Candida utilis ATCC 9950 Cell Walls and β(1,3)/(1,6)-Glucan Preparations Produced Using Agro-Waste as a Mycotoxins Trap

 , ,
, , 
Abstract
1. Introduction
2. Results
2.1. Chemical Composition of Studied Cell Walls and β-Glucan Preparations
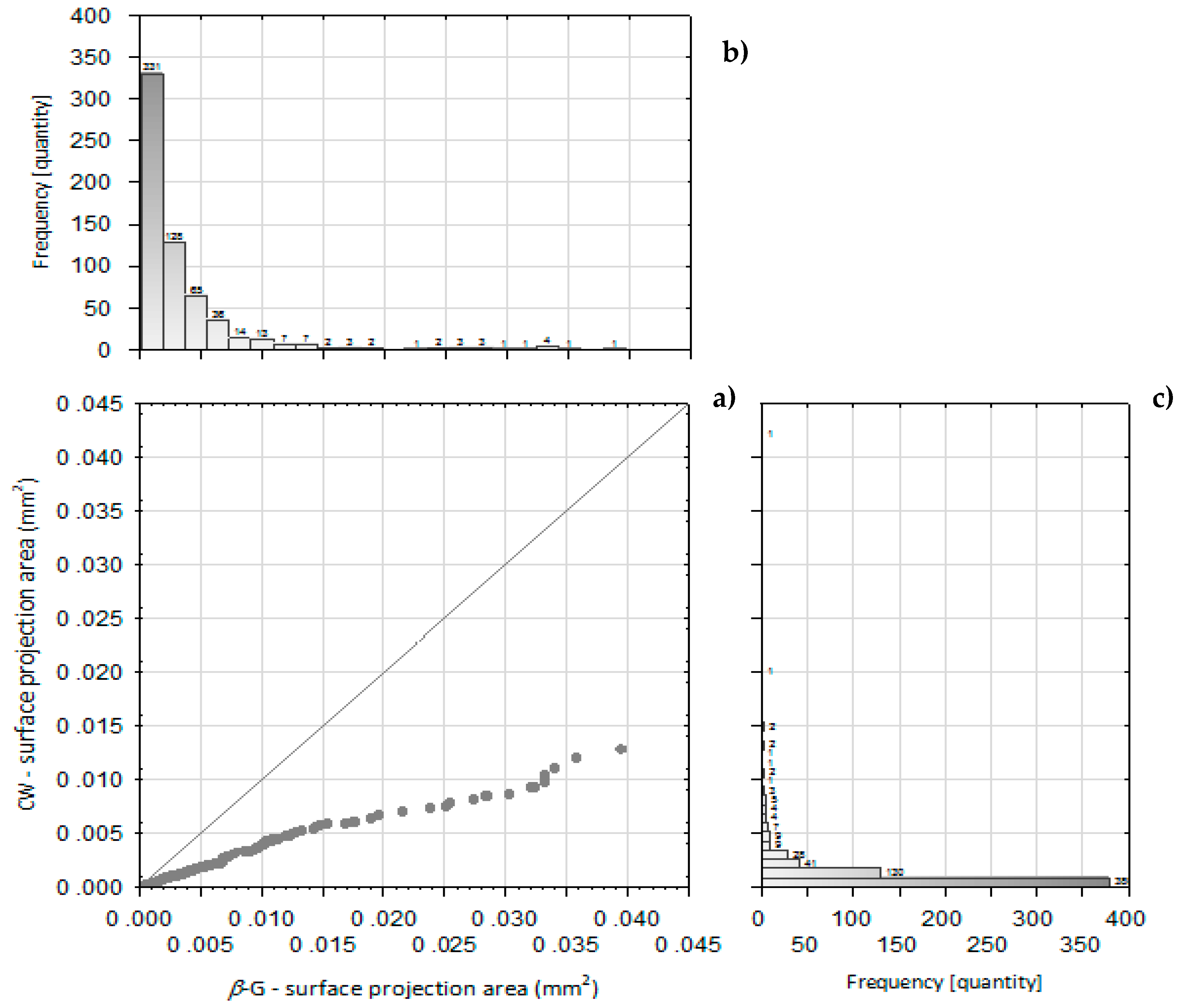
2.2. Microstructure and Particle Size of Isolated Powder Preparations
2.3. Results of Adsorption Tests
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Reagents and Analytical Standards
5.2. Production of Cell Walls and β(1,3)/(1,6)-Glucan Preparations
5.3. Determination of Particle Microstructure and Size of Powdered Cell Walls and β(1,3)/(1,6)-Glucan Preparations
5.4. Conditions of pH-Dependent Mycotoxin Adsorption by Studied Preparations
5.5. Mycotoxins Analysis with LC-MS
5.6. Estimation of Adsorption Efficiency, Adsorption Capacity and Distribution Coefficient
5.7. Data Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pinotti, L.; Ottoboni, M.; Giromini, C.; Dell’Orto, V.; Cheli, F.; Battilani, P. Mycotoxin Contamination in the EU Feed Supply Chain: A Focus on Cereal Byproducts. Toxins 2016, 8, 45. [Google Scholar] [CrossRef]
- Schatzmayr, G.; Streit, E. Global occurrence of mycotoxins in the food and feed chain: facts and figures. World Mycotoxin J. 2013, 6, 213–222. [Google Scholar] [CrossRef]
- Peng, Z.; Chen, L.; Xiao, J.; Zhou, X.; Nüssler, A.K.; Liu, L.; Liu, J.; Yang, W. Review of mechanisms of deoxynivalenol-induced anorexia: The role of gut microbiota. J. Appl. Toxicol. 2017, 8, 940–1029. [Google Scholar] [CrossRef]
