Evaluation of Resistance to Fescue Toxicosis in Purebred Angus Cattle Utilizing Animal Performance and Cytokine Response

 ,
,
Abstract
1. Introduction
2. Results
2.1. Changes in Forage Characteristics and Ergot Alkaloid Concentration
2.2. Distribution of T-Snip Genotypes Across FTSM Selected Animals
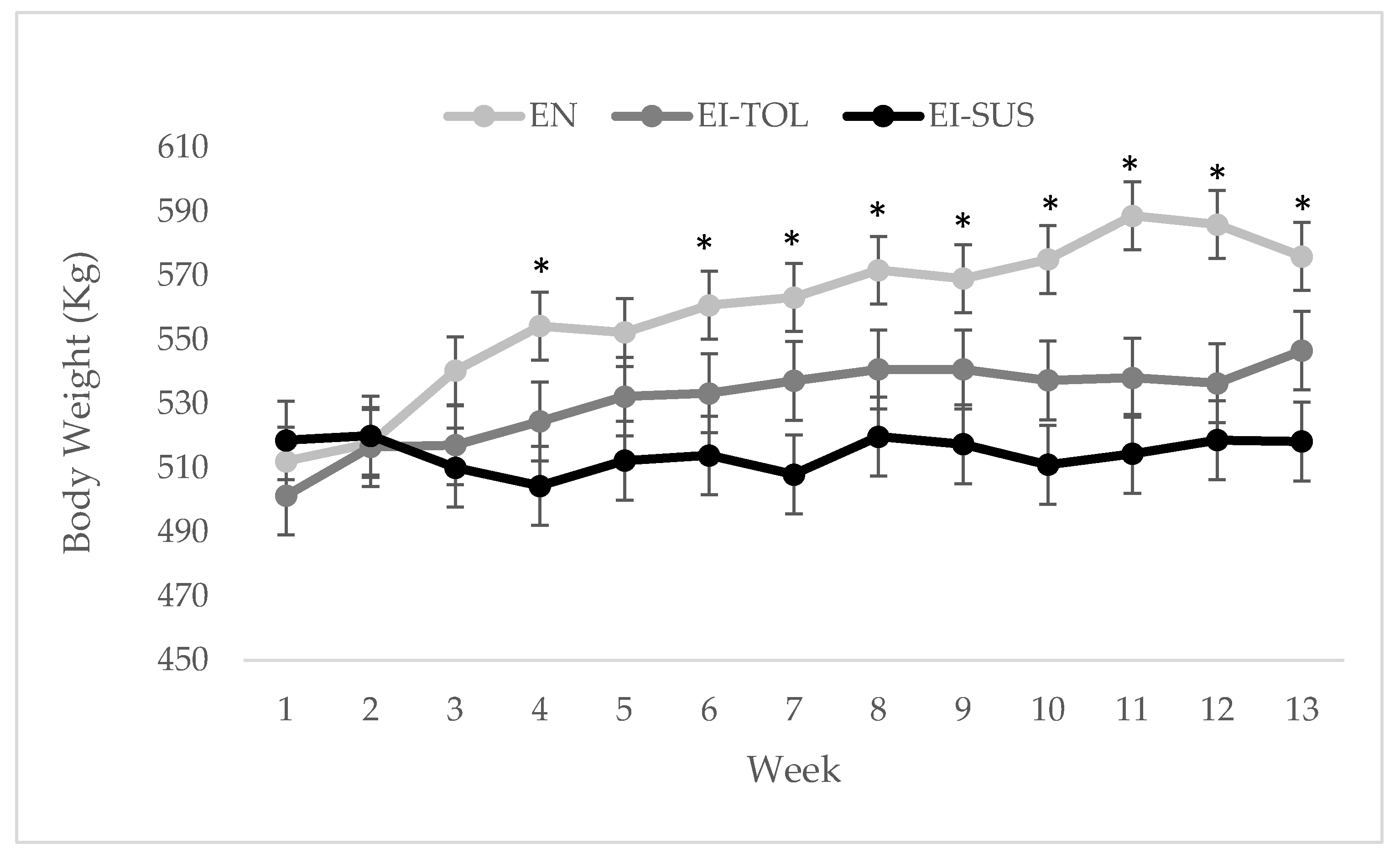
2.3. Phenotypic Variables of Animal Performance
2.4. Cytokine Response and Hormone Profiles in FTSM Selected Animals
3. Discussion
4. Conclusions
5. Materials and Methods
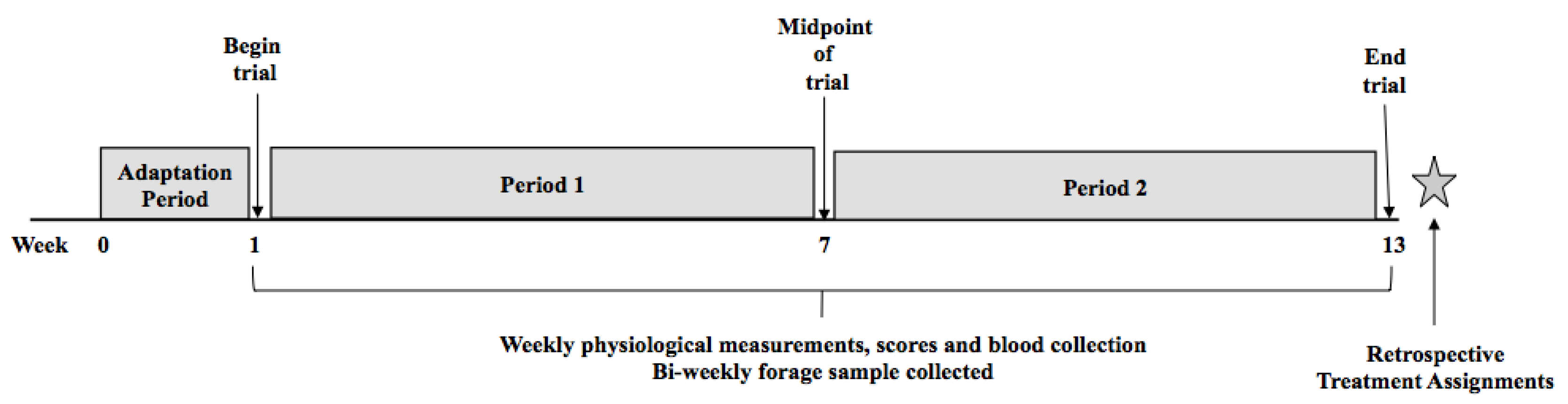
5.1. Animal Usage
Animal Management
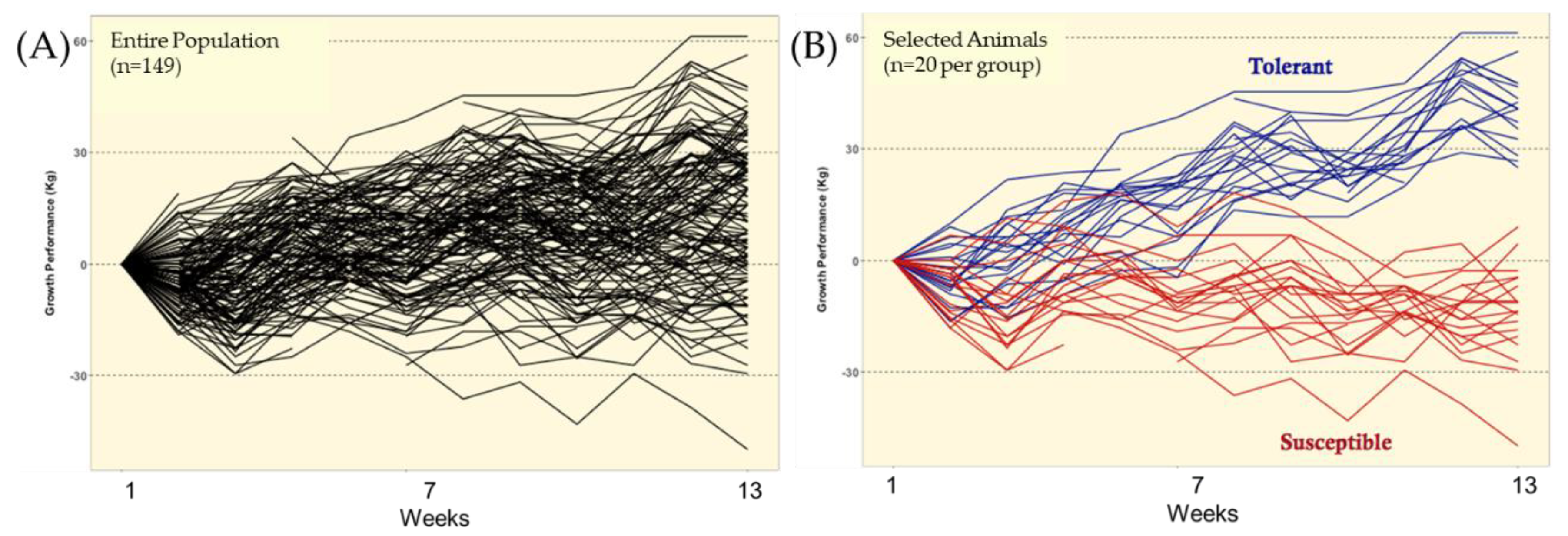
5.2. Fescue Toxicosis Selection Method (FTSM)
5.3. T-Snip Genotyping
5.4. Forage Management
5.5. Serum Assays and Analysis
5.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Strickland, J.R.; Looper, M.L.; Matthews, J.C.; Rosenkrans, C.F.; Flythe, M.D., Jr.; Brown, K.R. BOARD-INVITED REVIEW: St. Anthony’s fire in livestock: Causes, mechanisms, and potential solutions. J. Anim. Sci. 2011, 89, 1603–1626. [Google Scholar] [CrossRef]
- Miyazaki, S.; Ishizaki, I.; Ishizaka, M.; Kanbara, T.; Ishiguro-Takeda, Y. Lolitrem B residue in fat tissues of cattle consuming endophyte-infected perennial ryegrass straw. J. Vet. Diagn. Investig. 2004, 16, 340–342. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, C.W.; Ahn, G.C.; Park, E.K.; Kim, C.M.; Park, K.K. Ergovaline levels in tall fescue and its effect on performance of lactating cows. Anim. Feed. Sci. Technol. 2007, 136, 330–337. [Google Scholar] [CrossRef]
- McCulley, R.L.; Bush, L.P.; Carlisle, A.E.; Ji, H.; Nelson, J.A. Warming reduces tall fescue abundance but stimulates toxic alkaloid concentrations in transition zone pastures of the U.S. Front. Chem. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Galliou, J.M.; Khanal, P.; Mayberry, K.J.; Poore, M.H.; Poole, D.H.; Serão, N.V.L. Evaluation of a commercial genetic test for fescue toxicosis in pregnant Angus beef cattle. Transl. Anim. Sci. 2020, txaa181. [Google Scholar] [CrossRef]
- Grusie, T.; Cowwan, V.; Singh, J.; McKinnon, J.; Blakley, B. Proportions of predominant Ergot alkaloids (Claviceps purpurea) detected in Western Canadian grains from 2014–2016. World Mycotoxin J. 2018, 11, 259–264. [Google Scholar] [CrossRef]
- Poole, D.H.; Lyons, S.E.; Poole, R.K.; Poore, M.H. Ergot alkaloids induce vasoconstriction of bovine uterine and ovarian blood vessels. J. Anim. Sci. 2018, 96, 4812–4822. [Google Scholar] [CrossRef]
