

TAMs and PD-1 Networking in Gastric Cancer: A Review of the Literature

 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Search Strategy
2.2. Study Inclusion and Exclusion Criteria
2.3. Article Selection and Data Extraction
3. Results
3.1. Characteristics of Included Studies
3.2. Correlation of TAMs and PD-L1
3.3. Prognosis of PD-1+ TAMs
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- International Agency for Research on Cancer; World Health Organization. The Global Cancer Observatory. Stomach 2020. Available online: https://www.wcrf.org/ (accessed on 10 December 2023).
- González, C.A.; Agudo, A. Chapter 1-Epidemiology, Risk Factors and Pathogenesis, ESMO Gastrointestinal Tract Tumours 2021. Available online: https://oncologypro.esmo.org (accessed on 12 December 2023).
- Rihawi, K.; Ricci, A.D.; Rizzo, A.; Brocchi, S.; Marasco, G.; Pastore, L.V.; Lorena, F.; Llimpe, R.; Golfieri, R. Tumor-Associated Macrophages and Inflammatory Microenvironment in Gastric Cancer: Novel Translational Implications. Int. J. Mol. Sci. 2021, 22, 3805. [Google Scholar] [CrossRef] [PubMed]
- Taieb, J.; Bennouna, J.; Llorca, F.P.; Basile, D.; Samalin, E.; Zaanan, A. Treatment of gastric adenocarcinoma: A rapidly evolving landscape. Eur. J. Cancer 2023, 195, 113370. [Google Scholar] [CrossRef] [PubMed]
- Guan, W.L.; He, Y.; Xu, R.H. Gastric cancer treatment: Recent progress and future perspectives. J. Hematol. Oncol. 2023, 16, 57. [Google Scholar] [CrossRef] [PubMed]
- Depotte, L.; Palle, J.; Rasola, C.; Broudin, C.; Afrăsânie, V.A.; Mariani, A.; Zaanan, A. New developments and standard of care in the management of advanced gastric cancer. Clin. Res. Hepatol. Gastroenterol. 2023, 11, 102245. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Bai, Y.; Shen, M.; Li, Y. Therapeutic strategies for gastric cancer targeting immune cells: Future directions. Front. Immunol. 2022, 13, 992762. [Google Scholar] [CrossRef]
- Hashimoto, I.; Oshima, T. Claudins and Gastric Cancer: An Overview. Cancers 2022, 14, 290. [Google Scholar] [CrossRef]
- Li, J. Targeting claudins in cancer: Diagnosis, prognosis and therapy. Am. J. Cancer Res. 2021, 11, 3406–3424. [Google Scholar]
- Giulia Grizzi, G.; Venetis, K.; Denaro, N.; Bonomi, M.; Celotti, A.; Pagkali, A.; Hahne, J.C.; Tomasello, G.; Petrelli, F.; Fusco, N.; et al. Anti-Claudin Treatments in Gastroesophageal Adenocarcinoma Mainstream and Upcoming Strategies. J. Clin. Med. 2023, 12, 2973. [Google Scholar] [CrossRef]
- Shah, M.A.; Shitara, K.; Ajani, J.A.; Bang, Y.J.; Enzinger, P.; Ilson, D.; Lordick, F.; Van Cutsem, E.; Plazas, J.G.; Huang, J.; et al. Zolbetuximab plus CAPOX in CLDN18.2-positive gastric or gastroesophageal junction adenocarcinoma: The randomized, phase 3 GLOW trial. Nat. Med. 2023, 29, 2133–2141. [Google Scholar] [CrossRef]