- Zeidan, R.; Ul-Hassan, Z.; Al-Thani, R.; Balmas, V.; Jaoua, S. Application of Low-Fermenting Yeast Lachancea thermotolerans for the Control of Toxigenic Fungi Aspergillus parasiticus, Penicillium verrucosum and Fusarium graminearum and Their Mycotoxins. Toxins 2018, 10, 242. [Google Scholar] [CrossRef]
- Vila-Donat, P.; Marín, S.; Sanchis, V.; Ramos, A.J. A review of the mycotoxin adsorbing agents, with an emphasis on their multi-binding capacity, for animal feed decontamination. Chem. Toxicol. 2018, 114, 246–259. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, M.; Negi, B.; Kaushik, N.; Adhikari, A.; Al-Khedhairy, A.A.; Kaushik, N.K.; Choi, E.H. T-2 mycotoxin: toxicological effects and decontamination strategies. Oncotarget 2017, 8, 33933–33952. [Google Scholar] [CrossRef]
- Sharma, R.P. Immunotoxicity of mycotoxins. J. Dairy Sci. 1993, 76, 892–897. [Google Scholar] [CrossRef]
- Awad, W.A.; Ghareeb, K.; Böhm, J.; Zentek, J. Decontamination and detoxification strategies for the Fusarium mycotoxin deoxynivalenol in animal feed and the effectiveness of microbial biodegradation. Addit. Contam. A 2010, 27, 510–520. [Google Scholar] [CrossRef] [PubMed]
- Kabak, B.; Dobson, A.D.W.; Var, I. Strategies to prevent mycotoxin montamination of food and animal feed: a review. Crit. Rev. Food Sci. Nutr. 2006, 46, 593–619. [Google Scholar] [CrossRef]
- European Commission. Directive 2002/32/EC of the European Parliament and of the Council of 7 May 2002 on undesirable substances in animal feed. 2002. Available online: http://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:02002L0032-20131227 (accessed on 15 November 2018).