- Poole, R.K.; Devine, T.L.; Mayberry, K.J.; Eisemann, J.H.; Poore, M.H.; Long, N.M.; Poole, D.H. Impact of slick hair trait on physiological and reproductive performance in beef heifers consuming ergot alkaloids from endophyte-infected tall fescue. J. Anim. Sci. 2019, 97, 1456–1467. [Google Scholar] [CrossRef]
- Rottinghaus, G.E.; Garner, G.B.; Cornell, C.N.; Ellis, J.L. HPLC method for quantitating ergovaline in endophyte-infested tall fescue: Seasonal variation of ergovaline levels in stems with leaf sheaths, leaf blades, and seed heads. J. Agric. Food Chem. 1991, 39, 112–115. [Google Scholar] [CrossRef]
- Browning, R., Jr. Effects of endophyte-infected tall fescue on indicators of thermal status and growth in Hereford and Senepol steers. J. Anim. Sci. 2004, 82, 634–643. [Google Scholar] [CrossRef][Green Version]
- Burke, J.M.; Coleman, S.W.; Chase, C.C., Jr.; Riley, D.G.; Looper, M.L.; Brown, M.A. Interaction of breed type and endophyte-infected tall fescue on milk production and quality in beef cattle. J. Anim. Sci. 2010, 88, 2802–2811. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cole, N.A.; Brown, M.A.; Phillips, W.A. Genetic x environment interactions on blood constituents of Angus, Brahman, and reciprocal-cross cows and calves grazing common bermudagrass of endophyte-infected tall fescue. J. Anim. Sci. 2001, 79, 1151–1161. [Google Scholar] [CrossRef] [PubMed]
- Gray, K.; Smith, T.; Maltecca, C.; Overton, P.; Parish, J.A.; Cassady, J.P. Differences in hair coat shedding, and effects on calf weaning weight and BCS among Angus dams. Livest. Sci. 2011, 140, 68–71. [Google Scholar] [CrossRef]
- Campbell, B.T.; Kojima, C.J.; Cooper, T.A.; Bastin, B.C.; Wojakiewicz, L.; Kallenbach, R.L.; Schrick, F.N.; Waller, J.C. A Single Nucleotide Polymorphism in the Dopamine Receptor D2 Gene May Be Informative for Resistance to Fescue Toxicosis in Angus-Based Cattle. Anim. Biotechnol. 2014, 25, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hill, N.S.; Thompson, F.N.; Stuedemann, J.A.; Dawe, D.L.; Hiatt, E.E., 3rd. Urinary Alkaloid Excretion as a Diagnostic Tool for Fescue Toxicosis in Cattle. J. Vet. Diagn. Investig. 2000, 12, 210–217. [Google Scholar] [CrossRef]
- Drewnoski, M.E.; Poore, M.H.; Oliphant, E.J.; Marshall, B.; Green, J.T., Jr. Agronomic Performance of Stockpiled Tall Fescue Varies with Endophyte Infection Status. Online. Forage Grazinglands 2007, 5, 1–13. [Google Scholar] [CrossRef]
- Poore, M.H.; Drewnoski, M. Review: Utilization of stockpiled tall fescue in grazing systems for beef cattle. Prof. Anim. Sci. 2010, 26, 142–149. [Google Scholar] [CrossRef]