- Janjigian, Y.Y.; Shitara, K.; Moehler, M.; Garrido, M.; Salman, P.; Shen, L.; Wyrwicz, L.; Yamaguchi, K.; Skoczylas, T.; Bragagnoli, A.C.; et al. Nivolumab plus chemotherapy versus chemotherapy as first line treatment for advanced gastric cancer/gastroesophageal junction cancer/esophageal adenocarcinoma (CheckMate 649): A multicenter, randomized, open-label, phase 3 trial. Lancet 2021, 398, 27–40. [Google Scholar] [CrossRef]
- Bang, Y.J.; Kang, Y.K.; Catenacci, D.V.; Muro, K.; Fuchs, C.F.; Geva, R.; Hara, H.; Golan, T.; Garrido, M.; Jalal, S.I.; et al. Pembrolizumab alone or in combination with chemotherapy as first line therapy for patients with advanced gastric or gastroesophageal junction adenocarcinoma: Results from the phase II nonrandomized KEYNOTE-059 study. Gastric Cancer 2019, 22, 828–837. [Google Scholar] [CrossRef]
- Janjigian, Y.Y.; Kawazoe, A.; Bai, Y.; Xu, J.; Lonardi, S.; Metges, J.P.; Yanez, P.; Wyrwicz, L.S.; Shen, L.; Ostapenko, Y.; et al. Pembrolizumab plus trastuzumab and chemotherapy for HER2-positive gastric or gastro-oesophageal junction adenocarcinoma: Interim analyses from the phase 3 KEYNOTE-811 randomised placebo-controlled trial. Lancet 2023, 402, 10418. [Google Scholar] [CrossRef] [PubMed]
- Mellman, I.; Coukos, G.; Dranoff, G. Cancer immunotherapy comes of age. Nature 2011, 480, 480–489. [Google Scholar] [CrossRef] [PubMed]
- Mazzarella, L.; Duso, B.A.; Trapani, D.; Belli, C.; D’Amico, P.; Ferraro, E.; Viale, G.; Curigliano, G. The evolving landscape of’ next-generation’ immune checkpoint inhibitors: A review. Eur. J. Cancer 2019, 117, 14–31. [Google Scholar] [CrossRef] [PubMed]
- Robert, C.; Thomas, L.; Bondarenko, I.; O’Day, S.; Weber, S.; Garbe, C.; Lebbe, C.; Baurain, J.F.; Testori, A.; Grob, J.J.; et al. Ipilimumab plus dacarbazine for previously untreated metastatic melanoma. N. Engl. J. Med. 2011, 364, 2517–2526. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Chen, M.; Nie, H.; Yuan, Y. PD-1 and PD-L1 in cancer immunotherapy: Clinical implications and future considerations. Hum. Vaccin. Immunother. 2019, 15, 1111–1122. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.; Jiang, W.; Kim, B.Y.S.; Zhang, C.C.; Fu, Y.X.; Weissman, I.L. Phagocytosis checkpoints as new targets for cancer immunotherapy. Nat. Rev. Cancer 2019, 19, 568–586. [Google Scholar] [CrossRef]
- Takei, S.; Kawazoe, A.; Shitara, K. The New Era of Immunotherapy in Gastric Cancer. Cancers 2022, 14, 1054. [Google Scholar] [CrossRef]
- Narita, Y.; Muro, K. Updated Immunotherapy for Gastric Cancer. J. Clin. Med. 2023, 12, 2636. [Google Scholar] [CrossRef]
- Zhao, Q.; Cao, L.; Guan, L.; Bie, L.; Wang, S.; Xie, B.; Chen, X.; Shen, X.; Cao, F. Immunotherapy for gastric cancer: Dilemmas and prospect. Brief. Funct. Genom. 2019, 18, 107–112. [Google Scholar] [CrossRef]
