
Nutrient Solution Deprivation as a Tool to Improve Hydroponics Sustainability: Yield, Physiological, and Qualitative Response of Lettuce

 ,
,  ,
,  , , ,
, , ,  ,
,  ,
, 
 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Sampling and CIELab Colorimetric Index
2.3. Spad Index and Chlorophyll Fluorescence
2.4. Pigments Determination
2.5. Total Nitrogen and Mineral Profile Determination
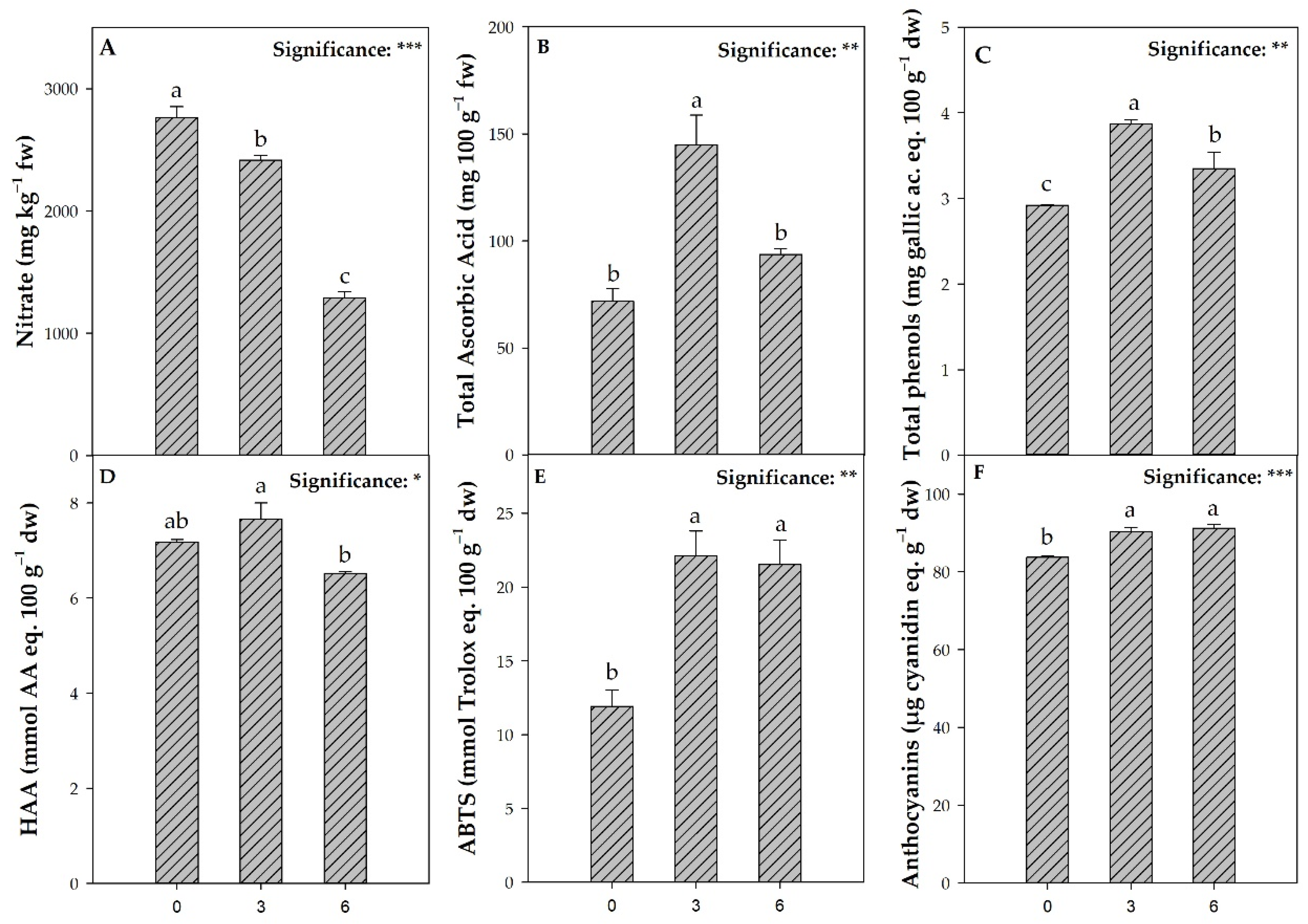
2.6. Total Phenol, Total Ascorbic Acid, Anthocyanins, Hydrogen Peroxide, and Antioxidant Activities Determination
2.7. Carbohydrates, Soluble Proteins, and Amino Acids Analysis
2.8. Statistics
3. Results
3.1. Yield and Yield Parameters
3.2. Colorimetric Parameters
3.3. Spad Index, Chlorophyll Fluorescence, and Hydrogen Peroxide
3.4. Mineral Composition
3.5. Quality Parameters
3.6. Organic Acid Profile and Carbohydrates Content
3.7. Soluble Proteins and Amino Acids Profile
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Vries, I.M. Origin and domestication of Lactuca sativa L. Genet. Resour. Crop Evol. 1997, 44, 165–174. [Google Scholar] [CrossRef]
- Barbosa, G.L.; Almeida Gadelha, F.D.; Kublik, N.; Proctor, A.; Reichelm, L.; Weissinger, E.; Wohlleb, G.M.; Halden, R.U. Comparison of land, water, and energy requirements of lettuce grown using hydroponic vs. conventional agricultural methods. Int. J. Environ. Res. Public Health 2015, 12, 6879–6891. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Moon, Y.; Tou, J.C.; Mou, B.; Waterland, N.L. Nutritional value, bioactive compounds and health benefits of lettuce (Lactuca sativa L.). J. Food Compos. Anal. 2016, 49, 19–34. [Google Scholar] [CrossRef]
- Zhou, W.; Chen, Y.; Xu, H.; Liang, X.; Hu, Y.; Jin, C.; Lu, L.; Lin, X. Short-term nitrate limitation prior to harvest improves phenolic compound accumulation in hydroponic-cultivated lettuce (Lactuca sativa L.) without reducing shoot fresh weight. J. Agric. Food Chem. 2018, 66, 10353–10361. [Google Scholar] [CrossRef] [PubMed]
- Sapkota, S.; Sapkota, S.; Liu, Z. Effects of nutrient composition and lettuce cultivar on crop production in hydroponic culture. Horticulturae 2019, 5, 72. [Google Scholar] [CrossRef]
- Damerum, A.; Chapman, M.A.; Taylor, G. Innovative breeding technologies in lettuce for improved post-harvest quality. Postharvest Biol. Technol. 2020, 168, 111266. [Google Scholar] [CrossRef]
- Miras-Moreno, B.; Corrado, G.; Zhang, L.; Senizza, B.; Righetti, L.; Bruni, R.; El-Nakhel, C.; Sifola, M.I.; Pannico, A.; De Pascale, S.; et al. The metabolic reprogramming induced by sub-optimal nutritional and light inputs in soilless cultivated green and red butterhead lettuce. Int. J. Mol. Sci. 2020, 21, 6381. [Google Scholar] [CrossRef]
- El-Nakhel, C.; Pannico, A.; Kyriacou, M.C.; Giordano, M.; De Pascale, S.; Rouphael, Y. Macronutrient deprivation eustress elicits differential secondary metabolites in red and green-pigmented butterhead lettuce grown in a closed soilless system. J. Sci. Food Agric. 2019, 99, 6962–6972. [Google Scholar] [CrossRef]
- Simko, I. Genetic variation in response to N, P, or K deprivation in baby leaf lettuce. Horticulturae 2020, 6, 15. [Google Scholar] [CrossRef]