- Thompson, R.W.; Fribourg, H.A.; Waller, J.C.; Sanders, W.L.; Reynolds, J.H.; Phillips, J.M.; Schmidt, S.P.; Crawford, R.J.; Allen, V.G.; Faulkner, D.B.; et al. Combined analysis of tall fescue steer grazing studies in the Eastern United States. J. Anim. Sci. 1993, 71, 1940–1946. [Google Scholar] [CrossRef]
- Brown, M.A.; Tharel, L.M.; Brown, A.H., Jr.; Miesner, J.R.; Jackson, W.G. Reproductive Performance of Angus and Brahman Cows Grazing Common Bermudagrass or Endophyte-lnfected Tall Fescue. Prof. Anim. Sci. 1992, 8, 58–65. [Google Scholar] [CrossRef]
- Burke, J.; Bishop, C.V.; Stormshak, F. Reproductive characteristics of endophyte-infected or novel tall fescue fed ewes. Livest. Sci. 2006, 104, 103–111. [Google Scholar] [CrossRef]
- Parish, J.A.; McCann, M.A.; Watson, R.H.; Paiva, N.N.; Hoveland, C.S.; Parks, A.H.; Upchurch, B.L.; Hill, N.S.; Bouton, J.H. Use of nonergot alkaloid-producing endophytes for alleviating tall fescue toxicosis in stocker cattle. J. Anim. Sci. 2003, 81, 2856–2868. [Google Scholar] [CrossRef] [PubMed]
- Osborn, T.G. Effect of Consuming Fungus-Infected and Fungus-Free Tall Fescue and Ergotamine Tartrate on Certain Physiological Variables of Cattle in Environmentally-Controlled Conditions. Ph.D. Thesis, Auburn University, Auburn, AL, USA, 1988. [Google Scholar]
- Browning, R., Jr. Physiological responses of Brahman and Hereford steers to an acute ergotamine challenge. J. Anim. Sci. 2000, 78, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Cole, N.A.; Stuedemann, J.A.; Thompson, F.N. Influence of both endophyte infestation in fescue pastures and calf genotype on subsequent feedlot performance of steers. Prof. Anim. Sci. 2001, 17, 174–182. [Google Scholar] [CrossRef]
- Fanning, M.D.; Spitzer, J.C.; Cross, D.L.; Thompson, F.N. A preliminary study of Growth, serum prolactin and reproductive performance of beef heifers grazing Acremonium coenophialum-infected tall fescue. Theriogenology 1992, 38, 375–384. [Google Scholar] [CrossRef]
- Smith, V.G.; Hacker, R.R.; Brown, R.G. Effect of Alterations in Ambient Temperature on Serum Prolactin Concentration in steers. J. Anim. Sci. 1977, 44, 645–649. [Google Scholar] [CrossRef] [PubMed]
- Igono, M.O.; Johnson, H.D.; Steevens, B.J.; Hainen, W.A.; Shanklin, M.D. Effect of season on milk temperature, milk growth hormone, prolactin, and somatic cell counts of lactating cattle. Int. J. Biometeorol. 1988, 32, 194–200. [Google Scholar] [CrossRef]
- Schams, D.; Reinhardt, V. Influence of the Season on Plasma Prolactin Level in Cattle from Birth to Maturity. Horm. Res. 1974, 5, 217–226. [Google Scholar] [CrossRef]