- Oya, Y.; Hayakawa, Y.; Koike, K. Tumor microenvironment in gastric cancers. Cancer Sci. 2020, 111, 2696–2707. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Lu, J.; Lou, J.; Shi, C.; Mo, S.; Shao, Y.; Ni, J.; Zhang, W.; Cheng, X. Gastric Cancer Tumor Microenvironment Characterization Reveals Stromal-Related Gene Signatures Associated With Macrophage Infiltration. Front. Genet. 2020, 11, 663. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.; Zhang, T.; Li, Q.; Chen, X.; Jiang, F.; Shen, X. The tumor immune-microenvironment in gastric cancer. Tumori 2022, 108, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Xiao, T.; Li, M.; Jia, Q. Tumor-associated macrophages: Potential therapeutic targets and diagnostic markers in cancer. Pathol. Res. Pract. 2023, 249. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Yang, Z.; Wang, J.; Wang, Q.; Zhang, H.; Ju, X. Research progress on tumour-associated macrophages in gastric cancer (Review). Oncol. Rep. 2021, 4, 35. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Fushida, S.; Yamamoto, Y.; Tsukada, T.; Kinoshita, J.; Oyama, K.; Miyashita, T.; Tajima, H.; Ninomiya, I.; Munesue, S.; et al. Tumor-associated macrophages of the M2 phenotype contribute to progression in gastric cancer with peritoneal dissemination. Gastric Cancer 2016, 19, 1052–1065. [Google Scholar] [CrossRef]
- Gordon, S.R.; Maute, R.L.; Dulken, B.W.; Hutter, G.; George, B.M.; McCracken, M.N.; Gupta, R.; Tsai, J.M.; Sinha, R.; Daniel Corey, D.; et al. PD-1 expression by tumor associated macrophages inhibits phagocytosis and tumor immunity. Nature 2017, 545, 495–499. [Google Scholar] [CrossRef]
- Bejarano, L.; Jordāo, M.J.C.; Joyce, J.A. Therapeutic Targeting of the Tumor Microenvironment. Cancer Discov. 2021, 11, 933–959. [Google Scholar] [CrossRef]
- Wanderley, C.W.; Colón, D.F.; Luiz, J.P.M.; Oliveira, F.F.; Viacava, P.R.; Leite, C.A.; Pereira, J.A.; Silva, C.M.; Silva, C.R.; Silva, R.L.; et al. Paclitaxel Reduces Tumor Growth by Reprogramming Tumor-Associated Macrophages to an M1 Profile in a TLR4-Dependent Manner. Cancer Res. 2018, 78, 5891–5900. [Google Scholar] [CrossRef]
- Pan, Y.; Yinda, Y.; Wang, X.; Zhang, T. Tumor-Associated Macrophages in Tumor Immunity. Front. Immunol. 2020, 11, 583084. [Google Scholar] [CrossRef]
- Saqib, U.; Sarkar, S.; Suk, K.; Mohammad, O.; Baig, M.S.; Savai, R. Phytochemicals as modulators of M1-M2 macrophages in inflammation. Oncotarget 2018, 9, 17937–17950. [Google Scholar] [CrossRef] [PubMed]
- Jayasingam, S.D.; Citartan, M.; Thang, T.H.; Zin, A.A.M.; Ang, K.C.; Ch’ng, E.S. Evaluating the Polarization of Tumor-Associated Macrophages Into M1 and M2 Phenotypes in Human Cancer Tissue: Technicalities and Challenges in Routine Clinical Practice. Front. Oncol. 2020, 9, 1512. [Google Scholar] [CrossRef] [PubMed]