- Frasetya, B.; Harisman, K.; Ramdaniah, N.A.H. The effect of hydroponics systems on the growth of lettuce. In Proceedings of the IOP Conference Series: Materials Science and Engineering, Bandung Barat, Indonesia, 20–21 April 2020; IOP Publishing: Suzhou, China, 2021; Volume 1098, p. 042115. [Google Scholar]
- Touliatos, D.; Dodd, I.C.; Mcainsh, M. Vertical farming increases lettuce yield per unit area compared to conventional horizontal hydroponics. Food Energy Secur. 2016, 5, 184–191. [Google Scholar] [CrossRef]
- Kotsiras, A.; Vlachodimitropoulou, A.; Gerakaris, A.; Bakas, N.; Darras, A.I. Innovative harvest practices of Butterhead, Lollo rosso and Batavia green lettuce (Lactuca sativa L.) types grown in floating hydroponic system to maintain the quality and improve storability. Sci. Hortic. 2016, 201, 1–9. [Google Scholar] [CrossRef]
- Tomasi, N.; Pinton, R.; Dalla Costa, L.; Cortella, G.; Terzano, R.; Mimmo, T.; Scampicchio, M.; Cesco, S. New “solutions” for floating cultivation system of ready-to-eat salad: A review. Trends Food Sci. Technol. 2015, 46, 267–276. [Google Scholar] [CrossRef]
- Ciriello, M.; Formisano, L.; El Nakhel, C.; Corrado, G.; Pannico, A.; De Pascale, S.; Rouphael, Y. Morpho-physiological responses and secondary metabolites modulation by preharvest factors of three hydroponically grown genovese basil cultivars. Front. Plant Sci. 2021, 12, 619. [Google Scholar] [CrossRef] [PubMed]
- Borgognone, D.; Rouphael, Y.; Cardarelli, M.; Lucini, L.; Colla, G. Changes in biomass, mineral composition, and quality of cardoon in response to NO3−:Cl− ratio and nitrate deprivation from the nutrient solution. Front. Plant Sci. 2016, 7, 978. [Google Scholar] [CrossRef]
- Gruda, N. Do soilless culture systems have an influence on product quality of vegetables? J. Appl. Bot. Food Qual. 2009, 82, 141–147. [Google Scholar] [CrossRef]
- Bian, Z.; Wang, Y.; Zhang, X.; Li, T.; Grundy, S.; Yang, Q.; Cheng, R. A review of environment effects on nitrate accumulation in leafy vegetables grown in controlled environments. Foods 2020, 9, 732. [Google Scholar] [CrossRef]
- Bot, J.L.; Jeannequin, B.; Fabre, R. Growth and nitrogen status of soilless tomato plants following nitrate withdrawal from the nutrient solution. Ann. Bot. 2001, 88, 361–370. [Google Scholar] [CrossRef]
- European Food Safety Authority. Nitrate in vegetables-scientific opinion of the panel on contaminants in the food chain. EFSA J. 2008, 6, 689. [Google Scholar] [CrossRef]
- Colla, G.; Kim, H.J.; Kyriacou, M.C.; Rouphael, Y. Nitrate in fruits and vegetables. Sci. Hortic. 2018, 237, 221–238. [Google Scholar] [CrossRef]
- European Union. Commission Regulation (EU) No 1258/2011 of 2 December 2011 amending Regulation (EC) No 1881/2006 as regards maximum levels for nitrates in foodstuffs Text with EEA relevance. Off. J. Eur. Union 2011, 54, 15–17. [Google Scholar]
- Santamaria, P.; Elia, A.; Parente, A.; Serio, F. Fertilization strategies for lowering nitrate content in leafy vegetables: Chicory and rocket salad cases. J. Plant. Nutr. 1998, 21, 1791–1803. [Google Scholar] [CrossRef]
- Santamaria, P.; Gonnella, M.; Elia, A.; Parente, A.; Serio, F. Ways of reducing rocket salad nitrate content. Acta Hortic. 2001, 548, 529–536. [Google Scholar] [CrossRef]
- Carillo, P.; Kyriacou, M.C.; El-Nakhel, C.; Pannico, A.; dell’Aversana, E.; D’Amelia, L.; Colla, G.; Caruso, G.; De Pascale, S.; Rouphael, Y. Sensory and functional quality characterization of protected designation of origin ‘Piennolo del Vesuvio’ cherry tomato landraces from Campania-Italy. Food Chem. 2019, 292, 166–175. [Google Scholar] [CrossRef]