- Wettemann, R.P.; Tucker, H.A. Relationship of Ambient Temperature to Serum Prolactin in Heifers. Proc. Soc. Exp. Biol. Med. 1974, 146, 908–911. [Google Scholar] [CrossRef]
- Foitzik, K.; Krause, K.; Nixon, A.J.; Ford, C.A.; Ohnemus, U.; Pearson, A.J.; Paus, R. Prolactin and its receptor are expressed in murine hair follicle epithelium show hair cycle-dependent expression, and induce catagen. Am. J. Pathol. 2003, 162, 1611–1621. [Google Scholar] [CrossRef]
- Nixon, A.J.; Ford, C.A.; Wildermoth, J.E.; Craven, A.J.; Ashby, M.G.; Pearson, A.J. Regulation of prolactin receptor expression in ovine skin in relation to circulating prolactin and wool follicle growth status. J. Endocrinol. 2002, 172, 605–614. [Google Scholar] [CrossRef]
- Aiken, G.E.; Klotz, J.; Looper, M.; Tabler, S.; Schrick, F. Disrupted hair follicle activity in cattle grazing endophyte-infected tall fescue in the summer insulates core body temperatures. Prof. Anim. Sci. 2011, 27, 336–343. [Google Scholar] [CrossRef]
- Koester, L.R.; Poole, D.H.; Serão, N.V.L.; Schmitz-Esser, S. Beef cattle that respond differently to fescue toxicosis have distinct gastrointestinal tract microbiota. PLoS ONE 2020, 15, e0229192. [Google Scholar] [CrossRef] [PubMed]
- Serão, N.V.L.; Kemp, R.A.; Mote, B.E.; Willson, P.; Harding, J.C.S.; Bishop, S.C.; Plastow, G.S.; Dekkers, J.C.M. Genetic and genomic basis of antibody response to porcine reproductive and respiratory syndrome (PRRS) in gilts and sows. Genet. Sel. Evol. 2016, 48, 51. [Google Scholar] [CrossRef] [PubMed]
- Poole, R.K.; Brown, A.R.; Poore, M.H.; Pickworth, C.L.; Poole, D.H. Effects of endophyte-infected tall fescue seed and protein supplementation on stocker steers: II. Adaptive and innate immune function. J. Anim. Sci. 2019, 97, 4160–4170. [Google Scholar] [CrossRef] [PubMed]
- Freeman, M.E.; Kanyicska, B.; Lerant, A.; Nagy, G. Prolactin: Structure, Function, and Regulation of Secretion. Physiol. Rev. 2000, 80, 1523–1631. [Google Scholar] [CrossRef]
- Lamberts, S.W.; MacLeod, R.M. Regulation of prolactin secretion at the level of the lactotroph. Physiol. Rev. 1990, 70, 279–318. [Google Scholar] [CrossRef]
- Sibley, D.R.; Creese, I. Interactions of ergot alkaloids with anterior pituitary D-2 dopamine receptors. Mol. Pharmacol. 1983, 23, 585–593. [Google Scholar]
- Porter, J.K.; Thompson, J.F.N. Effects of fescue toxicosis on reproduction in livestock. J. Anim. Sci. 1992, 70, 1594–1603. [Google Scholar] [CrossRef]