- Räihä, R.M.; Puolakkainen, P.A. Tumor-associated macrophages (TAMs) as biomarkers for gastric cancer: A review. Chronic Dis. Transl. Med. 2018, 4, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Gambardella, V.; Castillo, J.; Tarazona, N.; Valiente, F.G.; Ciarpaglini, C.M.; Cabeza-Segura, M.; Roselló, S.; Roda, D.; Huerta, M.; Cervantes, A.; et al. The role of tumor-associated macrophages in gastric cancer development and their potential as a therapeutic target. Cancer Treat. Rev. 2020, 86, 102015. [Google Scholar] [CrossRef] [PubMed]
- Piao, H.; Fu, L.; Wang, Y.; Liu, Y.; Wang, Y.; Meng, X.; Yang, D.; Xiao, X.; Zhang, J. A positive feedback loop between gastric cancer cells and tumor-associated macrophage induces malignancy progression. J. Exp. Clin. Cancer Res. 2022, 41, 174. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Xu, D.; Huang, C.; Guo, Y.; Wang, S.; Zhu, C.; Xu, J.; Zhang, Z.; Shen, Y.; Zhao, W.; et al. Regulatory T cells and M2 macrophages present diverse prognostic value in gastric cancer patients with different clinicopathologic characteristics and chemotherapy strategies. J. Transl. Med. 2019, 17, 192. [Google Scholar] [CrossRef] [PubMed]
- Ubukata, Y.; Ogata, K.; Sohda, M.; Yokobori, T.; Shimoda, Y.; Handa, T.; Nakazawa, N.; Kimura, A.; Kogure, N.; Sano, A.; et al. Role of PD-L1 Expression during the Progression of Submucosal. Gastric Cancer. Oncol. 2021, 99, 15–22. [Google Scholar] [CrossRef]
- Ju, X.; Zhang, H.; Zhou, Z.; Chen, M.; Wang, Q. Tumor-associated macrophages induce PD-L1 expression in gastric cancer cells through IL-6 and TNF-α signaling. Exp. Cell Res. 2020, 396, 112315. [Google Scholar] [CrossRef]
- Junttila, A.; Helminen, O.; Väyrynen, J.P.; Ahtiainen, M.; Kenessey, I.; Jalkanen, S.; Mecklin, J.P.; Kellokumpu, I.; Kuopio, T.; Böhm, J.; et al. Immunophenotype based on inflammatory cells, PD-1/PD-L1 signaling pathway and M2 macrophages predicts survival in gastric cancer. Br. J. Cancer 2020, 123, 1625–1632. [Google Scholar] [CrossRef]
- Huang, Y.K.; Wang, M.; Sun, Y.; Di Costanzo, N.; Mitchell, C.; Achuthan, A.; Hamilton, J.A.; Busuttil, R.A.; Boussioutas, A. Macrophage spatial heterogeneity in gastric cancer defined by multiplex immunohistochemistry. Nat. Commun. 2019, 10, 3928. [Google Scholar] [CrossRef]
- Harada, K.; Dong, X.; Estrella, J.S.; Correa, A.M.; Xu, Y.; Hofstetter, W.L.; Sudo, K.; Onodera, H.; Suzuki, K.; Suzuki, A.; et al. Tumor-associated macrophage infiltration is highly associated with PD-L1 expression in gastric adenocarcinoma. Gastric Cancer 2018, 21, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, R.; Cao, Y.; Gu, Y.; Lin, C.; Liu, X.; Lv, K.; He, X.; Fang, H.; Jin, K.; et al. Poor Clinical Outcomes and Immunoevasive Contexture in Intratumoral IL-10-Producing Macrophages Enriched Gastric Cancer Patients. Ann. Surg. 2022, 275, e626–e635. [Google Scholar] [CrossRef] [PubMed]
