Proteins in Soy Might Have a Higher Role in Cancer Prevention than Previously Expected: Soybean Protein Fractions Are More Effective MMP-9 Inhibitors Than Non-Protein Fractions, Even in Cooked Seeds


Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Materials
2.2. Soy Seed Soaking and Heat Treatment
2.3. Extraction and Quantification of Soluble Proteins
2.4. Extraction and Quantification of Non-Protein Compounds
2.4.1. Saponin Quantification
2.4.2. Polyphenolic Compounds
2.5. Half Maximal Effective Inhibition of MMPs (IC50)
2.6. Specific MMP-9 Inhibition
2.7. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis
2.8. Reverse Gelatin Zymography
2.9. In Vitro Colon Cancer Cell Assays
2.9.1. Cell Migration Assay
2.9.2. Cell Proliferation Assay
2.10. Substrate Gelatin Zymography
2.11. Statistical Analysis
3. Results and Discussion
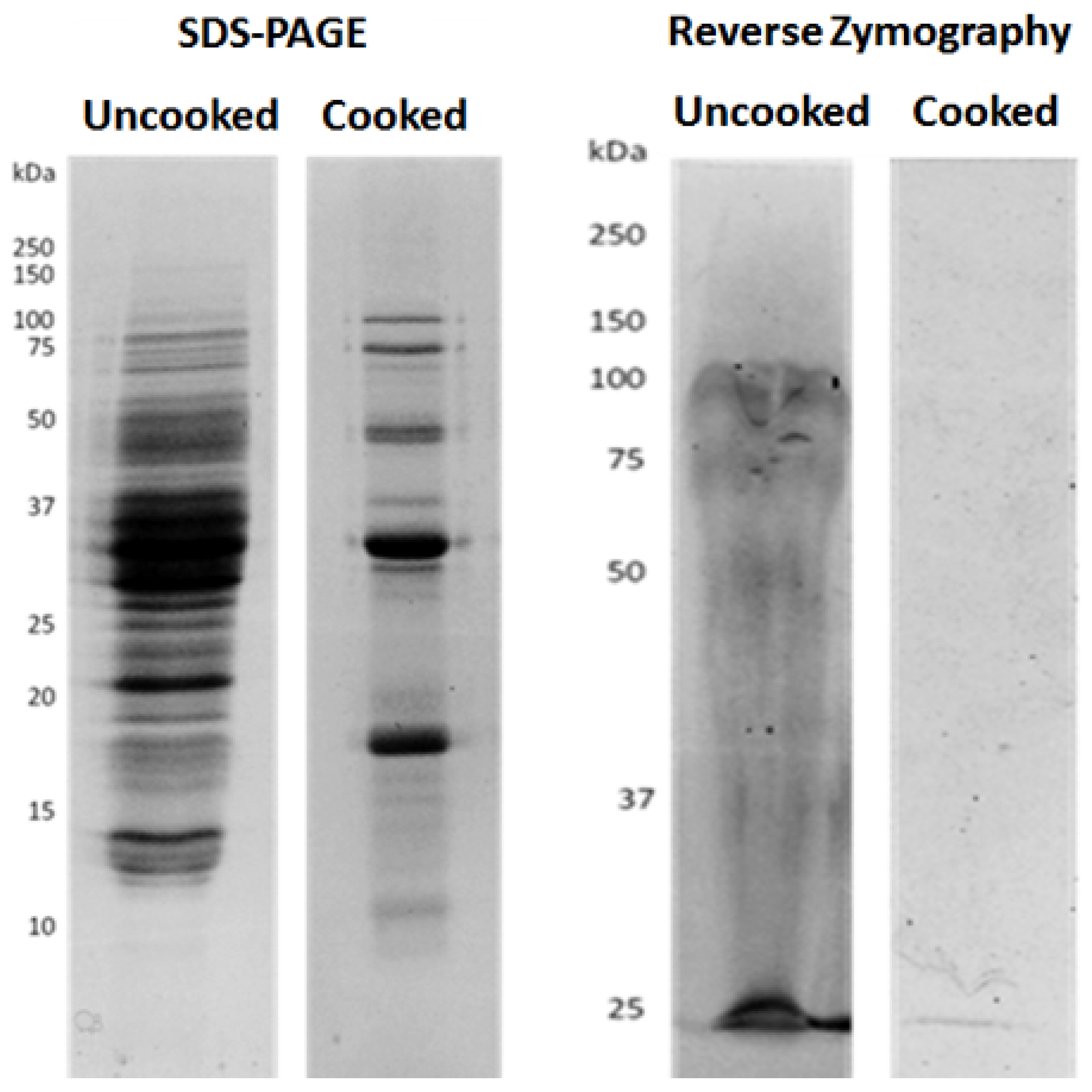
3.1. Cooking Affects the Amounts of Bioactive Compounds in Soy Seeds
3.2. Proteins in Soy are Better MMP Inhibitors Than Polyphenols and Saponins
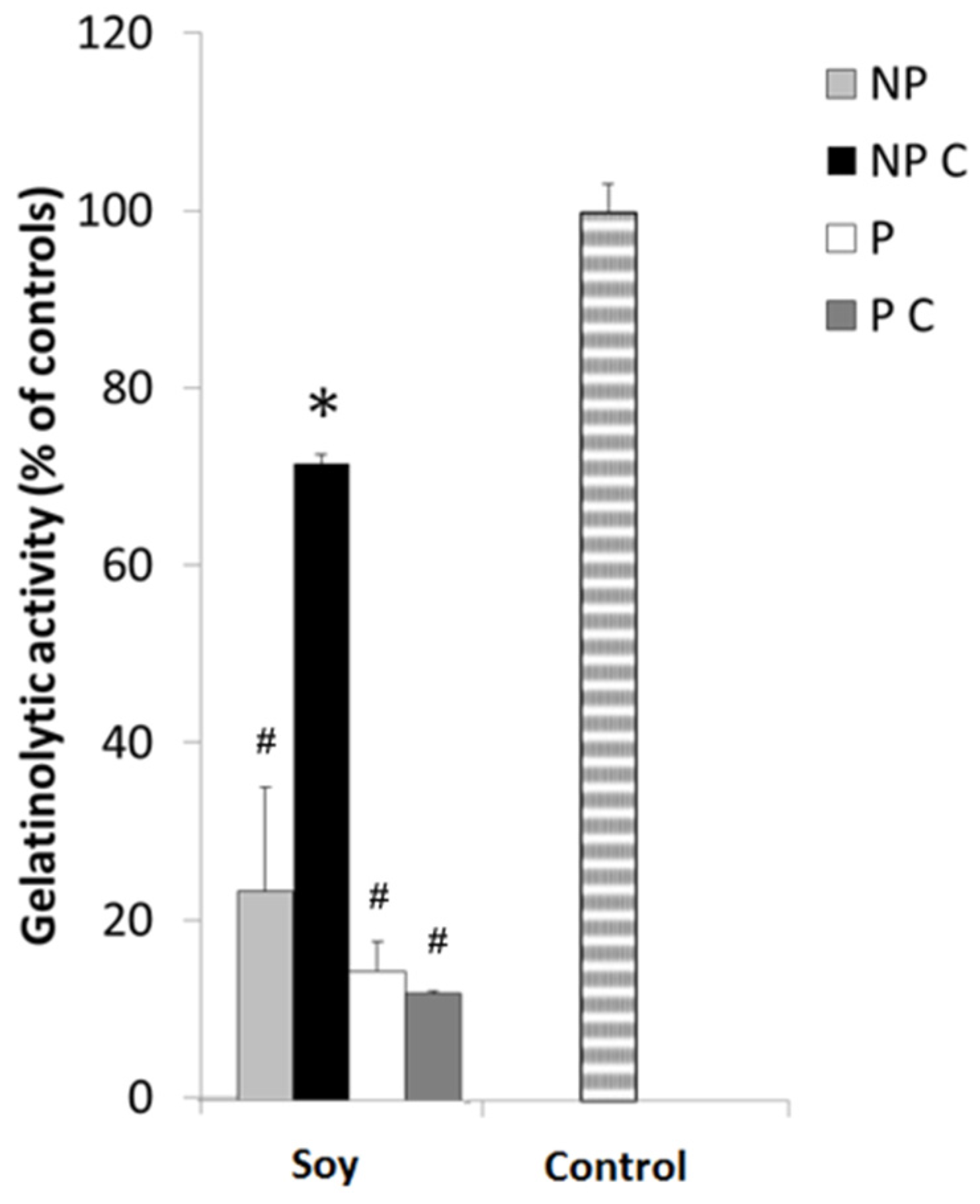
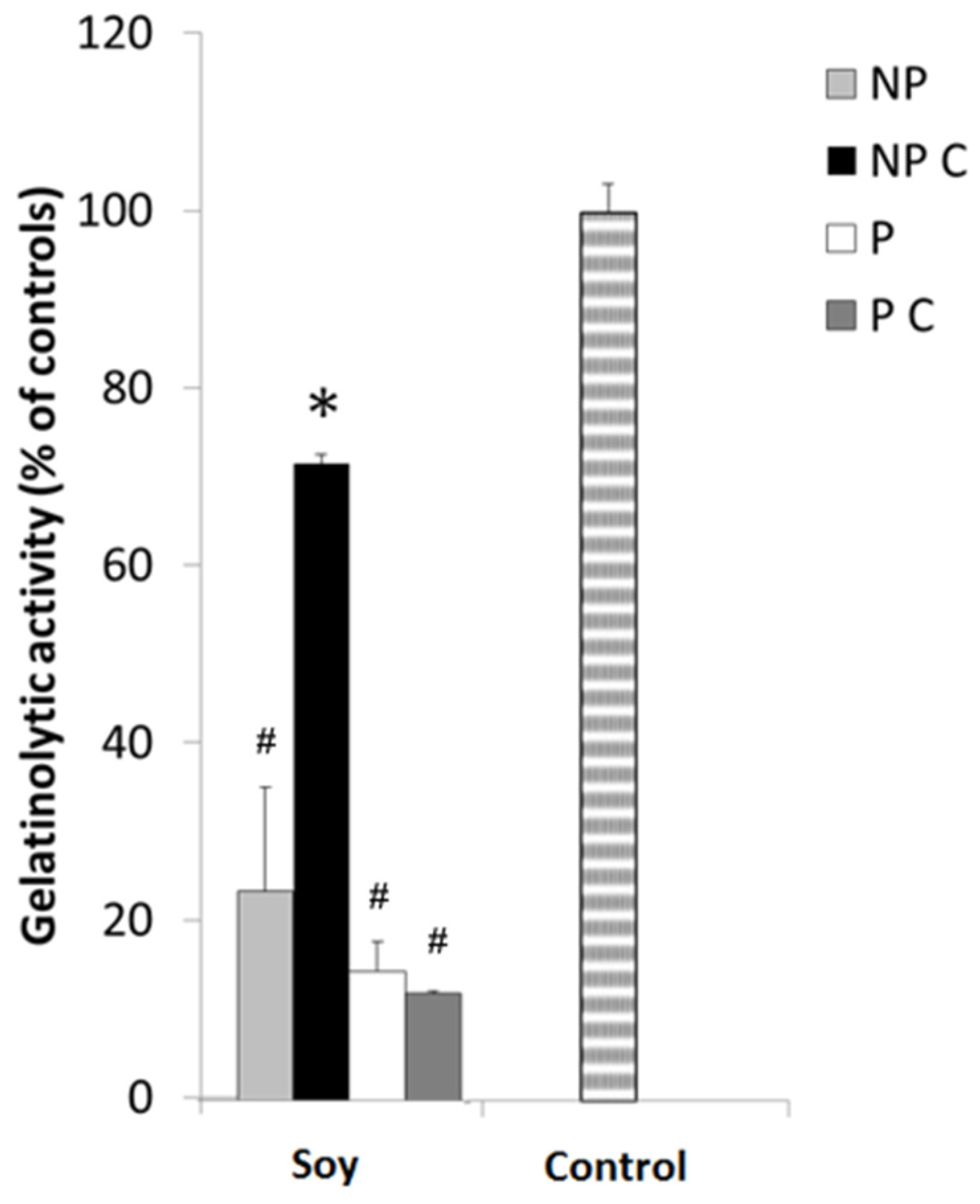
3.3. Cooking Affects MMP-9 Inhibitory Activities of Non-Protein Compounds, but Not of MMPI Proteins
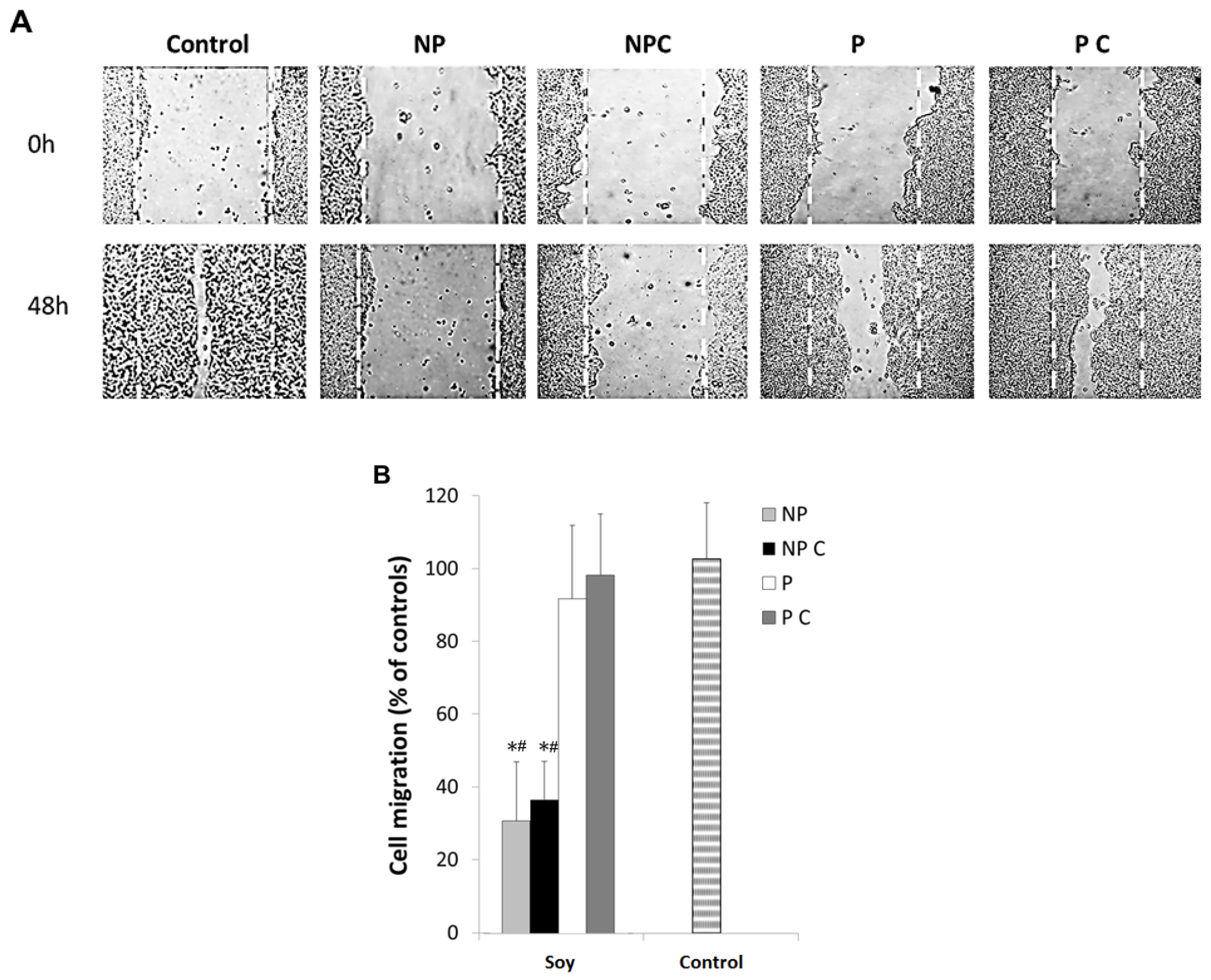
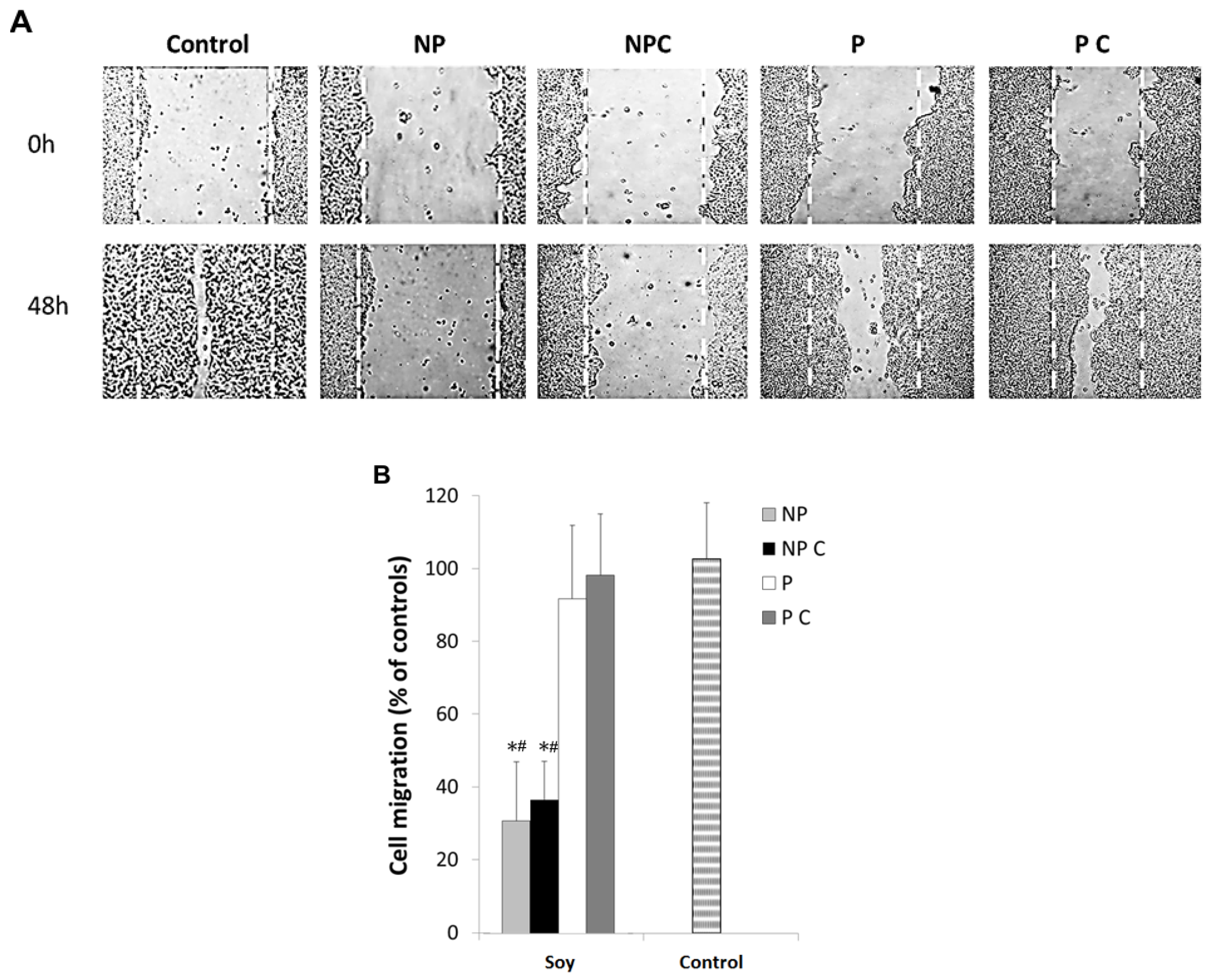
3.4. Soy Polyphenols Reduce Cancer Cell Migration Even without Inhibiting MMP-9
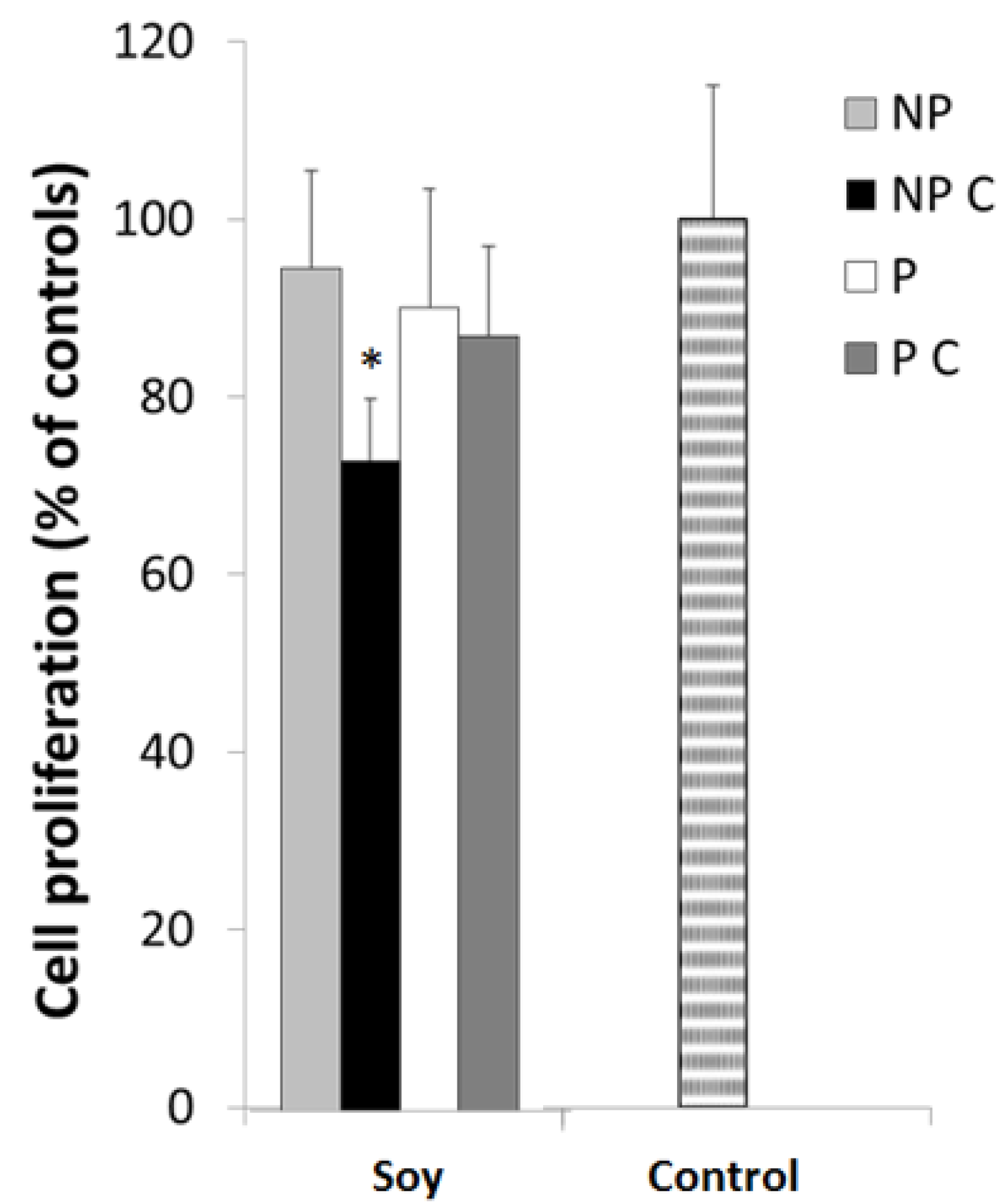
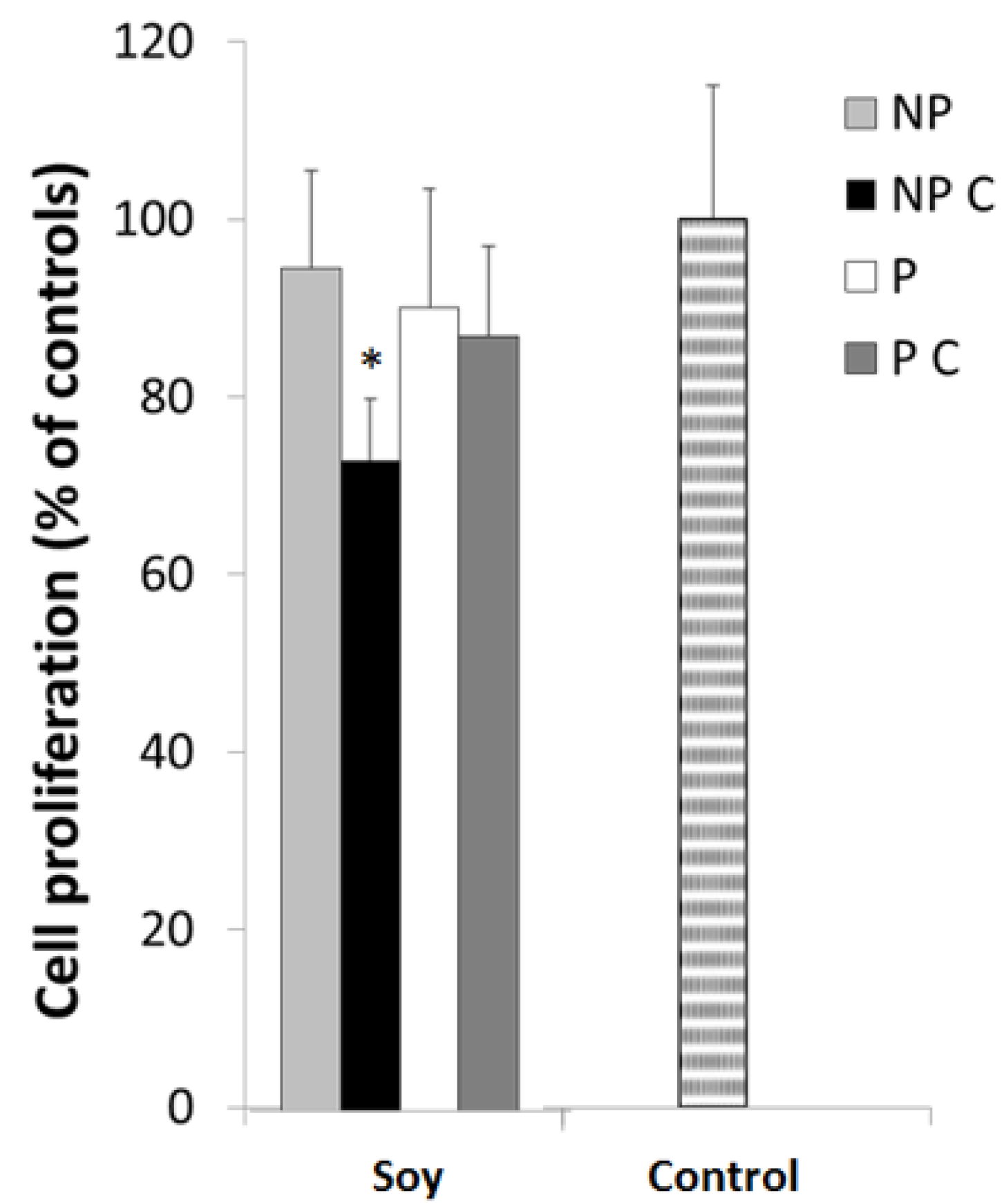
3.5. Cancer Cell Proliferation
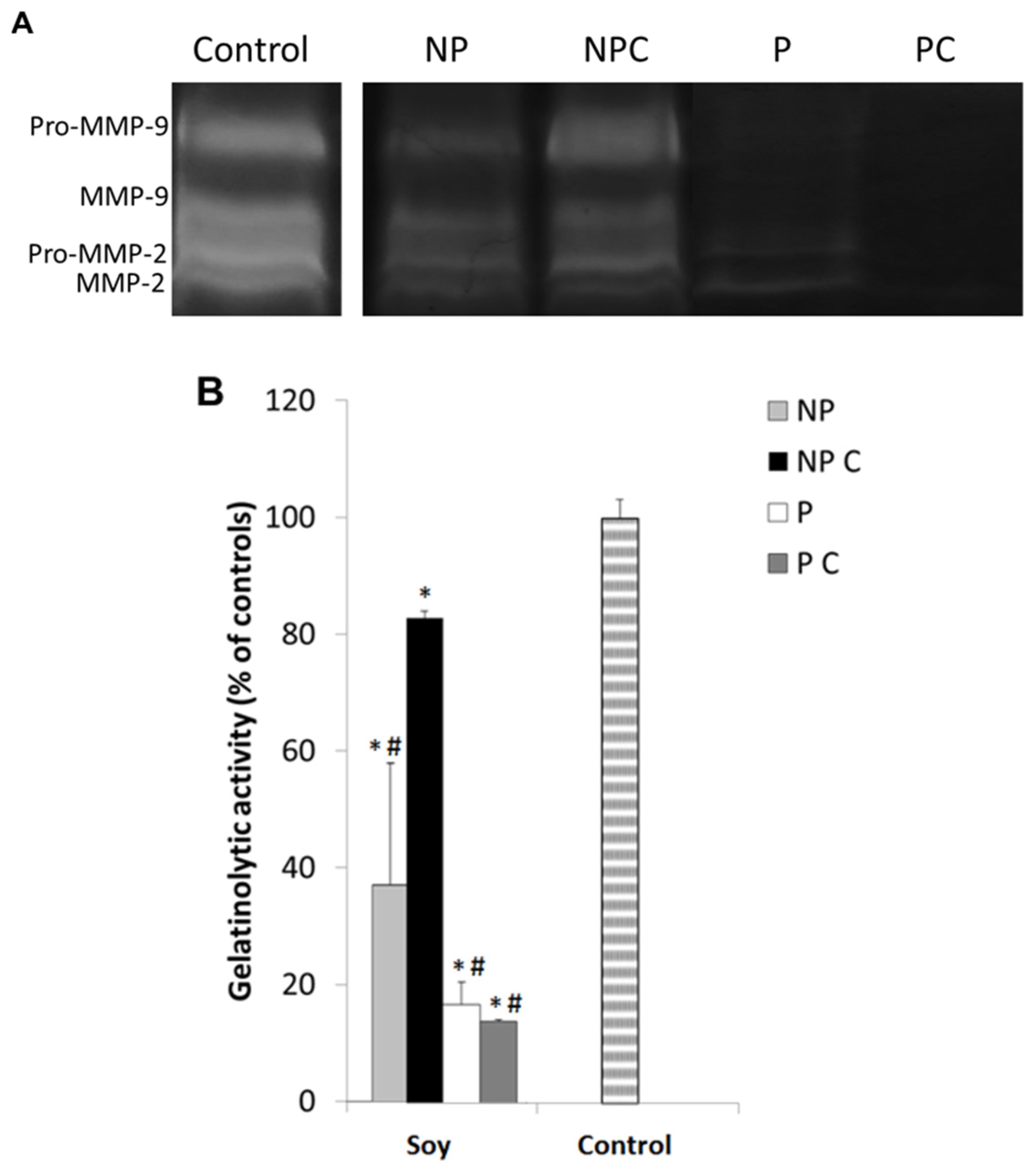
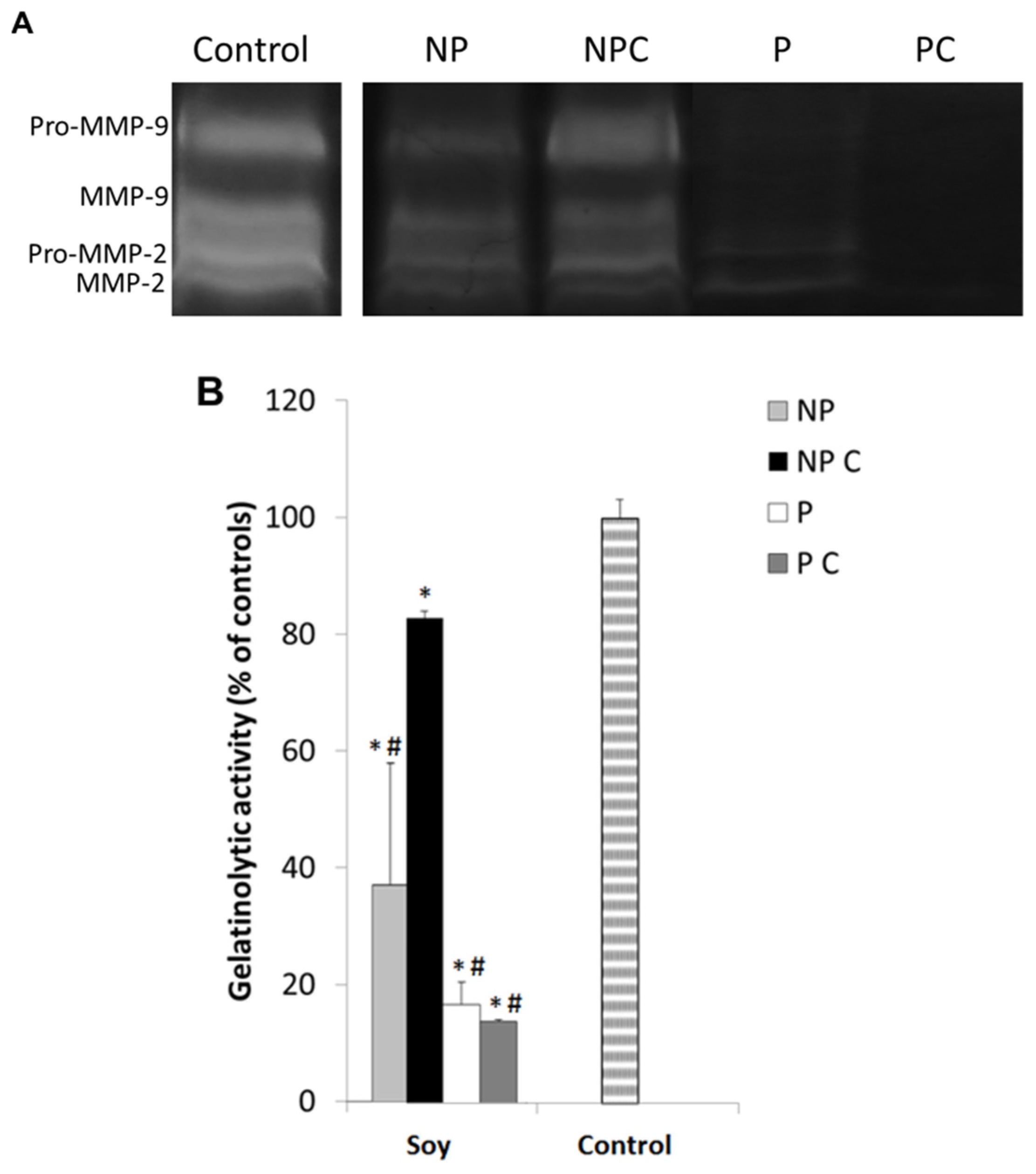
3.6. Soy Extracts Inhibit MMP-9 in HT-29 Extracellular Media, but Not MMP-2
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of interest
References
- Markle, B.; May, E.J.; Majumdar, A.P.N. Do nutraceutics play a role in the prevention and treatment of colorectal cancer? Cancer Metastasis Rev. 2010, 29, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Malla, N.; Sjøli, S.; Winberg, J.O.; Hadler-Olsen, E.; Uhlin-Hansen, L. Biological and pathobiological functions of gelatinase dimers and complexes. Connect. Tissue Res. 2008, 49, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Herszényi, L.; Hritz, I.; Lakatos, G.; Varga, M.Z.; Tulassay, Z. The behavior of matrix metalloproteinases and their inhibitors in colorectal cancer. Int. J. Mol. Sci. 2012, 13, 13240–13263. [Google Scholar] [CrossRef] [PubMed]