- Janeway, C.A., Jr.; Travers, P.; Walport, M.; Shlomchik, M. Immuno-Biology: The Immune System in Health and Disease, 5th ed.; Garland Publishing: New York, NY, USA, 2001; Print. [Google Scholar]
- Aiken, G.E.; Flythe, M.D. Vasoconstrictive responses by the carotid and auricular arteries in goats to ergot alkaloid exposure. Front. Chem. 2014, 2. [Google Scholar] [CrossRef]
- Jyotiranjan, T.; Mohapatra, S.; Mishra, C.; Dalai, N.; Kundu, A.K. Heat tolerance in goat—A genetic update. Pharma Innov. J. 2017, 6, 237–245. [Google Scholar]
- Iqbal, N.; Liu, X.; Yang, T.; Huang, Z.; Hanif, Q.; Asif, M.; Khan, Q.M.; Mansoor, S. Genomic variants identified from whole-genome resequencing of indicine cattle breeds from Pakistan. PLoS ONE 2019, 14, e0215065. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.; King, S.S.; Griswold, K.E.; Cazac, D.; Cross, D.L. Domperidone can ameliorate deleterious reproductive effects and reduced weight gain associated with fescue toxicosis in heifers. J. Anim. Sci. 2003, 81, 2568–2574. [Google Scholar] [CrossRef] [PubMed]
- Aiken, G.E.; Kirch, B.H.; Strickland, J.R.; Bush, L.P.; Looper, M.L.; Schrick, F.N. Hemodynamic responses of the caudal artery to toxic tall fescue in beef heifers. J. Anim. Sci. 2007, 85, 2337–2345. [Google Scholar] [CrossRef] [PubMed]
- Berghof, T.; Poppe, M.; Mulder, H.A. Opportunities to Improve Resilience in Animal Breeding Programs. Front. Genet. 2019, 9, 692. [Google Scholar] [CrossRef] [PubMed]
- Richards, M.W.; Spitzer, J.C.; Warner, M.B. Effect of Varying Levels of Postpartum Nutrition and Body Condition at Calving on Subsequent Reproductive Performance in Beef Cattle. J. Anim. Sci. 1986, 62, 300–306. [Google Scholar] [CrossRef]
- Olson, T.A.; Lucena, C.; Chase, C.C.; Hammond, A.C. Evidence of a major gene influencing hair length and heat tolerance in Bos taurus cattle. J. Anim. Sci. 2003, 81, 80–90. [Google Scholar] [CrossRef]
- Lyons, S.E.; Shaeffer, A.D.; Drewnoski, M.E.; Poore, M.H.; Poole, D.H. Effect of protein supplementation and forage allowance on the growth and reproduction of beef heifers grazing stockpiled tall fescue. J. Anim. Sci. 2016, 94, 1677–1688. [Google Scholar] [CrossRef]
- SAS. SAS System for Mixed Models; SAS Inst. Inc.: Cary, NC, USA, 1996. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2017. Available online: https://www.R-project.org/ (accessed on 9 August 2019).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forage Percentage | BBCFL 1 | UPRS 2 | Combined Locations | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| P-1 3 | P-2 4 | ES 5 | P-1 3 | P-2 4 | ES 5 | P-1 3 | P-2 4 | ES 5 | |||
