
JC Polyomavirus in Prostate Cancer—Friend or Foe?



Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Characteristics of Patients
2.2. Sample Collection
2.3. Tissue Selection
2.4. Isolation and Detection of EBV DNA
2.5. Isolation and Detection BKV and JCV
2.6. Detection of Anti-EBV Antibodies
2.7. Statistical Analysis
2.8. Ethics
3. Results
3.1. Evaluation of the Frequency EBV, JCV, and BKV in the Tumor Tissue of PCa Patients
3.2. Comparison of Patients with Single EBV Infection and Patients with EBV/JCV Co-Infection in the Context of Risk Group, Gleason Score, and TNM Classification
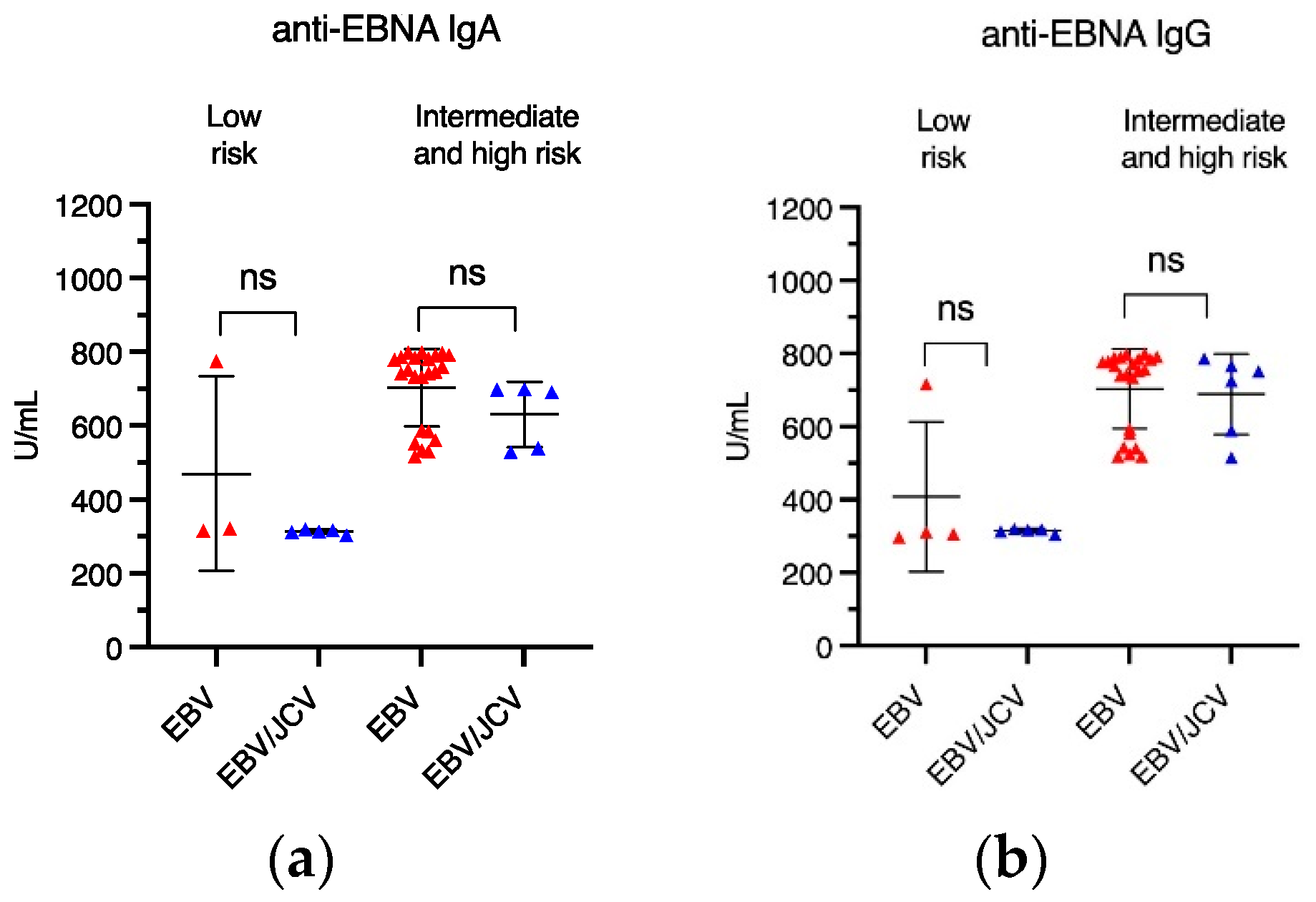