- European Commission. Commission Directive 2003/100/EC of 31 October 2003 amending annex I to directive 2002/32/EC of the European Parliament and of the Council on undesirable substances in animal feed. Off. J. Eur. Union 2003, 285, 33–37. [Google Scholar]
- European Commission. Commission Recommendation of 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding. Off. J. Eur. Union 2006, 229, 7–9. [Google Scholar]
- European Commission. Commission Recommendation of 27 March 2013 on the presence of T-2 and HT-2 toxin in cereals and cereal products. Off. J. Eur. Union 2013, 91, 12–15. [Google Scholar]
- De Mil, T.; Devreese, M.; De Baere, S.; Van Ranst, E.; Eeckhout, M.; De Backer, P.; Croubels, S.; Pfohl-Leszkowicz, A. Characterization of 27 Mycotoxin Binders and the Relation with in Vitro Zearalenone Adsorption at a Single Concentration. Toxins 2015, 7, 21–33. [Google Scholar] [CrossRef]
- Pereyra, C.; Cavaglieri, L.; Chiacchiera, S.; Dalcero, A. The corn influence on the adsorption levels of aflatoxin B1 and zearalenone by yeast cell wall. J. Appl. Microbiol. 2013, 114, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Merrill, M.L.; Bohnert, D.W.; Harmon, D.L.; Craig, A.M.; Schrick, F.N. The ability of a yeast-derived cell wall preparation to minimize the toxic effects of high-ergot alkaloid tall fescue straw in beef cattle. J. Anim. Sci. 2007, 85, 2596–2605. [Google Scholar] [CrossRef]
- Bhatti, S.A.; Khan, M.Z.; Hassan, Z.U.; Saleemi, M.K.; Saqib, M.; Khatoon, A.; Akhter, M. Comparative efficacy of bentonite clay, activated charcoal and Trichosporon mycotoxinivorans in regulating the feed-to-tissue transfer of mycotoxins. J. Sci. Food Agric. 2018, 98, 884–890. [Google Scholar] [CrossRef]
- Qu, D.; Huang, X.; Han, J.; Man, N. Efficacy of mixed adsorbent in ameliorating ochratoxicosis in broilers fed ochratoxin A contaminated diets. Ital. J. Anim. Sci. 2017, 16, 573–579. [Google Scholar] [CrossRef]
- European Commission. Commission regulation (EC) No. 386/2009 of 12 May 2009 amending Regulation (EC) No. 1831/2003 of the European Parliament and of the Council as regards the establishment of a new functional group of feed additives. Off. J. Eur. Union 2009, 118, 66. [Google Scholar]
- Huwig, A.; Freimund, S.; Käppeli, O.; Dutler, H. Mycotoxin detoxication of animal feed by different adsorbents. Toxicol. Lett. 2001, 122, 179–188. [Google Scholar] [CrossRef]
- Joannis-Cassan, C.; Tozlovanu, M.; Hadjeba-Medjdoub, K.; Ballet, N.; Pfohl-Leszkowicz, A. Binding of Zearalenone, Aflatoxin B1, and Ochratoxin A by Yeast-Based Products: A Method for Quantification of Adsorption Performance. J. Prot. 2011, 74, 1175–1185. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.Z.; Wang, G.Y.; Yang, Z.B.; Zhang, G.G.; Wu, Y.B.; Li, Z. Effects of Fusarium mycotoxins with yeast cell wall absorbent on hematology, serum biochemistry, and oxidative stress in broiler chickens. J. Appl. Poult. Res. 2014, 23, 165–173. [Google Scholar] [CrossRef]
- Magnoli, A.; Poloni, V.; Rojo, M.; Combina, M.; Chiacchiera, S.; Dalcero, A.; Rodriguez, M.; Cavaglieri, L. Novel yeast isolated from broilers’ feedstuff, gut and faeces as aflatoxin B 1 adsorbents. J. Appl. Microbiol. 2016, 121, 1766–1776. [Google Scholar] [CrossRef] [PubMed]