- Ivanović, T.; Božić, D.; Benzon, B.; Čapkun, V.; Vukojević, K.; Durdov, G.M. Histological Type, Cytotoxic T Cells and Macrophages in the Tumor Microenvironment Affect the PD-L1 Status of Gastric Cancer. Biomedicines 2023, 11, 709. [Google Scholar] [CrossRef] [PubMed]
- Kono, Y.; Saito, H.; Miyauchi, W.; Shimizu, S.; Murakami, Y.; Shishido, Y.; Miyatani, K.; Matsunaga, T.; Fukumoto, Y.; Nakayama, Y.; et al. Increased PD-1-positive macrophages in the tissue of gastric cancer are closely associated with poor prognosis in gastric cancer patients. BMC Cancer 2020, 20, 175. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Li, B.; Wei, Y.; Zhao, Y.; Wang, L.; Zhang, P.; Yang, J.; He, W.; Chen, H.; Jiao, Z.; et al. Tumor-derived exosomes induce PD1+ macrophage population in human gastric cancer that promotes disease progression. Oncogenesis 2018, 7, 41. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Zhang, J.; Fan, X.; Zheng, Z.; Jiang, X.; Chen, D.; Lu, Y.; Li, Y.; Wang, M.; Hu, M.; et al. Immune Profiling in Gastric Cancer Reveals the Dynamic Landscape of Immune Signature Underlying Tumor Progression. Front. Immunol. 2022, 13, 935552. [Google Scholar] [CrossRef]
- Huck, B.R.; Kötzner, L.; Urbahns, K. Small Molecules Drive Big Improvements in ImmunoOncology Therapies. Angew. Chem. Int. Ed. Engl. 2018, 57, 4412–4428. [Google Scholar] [CrossRef]
- Anfray, C.; Ummarino, A.; Andón, F.T.; Allavena, P. Current Strategies to Target Tumor-Associated-Macrophages to Improve Anti-Tumor Immune Responses. Cells 2020, 9, 46. [Google Scholar] [CrossRef]
- Li, M.; He, L.; Zhu, J.; Zhang, P.; Liang, S. Targeting tumor-associated macrophages for cancer treatment. Cell Biosci. 2022, 12, 85. [Google Scholar] [CrossRef]
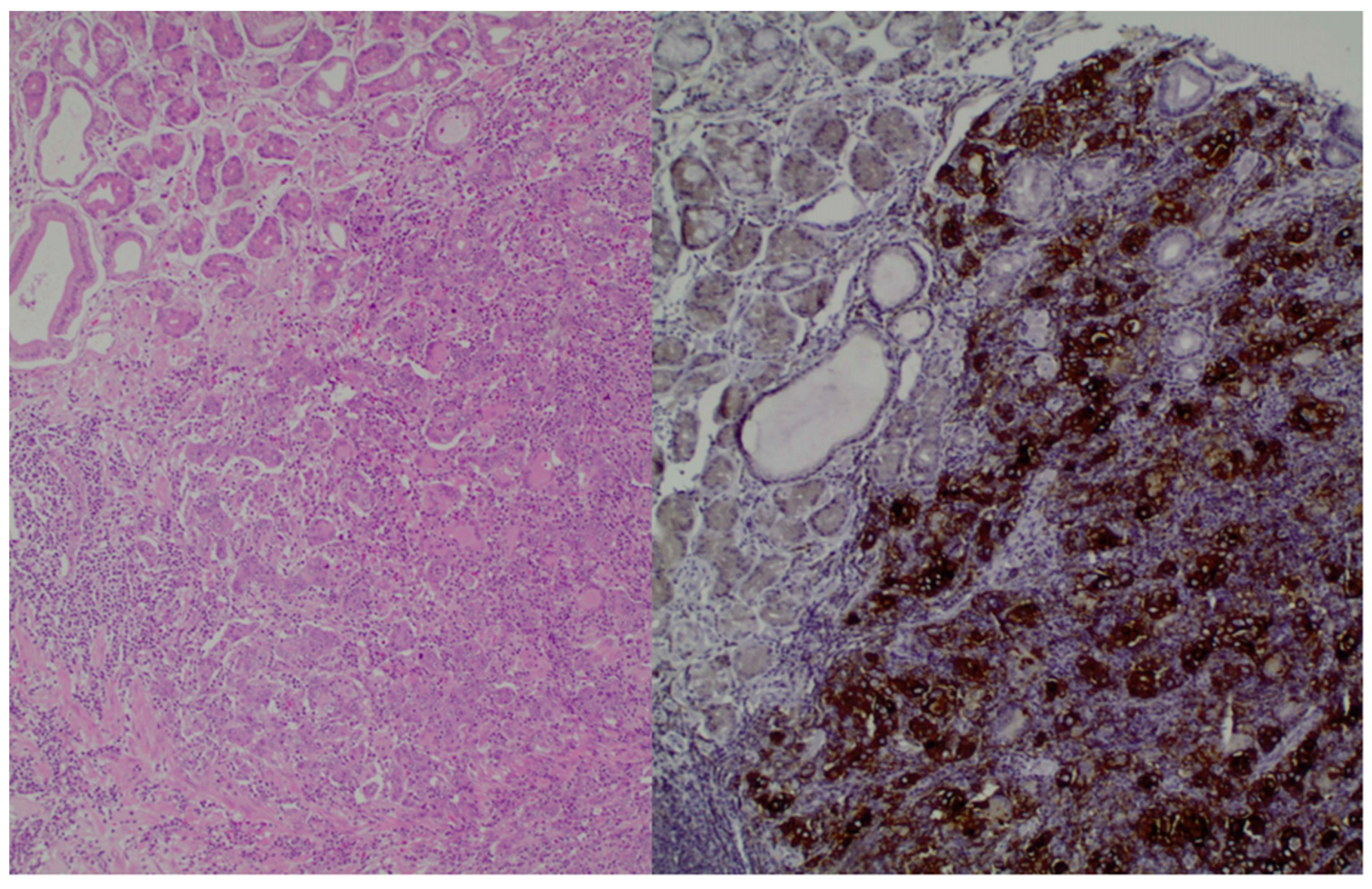


{kind=link}
{kind=link}
| Author | Gender n(%) | Sample Size | Used in the Analysis | T-Stage | Lymphnodes | N-Stage | Metastasis | AJCC Staging 8th Edition | Surgery | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Male | Female | T1 | T2 | T3 | T4 | YES | NO | N0 | N1 | N2 | N3 | YES | NO | I | II | III | IV | YES | NO | ||||
| Correlation of TAMS and PD-L1 | |||||||||||||||||||||||
| 1 | Ubukata Y (2020) [39] | 76 (71) | 31 (29) | 635 | 107 | 107 | 0 | 0 | 0 | 45 | 62 | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | 107 | 0 |
| 2 | Ju X (2020) [40] | NA | NA | 95 | 95 | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA |
| 3 | Junttila A (2020) [41] | 61 (51.7) | 57 (48.3) | 132 | 118 | 34 | 52 | 32 | 4 | 40 | 78 | 40 | 53 | 18 | 7 | 4 | 114 | 32 | 51 | 31 | 4 | 118 | 0 |
| 4 | Huang Y.K (2019) [42] | 36 (64.3) | 20 (35.7) | 56 | 56 | 3 | 13 | 39 | 1 | 33 | 23 | 23 | 18 | 10 | 5 | 3 | 53 | NA | NA | NA | NA | NA | NA |
| 5 | Harada K (2017) [43] | 134 (62) | 82 (38) | 217 | 216 | 31 | 104 | 66 | 14 | 144 | 67 | 67 | 84 | 27 | 33 | 59 | 154 | 67 | 49 | 38 | 59 | 216 | 0 |
| 6 | Zhang H (2022) [44] | 575 (67.5) | 270 (32.5) | 932 | 852 | 110 | 140 | 261 | 334 | 550 | 302 | 302 | 152 | 162 | 230 | 0 | 852 | 169 | 224 | 448 | 0 | 852 | 0 |