3.3. Frequency of Anti-EBV Antibodies in PCa Patients with EBV Single Infection Compared with EBV/JCV Co-Infection
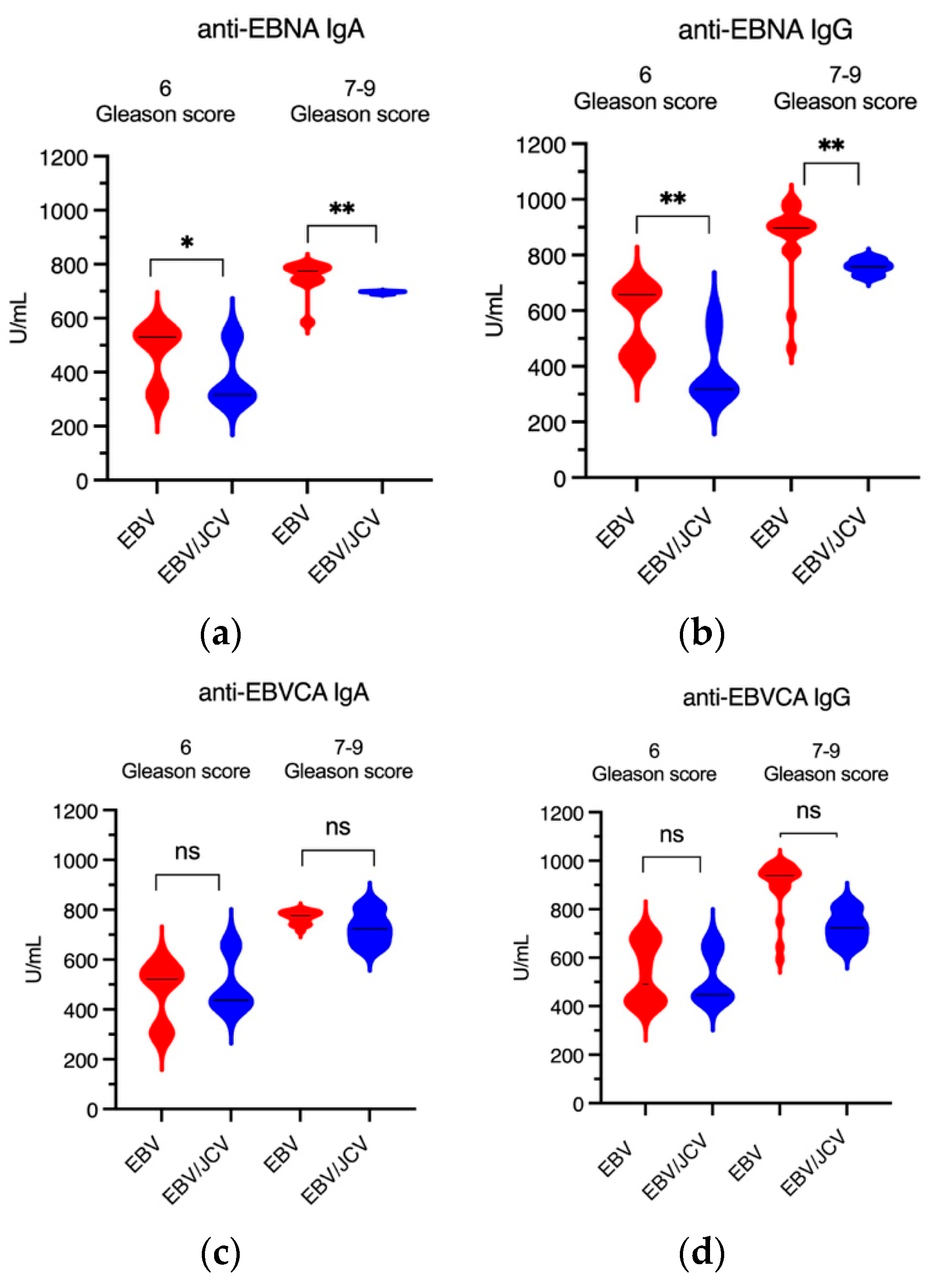
3.4. Antibody Levels for EBVCA IgA and IgG as Well as EBNA1 IgA and IgG in PCa Patients
3.5. Comparison of EBV Load in Tumor Tissue in PCa Patients with Single EBV Infection and in Patients with EBV/JCV Co-Infection
4. Discussion
4.1. Limitation of Our Study
4.2. Future Research Directions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. International Agency for Research of Cancer Global Cancer Observatory. 2022. Available online: https://gco.iarc.who.int/media/globocan/factsheets/cancers/27-prostate-fact-sheet.pdf (accessed on 6 March 2025).