- Mendieta, C.R.; Gómez, G.V.; Del Río, J.C.G.; Cueva, A.C.; Arce, J.M.; Ávila, E.G. Effect of the Addition of Saccharomyces cerevisiae yeast cell wall to diets with mycotoxinson on the performance and immune responses of broilers. J. Poult. Sci. 2018, 55, 38–46. [Google Scholar] [CrossRef]
- Liu, N.; Wang, J.Q.; Jia, S.C.; Chen, Y.K.; Wang, J.P. Effect of yeast cell wall on the growth performance and gut health of broilers challenged with aflatoxin B1 and necrotic enteritis. Poult. Sci. 2018, 97, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Pfohl-Leszkowicz, A.; Hadjeba-Medjdoub, K.; Ballet, N.; Schrickx, J.; Fink-Gremmels, J. Assessment and characterisation of yeast-based products intended to mitigate ochratoxin exposure using in vitro and in vivo models. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2015, 32, 604–616. [Google Scholar] [CrossRef] [PubMed]
- Armando, M.; Pizzolitto, R.; Dogi, C.; Cristofolini, A.; Merkis, C.; Poloni, V.; Dalcero, A.; Cavaglieri, L. Adsorption of ochratoxin A and zearalenone by potential probiotic Saccharomyces cerevisiae strains and its relation with cell wall thickness. J. Appl. Microbiol. 2012, 113, 256–264. [Google Scholar] [CrossRef]
- Keller, L.; Abrunhosa, L.; Keller, K.; Rosa, C.A.; Cavaglieri, L.; Venâncio, A.; Battilani, P.; Rosa, C.A.R. Zearalenone and Its Derivatives α-Zearalenol and β-Zearalenol Decontamination by Saccharomyces cerevisiae Strains Isolated from Bovine Forage. Toxins 2015, 7, 3297–3308. [Google Scholar] [CrossRef]
- Luo, Y.; Wang, J.; Liu, B.; Wang, Z.; Yuan, Y.; Yue, T. Effect of Yeast Cell Morphology, Cell Wall Physical Structure and Chemical Composition on Patulin Adsorption. PLoS ONE 2015, 10, e0136045. [Google Scholar] [CrossRef]
- Pereyra, C.; Gil, S.; Cristofolini, A.; Bonci, M.; Makita, M.; Monge, M.; Montenegro, M.; Cavaglieri, L. The production of yeast cell wall using an agroindustrial waste influences the wall thickness and is implicated on the aflatoxin B1 adsorption process. Res. Int. 2018, 111, 306–313. [Google Scholar] [CrossRef]
- Devegowda, G.; Aravind, B.I.R.; Morton, M.G. Saccharomyces cerevisiae and mannan-oligosaccharides to counteract aflatoxins in broilers. Proc. Aust. Poult. Sci. Symp. 1996, 8, 103–106. [Google Scholar]
- Faucet-Marquis, V.; Joannis-Cassan, C.; Hadjeba-Medjdoub, K.; Ballet, N.; Pfohl-Leszkowicz, A. Development of an in vitro method for the prediction of mycotoxin binding on yeast-based products: case of aflatoxin B1, zearalenone and ochratoxin A. Appl. Microbiol. Biotechnol. 2014, 98, 7583–7596. [Google Scholar] [CrossRef]
- Klis, F.M.; Boorsma, A.; De Groot, P.W.J. Cell wall construction inSaccharomyces cerevisiae. Yeast 2006, 23, 185–202. [Google Scholar] [CrossRef]
- Pereyra, C.M.; Cavaglieri, L.R.; Keller, K.M.; Chiacchera, S.W.; Rosa, C.A.D.R.; Dalcero, A.M. In vitro ochratoxin A adsorption by commercial yeast cell walls. Rev. Bras. Med. Vet. 2015, 37, 25–28. [Google Scholar]