- Corrado, G.; Formisano, L.; De Micco, V.; Pannico, A.; Giordano, M.; El-Nakhel, C.; Chiaiese, P.; Sacchi, R.; Rouphael, Y. Understanding the morpho-anatomical, physiological, and functional response of sweet basil to isosmotic nitrate to chloride ratios. Biology 2020, 9, 158. [Google Scholar] [CrossRef]
- Chatzigianni, M.; Ntatsi, G.; Theodorou, M.; Stamatakis, A.; Livieratos, I.; Rouphael, Y.; Savvas, D. Functional quality, mineral composition and biomass production in hydroponic spiny chicory (Cichorium spinosum L.) are modulated interactively by ecotype, salinity and nitrogen supply. Front. Plant. Sci. 2019, 10, 1040. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant. Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Formisano, L.; Ciriello, M.; El-Nakhel, C.; De Pascale, S.; Rouphael, Y. Dataset on the effects of anti-insect nets of different porosity on mineral and organic acids profile of Cucurbita pepo L. fruits and leaves. Data 2021, 6, 50. [Google Scholar] [CrossRef]
- Pannico, A.; El-Nakhel, C.; Kyriacou, M.C.; Giordano, M.; Stazi, S.R.; De Pascale, S.; Rouphael, Y. Combating micronutrient deficiency and enhancing food functional quality through selenium fortification of select lettuce genotypes grown in a closed soilless system. Front. Plant. Sci. 2019, 10, 1495. [Google Scholar] [CrossRef] [PubMed]
- Kyriacou, M.C.; El-Nakhel, C.; Graziani, G.; Pannico, A.; Soteriou, G.A.; Giordano, M.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Functional quality in novel food sources: Genotypic variation in the nutritive and phytochemical composition of thirteen microgreens species. Food Chem. 2019, 277, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Kampfenkel, K.; Van Montagu, M.; Inzé, D. Extraction and determination of ascorbate and dehydroascorbate from plant tissue. Anal. Biochem. 1995, 225, 165–167. [Google Scholar] [CrossRef]
- Rouphael, Y.; Corrado, G.; Colla, G.; De Pascale, S.; Dell’Aversana, E.; D’Amelia, L.I.; Fusco, G.M.; Carillo, P. Biostimulation as a means for optimizing fruit phytochemical content and functional quality of tomato landraces of the San Marzano area. Foods 2021, 10, 926. [Google Scholar] [CrossRef] [PubMed]
- Dell’Aversana, E.; Hessini, K.; Ferchichi, S.; Fusco, G.M.; Woodrow, P.; Ciarmiello, L.F.; Abdelly, C.; Carillo, P. Salinity duration differently modulates physiological parameters and metabolites profile in roots of two contrasting barley genotypes. Plants 2021, 10, 307. [Google Scholar] [CrossRef] [PubMed]
- Fogliano, V.; Verde, V.; Randazzo, G.; Ritieni, A. Method for measuring antioxidant activity and its application to monitoring the antioxidant capacity of wines. J. Agric. Food Chem. 1999, 47, 1035–1040. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, N.; Ke, R.; Yang, M.; Rice-Evans, C. Screening of dietary carotenoids and carotenoid-rich fruit extracts for antioxidant activities applying 2, 2′-azinobis (3-ethylenebenzothiazoline-6-sulfonic acid radical cation decolorization assay. Methods Enzymol. 1999, 299, 379–389. [Google Scholar]