- Baugh, M.D.; Perry, M.J.; Hollander, A.P.; Davies, D.R.; Cross, S.S.; Lobo, A.J.; Taylor, C.J.; Evans, G.S. Matrix metalloproteinase levels are elevated in inflammatory bowel disease. Gastroenterology 1999, 117, 814–822. [Google Scholar] [CrossRef]
- Parks, W.C.; Wilson, C.L.; López-Boado, Y.S. Matrix metalloproteinases as modulators of inflammation and innate immunity. Nat. Rev. Immunol. 2004, 4, 617–629. [Google Scholar] [CrossRef] [PubMed]
- Murphy, G.; Nagase, H. Progress in matrix metalloproteinase research. Mol. Aspects Med. 2009, 29, 290–308. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.K.; Vansaun, M.N.; Shim, J.H.; Matrisian, L.M.; Gorden, D.L. Increased metastases are associated with inflammation and matrix metalloproteinase-9 activity at incision sites in a murine model of peritoneal dissemination of colorectal cancer. J. Surg. Res. 2013, 180, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Garg, P.; Vijay-Kumar, M.; Wang, L.; Gewirtz, A.T.; Merlin, D.; Sitaraman, S.V. Matrix metalloproteinase-9-mediated tissue injury overrides the protective effect of matrix metalloproteinase-2 during colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Moore, B.A.; Manthey, C.L.; Johnson, D.L.; Bauer, A.J. Matrix Metalloproteinase-9 Inhibition Reduces Inflammation and Improves Motility in Murine Models of Postoperative Ileus. Gastroenterology 2011, 141, 1283–1292. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, M.; Eckhardt, S.G. Development of Matrix Metalloproteinase Inhibitors in Cancer Therapy. J. Natl. Cancer Inst. 2001, 93, 178–193. [Google Scholar] [CrossRef] [PubMed]
- Coussens, L.M.; Fingleton, B.; Matrisian, L.M. Matrix Metalloproteinase Inhibitors and Cancer: Trials and Tribulations. Science 2002, 295, 2387–2393. [Google Scholar] [CrossRef] [PubMed]
- Mook, O.R.F.; Frederiks, W.M.; van Noorden, C.J.F. The role of gelatinases in colorectal cancer progression and metastasis. Biochim. Biophys. Acta 2004, 1705, 69–89. [Google Scholar] [CrossRef] [PubMed]
