR1 Domain Protein 1 (COMMD1) Modulates the Aggregation of Misfolded Protein Species in a Client-Specific Manner. PLoS ONE 2014, 9, e92408. [Google Scholar]
- Daniloski, Z.; Jordan, T.X.; Wessels, H.H.; Hoagland, D.A.; Kasela, S.; Legut, M.; Maniatis, S.; Mimitou, E.P.; Lu, L.; Geller, E.; et al. Identification of Required Host Factors for SARS-CoV-2 Infection in Human Cells. Cell 2021, 184, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; He, S.X.; Zheng, X.; Huang, S.; Chen, H.; Chen, H.; Luo, W.; Guo, Z.; He, X.; Zhao, Q. Transcriptional analysis of the expression, prognostic value and immune infiltration activities of the COMMD protein family in hepatocellular carcinoma. BMC Cancer 2021, 21, 1001. [Google Scholar] [CrossRef]
- Matsuda, H.; Campion, C.G.; Fujiwara, K.; Ikeda, J.; Cossette, S.; Verissimo, T.; Ogasawara, M.; Gaboury, L.; Saito, K.; Yamaguchi, K.; et al. HCaRG/COMMD5 inhibits ErbB receptor-driven renal cell carcinoma. Oncotarget 2017, 8, 69559–69576. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Huang, W.; Sun, Y.; Liang, H.; Chen, M.; Wu, X.; Wang, X.; Zhang, L.; Cheng, X.; Fan, Y.; et al. Prognosis and modulation mechanisms of COMMD6 in human tumours based on expression profiling and comprehensive bioinformatics analysis. Br. J. Cancer 2019, 121, 699–709. [Google Scholar] [CrossRef]
- Riera-Romo, M. COMMD1: A Multifunctional Regulatory Protein. J. Cell Biochem. 2018, 119, 34–51. [Google Scholar] [CrossRef]
- Burstein, E.; Hoberg, J.E.; Wilkinson, A.S.; Rumble, J.M.; Csomos, R.A.; Komarck, C.M.; Maine, G.N.; Wilkinson, J.C.; Mayo, M.W.; Duckett, C.S. COMMD Proteins, a Novel Family of Structural and Functional Homologs of MURR1. J. Biol. Chem. 2005, 280, 22222–22232. [Google Scholar] [CrossRef] [PubMed]
- Healy, M.D.; Hospenthal, M.K.; Hall, R.J.; Chandra, M.; Chilton, M.; Tillu, V.; Chen, K.E.; Celligoi, J.; McDonald, F.J.; Cullen, P.J.; et al. Structural insights into the architecture and membrane interactions of the conserved COMMD proteins. eLife 2018, 7, e35898. [Google Scholar] [CrossRef] [PubMed]
- Narindrasorasak, S.; Kulkarni, P.; Deschamps, P.; She, Y.M.; Sarkar, B. Characterization and Copper Binding Properties of Human COMMD1 (MURR1). Biochemistry 2007, 46, 3116–3128. [Google Scholar] [CrossRef]
- AlphaFold Protein Structure Database. Available online: Alphafold.ebi.ac.uk (accessed on 1 October 2021).
- Phillips-Krawczak, C.A.; Singla, A.; Starokadomskyy, P.; Deng, Z.; Osborne, D.G.; Li, H.; Dick, C.J.; Gomez, T.S.; Koenecke, M.; Zhang, J.S.; et al. COMMD1 is linked to the WASH complex and regulates endosomal trafficking of the copper transporter ATP7A. Mol. Biol. Cell 2015, 26, 91–103. [Google Scholar] [CrossRef]
- Schou, K.B.; Andersen, J.S.; Pedersen, L.B. A divergent calponin homology (NN–CH) domain defines a novel family: Implications for evolution of ciliary IFT complex B proteins. Bioinformatics 2014, 30, 899–902. [Google Scholar] [CrossRef] [PubMed]
- Kirkwood, K.J.; Ahmad, Y.; Larance, M.; Lamond, A.I. Characterization of native protein complexes and protein isoform variation using size-fractionation-based quantitative proteomics. Mol. Cell Proteom. 2013, 12, 3851–3873. [Google Scholar] [CrossRef]
- Kendall, A.K.; Xie, B.; Xu, P.; Wang, J.; Burcham, R.; Frazier, M.N.; Binshtein, E.; Wei, H.; Graham, T.R.; Nakagawa, T.; et al. Mammalian Retromer Is an Adaptable Scaffold for Cargo Sorting from Endosomes. Structure 2020, 28, 393–405. [Google Scholar] [CrossRef]
- Harbour, M.E.; Breusegem, S.Y.; Seaman, M.N.J. Recruitment of the endosomal WASH complex is mediated by the extended ‘tail’ of Fam21 binding to the retromer protein Vps35. Biochem. J. 2012, 442, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Schrödinger, LLC. The PyMOL Molecular Graphics System, Version 2.0; Schrödinger, LLC.: New York, NY, USA, 2015. [Google Scholar]
- Zhang, T.J.; Zhang, K.; Qi, L.; Hu, Q.; Shen, Z.; Liu, B.; Deng, J.; Zhang, C.; Zhang, Y. DENN domain-containing protein FAM45A regulates the homeostasis of late/multivesicular endosomes. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 916–929. [Google Scholar] [CrossRef] [PubMed]
- GnomAD Genome Aggregation Database v2.1.1. Available online: Gnomad.broadinstitute.org (accessed on 1 October 2021).