- Song, P.; Feng, G.; Brooks, J.; Zhou, B.; Zhou, H.; Zhao, Z.; Li, Y. Environmental risk of chlorine-controlled clogging in drip irrigation system using reclaimed water: The perspective of soil health. J. Clean. Prod. 2019, 232, 1452–1464. [Google Scholar] [CrossRef]
- Lonigro, A.; Montemurro, N.; Laera, G. Effects of residual disinfectant on soil and lettuce crop irrigated with chlorinated water. Sci. Total Environ. 2017, 584, 595–602. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C.; Petropoulos, S.A.; De Pascale, S.; Colla, G. Improving vegetable quality in controlled environments. Sci. Hortic. 2018, 234, 275–289. [Google Scholar] [CrossRef]
- Mou, B. Nutrient content of lettuce and its Improvement. Curr. Nutr. Food Sci. 2009, 5, 242–248. [Google Scholar] [CrossRef]
- Fallovo, C.; Rouphael, Y.; Rea, E.; Battistelli, A.; Colla, G. Nutrient solution concentration and growing season affect yield and quality of Lactuca sativa L. var. acephala in floating raft culture. J. Sci. Food Agric. 2009, 89, 1682–1689. [Google Scholar] [CrossRef]
- Genuncio, G.d.C.; Gomes, M.; Ferrari, A.C.; Majerowicz, N.; Zonta, E. Hydroponic lettuce production in different concentrations and flow rates of nutrient solution. Hortic. Bras. 2012, 30, 526–530. [Google Scholar] [CrossRef]
- Samborska, I.A.; Kalaji, H.M.; Sieczko, L.; Goltsev, V.; Borucki, W.; Jajoo, A. Structural and functional disorder in the photosynthetic apparatus of radish plants under magnesium deficiency. Funct. Plant. Biol. 2018, 45, 668–679. [Google Scholar] [CrossRef]
- Hawkesford, M.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Møller, I.S.; White, P. Functions of macronutrients. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2011; pp. 135–189. ISBN 9780123849052. [Google Scholar]
- Rouphael, Y.; Kyriacou, M.C. Enhancing quality of fresh vegetables through salinity eustress and biofortification applications facilitated by soilless cultivation. Front. Plant. Sci. 2018, 9, 1254. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Cardarelli, M.; Svecova, E.; Rea, E.; Lucini, L. Effects of saline stress on mineral composition, phenolic acids and flavonoids in leaves of artichoke and cardoon genotypes grown in floating system. J. Sci. Food Agric. 2013, 93, 1119–1127. [Google Scholar] [CrossRef]
- Tegeder, M.; Masclaux-Daubresse, C. Source and sink mechanisms of nitrogen transport and use. New Phytol. 2018, 217, 35–53. [Google Scholar] [CrossRef] [PubMed]
- Gerendás, J.; Führs, H. The significance of magnesium for crop quality. Plant. Soil 2013, 368, 101–128. [Google Scholar] [CrossRef]
- Schreiber, U.; Bilger, W.; Neubauer, C. Chlorophyll fluorescence as a nonintrusive indicator for rapid assessment of in vivo photosynthesis. In Ecophysiology of Photosynthesis; Schulze, E., Caldwell, M., Eds.; Springer: Heidelberg, Germany, 1995; pp. 49–70. [Google Scholar]
- Lin, K.H.; Huang, M.Y.; Huang, W.D.; Hsu, M.H.; Yang, Z.W.; Yang, C.M. The effects of red, blue, and white light-emitting diodes on the growth, development, and edible quality of hydroponically grown lettuce (Lactuca sativa L. var. capitata). Sci. Hortic. 2013, 150, 86–91. [Google Scholar] [CrossRef]
- Rambo, L.; Ma, B.L.; Xiong, Y.; da Silvia, P.R.F. Leaf and canopy optical characteristics as crop-n-status indicators for field nitrogen management in corn. J. Plant. Nutr. Soil Sci. 2010, 173, 434–443. [Google Scholar] [CrossRef]