- Wojciechowska, U.; Didkowska, J.A.; Barańska, K.; Miklewska, M.; Michałek, I.; Olasek, P.; Jawołowska, A. Cancer in Poland in 2022; Maria Skłodowska-Curie National Institute of Oncology: Warsaw, Poland, 2024; Available online: https://onkologia.org.pl (accessed on 12 February 2025).
- Huang, L.; La Bonte, M.J.; Craig, S.G.; Finn, S.P.; Allott, E.H. Inflammation and Prostate Cancer: A Multidisciplinary Approach to Identifying Opportunities for Treatment and Prevention. Cancers 2022, 14, 1367. [Google Scholar] [CrossRef] [PubMed]
- Tafuri, A.; Ditonno, F.; Panunzio, A.; Gozzo, A.; Porcaro, A.B.; Verratti, V.; Cerruto, M.A.; Antonelli, A. Prostatic Inflammation in Prostate Cancer: Protective Effect or Risk Factor? Uro 2021, 1, 54–59. [Google Scholar] [CrossRef]
- Maitland, N.J.; Collins, A.T. Inflammation as the Primary Aetiological Agent of Human Prostate Cancer: A Stem Cell Connection? J. Cell Biochem. 2008, 105, 931–939. [Google Scholar] [CrossRef]
- Fernandes, R.; Costa, C.; Fernandes, R.; Barros, A.N. Inflammation in Prostate Cancer: Exploring the Promising Role of Phenolic Compounds as an Innovative Therapeutic Approach. Biomedicines 2023, 11, 3140. [Google Scholar] [CrossRef]
- Gurel, B.; Lucia, M.S.; Thompson, I.M.; Goodman, P.J.; Tangen, C.M.; Kristal, A.R.; Parnes, H.L.; Hoque, A.; Lippman, S.M.; Sutcliffe, S. Chronic inflammation in benign prostate tissue is associated with high-grade prostate cancer in the placebo arm of the prostate cancer prevention trial. Cancer Epidemiol. Prev. Biomark. 2014, 23, 847–856. [Google Scholar] [CrossRef]
- Grisanzio, C.; Signoretti, S. P63 in Prostate Biology and Pathology. J. Cell Biochem. 2008, 103, 1354–1368. [Google Scholar] [CrossRef] [PubMed]
- Vogelstein, B.; Papadopoulos, N.; Velculescu, V.E.; Zhou, S.; Diaz, L.A.; Kinzler, K.W. Cancer Genome Landscapes. Science 2013, 339, 1546–1558. [Google Scholar] [CrossRef] [PubMed]
- Caini, S.; Gandini, S.; Dudas, M.; Bremer, V.; Severi, E.; Gherasim, A. Sexually transmitted infections and prostate cancer risk: A systematic review and meta-analysis. Cancer Epidemiol. 2014, 38, 329–338. [Google Scholar] [CrossRef]
- Akram, N.; Imran, M.; Noreen, M.; Ahmed, F.; Atif, M.; Fatima, Z.; Bilal Waqar, A. Oncogenic role of tumor viruses in humans. Viral Immunol. 2017, 30, 20–27. [Google Scholar] [CrossRef]
- Moore, P.S.; Chang, Y. Why do viruses cause cancer? Highlights of the first century of human tumour virology. Nat. Rev. Cancer 2010, 10, 878–889. [Google Scholar] [CrossRef]
- Chang, Y.; Moore, P.S.; Weiss, R.A. Human oncogenic viruses: Nature and discovery. Phil. Trans. R. Soc. B 2017, 372, 20160264. [Google Scholar] [CrossRef] [PubMed]
- MacLennan, S.A.; Marra, M.A. Oncogenic Viruses and the Epigenome: How Viruses Hijack Epigenetic Mechanisms to Drive Cancer. Int. J. Mol. Sci. 2023, 24, 9543. [Google Scholar] [CrossRef]
- Abumisimir, B.; Mahasneh, I.; Kasmi, Y.; Saif, I.; Hammou, R.A.; Mustafa, M. Prostate Cancer and Viral Infections: Epidemiological and Clinical Indications; Ennaji, M.M., Ed.; Academic Press: Cambridge, MA, USA, 2023; pp. 263–272. [Google Scholar]