- The Human Protein Atlas. Available online: www.proteinatlas.org (accessed on 1 October 2021).
- Linardopoulou, E.V.; Parghi, S.S.; Friedman, C.; Osborn, G.E.; Parkhurst, S.M.; Trask, B.J. Human Subtelomeric WASH Genes Encode a New Subclass of the WASP Family. PLoS Gen. 2007, 3, e237. [Google Scholar] [CrossRef] [PubMed]
- Rojas, R.; van Vlijmen, T.; Mardones, G.A.; Prabhu, Y.; Rojas, A.L.; Mohammed, S.; Heck, A.J.R.; Raposo, G.; van der Sluijs, P.; Bonifacino, J.S. Regulation of retromer recruitment to endosomes by sequential action of Rab5 and Rab7. J. Cell Biol. 2008, 183, 513–526. [Google Scholar] [CrossRef]
- Campion, C.G.; Zaoui, K.; Verissimo, T.; Cossette, S.; Matsuda, H.; Solban, N.; Hamet, P.; Tremblay, J. COMMD5/HCaRG Hooks Endosomes on Cytoskeleton and Coordinates EGFR Trafficking. Cell Rep. 2018, 24, 670–684. [Google Scholar] [CrossRef] [PubMed]
- Priya, A.; Kalaidzidis, I.V.; Kalaidzidis, Y.; Lambright, D.; Datta, S. Molecular Insights into Rab7-Mediated Endosomal Recruitment of Core Retromer: Deciphering the Role of Vps26 and Vps35. Traffic 2015, 16, 68–84. [Google Scholar] [CrossRef] [PubMed]
- Duleh, S.N.; Welch, M.D. WASH and the Arp2/3 complex regulate endosome shape and trafficking. Cytoskeleton 2010, 67, 193–206. [Google Scholar] [CrossRef] [PubMed]
- Gomez, T.S.; Billadeau, D.D. A FAM21-Containing WASH Complex Regulates Retromer-Dependent Sorting. Dev. Cell 2009, 17, 699–711. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hodgkinson, V.; Zhu, S.; Weisman, G.A.; Petris, M.J. Advances in the Understanding of Mammalian Copper Transporters. Adv. Nutr. 2011, 2, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Materia, S.; Cater, M.A.; Klomp, L.W.J.; Mercer, J.F.B.; La Fontaine, S. Clusterin and COMMD1 Independently Regulate Degradation of the Mammalian Copper ATPases ATP7A and ATP7B. J. Biol. Chem. 2012, 287, 2485–2499. [Google Scholar] [CrossRef] [PubMed]
- Voineagu, I.; Huang, L.; Winden, K.; Lazaro, M.; Haan, E.; Nelson, J.; McGaughran, J.; Nguyen, L.S.; Friend, K.; Hackett, A.; et al. CCDC22: A novel candidate gene for syndromic X-linked intellectual disability. Mol. Psychiatry 2012, 17, 4–7. [Google Scholar] [CrossRef]
- Tao, T.Y.; Liu, F.; Klomp, L.; Wijmenga, C.; Gitlin, J.D. The Copper Toxicosis Gene Product Murr1 Directly Interacts with the Wilson Disease Protein. J. Biol. Chem. 2003, 278, 41593–41596. [Google Scholar] [CrossRef] [PubMed]
- De Bie, P.; van de Sluis, B.; Burstein, E.; van de Berghe, P.V.E.; Muller, P.; Berger, R.; Gitlin, J.D.; Wijmenga, C.; Klomp, L.W.J. Distinct Wilson-disease mutations in ATP7B are associated with enhanced binding to COMMD1 and reduced stability of ATP7B. Gastroenterology 2007, 133, 1316–1326. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.; Ke, Y.; Ly, K.; McDonald, F.J. COMMD1 regulates the delta epithelial sodium channel (dENaC) through trafficking and ubiquitination. Biochem. Biophys. Res. Commun. 2011, 411, 506–511. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.F.; Swart, M.; Ke, Y.; Ly, K.; McDonald, F.J. Functional interaction of COMMD3 and COMMD9 with the epithelial sodium channel. Am. J. Physiol. Renal Physiol. 2013, 305, F80–F89. [Google Scholar] [CrossRef] [PubMed]