| Percent fescue (%) | 72.5 | 56.0 | 64.3 | 72.9 | 63.3 | 68.1 | 72.7 | 59.7 | 66.2 | ||
| Tiller infection 6 (%) | 84.2 | 77.5 | 80.8 | 82.5 | 90.0 | 86.3 | 83.3 | 83.8 | 83.5 | ||
| Ergot alkaloid analysis 7 | |||||||||||
| Ergosine (μg/kg) | – 8 | – | – | – | 333.3 ± 322 | 166.7 ± 230 | – | 166.7 ± 230 | 83.3 ± 163 | ||
| Ergotamine (μg/kg) | – | – | – | – | 175.0 ± 202 | 87.5 ± 44 | – | 87.5 ± 142 | 43.8 ± 101 | ||
| Ergocornine (μg/kg) | – | – | – | – | 53.3 ± 61 | 26.7 ± 43 | – | 26.7 ± 43 | 13.3 ± 30 | ||
| Ergocryptine (μg/kg) | – | – | – | – | 150.0 ± 93 | 75.0 ± 66 | – | 75.0 ± 66 | 37.5 ± 47 | ||
| Ergocristine (μg/kg) | – | – | – | – | 48.3 ± 51 | 24.2 ± 36 | – | 24.2 ± 36 | 12.1 ± 25 | ||
| Ergovaline (μg/kg) | 263.3 ± 52 | 106.7 ± 41 | 185.0 ± 38 | 436.7 ± 83 | 196.7 ± 102 | 316.7 ± 64 | 350 ± 58 | 151.7 ± 55 | 250.8 ± 42 | ||
| Total (μg/kg) | 263.3 ± 52 | 106.7 ± 41 | 185.0 ± 38 | 436.7 ± 83 | 956.7 ± 43 | 696.7 ± 45 | 350.0 ± 58 | 531.7 ± 25 | 440.8 ± 37 | ||
| FTSM 1 | T-Snip Genotypes | |||
|---|---|---|---|---|
| 1 | 2 | 3 | 4 | |
| TOL | 0.150 a (0.08) | 0.100 a (0.07) | 0.600 a (0.11) | 0.15 a (0.08) |
| SUS | 0.263 a (0.10) | 0.368 b (0.11) | 0.316 b (0.11) | 0.05 a (0.05) |
| Item | Treatment 1,2 | SEM | p Value | ||
|---|---|---|---|---|---|
| EN | EI-TOL | EI-SUS | |||
| Initial BW, kg | 512.1 | 501.4 | 518.6 | 10.6 | 0.5082 |
| Final BW, kg | 576.0 a,* | 546.6 a,b,* | 518.3 b | 12.3 | 0.0004 |
| ADG, kg/d | 0.703 a | 0.497 b | −0.003 c | 0.05 | <0.0001 |
| BCS 3 | 6.09 a | 5.50 b | 5.59 b | 0.06 | <0.0001 |
| HCS 4 | 1.89 a,* | 1.98 a,* | 2.34 b | 0.04 | <0.0001 |
| HSS 5 | 1.99 a | 2.36 b | 2.64 c | 0.04 | <0.0001 |
| Rectal temp., °C | 38.6 a | 38.7 b | 38.9 c | 0.02 | <0.0001 |
| Time | P-1 1 | P-2 2 | ES 3 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatment 4,5 | EI-TOL | EI-SUS | SEM | p-Value | EI-TOL | EI-SUS | SEM | p Value | EI-TOL | EI-SUS | SEM | p-Value |
| ADG, kg/d | 1.8 a | −0.55 b | 0.13 | <0.0001 | 0.5 | 0.5 | 0.2 | 0.8726 | 1.2 a | −0.025 b | 0.1 | <0.0001 |
| ∆BCS 6 | 0.488 a | 0.013 b | 0.09 | 0.0008 | 0.613 * | 0.363 * | 0.09 | 0.0625 | 1.05 a | 0.375 b | 0.10 | <0.0001 |
| ∆HCS 7 | −2.013 a | −0.913 b | 0.174 | <0.0001 | −0.300 a | −1.150 b | 0.207 | 0.0062 | −2.313 | −2.063 | 0.226 | 0.4398 |
| ∆HSS 8 | −1.925 a | −0.613 b | 0.198 | <0.0001 | −0.763 a | −1.838 b | 0.202 | 0.0006 | −2.688 | −2.450 | 0.248 | 0.5027 |
| ∆RT 9, °C | −0.250 | 0.105 | 0.176 | 0.1630 | 0.505 | 0.230 | 0.126 | 0.1299 | 0.250 | 0.325 | 0.179 | 0.7680 |