- Zucker, S.; Vacirca, J. Role of matrix metalloproteinases (MMPs) in colorectal cancer. Cancer Metastasis Rev. 2004, 23, 101–117. [Google Scholar] [CrossRef] [PubMed]
- Sang, S.; Ju, J.; Lambert, J.D.; Lin, Y.; Hong, J.; Bose, M.; Wang, S.; Bai, N.; He, K.; Reddy, B.S.; et al. Wheat bran oil and its fractions inhibit human colon cancer cell growth and intestinal tumorigenesis in Apcmin/+ mice. J. Agric. Food Chem. 2006, 54, 9792–9797. [Google Scholar] [CrossRef] [PubMed]
- Ndinguri, M.W.; Bhowmick, M.; Tokmina-Roszyk, D.; Robichaud, T.K.; Fields, G.B. Peptide-based selective inhibitors of matrix metalloproteinase-mediated activities. Molecules 2012, 17, 14230–14248. [Google Scholar] [CrossRef] [PubMed]
- Gaultier, F.; Foucault-Bertaud, A.; Lamy, E.; Ejeil, A.L.; Dridi, S.M.; Piccardi, N.; Piccirilli, A.; Msika, P.; Godeau, G.; Gogly, B. Effects of a vegetable extract from Lupinus albus (LU105) on the production of matrix metalloproteinases (MMP1, MMP2, MMP9) and tissue inhibitor of metalloproteinases (TIMP1, TIMP2) by human gingival fibroblasts in culture. Clin. Oral Investig. 2003, 7, 198–205. [Google Scholar] [PubMed]
- Craig, W.J.; Mangels, A.R. Position of the American Dietetic Association: Vegetarian diets. J. Am. Diet. Assoc. 2009, 109, 1266–1282. [Google Scholar] [PubMed]
- Lima, A.I.G.; Mota, J.; Monteiro, S.A.V.S.; Ferreira, R.M.S.B. Legume seeds and colorectal cancer revisited: Protease inhibitors reduce MMP-9 activity and colon cancer cell migration. Food Chem. 2016, 197, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Adlercreutz, H.; Mazur, W. Phyto-oestrogens and Western diseases. Ann. Med. 1997, 29, 95–120. [Google Scholar] [CrossRef] [PubMed]
- Fournier, D.B.; Erdman, J.W., Jr.; Gordon, G.B. Soy, its components, and cancer prevention: A review of the in vitro, animal, and human data. Cancer Epidemiol. Biomark. Prev. 1998, 7, 1055–1065. [Google Scholar]
- Messina, M.J.; Persky, V.; Setchell, K.D.; Barnes, S. Soy intake and cancer risk: A review of the in vitro and in vivo data. Nutr. Cancer 1994, 21, 113–131. [Google Scholar] [CrossRef] [PubMed]