- Ware, A.W.; Cheung, T.T.; Rasulov, S.; Burstein, E.; McDonald, F.J. Epithelial Na+ Channel: Reciprocal Control by COMMD10 and Nedd4-2. Front. Physiol. 2018, 9, 793. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Patel, S.V.; Snyder, P.M. Nedd4-2 Catalyzes Ubiquitination and Degradation of Cell Surface ENaC. J. Biol. Chem. 2007, 282, 20207–20212. [Google Scholar] [CrossRef] [PubMed]
- Ke, Y.; Butt, A.G.; Swart, M.; Liu, Y.F.; McDonald, F.J. COMMD1 downregulates the epithelial sodium channel through Nedd4–2. Am. J. Physiol. Renal Physiol. 2010, 298, F1445–F1456. [Google Scholar] [CrossRef] [PubMed]
- Solban, N.; Jia, H.P.; Richard, S.; Tremblay, S.; Devlin, A.M.; Peng, J.; Gossard, F.; Guo, D.F.; Morel, G.; Hamet, P.; et al. HCaRG, a novel calcium-regulated gene coding for a nuclear protein, is potentially involved in the regulation of cell proliferation. J. Biol. Chem. 2000, 275, 32234–32243. [Google Scholar] [CrossRef] [PubMed]
- Groppelli, E.; Len, A.C.; Granger, L.A.; Jolly, C. Retromer regulates HIV-1 envelope glycoprotein trafficking and incorporation into virions. PLoS Pathog. 2014, 10, e1004518. [Google Scholar] [CrossRef] [PubMed]
- Popa, A.; Zhang, W.; Harrison, M.S.; Goodner, K.; Kazakov, T.; Goodwin, E.C.; Lipovsky, A.; Burd, C.G.; DiMaio, D. Direct binding of retromer to human papillomavirus type 16 minor capsid protein L2 mediates endosome exit during viral infection. PLoS Pathog. 2015, 11, e1004699. [Google Scholar] [CrossRef]
- Mirrashidi, K.M.; Elwell, C.A.; Verschueren, E.; Johnson, J.R.; Frando, A.; von Dollen, J.; Rosenberg, O.; Gulbahce, N.; Jang, G.; Johnson, T.; et al. Global mapping of the Inc-human Interactome reveals that Retromer restricts chlamydia infection. Cell Host Microbe. 2015, 18, 109–121. [Google Scholar] [CrossRef]
- Yao, J.; Yang, F.; Sun, X.; Wang, S.; Gan, N.; Liu, Q.; Liu, D.; Zhang, X.; Niu, D.; Wei, Y.; et al. Mechanism of inhibition of retromer transport by the bacterial effector RidL. Proc. Natl. Acad. Sci. USA 2018, 115, E1446–E1454. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; You, N.; Huang, X.; Gu, H.; Wu, K.; Mi, N.; Li, J. COMMD7 Regulates NF-kB Signaling Pathway in Hepatocellular Carcinoma Stem-like Cells. Mol. Ther. Oncolytics 2019, 12, 112–123. [Google Scholar] [CrossRef]
- Esposito, E.; Napolitano, G.; Pescatore, A.; Calculli, G.; Incoronato, M.R.; Leonardi, A.; Ursini, M.V. COMMD7 as a novel NEMO interacting protein involved in the termination of NF-κB signaling. J. Cell Physiol. 2016, 231, 152–161. [Google Scholar] [CrossRef] [PubMed]
- You, N.; Li, J.; Huang, X.; Wu, K.; Tang, Y.; Wang, L.; Li, H.; Mi, N.; Zheng, L. COMMD7 promotes hepatocellular carcinoma through regulating CXCL10. Biomed. Pharmacother. 2017, 88, 653–657. [Google Scholar] [CrossRef] [PubMed]
- Sperger, J.M.; Chen, X.; Draper, J.S.; Antosiewicz, J.E.; Chon, C.H.; Jones, S.B.; Brooks, J.D.; Andrews, P.W.; Brown, P.O.; Thomson, J.A. Gene expression patterns in human embryonic stem cells and human pluripotent germ cell tumors. Proc. Natl. Acad. Sci. USA 2003, 100, 13350–13355. [Google Scholar] [CrossRef]