- Mahlangu, R.I.S.; Maboko, M.M.; Sivakumar, D.; Soundy, P.; Jifon, J. Lettuce (Lactuca sativa L.) growth, yield and quality response to nitrogen fertilization in a non-circulating hydroponic system. J. Plant. Nutr. 2016, 39, 1766–1775. [Google Scholar] [CrossRef]
- Fallovo, C.; Rouphael, Y.; Cardarelli, M.; Rea, E.; Battistelli, A.; Colla, G. Yield and quality of leafy lettuce in response to nutrient solution composition and growing season. J. Food Agric. Environ. 2009, 7, 456–462. [Google Scholar]
- Pathare, P.B.; Opara, U.L.; Al-Said, F.A.J. Colour measurement and analysis in fresh and processed foods: A review. Food Bioprocess. Technol. 2013, 6, 36–60. [Google Scholar] [CrossRef]
- Roshanak, S.; Rahimmalek, M.; Goli, S.A.H. Evaluation of seven different drying treatments in respect to total flavonoid, phenolic, vitamin C content, chlorophyll, antioxidant activity and color of green tea (Camellia sinensis or C. assamica) leaves. J. Food Sci. Technol. 2016, 53, 721–729. [Google Scholar] [CrossRef]
- Gershenzon, J. Changes in the levels of plant secondary metabolites under water and nutrient stress. In Phytochemical Adaptations to Stress; Springer: Heidelberg, Germany, 1984; pp. 273–320. [Google Scholar]
- Ramakrishna, A.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant. Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [CrossRef]
- Rouphael, Y.; Cardarelli, M.; Lucini, L.; Rea, E.; Colla, G. Nutrient solution concentration affects growth, mineral composition, phenolic acids, and flavonoids in leaves of artichoke and cardoon. HortScience 2012, 47, 1424–1429. [Google Scholar] [CrossRef]
- Pannico, A.; Graziani, G.; El-Nakhel, C.; Giordano, M.; Ritieni, A.; Kyriacou, M.C.; Rouphael, Y. Nutritional stress suppresses nitrate content and positively impacts ascorbic acid concentration and phenolic acids profile of lettuce microgreens. Italus Hortus 2020, 27, 41–52. [Google Scholar] [CrossRef]
- Baslam, M.; Morales, F.; Garmendia, I.; Goicoechea, N. Nutritional quality of outer and inner leaves of green and red pigmented lettuces (Lactuca sativa L.) consumed as salads. Sci. Hortic. 2013, 151, 103–111. [Google Scholar] [CrossRef]
- Yoshida, T.; Sakuma, K.; Kumagai, H. Nutritional and taste characteristics of low-potassium lettuce developed for patients with chronic kidney diseases. Hong Kong J. Nephrol. 2014, 16, 42–45. [Google Scholar] [CrossRef][Green Version]
- Mondal, M.F.; Asaduzzaman, M.; Ueno, M.; Kawaguchi, M.; Yano, S.; Ban, T.; Tanaka, H.; Asao, T. Reduction of potassium (K) content in strawberry fruits through KNO3 management of hydroponics. Hortic. J. 2016, 86, 26–36. [Google Scholar] [CrossRef]
- Ogawa, A.; Eguchi, T.; Toyofuku, K. Cultivation methods for leafy vegetables and tomatoes with low potassium content for dialysis patients. Environ. Control. Biol. 2012, 50, 407–414. [Google Scholar] [CrossRef]
- Atsushi, O. Cultivation methods for leafy vegetables and tomatoes with low potassium content for dialysis patients and the change of those qualities. Potassium Improv. Qual. Fruits Veg. Hydroponic Nutr. Manag. 2019, 69, 2–17. [Google Scholar] [CrossRef]
- Asao, T.; Asaduzzaman, M.; Mondal, M.F.; Tokura, M.; Adachi, F.; Ueno, M.; Kawaguchi, M.; Yano, S.; Ban, T. Impact of reduced potassium nitrate concentrations in nutrient solution on the growth, yield and fruit quality of melon in hydroponics. Sci. Hortic. 2013, 164, 221–231. [Google Scholar] [CrossRef]