- Dalianis, T.; Hirsch, H.H. Human polyomaviruses in disease and cancer. Virology 2013, 437, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Frisque, R.J.; Bream, G.L.; Cannella, M.T. Human polyomavirus JC virus genome. J. Virol. 1984, 51, 458–469. [Google Scholar] [CrossRef]
- Kean, J.M.; Rao, S.; Wang, M.; Garcea, R.L. Seroepidemiology of human polyomaviruses. PLoS Pathog. 2009, 5, e1000363. [Google Scholar] [CrossRef] [PubMed]
- Delbue, S.; Comar, M.; Ferrante, P. Review on the role of the human Polyomavirus JC in the development of tumors. Infect. Agent. Cancer 2017, 12, 10. [Google Scholar] [CrossRef]
- Zambrano, A.; Kalantari, M.; Simoneau, A.; Jensen, J.L.; Villarreal, L.P. Detection of human polyomaviruses and papillomaviruses in prostatic tissue reveals the prostate as a habitat for multiple viral infections. Prostate 2002, 53, 263–276. [Google Scholar] [CrossRef]
- Gorish, B.M.T. JC Polyoma Virus as a Possible Risk Factor for Prostate Cancer Development—Immunofluorescence and Molecular Based Case Control Study. Cancer Control 2022, 29, 10732748221140785. [Google Scholar] [CrossRef]
- Villani, S.; Gagliano, N.; Procacci, P.; Sartori, P.; Comar, M.; Provenzano, M.; Favi, E.; Ferraresso, M.; Ferrante, P.; Delbue, S. Characterization of an in vitro model to study the possible role of polyomavirus BK in prostate cancer. J. Cell Physiol. 2019, 234, 11912–11922. [Google Scholar] [CrossRef]
- Delbue, S.; Matei, D.V.; Carloni, C.; Pecchenini, V.; Carluccio, S.; Villani, S.; Tringali, V.; Brescia, A.; Ferrante, P. Evidence supporting the association of polyomavirus BK genome with prostate cancer. Med. Microbiol. Immunol. 2013, 202, 425–430. [Google Scholar] [CrossRef]
- Ahmed, K.; Sheikh, A.; Fatima, S.; Haider, G.; Ghias, K.; Abbas, F.; Mughal, N.; Abidi, S.H. Detection and characterization of latency stage of EBV and histopathological analysis of prostatic adenocarcinoma tissues. Sci. Rep. 2022, 12, 10399. [Google Scholar] [CrossRef] [PubMed]
- Moens, U.; Prezioso, C.; Pietropaolo, V. Functional Domains of the Early Proteins and Experimental and Epidemiological Studies Suggest a Role for the Novel Human Polyomaviruses in Cancer. Front. Microbiol. 2022, 13, 834368. [Google Scholar] [CrossRef]
- Kiś, J.; Góralczyk, M.; Sikora, D.; Stępień, E.; Drop, B.; Polz-Dacewicz, M. Can the Epstein–Barr Virus Play a Role in the Development of Prostate Cancer? Cancers 2024, 16, 328. [Google Scholar] [CrossRef] [PubMed]
- Epstein, J.I.; Egevad, L.; Amin, M.B.; Delahunt, B.; Srigley, J.R.; Humphrey, P.A.; Grading Committee. The 2014 International Society of Urological Pathology (ISUP) Consensus Conference on Gleason Grading of Prostatic Carcinoma: Definition of Grading Patterns and Proposal for a New Grading System. Am. J. Surg. Pathol. 2016, 40, 244–252. [Google Scholar] [CrossRef]
- Epstein, J.I.; Zelefsky, M.J.; Sjoberg, D.D.; Nelson, J.B.; Egevad, L.; Magi-Galluzzi, C.; Vickers, A.J.; Parwani, A.V.; Reuter, V.E.; Fine, S.W.; et al. A Contemporary Prostate Cancer Grading System: A Validated Alternative to the Gleason Score. Eur. Urol. 2016, 69, 428–435. [Google Scholar] [CrossRef]
- EAU. Guidelines for Prostate Cancer. Available online: https://uroweb.org/guidelines/prostate-cancer/chapter/classification-and-staging-systems (accessed on 8 February 2025).