- Bzducha-Wróbel, A.; Błażejak, S.; Kieliszek, M.; Pobiega, K.; Falana, K.; Janowicz, M. Modification of the cell wall structure of Saccharomyces cerevisiae strains during cultivation on waste potato juice water and glycerol towards biosynthesis of functional polysaccharides. J. Biotechnol. 2018, 281, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bzducha-Wróbel, A.; Pobiega, K.; Błażejak, S.; Kieliszek, M. The scale-up cultivation of Candida utilis in waste potato juice water with glycerol affects biomass and β(1,3)/(1,6)-glucan characteristic and yield. Appl. Microbiol. Biotechnol. 2018, 102, 9131–9145. [Google Scholar] [CrossRef] [PubMed]
- Bzducha-Wróbel, A.; Błażejak, S.; Molenda, M.; Reczek, L. Erratum to: Biosynthesis of β(1,3)/(1,6)-glucans of cell wall of the yeast Candida utilis ATCC 9950 strains in the culture media supplemented with deproteinated potato juice water and glycerol. Eur. Food Res. Technol. 2015, 240, 1281–1282. [Google Scholar] [CrossRef]
- Boudergue, C.; Burel, C.; Dragacci, S.; Favrot, M.; Fremy, J.; Massimi, C.; Prigent, P.; Debongnie, P.; Pussemier, L.; Boudra, H.; et al. Review of mycotoxin-detoxifying agents used as feed additives: mode of action, efficacy and feed/food safety. EFSA Support. Publ. 2009, 6. [Google Scholar] [CrossRef]
- Yiannikouris, A.; François, J.; Poughon, L.; Dussap, C.G.; Jeminet, G.; Bertin, G.; Jouany, J.P. Influence of pH on complexing of model beta-d-glucans with zearalenone. J. Prot. 2004, 67, 2741–2746. [Google Scholar]
- Tabari, D.G.; Kermanshahi, H.; Golian, A.; Heravi, R.M.; Department of Animal Science, Faculty of Agriculture, Ferdowsi University of Mashhad, Mashhad, Iran. In Vitro Binding Potentials of Bentonite, Yeast Cell Wall and Lactic Acid Bacteria for Aflatoxin B1 and Ochratoxin A. Iran. J. Toxicol. 2018, 12, 7–13. [Google Scholar]
- Guo, C.; Yuan, Y.; Yue, T.; Hatab, S.; Wang, Z. Binding mechanism of patulin to heat-treated yeast cell. Lett. Appl. Microbiol. 2012, 55, 453–459. [Google Scholar] [CrossRef]
- Petruzzi, L.; Sinigaglia, M.; Corbo, M.R.; Campaniello, D.; Speranza, B.; Bevilacqua, A. Decontamination of ochratoxin A by yeasts: possible approaches and factors leading to toxin removal in wine. Appl. Microbiol. Biotechnol. 2014, 98, 6555–6567. [Google Scholar] [CrossRef] [PubMed]
- Cole, R.J.; Jarvis, B.B.; Schweikert, M.A. Handbook of Secondary Fungal Metabolites; Academic Press: San Diego, CA, USA, 2003; Volume III, pp. 199–324, 545–624. [Google Scholar]
- Lopez-Ribot, J.L.; Casanova, M.; Martinez, J.P.; Sentandreu, R. Characterization of cell wall proteins of yeast and hydrophobic mycelial cells of Candida albicans. Infect. Immun. 1991, 59, 2324–2332. [Google Scholar] [PubMed]
- Fruhauf, S.; Schwartz, H.; Ottner, F.; Krska, R.; Vekiru, E. Yeast cell based feed additives: studies on aflatoxin B 1 and zearalenone. Addit. Contam. A 2012, 29, 217–231. [Google Scholar] [CrossRef] [PubMed]
- Piotrowska, M.; Masek, A.; Manderville, R.A. Saccharomyces Cerevisiae Cell Wall Components as Tools for Ochratoxin A Decontamination. Toxins 2015, 7, 1151–1162. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, B.; Gonçalves, C.; Rosim, R.; Oliveira, C.; Corassin, C. Evaluations of Different Sources of Saccharomyces cerevisiae to Binding Capacity of Aflatoxin B1 Utilizing their Adsorption Isotherms. J. Chem. Nanotechnol. 2017, 3, 126–132. [Google Scholar] [CrossRef]