| Treatment 1,2 | EI-TOL | EI-SUS | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Location | BBCFL 3 | UPRS 4 | BBCFL 3 | UPRS 4 | p-Values | |||||||||||
| Week | 1 | 7 | 13 | 1 | 7 | 13 | 1 | 7 | 13 | 1 | 7 | 13 | SEM | TRT | LOC | TIME |
| Growth factors 5 | ||||||||||||||||
| FGF1 | 196.5 * | 110.3 * | 77.3 * | 233.8 * | 145.2 * | 120.0 * | 98.0 * | 69.3 * | 52.3 * | 203.8 * | 163.0 * | 99.8 * | 60.4 | 0.3381 | 0.0793 | 0.0716 |
| FGF2 | 9.4 | 28.1 | 46.8 | 27.9 | 16.6 | 16.1 | 34.9 | 26.6 | 19.9 | 25.0 | 1937 | 7.3 | 14.8 | 0.8324 | 0.3327 | 0.9849 |
| IGF1 | 6.8 | 21.3 | 10.4 | 21.2 | 4.3 | 6.5 | 18.7 | 14.1 | 3.9 | 10.2 | 8.8 | 9.3 | 4.2 | 0.7330 | 0.3602 | 0.1441 |
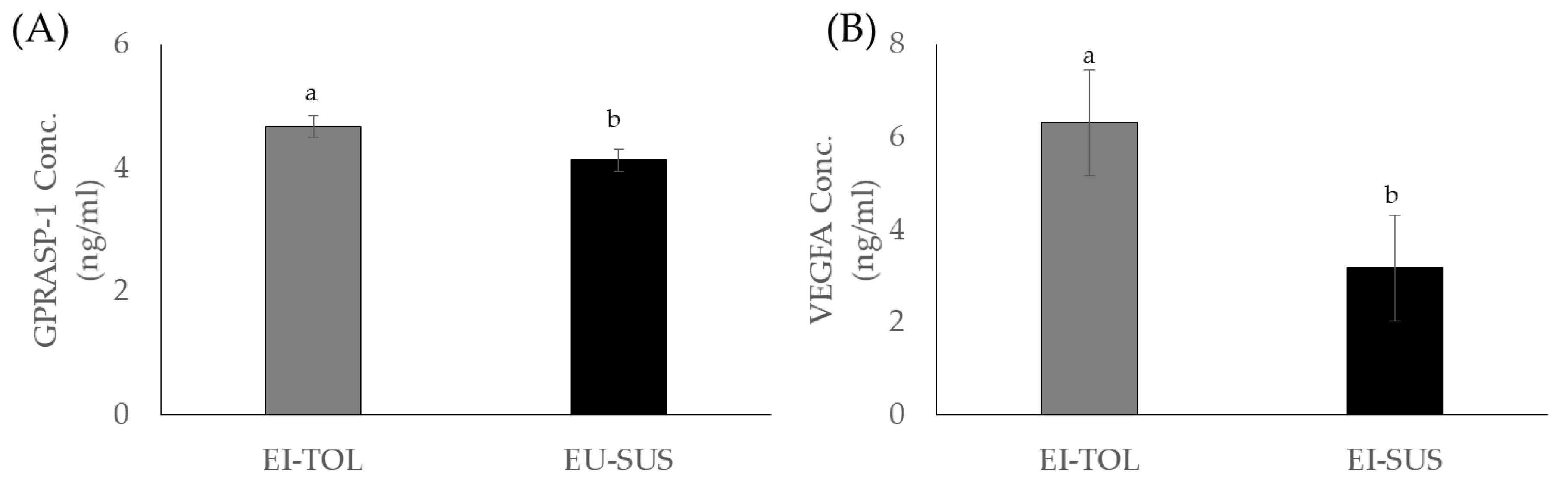
| VEGFA | 572.64 | 429.9 | 441.0 | 1117.2 | 512.4 | 711.2 | 208.7 | 152.9 | 220.1 | 253.64 | 769.8 | 297.2 | 114.2 | 0.0444 | 0.1070 | 0.8319 |
| Chemokines 5 | ||||||||||||||||
| CCL2 | 856.4 | 2817.6 | 269.7 | 2012.2 | 1946.2 | 1979.8 | 269.2 | 2124.8 | 277.7 | 1022.8 | 1622.4 | 1374.8 | 898.7 | 0.3210 | 0.2984 | 0.1452 |
| CCL4 | 8.7 | 11.9 | 141.3 | 16.8 a | 616.7 b | 754.9 b | 9.8 | 10.8 | 191.8 | 7.6 a | 171.1 a,b | 404.4 b | 192.7 | 0.2831 | 0.0245 | 0.0428 |
| CXCL9 | 35.7 | 41.6 | 73.2 | 41.8 | 250.9 | 220.1 | 98.2 | 42.2 | 62.8 | 75.9 | 56.3 | 150.1 | 70.2 | 0.4644 | 0.0708 | 0.4237 |
| CXCL10 | 34.7 | 44.3 | 68.1 | 45.1 | 91.0 | 104.6 | 42.2 | 34.9 | 57.5 | 80.8 | 54.2 | 67.1 | 27.7 | 0.5909 | 0.0929 | 0.4521 |
| Cytokine receptors antagonists 5 | ||||||||||||||||
| IL36RN | 69.2 | 94.2 | 167.4 | 109.1 | 562.9 | 512.9 | 67.8 | 73.8 | 110.0 | 205.8 | 252.2 | 357.6 | 136.7 | 0.3590 | 0.0044 | 0.1922 |
| GPRASP-1 | 4.1 a | 6.1 b | 4.2 a | 4.2 | 4.7 | 4.7 | 3.8 a | 5.2 b | 3.8 a | 4.2 | 4.0 | 3.8 | 0.4 | 0.0343 | 0.3268 | 0.0033 |
| NCAM1 | 33.8 | 33.7 | 30.7 | 33.1 | 32.3 | 42.4 | 29.9 | 31.9 | 26.9 | 30.6 | 35.0 | 39.4 | 3.8 | 0.3428 | 0.0496 | 0.5422 |
| Treatment 1,2 | EI-TOL | EI-SUS | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Location | BBCFL 3 | UPRS 4 | BBCFL 3 | UPRS 4 | p-Values | ||||||||||||
| Week | 1 | 7 | 13 | 1 | 7 | 13 | 1 | 7 | 13 | 1 | 7 | 13 | SEM | TRT | LOC | TIME | |
| Anti-inflammatory 5 | |||||||||||||||||
| IL4 | 89.7 | 256.8 | 30.1 | 286.1 | 215.8 | 407.7 | 48.9 | 47.8 | 26.4 | 198.2 | 110.1 | 95.2 | 133.9 | 0.1141 | 0.0909 | 0.9801 | |
| IL13 | 6.7 | 7.4 | 5.0 | 13.1 | 9.5 | 10.1 | 9.5 | 10.7 | 2.1 | 26.1 | 10.4 | 6.0 | 5.1 | 0.4710 | 0.0672 | 0.1021 | |
| Pro-inflammatory 5 | |||||||||||||||||