- Messina, M.; Gugger, E.T.; Alekel, D.L. Soy protein, soybean isoflavones, and bone health: A review of the animal and human data. In Handbook of Nutraceuticals and Functional Foods; Wildman, R., Ed.; CRC Press: Boca Raton, FL, USA, 2001; pp. 77–98. [Google Scholar]
- Anderson, J.J.; Garner, S.C. Phytoestrogens and bone. Baillieres Clin. Endocrinol. Metab. 1998, 12, 543–557. [Google Scholar] [CrossRef]
- Erdman, J.W., Jr. Soy protein and cardiovascular disease: A statement for healthcare professionals from the nutrition committee of the AHA. Circulation 2000, 102, 2555–2559. [Google Scholar] [CrossRef] [PubMed]
- Messina, M.; Barnes, S. The role of soy products in reducing risk of cancer. J. Natl. Cancer Inst. 1991, 83, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Champ, M.M. Non-nutrient bioactive substances of pulses. Br. J. Nutr. 2002, 88, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Campos-Veja, R.; Loarca-Piña, G.; Oomah, B.D. Minor components of pulses and their potential impact on human health. Food Res. Int. 2010, 43, 461–482. [Google Scholar] [CrossRef]
- Xu, B.; Chang, S.K. Comparative study on antiproliferation properties and cellular antioxidant activities of commonly consumed food legumes against nine human cancer cell lines. Food Chem. 2012, 134, 1287–1296. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, A.R. The Bowman-Birk inhibitor from soybeans as an anticarcinogenic agent. Am. J. Clin. Nutr. 1998, 68, 1406–1412. [Google Scholar]
- Kyle, S.; James, K.A.R.; McPherson, M.J. Recombinant production of the therapeutic peptide lunasin. Microb. Cell Fact. 2012, 11, 28. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.H.; Han, I.H.; Sung, M.K.; Yoo, H.; Kim, Y.G.; Kim, J.S.; Kawada, T.; Yu, R. Soybean saponin inhibits tumor cell metastasis by modulating expressions of MMP-2, MMP-9 and TIMP-2. Cancer Lett. 2008, 261, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, A.R.; Wan, X.S. Effects of the Bowman-Birk inhibitor on growth, invasion, and clonogenic survival of human prostate epithelial cells and prostate cancer cells. Prostate 2002, 50, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Fereidunian, A.; Sadeghalvad, M.; Oscoie, M.O.; Mostafaie, A. Soybean