- Fan, Y.; Zhang, L.; Sun, Y.; Yang, M.; Wang, X.; Wu, X.; Huang, W.; Chen, L.; Pan, S.; Guan, J. Expression profile and bioinformatics analysis of COMMD10 in BALB/C mice and human. Cancer Gene Ther. 2020, 27, 216–225. [Google Scholar] [CrossRef]
- Yang, S.S.; Li, X.M.; Yang, M.; Ren, X.L.; Hu, J.L.; Zhu, X.H.; Wang, F.F.; Zeng, Z.C.; Li, J.Y.; Cheng, Z.Q.; et al. FMNL2 destabilises COMMD10 to activate NF-kB pathway in invasion and metastasis of colorectal cancer. Br. J. Cancer 2017, 117, 1164–1175. [Google Scholar] [CrossRef] [PubMed]
- You, N.; Li, J.; Gong, Z.; Huang, X.; Wang, W.; Wang, L.; Wu, K.; Zheng, L. COMMD7 Functions as Molecular Target in Pancreatic Ductal Adenocarcinoma. Mol. Carcinog. 2017, 56, 607–624. [Google Scholar] [CrossRef] [PubMed]
- Van de Sluis, B.; Muller, P.; Duran, K.; Chen, A.; Groot, A.J.; Klomp, L.W.; Liu, P.P.; Wijmenga, C. Increased Activity of Hypoxia-Inducible Factor 1 Is Associated with Early Embryonic Lethality in Commd1 Null Mice. Mol. Cell Biol. 2007, 27, 4142–4156. [Google Scholar] [CrossRef]
- Mu, P.; Akashi, T.; Lu, F.; Kishida, S.; Kadomatsu, K. A novel nuclear complex of DRR1, F-actin and COMMD1 involved in NF-κB degradation and cell growth suppression in neuroblastoma. Oncogene 2017, 36, 5745–5756. [Google Scholar] [CrossRef] [PubMed]
- Vonk, W.I.M.; Wijmenga, C.; Berger, R.; van de Sluis, B.; Klomp, L.W.J. Cu,Zn Superoxide Dismutase Maturation and Activity Are Regulated by COMMD1. J. Biol. Chem. 2010, 285, 28991–29000. [Google Scholar] [CrossRef] [PubMed]
- Dumoulin, B.; Ufer, C.; Stehling, S.; Heydeck, D.; Kuhn, H.; Sofi, S. Identification of the COMM-domain containing protein 1 as specific binding partner for the guanine-rich RNA sequence binding factor 1. Biochem. Biophys. Acta Gen. Subj. 2020, 1864, 129678. [Google Scholar] [CrossRef] [PubMed]
- You, N.; Li, J.; Huang, X.; Wu, K.; Tang, Y.; Wang, L.; Li, H.; Mi, N.; Zheng, L. COMMD7 activates CXCL10 production by regulating NF-κB and the production of reactive oxygen species. Mol. Med. Rep. 2018, 17, 6784–6788. [Google Scholar] [CrossRef]
- Matsuda, H.; Hamet, P.; Tremblay, J. Hypertension-related, calcium-regulated gene (HCaRG/COMMD5) and kidney diseases: HCaRG accelerates tubular repair. J. Nephrol. 2014, 27, 351–360. [Google Scholar] [CrossRef] [PubMed][Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laulumaa, S.; Varjosalo, M. Commander Complex—A Multifaceted Operator in Intracellular Signaling and Cargo. Cells 2021, 10, 3447. https://doi.org/10.3390/cells10123447
Laulumaa S, Varjosalo M. Commander Complex—A Multifaceted Operator in Intracellular Signaling and Cargo. Cells. 2021; 10(12):3447. https://doi.org/10.3390/cells10123447
Chicago/Turabian StyleLaulumaa, Saara, and Markku Varjosalo. 2021. "Commander Complex—A Multifaceted Operator in Intracellular Signaling and Cargo" Cells 10, no. 12: 3447. https://doi.org/10.3390/cells10123447
APA StyleLaulumaa, S., & Varjosalo, M. (2021). Commander Complex—A Multifaceted Operator in Intracellular Signaling and Cargo. Cells, 10(12), 3447. https://doi.org/10.3390/cells10123447