- Asaduzzaman, M.; Talukder, M.R.; Tanaka, H.; Ueno, M.; Kawaguchi, M.; Yano, S.; Ban, T.; Asao, T. Production of low-potassium content melon through hydroponic nutrient management using perlite substrate. Front. Plant. Sci. 2018, 9, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Colonna, E.; Rouphael, Y.; Barbieri, G.; De Pascale, S. Nutritional quality of ten leafy vegetables harvested at two light intensities. Food Chem. 2016, 199, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Petropoulos, S.A.; Constantopoulou, E.; Karapanos, I.; Akoumianakis, C.A.; Passam, H.C. Diurnal variation in the nitrate content of parsley foliage. Int. J. Plant. Prod. 2011, 5, 431–438. [Google Scholar]
- Santamaria, P. Nitrate in vegetables: Toxicity, content, intake and EC regulation. J. Sci. Food Agric. 2006, 86, 10–17. [Google Scholar] [CrossRef]
- Cavaiuolo, M.; Ferrante, A. Nitrates and glucosinolates as strong determinants of the nutritional quality in rocket leafy salads. Nutrients 2014, 6, 1519–1538. [Google Scholar] [CrossRef]
- Gent, M.P.N. Rate of change of composition of lettuce in response to nitrogen depletion or re-supply. J. Sci. Food Agric. 2012, 92, 3007–3015. [Google Scholar] [CrossRef]
- Scheible, W.R.; González-Fontes, A.; Lauerer, M.; Müller-Röber, B.; Caboche, M.; Stitt, M. Nitrate acts as a signal to induce organic acid metabolism and repress starch metabolism in tobacco. Plant. Cell 1997, 9, 783–798. [Google Scholar] [CrossRef]
- Atkinson, N.J.; Urwin, P.E. The interaction of plant biotic and abiotic stresses: From genes to the field. J. Exp. Bot. 2012, 63, 3523–3544. [Google Scholar] [CrossRef] [PubMed]
- Becker, C.; Urlić, B.; Špika, M.J.; Kläring, H.P.; Krumbein, A.; Baldermann, S.; Ban, S.G.; Perica, S.; Schwarz, D. Nitrogen limited red and green leaf lettuce accumulate flavonoid glycosides, caffeic acid derivatives, and sucrose while losing chlorophylls, ?-carotene and xanthophylls. PLoS ONE 2015, 10, e0142867. [Google Scholar] [CrossRef]
- Sudhakar, C.; Lakshmi, A.; Giridarakumar, S. Changes in the antioxidant enzyme efficacy in two high yielding genotypes of mulberry (Morus alba L.) under NaCl salinity. Plant. Sci. 2001, 161, 613–619. [Google Scholar] [CrossRef]
- Formisano, L.; El-Nakhel, C.; Corrado, G.; De Pascale, S.; Rouphael, Y. Biochemical, physiological, and productive response of greenhouse vegetables to suboptimal growth environment induced by insect nets. Biology 2020, 9, 432. [Google Scholar] [CrossRef]
- Carillo, P. GABA shunt in durum wheat. Front. Plant. Sci. 2018, 9, 100. [Google Scholar] [CrossRef]
- Annunziata, M.G.; Ciarmiello, L.F.; Woodrow, P.; Dell’aversana, E.; Carillo, P. Spatial and temporal profile of glycine betaine accumulation in plants under abiotic stresses. Front. Plant. Sci. 2019, 10, 230. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant. Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant. Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Bown, A.W.; Shelp, B.J. Does the GABA shunt regulate cytosolic GABA? Trends Plant. Sci. 2020, 25, 422–424. [Google Scholar] [CrossRef]
- Bown, A.W.; Shelp, B.J. Plant GABA: Not just a Metabolite. Trends Plant. Sci. 2016, 21, 811–813. [Google Scholar] [CrossRef] [PubMed]