Bowman-Birk Protease Inhibitor (BBI): Identification of the Mechanisms of BBI Suppressive Effect on Growth of Two Adenocarcinoma Cell Lines: AGS and HT29. Arch. Med. Res. 2014, 45, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, A.R. The evidence for soybean products as cancer preventive agents. J. Nutr. 1995, 125, 733–743. [Google Scholar]
- Losso, J.N. The biochemical and functional food properties of the bowman-birk inhibitor. Crit. Rev. Food Sci. Nutr. 2008, 48, 94–118. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Jeong, H.J.; de Lumen, B.O. In Vitro Digestibility of the Cancer-Preventive Soy Peptides Lunasin and BBI. J. Agric. Food Chem. 2007, 55, 10703–10706. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Pan, Y.; Cheng, Y.; Li, H.; Liu, D.; Li, H. Lunasin suppresses the migration and invasion of breast cancer cells by inhibiting matrix metalloproteinase-2/-9 via the FAK/Akt/ERK and NF-κB signaling pathways. Oncol. Rep. 2016, 36, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Chang, S.K.C. Effect of soaking, boiling, and steaming on total phenolic contentand antioxidant activities of cool season food legumes. Food Chem. 2008, 110, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Makkar, H.P.S.; Becker, K. Nutrients and antiquality factors in different morphological parts of the Moringa oleifera tree. J. Agric. Sci. 1997, 128, 311–322. [Google Scholar] [CrossRef]
- Hiai, S.; Oura, H.; Nakajima, T. Color reaction of some sapogenins and saponins with vanillin and sulfuric acid. Planta Med. 1976, 29, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin–Ciocalteau reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef] [PubMed]
- Bouhdid, S.; Abrini, J.; Amensour, M.; Zhiri, A.; Espuny, M.J.; Manresa, A. Functional and ultrastructural changes in Pseudomonas aeruginosa and Staphylococcus aureus cells induced by Cinnamomum verum essential oil. J. Appl. Microbiol. 2010, 109, 1139–1149. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, S.P.; Li, H.; Taniguchi, G.T. Zymography and Reverse Zymography for Detecting MMPs and TIMPs. Methods Mol. Biol. 2001, 622, 257–269. [Google Scholar]