- Abidi, S.H.; Bilwani, F.; Ghias, K.; Abbas, F. Viral etiology of prostate cancer: Genetic alterations and immune response. A literature review. Int. J. Surg. 2018, 52, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Balis, V.; Sourvinos, G.; Soulitzis, N.; Giannikaki, E.; Sofras, F.; Spandidos, D.A. Prevalence of BK virus and human papillomavirus in human prostate cancer. Int. J. Biol. Markers 2007, 22, 245–251. [Google Scholar] [CrossRef]
- Padgett, B.L.; Zu Rhein, G.M.; Walker, D.L.; Echroade, R.; Dessel, B. Cultivation of papova-like virus from human brain with progressive multifocal leukoencephalopathy. Lancet 1971, 1, 1257–1260. [Google Scholar] [CrossRef]
- Gardner, S.D.; Field, A.M.; Coleman, D.V.; Hulme, B. New human papovavirus (B.K.) isolated from urine after renal transplantation. Lancet 1971, 1, 1253–1257. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans. In Malaria and Some Polyomaviruses (SV40, BK, JC, and Merkel Cell Viruses); International Agency for Research on Cancer: Lyon, France, 2013; p. 104. [Google Scholar]
- Jiang, M.; Abend, J.R.; Johnson, S.F.; Imperiale, M.J. The role of polyomaviruses in human disease. Virology 2009, 384, 266–273. [Google Scholar] [CrossRef]
- Prado, J.C.M.; Monezi, T.A.; Amorim, A.T.; Lino, V.; Paladino, A.; Boccardo, E. Human polyomaviruses and cancer: An overview. Clinics 2018, 73 (Suppl. S1), e558s. [Google Scholar] [CrossRef]
- Anzivino, E.; Rodio, D.M.; Mischitelli, M.; Bellizzi, A.; Sciarra, A.; Salciccia, S.; Gentile, V.; Pietropaolo, V. High Frequency of JCV DNA Detection in Prostate Cancer Tissues. Cancer Genom. Proteom. 2015, 12, 189–200. [Google Scholar]
- Shen, C.; Tung, C.; Chao, C.; Jou, Y.; Huang, S.; Meng, M.; Chang, D.; Chen, P. The differential presence of human polyomaviruses, JCPyV and BKPyV, in prostate cancer and benign prostate hypertrophy tissues. BMC Cancer 2021, 21, 1141. [Google Scholar] [CrossRef]
- Liu, W.; Chen, G.; Gong, X.; Wang, Y.; Zheng, Y.; Liao, X.; Liao, W.; Song, N.L.; Xu, J.; Zhang, X. The diagnostic value of EBV-DNA and EBV-related antibodies detection for nasopharyngeal carcinoma: A metaanalysis. Cancer Cell Int. 2021, 21, 164–177. [Google Scholar] [CrossRef] [PubMed]
- Sivachandran, N.; Wang, X.; Frappier, L. Functions of the Epstein-Barr virus EBNA1 protein in viral reactivation and lytic infection. J. Virol. 2012, 86, 6146–6158. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, S.; Hu, J.; Luo, X.; Li, N.; Bode, A.M.; Cao, Y. Epstein-Barr virus lytic reactivation regulation and its pathogenic role in carcinogenesis. Int. J. Biol. Sci. 2016, 12, 1309–1318. [Google Scholar] [CrossRef]