| IFNAR1 | 20.0 a | 44.8 a | 460.2 b | 71.5 a | 304.5 b | 518.2 b | 44.4a | 52.4a | 362.7 b | 29.2 a | 259.9 b | 306.5 b | 49.5 | 0.3921 | 0.2361 | 0.0001 | |
| IFNG | 1.7 | 1.8 | 1.8 | 2.5 | 3.2 | 4.5 | 3.9 | 2.0 | 1.2 | 4.3 | 3.5 | 2.9 | 1.5 | 0.6638 | 0.1083 | 0.8552 | |
| IL1A | 72.0 | 53.3 | 209.0 | 99.2 | 273.7 | 380.6 | 146.0 | 65.6 | 116.6 | 162.4 | 159.3 | 266.7 | 85.1 | 0.5768 | 0.0287 | 0.1060 | |
| IL15 | 13.1 | 41.5 | 4.9 | 25.8 | 22.3 | 32.6 | 9.7 | 14.7 | 6.1 | 12.6 | 30.1 | 11.5 | 12.7 | 0.1847 | 0.2822 | 0.2296 | |
| IL2 | 107.6 | 218.8 | 255.0 | 100.4 | 191.0 | 87.8 | 216.0 | 110.1 | 143.8 | 30.6 | 43.1 | 60.0 | 93.8 | 0.2878 | 0.1113 | 0.9128 | |
| IL21 | 60.9 | 54.0 | 92.4 | 89.2 | 177.5 | 178.2 | 118.1 | 68.3 | 48.7 | 167.7 | 66.4 | 132.6 | 58.6 | 0.8095 | 0.0795 | 0.8667 | |
| TNF | 43.0 | 52.0 | 77.3 | 86.3 | 175.5 | 263.5 | 113.9 | 64.2 | 53.1 | 176.4 | 64.2 | 130.4 | 70.5 | 0.7046 | 0.0523 | 0.7093 | |
| Reproductive hormones 5 | |||||||||||||||||
| Prolactin | 55.0 a | 102.4 b | 74.5 a,b | 51.0 a | 166.0 b | 140.7 a,b | 89.5 a | 167.5 b | 105.7 a,b | 60.6 a | 217.6 b | 189.4 a,b | 38.8 | 0.0787 | 0.0918 | 0.0019 | |
| Progesterone | 5.0a | 2.3 b | 2.9 b | 2.9 | 1.8 | 3.0 | 3.8 a | 2.5 b | 3.1a,b | 2.4 | 2.3 | 2.4 | 0.5 | 0.0895 | 0.0018 | 0.0003 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poole, D.H.; Mayberry, K.J.; Newsome, M.; Poole, R.K.; Galliou, J.M.; Khanal, P.; Poore, M.H.; Serão, N.V.L. Evaluation of Resistance to Fescue Toxicosis in Purebred Angus Cattle Utilizing Animal Performance and Cytokine Response. Toxins 2020, 12, 796. https://doi.org/10.3390/toxins12120796
Poole DH, Mayberry KJ, Newsome M, Poole RK, Galliou JM, Khanal P, Poore MH, Serão NVL. Evaluation of Resistance to Fescue Toxicosis in Purebred Angus Cattle Utilizing Animal Performance and Cytokine Response. Toxins. 2020; 12(12):796. https://doi.org/10.3390/toxins12120796
Chicago/Turabian StylePoole, Daniel H., Kyle J. Mayberry, McKayla Newsome, Rebecca K. Poole, Justine M Galliou, Piush Khanal, Matthew H. Poore, and Nick V. L. Serão. 2020. "Evaluation of Resistance to Fescue Toxicosis in Purebred Angus Cattle Utilizing Animal Performance and Cytokine Response" Toxins 12, no. 12: 796. https://doi.org/10.3390/toxins12120796
APA StylePoole, D. H., Mayberry, K. J., Newsome, M., Poole, R. K., Galliou, J. M., Khanal, P., Poore, M. H., & Serão, N. V. L. (2020). Evaluation of Resistance to Fescue Toxicosis in Purebred Angus Cattle Utilizing Animal Performance and Cytokine Response. Toxins, 12(12), 796. https://doi.org/10.3390/toxins12120796