- Carmichael, J.; DeGraff, W.G.; Gazdar, A.F.; Minna, J.D.; Mitchell, J.B. Evaluation of a tetrazolium-based semiautomated colorimetric assay: Assessment of chemosensitivity testing. Cancer Res. 1987, 47, 936–942. [Google Scholar] [PubMed]
- Toth, M.; Sohail, A.; Fridman, R. Assessment of Gelatinases (MMP-2 and MMP-9) by Gelatin Zymography. Methods Mol. Biol. 2012, 878, 121–135. [Google Scholar] [PubMed]
- Messina, M.J.; Loprinzi, C.L. Soy for breast cancer survivors: A critical review of the literature. J. Nutr. 2001, 131, 3095–3108. [Google Scholar]
- Takezaki, T.; Gao, C.M.; Ding, J.H.; Liu, T.K.; Li, M.S.; Tajima, K. Comparative study of lifestyles of residents in high and low risk areas for gastric cancer in Jiangsu Province, China; with special reference to allium vegetables. J. Epidemiol. 1999, 9, 297–305. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lee, S.A.; Shu, X.O.; Li, H.; Yang, G.; Cai, H.; Wen, W.; Ji, B.T.; Gao, J.; Gao, Y.T.; Zheng, W. Adolescent and adult soy food intake and breast cancer risk: Results from the Shanghai Women’s Health Study. Am. J. Clin. Nutr. 2009, 89, 1920–1926. [Google Scholar] [CrossRef] [PubMed]
- Patisaul, H.B.; Jefferson, W. The pros and cons of phytoestrogens. Front. Neuroendocrinol. 2010, 31, 400–419. [Google Scholar] [CrossRef] [PubMed]
- Rochfort, S.; Panozzo, J. Phytochemicals for health, the role of pulses. J. Agric. Food Chem. 2007, 55, 7981–7994. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, M.; Hoshino, M.; Katsube, Y.; Hayashi, H.; Kushiro, T.; Ebizuka, Y. Identification of beta-amyrin and sophoradiol 24-hydroxylase by expressed sequence tag mining and functional expression assay. FEBS J. 2006, 273, 948–959. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, R.S.; Guo, J.; Copeland, J.; Browning, J.D., Jr.; Sleper, D.; Rottinghaus, G.E.; Berhow, M.A. Environmental influences on isoflavones and saponins in soybeans and their role in colon cancer. J. Nutr. 2005, 135, 1239–1242. [Google Scholar] [PubMed]