- Michaeli, S.; Fromm, H. Closing the loop on the GABA shunt in plants: Are GABA metabolism and signaling entwined? Front. Plant. Sci. 2015, 6, 419. [Google Scholar] [CrossRef]
- Takayama, M.; Ezura, H. How and why does tomato accumulate a large amount of GABA in the fruit? Front. Plant. Sci. 2015, 6, 612. [Google Scholar] [CrossRef]
- Woodrow, P.; Ciarmiello, L.F.; Annunziata, M.G.; Pacifico, S.; Iannuzzi, F.; Mirto, A.; D’Amelia, L.; Dell’Aversana, E.; Piccolella, S.; Fuggi, A.; et al. Durum wheat seedling responses to simultaneous high light and salinity involve a fine reconfiguration of amino acids and carbohydrate metabolism. Physiol. Plant. 2017, 159, 290–312. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Days of Deprivation (DD) | Leaf Area | Leaf Number | Fresh Yield | Dry Biomass | Dry Matter |
|---|---|---|---|---|---|
| (cm2 Plant−1) | (no. Plant) | (g Plant−1) | (g Plant−1) | (%) | |
| 0 | 1481 ± 13 a | 17.61 ± 0.20 | 100.2 ± 0.45 a | 3.76 ± 0.15 a | 3.71 ± 0.14 |
| 3 | 1435 ± 27 a | 17.33 ± 0.17 | 95.42 ± 0.85 b | 3.42 ± 0.01 ab | 3.54 ± 0.04 |
| 6 | 1312 ± 23 b | 17.06 ± 0.29 | 86.98 ± 0.97 c | 3.37 ± 0.04 b | 3.80 ± 0.09 |
| Significance | ** | ns | *** | * | ns |
| Days of Deprivation (DD) | L | a | b | Chroma | Hue Angle |
|---|---|---|---|---|---|
| 0 | 47.61 ± 0.43 a | −9.80 ± 0.20 b | 32.63 ± 0.53 a | 34.08 ± 0.56 a | 253.3 ± 0.07 |
| 3 | 47.44 ± 0.28 a | −9.07 ± 0.03 b | 30.73 ± 0.21 a | 32.06 ± 0.21 a | 253.6 ± 0.04 |
| 6 | 44.25 ± 0.84 b | −7.74 ± 0.32 a | 27.23 ± 0.78 b | 28.32 ± 0.83 b | 254.2 ± 0.34 |
| Significance | ** | ** | *** | *** | ns |
| Days of Deprivation (DD) | Total Chlorophylls | Carotenoids | Chlorophyll a/b Ratio | Chlorophyll/Carotenoids Ratio | SPAD Index | Fv/Fm | H2O2 |
|---|---|---|---|---|---|---|---|
| (mg g−1 fw) | (mg g−1 fw) | (µmol g−1 dw) | |||||
| 0 | 0.740 ± 0.03 | 0.238 ± 0.01 | 2.05 ± 0.10 | 3.11 ± 0.03 | 27.08 ± 0.52 a | 0.843 ± 0.00 a | 35.16 ± 3.63 b |
| 3 | 0.749 ± 0.03 | 0.248 ± 0.01 | 2.33 ± 0.01 | 3.02 ± 0.08 | 25.27 ± 0.23 b | 0.834 ± 0.00 b | 37.04 ± 3.21 b |
| 6 | 0.828 ± 0.06 | 0.268 ± 0.01 | 2.33 ± 0.10 | 3.09 ± 0.14 | 26.39 ± 0.25 ab | 0.829 ± 0.00 b | 64.69 ± 2.46 a |
| Significance | ns | ns | ns | ns | * | ** | ** |
| Days of Deprivation (DD) | N | p | K | S |
|---|---|---|---|---|
| (mg g−1 dw) | (mg g−1 dw) | (mg g−1 dw) | (mg g−1 dw) | |
| 0 | 46.40 ± 1.28 a | 5.90 ± 0.20 ab | 104.6 ± 0.37 a | 1.67 ± 0.07 |
| 3 | 43.95 ± 0.07 ab | 6.62 ± 0.07 a | 102.8 ± 1.37 a | 1.85 ± 0.01 |
| 6 | 41.11 ± 0.95 b | 5.37 ± 0.33 b | 75.02 ± 1.84 b | 1.72 ± 0.10 |
| Significance | * | * | *** | ns |
| Days of Deprivation (DD) | Malate | Tartrate | Oxalate | Citrate | Isocitrate | Glucose | Fructose | Sucrose | Starch |
|---|---|---|---|---|---|---|---|---|---|
| (mg g−1 dw) | (mg g−1 dw) | (mg g−1 dw) | (mg g−1 dw) | (mg g−1 dw) | (µmol g−1 dw) | (µmol g−1 dw) | (µmol g−1 dw) | (µmol g−1 dw) | |
| 0 | 30.35 ± 0.87 a | 2.78 ± 0.08 a | 2.00 ± 0.06 a | 6.19 ± 0.19 | 1.03 ± 0.08 a | 331.9 ± 3.02 b | 291.3 ± 3.82 | 46.35 ± 5.93 b | 44.37 ± 1.01 b |
| 3 | 28.66 ± 0.64 ab | 2.38 ± 0.04 b | 1.63 ± 0.02 b | 5.72 ± 0.36 | 0.74 ± 0.06 b | 341.6 ± 0.67 ab | 291.2 ± 4.73 | 64.36 ± 0.63 a | 53.01 ± 1.05 a |
| 6 | 26.74 ± 0.41 b | 2.49 ± 0.02 b | 1.83 ± 0.06 ab | 5.49 ± 0.45 | 0.60 ± 0.04 b | 360.4 ± 7.63 a | 284.1 ± 1.09 | 61.86 ± 1.13 a | 50.61 ± 0.71 a |