| 7 | Ivanovic T (2023) [45] | 66 (68) | 31 (32) | 97 | 97 | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | 3 | 94 | 14 | 31 | 49 | 3 | 97 | 0 |
| 2164 | 1541 | ||||||||||||||||||||||
| Prognosis of PD-1+ TAMS | |||||||||||||||||||||||
| 1 | Kono Y (2020) [46] | 75 (73.5) | 27 (26.5) | 102 | 102 | 12 | NA | NA | NA | 48 | 54 | NA | NA | NA | NA | 85 | 17 | NA | NA | NA | 0 | 102 | 0 |
| 2 | Wang F (2018) [47] | 17 (65.4) | 9 (34.6) | 26 | 26 | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | 5 | 9 | 9 | 3 | NA | NA |
| 3 | Wei Y (2022) [48] | 8 (80) | 2 (20) | 10 | 10 | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | 1 | 9 | NA | NA | 6 | 2 | 10 | 0 |
| 138 | 138 | ||||||||||||||||||||||
| Correlation of TAMS and PD-L1 | ||||
|---|---|---|---|---|
| Author | Definition of PD-L1 | Definition of TAMS | Method of Analysis | |
| 1 | Ubukata Y (2020) [39] | No/low and high expression | CD163 level was determined | Immunohistochemistry |
| 2 | Ju X (2020) [40] | 0 (no staining), 1 (weak staining), 2 (moderate to intense staining). | CD68 in macrophage was scored as 1 (<1%), 2 (1–9%), 3, or 4 (>20%) | Immunohistochemistry |
| 3 | Junttila A (2020) [41] | PD-L1 CPS was ≤1% or >1%/Divided into two groups | Clever-1 was defined to be high when median ≥ 15 in macrophages | Immunohistochemistry |
| 4 | Huang Y.K (2019) [42] | Low or high expression | Macrophages were CD68 positive/M1-like TAM populations were identified based on the absence of CD163 and CD206/M2-like TAM populations were identified by the presence of CD163 and/or CD206 | Immunohistochemistry |
| 5 | Harada K (2017) [43] | PD-L1 positive if >1% of tumor cells expressed PD-L1 | The density of CD68- and CD163-positive cells was evaluated | Immunohistochemistry |
| 6 | Zhang H (2022) [44] | Positive or negative PD-L1 expression | For the CD68+ macrophages density, ≤66/HPF (at ×200 magnification) was defined as low and ≥67/HPF was defined as high/For the IL-10+ CD68+ cells density, ≤9/HPF was defined as low and ≥10/HPF was defined as high. | Immunohistochemistry |
| 7 | Ivanovic T (2023) [45] | PD-L1-positive tumor cells estimated as negative (0), low | The density of macrophage infiltration was valued as 0 (no infiltration), 1 (sparse), 2 (peripheral) and 3 (dense and intermingled with tumor cells) | Immunohistochemistry |
| Prognosis of PD-1+ TAMs | |||
|---|---|---|---|
| Author | Definition of PD1 + Macrophages | Method of Analysis | |
| 1 | Kono Y (2020) [46] | The frequency of PD-1+ macrophages was represented by the ratio of the number of PD-1+ CD68+ cells to that of CD68+ cells. | Immunohistochemistry |
| 2 | Wang F (2018) [47] | PD+ or PD- maxrophages | Immunohistochemistry |