- Gurfinkel, D.M.; Rao, A.V. Soya saponins: The relationship between chemical structure and colon anticarcinogenic activity. Nutr. Cancer 2003, 47, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.X.; Huang, G.Q.; Zhang, S.H. Soya saponins inhibit the proliferation of Hela cells by inducing apoptosis. Exp. Toxicol. Pathol. 2007, 59, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.Y.; Chen, Y.H.; Chien, Y.W.; Huang, W.H.; Lin, S.H. Effect of soy saponin on the growth of human colon cancer cells. World J. Gastroenterol. 2010, 16, 3371–3376. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.H.; Sung, M.K.; Kawada, T.; Yoo, H.; Kim, Y.K.; Kim, J.S. Soybean saponins suppress the release of proinflammatory mediators by LPS-stimulated peritoneal macrophages. Cancer Lett. 2005, 230, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, L.; Gomes, J.C.; Stringheta, P.C.; Gontijo, A.M.; Padovani, C.R.; Ribeiro, L.R.; Salvadori, D.M. Black bean (Phaseolus vulgaris L.) as a protective agent against DNA damage in mice. Food Chem. Toxicol. 2003, 41, 1671–1676. [Google Scholar] [CrossRef]
- Cardador-Martinez, A.; Loarca-Pina, G.; Oomah, B.D. Antioxidant activity in common beans (Phaseolus vulgaris L.). J. Agric. Food Chem. 2002, 50, 6975–6980. [Google Scholar] [CrossRef] [PubMed]
- Trock, B.J.; Hilakivi-Clarke, L.; Clarke, R. Meta-analysis of soy intake and breast cancer risk. J. Natl. Cancer Inst. 2006, 98, 459–471. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Spitznagel, E.L. Meta-analysis of soy food and risk of prostate cancer in men. Int. J. Cancer 2005, 117, 667–669. [Google Scholar] [CrossRef] [PubMed]
- Cassidy, A.; Albertazzi, P.; Lise Nielsen, I.; Hall, W.; Williamson, G.; Tetens, I.; Atkins, S.; Cross, H.; Manios, Y.; Wolk, A.; et al. Critical review of health effects of soyabean phyto-oestrogens in post-menopausal women. Proc. Nutr. Soc. 2006, 65, 76–92. [Google Scholar] [CrossRef] [PubMed]
- Messina, M.; McCaskill-Stevens, W.; Lampe, J.W. Addressing the soy and breast cancer relationship: Review, commentary, and workshop proceedings. J. Natl. Cancer Inst. 2006, 98, 1275–1284. [Google Scholar] [CrossRef] [PubMed]
- Ware, J.H.; Wan, X.S.; Newberne, P.; Kennedy, A.R. Bowman-Birk inhibitor concentrate reduces colon inflammation in mice with dextran sulfate sodium-induced ulcerative colitis. Dig. Dis. Sci. 1999, 44, 986–990. [Google Scholar] [CrossRef] [PubMed]