| Significance | * | ** | ** | ns | ** | * | ns | * | ** |
| Chemical Compounds | Days of Deprivation (DD) | |||
|---|---|---|---|---|
| 0 | 3 | 6 | Sig. | |
| Soluble proteins (mg g−1 dw) | 6.99 ± 0.17 a | 6.70 ± 0.32 a | 5.13 ± 0.37 b | ** |
| Free amino acids (µmol g−1 dw) | ||||
| Ala | 6.92 ± 0.17 | 6.47 ± 0.07 | 6.94 ± 0.38 | ns |
| Arg | 3.22 ± 0.07 ab | 3.06 ± 0.21 b | 4.07 ± 0.32 a | * |
| Asn | 73.68 ± 0.34 a | 72.26 ± 0.55 a | 64.30 ± 2.61 b | * |
| Asp | 2.16 ± 0.02 | 2.11 ± 0.12 | 1.89 ± 0.24 | ns |
| GABA | 4.19 ± 0.10 c | 5.37 ± 0.39 b | 7.08 ± 0.08 a | *** |
| Gln | 50.83 ± 0.66 a | 48.54 ± 1.51 ab | 45.58 ± 0.88 b | * |
| Glu | 16.43 ± 0.15 a | 15.16 ± 0.37 b | 13.93 ± 0.28 c | ** |
| Gly | 0.49 ± 0.01 b | 0.56 ± 0.02 ab | 0.65 ± 0.03 a | ** |
| His | 0.60 ± 0.01 | 0.62 ± 0.04 | 0.67 ± 0.04 | ns |
| Ile | 2.33 ± 0.04 b | 2.31 ± 0.04 b | 2.76 ± 0.14 a | * |
| Leu | 2.21 ± 0.03 b | 2.34 ± 0.11 ab | 2.72 ± 0.13 a | * |
| Lys | 0.75 ± 0.02 | 0.76 ± 0.04 | 0.87 ± 0.05 | ns |
| MEA | 4.96 ± 0.10 | 4.66 ± 0.29 | 5.45 ± 0.30 | ns |
| Met | 0.05 ± 0.00 | 0.04 ± 0.00 | 0.06 ± 0.01 | ns |
| Orn | 0.58 ± 0.01 | 0.56 ± 0.02 | 0.58 ± 0.02 | ns |
| Phe | 1.39 ± 0.01 | 1.36 ± 0.08 | 1.39 ± 0.03 | ns |
| Pro | 3.41 ± 0.14 | 3.20 ± 0.04 | 3.17 ± 0.18 | ns |
| Ser | 6.38 ± 0.06 | 5.86 ± 0.23 | 5.33 ± 0.45 | ns |
| Thr | 2.07 ± 0.02 | 1.99 ± 0.05 | 1.67 ± 0.21 | ns |
| Trp | 0.85 ± 0.04 | 0.75 ± 0.05 | 0.71 ± 0.06 | ns |
| Tyr | 3.52 ± 0.09 | 3.54 ± 0.21 | 3.79 ± 0.26 | ns |
| Val | 2.41 ± 0.02 | 2.53 ± 0.14 | 2.80 ± 0.15 | ns |
| Total AA | 189.4 ± 1.00 a | 184.1 ± 2.71 a | 176.4 ± 0.78 b | ** |
| Essential AA | 15.89 ± 0.23 | 15.77 ± 0.73 | 17.73 ± 0.86 | ns |
| BCAAs | 6.95 ± 0.08 b | 7.18 ± 0.28 ab | 8.28 ± 0.42 a | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciriello, M.; Formisano, L.; Pannico, A.; El-Nakhel, C.; Fascella, G.; Duri, L.G.; Cristofano, F.; Gentile, B.R.; Giordano, M.; Rouphael, Y.; et al. Nutrient Solution Deprivation as a Tool to Improve Hydroponics Sustainability: Yield, Physiological, and Qualitative Response of Lettuce. Agronomy 2021, 11, 1469. https://doi.org/10.3390/agronomy11081469
Ciriello M, Formisano L, Pannico A, El-Nakhel C, Fascella G, Duri LG, Cristofano F, Gentile BR, Giordano M, Rouphael Y, et al. Nutrient Solution Deprivation as a Tool to Improve Hydroponics Sustainability: Yield, Physiological, and Qualitative Response of Lettuce. Agronomy. 2021; 11(8):1469. https://doi.org/10.3390/agronomy11081469
Chicago/Turabian StyleCiriello, Michele, Luigi Formisano, Antonio Pannico, Christophe El-Nakhel, Giancarlo Fascella, Luigi Giuseppe Duri, Francesco Cristofano, Beniamino Riccardo Gentile, Maria Giordano, Youssef Rouphael, and et al. 2021. "Nutrient Solution Deprivation as a Tool to Improve Hydroponics Sustainability: Yield, Physiological, and Qualitative Response of Lettuce" Agronomy 11, no. 8: 1469. https://doi.org/10.3390/agronomy11081469
APA StyleCiriello, M., Formisano, L., Pannico, A., El-Nakhel, C., Fascella, G., Duri, L. G., Cristofano, F., Gentile, B. R., Giordano, M., Rouphael, Y., Fusco, G. M., Woodrow, P., & Carillo, P. (2021). Nutrient Solution Deprivation as a Tool to Improve Hydroponics Sustainability: Yield, Physiological, and Qualitative Response of Lettuce. Agronomy, 11(8), 1469. https://doi.org/10.3390/agronomy11081469