- Jouany, J.P.; Yiannikouris, A.; Bertin, G. The chemical bonds between mycotoxins and cell wall components of Saccharomyces cerevisiae have been identified. Archiva Zootechnica 2005, 8, 26–50. [Google Scholar]
- Yiannikouris, A.; François, J.; Poughon, L.; Dussap, C.G.; Bertin, G.; Jeminet, G.; Jouany, J.P. Alkali extraction of beta-d-glucans from Saccharomyces cerevisiae cell wall and study of their adsorptive properties toward zearalenone. J. Agric. Food Chem. 2004, 52, 3666–3673. [Google Scholar] [CrossRef] [PubMed]
- Sabater-Vilar, M.; Malekinejad, H.; Selman, M.H.J.; Van Der Doelen, M.A.M.; Fink-Gremmels, J.; Doelen, M.A.M. In vitro assessment of adsorbents aiming to prevent deoxynivalenol and zearalenone mycotoxicoses. Mycopathologia 2007, 163, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Döll, S.; Danicke, S.; Valenta, H.; Flachowsky, G. In vitro studies on the evaluation of mycotoxin detoxifying agents for their efficacy on deoxynivalenol and zearalenone. Arch. Anim. Nutr. 2004, 58, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Avantaggiato, G.; Solfrizzo, M.; Visconti, A. Recent advances on the use of adsorbent materials for detoxification of Fusarium mycotoxins. Addit. Contam. 2005, 22, 379–388. [Google Scholar] [CrossRef]
- Kong, C.; Shin, S.Y.; Kim, B.G. Evaluation of mycotoxin sequestering agents for aflatoxin and deoxynivalenol: an in vitro approach. SpringerPlus 2014, 3, 346. [Google Scholar] [CrossRef] [PubMed]
- Sletmoen, M.; Stokke, B.T. Higher order structure of (1,3)-β-D-glucans and its influence on their biological activities and complexation abilities. Biopolymers 2008, 89, 310–321. [Google Scholar] [CrossRef]
- Zechner-Krpan, V.; Petravić-Tominac, V.; Gospodarić, I.; Sajli, L.; Ðaković, S.; Filipović-Grčić, J. Characterization of β-glucans isolated from brewer’s yeast and dried by different methods. Food Technol. Biotechnol. 2010, 48, 189–197. [Google Scholar]
- Hunter, K.W.; Gult, R.A.; Berner, M.D. Preparation of microparticulate β-glucan from Saccharomyces cerevisiae for use in immune potentiation. Lett. App. Microbiol. 2002, 35, 267–271. [Google Scholar] [CrossRef]
- Hromádková, Z.; Ebringerová, A.; Sasinková, V.; Šandula, J.; Hříbalová, V.; Omelková, J. Influence of the drying method on the physical properties and immunomodulatory activity of the particulate (1, 3)-β-D-glucan from Saccharomyces cerevisiae. Carbohydr. Polym. 2003, 5, 9–15. [Google Scholar] [CrossRef]
- Zechner-Krpan, V.; Petravić-Tominac, V.; Galović, P.; Galović, V.; Filipović-Grčić, J.; Srečec, S. Application of different drying methods on β-glucan isolated from spent brewer’s yeast using alkaline. Agric. Conspec. Sci. 2010, 75, 45–50. [Google Scholar]
- Janowicz, M.; Lenart, A. The impact of high pressure and drying processing on internal structure and quality of fruit. Eur. Food Res. Technol. 2018, 244, 1329–1340. [Google Scholar] [CrossRef]
- Bryła, M.; Waśkiewicz, A.; Podolska, G.; Szymczyk, K.; Jędrzejczak, R.; Damaziak, K.; Sułek, A.; Battilani, P. Occurrence of 26 Mycotoxins in the Grain of Cereals Cultivated in Poland. Toxins 2016, 8, 160. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Preparation | Total Sugars | β(1,3)/(1,6)-Glucan | Protein |
|---|---|---|---|
| [g 100g Preparation] | |||
| CW | 68.7 ± 4.0 a1 | 62.9 ± 0.7 a | 7.6 ± 0.5 b |