| 3 | Wei Y (2022) [48] | High or low expression of PD-L1 in macrophages/Tumor macrophages expressed higher levels of CD68 and PPARg and lower levels of HLA-DR. | Mass cytometry by time of flight (CyTOF) combined with genomic bioinformatic analysis |
| Correlation of TAMS and PD-L1 | |||
|---|---|---|---|
| Author | Statistical Methods Were Used | Statistically Significant Cutoff | |
| 1 | Ubukata Y (2020) [39] | Student’s t-test and the χ2 test | p values < 0.05 |
| 2 | Ju X (2020) [40] | Kaplan-Meier methods, Log-Rank test and Student t-test | p values < 0.05 |
| 3 | Junttila A (2020) [41] | Kaplan-Meier method, Log-Rank, Chi-square test and Univariate and multivariate Cox proportional hazards regression models | p values < 0.05 |
| 4 | Huang Y.K (2019) [42] | Mann–Whitney U test (two-tailed), Spearman and Pearson correlation, Chi-square analysis and Kaplan–Meier analysis (Log-rank, Mantel-Cox test) | p values < 0.05 |
| 5 | Harada K (2017) [43] | Kruskal Wallis test, Pearson chi-square test, Fisher’s exact and Cox regression model test | p values < 0.05 |
| 6 | Zhang H (2022) [44] | Chi-squared test, Student t-test, Kaplan–Meier curves, Cox proportional hazards regression model and Spearman correlation analysis | p values < 0.05 |
| 7 | Ivanovic T (2023) [45] | t-test, Mann–Whitney U test, χ2 test, Pearson correlation coefficient, Kaplan–Meier curve and Log-rank test | p values < 0.05 |
| Prognosis of PD-1+ TAMs | |||
| 1 | Kono Y (2020) [46] | Paired t-test, Mann-Whitney U test, Kaplan-Meier method, Log-rank test and Cox’s proportional hazards model | p values < 0.05 |
| 2 | Wang F (2018) [47] | Bonferroni post test and Pearson’s correlation analysis | p values < 0.05 |
| 3 | Wei Y (2022) [48] | Paired sample t-test | p values < 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yerolatsite, M.; Torounidou, N.; Gogadis, A.; Kapoulitsa, F.; Ntellas, P.; Lampri, E.; Tolia, M.; Batistatou, A.; Katsanos, K.; Mauri, D. TAMs and PD-1 Networking in Gastric Cancer: A Review of the Literature. Cancers 2024, 16, 196. https://doi.org/10.3390/cancers16010196
Yerolatsite M, Torounidou N, Gogadis A, Kapoulitsa F, Ntellas P, Lampri E, Tolia M, Batistatou A, Katsanos K, Mauri D. TAMs and PD-1 Networking in Gastric Cancer: A Review of the Literature. Cancers. 2024; 16(1):196. https://doi.org/10.3390/cancers16010196
Chicago/Turabian StyleYerolatsite, Melina, Nanteznta Torounidou, Aristeidis Gogadis, Fani Kapoulitsa, Panagiotis Ntellas, Evangeli Lampri, Maria Tolia, Anna Batistatou, Konstantinos Katsanos, and Davide Mauri. 2024. "TAMs and PD-1 Networking in Gastric Cancer: A Review of the Literature" Cancers 16, no. 1: 196. https://doi.org/10.3390/cancers16010196
APA StyleYerolatsite, M., Torounidou, N., Gogadis, A., Kapoulitsa, F., Ntellas, P., Lampri, E., Tolia, M., Batistatou, A., Katsanos, K., & Mauri, D. (2024). TAMs and PD-1 Networking in Gastric Cancer: A Review of the Literature. Cancers, 16(1), 196. https://doi.org/10.3390/cancers16010196