- Martinez-Fierro, M.L.; Leach, R.J.; Gomez-Guerra, L.S.; Garza-Guajardo, R.; Johnson-Pais, T.; Beuten, J.; Morales-Rodriguez, I.B.; Hernandez-Ordoñez, M.A.; Calderon-Cardenas, G.; Ortiz-Lopez, R.; et al. Identification of viral infections in the prostate and evaluation of their association with cancer. BMC Cancer 2010, 10, 326. [Google Scholar] [CrossRef]
- Mundo, L.; Leoncini, L.; Accardi-Gheit, R. Epstein–Barr Virus Infection in Cancer. Cancers 2023, 15, 4659. [Google Scholar] [CrossRef]
- Wang, L.; Ning, S. New Look of EBV LMP1 Signaling Landscape. Cancers 2021, 13, 5451. [Google Scholar] [CrossRef]
- Chakravorty, S.; Yan, B.; Wang, C.; Wang, L.; Quaid, J.T.; Lin, C.F.; Briggs, S.D.; Majumder, J.; Canaria, D.A.; Chauss, D.; et al. Integrated Pan-Cancer Map of EBV-Associated Neoplasms Reveals Functional Host–Virus Interactions. Cancer Res. 2019, 79, 6010–6023. [Google Scholar] [CrossRef]
- Calcinotto, A.; Spataro, C.; Zagato, E.; Di Mitri, D.; Gil, V.; Crespo, M.; De Bernardis, G.; Losa, M.; Mirenda, M.; Pasquini, E.; et al. IL-23 secreted by myeloid cells drives castration-resistant prostate cancer. Nature 2018, 559, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.M. Viral oncogenes, noncoding RNAs, and RNA splicing in human tumor viruses. Int. J. Biol. Sci. 2010, 6, 730–755. [Google Scholar] [CrossRef]
- Barbier, M.T.; Del Valle, L. Co-Detection of EBV and Human Polyomavirus JCPyV in a Case of AIDS-Related Multifocal Primary Central Nervous System Diffuse Large B-Cell Lymphoma. Viruses 2023, 15, 755. [Google Scholar] [CrossRef] [PubMed]
- Darbinyan, A.; White, M.K.; Akan, S.; Radhakrishnan, S.; Del Valle, L.; Amini, S.; Khalili, K. Alterations of DNA damage repair pathways resulting from JCV infection. Virology 2007, 364, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Del Valle, L.; Gordon, J.; Assimakopoulou, M.; Enam, S.; Geddes, J.F.; Varakis, J.N.; Katsetos, C.D.; Croul, S.; Khalili, K. Detection of JC Virus DNA sequences and expression of the viral regulatory protein T-Antigen in tumors of the Central Nervous System. Cancer Res. 2001, 61, 4287–4293. [Google Scholar]
- Ripple, M.J.; Parker Struckhoff, A.; Trillo-Tinoco, J.; Li, L.; Margolin, D.A.; McGoey, R.; Del Valle, L. Activation of c-Myc and Cyclin D1 by JCV T-Antigen and beta-catenin in colon cancer. PLoS ONE 2014, 9, e106257. [Google Scholar] [CrossRef]
- Nguyen, L.; Papenhausen, P.; Shao, H. The Role of c-MYC in B-Cell Lymphomas: Diagnostic and Molecular Aspects. Genes 2017, 8, 116. [Google Scholar] [CrossRef]
- Escobedo-Bonilla, C.M. Mini Review: Virus Interference: History, Types and Occurrence in Crustaceans. Front. Immunol. 2021, 12, 674216. [Google Scholar] [CrossRef]
- Du, Y.; Wang, C.; Zhang, Y. Viral Coinfections. Viruses 2022, 14, 2645. [Google Scholar] [CrossRef]