| β-G | 93.4 ± 3.1 b | 84.8 ± 1.3 b | 2.0 ± 1.3 a |
| Preparation | Alkali Insoluble Polysaccharides | β(1,3)-Glucan Insoluble in Alkali | β(1,6)-Glucan Insoluble in Alkali | Alkali Soluble Polysaccharides |
|---|---|---|---|---|
| [g 100g preparation] | ||||
| β-G | 75.6 ± 2.5 | 56.2 ± 5.9 (74.3%)* | 19.5 ± 0.5 (25.6)* | 24.4 ± 2.9 |
| Preparation | d10* [um] | d50** [um] | d90*** [um] | D[4,3] **** [um] |
|---|---|---|---|---|
| CW | 7.76 ± 0.56 a | 62.58 ± 4.14 A | 201.91 ± 15.01 A | 85.42 ± 6.29 a |
| β-G | 16.10 ± 0.46 b | 84.98 ± 2.38 B | 208.98 ± 4.70 B | 101.52 ± 2.46 b |
| Adsorption Characteristic | pH | CW | β-G | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AFB1 | ZEN | OTA | DON | NIV | T-2 | FB1 | AFB1 | ZEN | OTA | DON | NIV | T-2 | FB1 | ||
| Mads (ng) | 3.0 | 248 de | 821 h | 446 f | 139 abcd | 41 ab | 72 abc | 101 abc | 170 cde | 743 h | 247 de | 112 abc | 36 ab | 106 abc | 194 cde |
| %ads | 25 DE | 82 H | 45 F | 14 ABCD | 4 AB | 7 ABC | 12 ABC | 17 CDE | 74 H | 24 DE | 11 ABC | 4 AB | 11 ABC | 19 CDE | |
| Cads (µg/g) | 12 lm | 41.1 p | 22 n | 7 ijkl | 2 ij | 5 ijk | 5 ijk | 9 klm | 37 p | 12 lm | 6 ijk | 2 ij | 5 ijk | 10 klm | |
| Kd (mL/g) | 33 | 459 | 81 | 16 | 4 | 8 | 13 | 21 | 289 | 32 | 13 | 4 | 12 | 24 | |
| Mads (ng) | 6.0 | 252 de | 756 h | 117 abc | 215 cd | 95 b | 161 bcd | 114 abc | 291 e | 614 g | 174 cde | 157 bcd | 121 abc | 77 abc | 21 a |
| %ads | 25 DE | 76 H | 12 ABC | 22 CD | 10 B | 16 BCD | 13 ABC | 29 E | 61 G | 17 CDE | 16 BCD | 12 ABC | 8 ABC | 2 A | |
| Cads (µg/g) | 13 lm | 38 p | 6 ijk | 11 kl | 5 j | 8 jki | 6 ijk | 15 m | 31 o | 9 klm | 8 jkl | 6 ijk | 4 ijk | 1 i | |
| Kd (mL/g) | 34 | 310 | 13 | 22 | 11 | 19 | 14 | 41 | 159 | 21 | 19 | 13 | 8 | 2 | |
| Mycotoxin | RSD (%) |
|---|---|
| AFB1 | 5.1 (n = 5; 450 ng/mL) 6.9 (n = 5; 250 ng/mL) |
| ZEN | 4.2 (n = 5; 450 ng/mL) 12.2 (n = 5; 100 ng/mL) |
| OTA | 4.0 (n = 5; 450 ng/mL) 7.4 (n = 5; 200 ng/mL) |
| DON | 5.5 (n = 5; 450 ng/mL) |
| NIV | 4.5 (n = 5; 450 ng/mL) |
| T-2 toxin | 4.8 (n = 5; 450 ng/mL) |
| FB1 | 5.9 (n = 5; 450 ng/mL) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bzducha-Wróbel, A.; Bryła, M.; Gientka, I.; Błażejak, S.; Janowicz, M. Candida utilis ATCC 9950 Cell Walls and β(1,3)/(1,6)-Glucan Preparations Produced Using Agro-Waste as a Mycotoxins Trap. Toxins 2019, 11, 192. https://doi.org/10.3390/toxins11040192
Bzducha-Wróbel A, Bryła M, Gientka I, Błażejak S, Janowicz M. Candida utilis ATCC 9950 Cell Walls and β(1,3)/(1,6)-Glucan Preparations Produced Using Agro-Waste as a Mycotoxins Trap. Toxins. 2019; 11(4):192. https://doi.org/10.3390/toxins11040192
Chicago/Turabian StyleBzducha-Wróbel, Anna, Marcin Bryła, Iwona Gientka, Stanisław Błażejak, and Monika Janowicz. 2019. "Candida utilis ATCC 9950 Cell Walls and β(1,3)/(1,6)-Glucan Preparations Produced Using Agro-Waste as a Mycotoxins Trap" Toxins 11, no. 4: 192. https://doi.org/10.3390/toxins11040192
APA StyleBzducha-Wróbel, A., Bryła, M., Gientka, I., Błażejak, S., & Janowicz, M. (2019). Candida utilis ATCC 9950 Cell Walls and β(1,3)/(1,6)-Glucan Preparations Produced Using Agro-Waste as a Mycotoxins Trap. Toxins, 11(4), 192. https://doi.org/10.3390/toxins11040192