- Goto, H.; Ihira, H.; Morishita, K.; Tsuchiya, M.; Ohta, K.; Yumine, N.; Tsurudome, M.; Nishio, M. Enhanced growth of influenza A virus by coinfection with human parainfluenza virus type 2. Med. Microbiol. Immunol. 2016, 205, 209–218. [Google Scholar] [CrossRef]
- Chu, C.-J.; Lee, S.-D. Hepatitis B virus/hepatitis C virus coinfection: Epidemiology, clinical features, viral interactions and treatment. J. Gastroenterol. Hepatol. 2008, 23, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Moens, U.; Van Ghule, M.; Ehlers, B. Are human polyomaviruses co-factors for cancers induced by other oncoviruses? Rev. Med. Virol. 2014, 24, 343–360. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, N.J.; Glenn, W.K.; Sahrudin, A.; Orde, M.M.; Delprado, W.; Lawson, J.S. Human papillomavirus and Epstein Barr virus in prostate cancer: Koilocytes indicate potential oncogenic influences of human papillomavirus in prostate cancer. Prostate 2013, 73, 236–241. [Google Scholar] [CrossRef]
- Nahand, J.S.; Khanaliha, K.; Mirzaei, H.; Moghoofei, M.; Baghi, H.B.; Esghaei, M.; Khatami, A.R.; Fatemipour, M.; Bokharaei-Salim, F. Possible role of HPV/EBV coinfection in anoikis resistance and development in prostate cancer. BMC Cancer 2021, 21, 926. [Google Scholar] [CrossRef] [PubMed]
- Pietropaolo, V.; Prezioso, C.; Bagnato, F.; Antonelli, G. John Cunningham virus: An overview on biology and disease of the etiological agent of the progressive multifocal leukoencephalopathy. New Microbiol. 2018, 41, 179–186. [Google Scholar]
- Ferreira, A.D.; Tayyar, Y.; Idris, A.; McMillan, N.A.J. A “hit-and-run” affair—A possible link for cancer progression in virally driven cancers. Biochim. Biophys. Acta (BBA)—Rev. Cancer 2021, 1875, 188476. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PCa Patients | |||||
|---|---|---|---|---|---|
| EBV Single Infection | EBV/JCV Co-Infection | ||||
| n | % | n | % | ||
| Total | 41 | 35.7 | 16 | 13.9 | |
| Age | <60 | 6 | 14.6 | 1 | 6.3 |
| >60 | 35 | 85.4 | 15 | 93.8 | |
| p | 0.6599 | ||||
| Place of residence | Urban | 28 | 68.3 | 9 | 56.3 |
| Rural | 13 | 31.7 | 7 | 43.8 | |
| p | 0.5379 | ||||
| Smoking | Never | 10 | 24.4 | 2 | 12.5 |
| Ever | 31 | 75.6 | 14 | 87.5 | |
| p | 0.4767 | ||||
| Alcohol abuse | never | 14 | 34.2 | 5 | 31.3 |
| ≤drink per week | 23 | 56.1 | 10 | 62.5 | |
| >drink per week | 4 | 9.8 | 1 | 6.3 | |
| p | 0.9999 | ||||
| PCa Patients | |||||
|---|---|---|---|---|---|
| EBV Single Infection N = 41 | EBV/JCV Co-Infection N = 16 | ||||
| n | % | n | % | ||
| Risk | Low | 10 | 24.4 | 9 | 56.2 |
| Intermediate/high | 31 | 75.6 | 7 | 43.8 | |
| p | 0.0307 * | ||||
| Gleason score | 6 | 17 | 41.5 | 12 | 75.0 |
| 7–9 | 24 | 58.5 | 4 | 25.0 | |
| p | 0.0379 * | ||||
| T | T1 | 11 | 26.8 | 10 | 62.5 |
| T2 | 30 | 73.2 | 6 | 37.5 | |
| T3 | 0 | 0.0 | 0 | 0.0 | |
| T4 | 0 | 0.0 | 0 | 0.0 | |
| p | 0.0166 * | ||||
| N | N0 | 41 | 100.0 | 16 | 100.0 |
| M | M0 | 41 | 100.0 | 16 | 100.0 |
| EBV-Positive | EBV/JCV Co-Infection | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Low Risk | Intermediate/ High Risk | p | Low Risk | Intermediate/ High Risk | p | |||||
| n | (%) | n | (%) | n | (%) | n | (%) | |||
| n = 10 | n = 30 | n = 9 | n = 7 | |||||||
| EBVCA IgA | 7 | 70.00 | 26 | 86.67 | 0.2297 | 5 | 55.56 | 6 | 85.71 | 0.1967 |
| EBVCA IgG | 9 | 90.00 | 25 | 83.33 | 0.9090 | 5 | 55.56 | 6 | 85.71 | 0.1967 |
| EBNA IgA | 3 | 30.00 | 23 | 83.33 | 0.0036 * | 5 | 55.56 | 5 | 71.43 | 0.5153 |
| EBNA IgG | 4 | 40.00 | 23 | 83.33 | 0.0159 * | 5 | 55.56 | 6 | 85.71 | 0.1967 |
| EBV Single Infection | EBV/JCV Co-Infection | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 6 Gleason Score | 7–9 Gleason Score | p | 6 Gleason Score | 7–9 Gleason Score | p | |||||
| n | (%) | n | (%) | n | (%) | n | (%) | |||
| n = 18 | n = 23 | n = 12 | n = 4 | |||||||
| EBVCA IgA | 12 | 66.67 | 13 | 56.52 | 0.5087 | 7 | 58.33 | 4 | 100.00 | 0.1195 |
| EBVCA IgG | 12 | 66.67 | 22 | 95.65 | 0.0313 * | 8 | 66.67 | 3 | 75.00 | 0.7555 |
| EBNA IgA | 8 | 44.40 | 18 | 78.26 | 0.0486 * | 7 | 58.33 | 3 | 75.00 | 0.5510 |
| EBNA IgG | 10 | 55.56 | 17 | 73.91 | 0.2186 | 7 | 58.33 | 4 | 100.00 | 0.1195 |
| EBV Single Infection | EBV/JCV Co-Infection | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| T1 | T2 | p | T1 | T2 | p | |||||
| n | (%) | n | (%) | n | (%) | n | (%) | |||
| n = 11 | n = 30 | n = 10 | n = 6 | |||||||
| EBVCA IgA | 7 | 63.64 | 26 | 86.67 | 0.0992 | 6 | 60.00 | 5 | 83.33 | 0.3296 |
| EBVCA IgG | 9 | 81.82 | 25 | 83.33 | 0.9090 | 5 | 50.00 | 6 | 100.00 | 0.0367 * |
| EBNA IgA | 3 | 27.27 | 23 | 76.67 | 0.0036 * | 6 | 60.00 | 4 | 66.67 | 0.7897 |
| EBNA IgG | 4 | 36.36 | 23 | 76.67 | 0.0159 * | 6 | 60.00 | 5 | 83.33 | 0.3296 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiś, J.; Sikora, D.; Jarosz, M.J.; Polz-Dacewicz, M. JC Polyomavirus in Prostate Cancer—Friend or Foe? Cancers 2025, 17, 1725. https://doi.org/10.3390/cancers17101725
Kiś J, Sikora D, Jarosz MJ, Polz-Dacewicz M. JC Polyomavirus in Prostate Cancer—Friend or Foe? Cancers. 2025; 17(10):1725. https://doi.org/10.3390/cancers17101725
Chicago/Turabian StyleKiś, Jacek, Dominika Sikora, Mirosław J. Jarosz, and Małgorzata Polz-Dacewicz. 2025. "JC Polyomavirus in Prostate Cancer—Friend or Foe?" Cancers 17, no. 10: 1725. https://doi.org/10.3390/cancers17101725
APA StyleKiś, J., Sikora, D., Jarosz, M. J., & Polz-Dacewicz, M. (2025). JC Polyomavirus in Prostate Cancer—Friend or Foe? Cancers, 17(10), 1725. https://doi.org/10.3390/cancers17101725