- Roy, F.; Boye, J.I.; Simpson, B.K. Bioactive proteins and peptides in pulse crops: Pea, chickpea and lentil. Food Res. Int. 2010, 43, 432–442. [Google Scholar] [CrossRef]
- Muzquiz, M.; Varela, A.; Burbano, C.; Cuadrado, C.; Guillamón, E.; Pedrosa, M. Bioactive compounds in legumes: Pronutritive and antinutritive actions. Implications for nutrition and health. Phytochem. Rev. 2012, 11, 227–244. [Google Scholar] [CrossRef]
- El-Adawy, T. Nutritional composition and antinutritional factors of chickpeas (Cicer arietinum L.) undergoing different cooking methods and germination. Plant Foods Hum. Nutr. 2002, 57, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Clemente, A.; Sánchez-Vioque, R.; Vioque, J.; Bautista, J.; Millán, F. Effect of cooking on protein quality of chickpea (Cicer arietinum) seeds. Food Chem. 1998, 62, 1–6. [Google Scholar] [CrossRef]
- Nicoli, M.C.; Anese, M.; Parpinel, M. Influence of processing on the antioxidant properties of fruit and vegetables. Trends Food Sci. Technol. 1999, 10, 94–100. [Google Scholar] [CrossRef]
- Singh, U. Antinutritional factors of chickpea and pigeonpea and their removal by processing. Plant Foods Hum. Nutr. 1988, 38, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Chang, S.K. Reduction of antiproliferative capacities, cell-based antioxidant capacities and phytochemical contents of common beans and soybeans upon thermal processing. Food Chem. 2011, 129, 974–981. [Google Scholar] [CrossRef] [PubMed]
- Ranilla, L.G.; Genovese, M.I.; Lajolo, F.M. Effect of different cooking conditions on phenolic compounds and antioxidant capacity of some selected Brazilian bean (Phaseolus vulgaris L.) cultivars. J. Agric. Food Chem. 2009, 57, 5734–5742. [Google Scholar] [CrossRef] [PubMed]
- Bourguet, E.; Hornebeck, W.; Sapi, J.; Alix, A.J.P.; Moroy, G. Pharmacomodulation of broad spectrum matrix metalloproteinase inhibitors towards regulation of gelatinases. In Enzyme Inhibition and Bioapplications; Sharma, R., Ed.; InTech: Rijeka, Croatia, 2012. [Google Scholar]
- Ferlay, J.; Steliarova-Foucher, E.; Lortet-Tieulent, J.; Rosso, S.; Coebergh, J.W.W.; Comber, H.; Forman, D.; Bray, F. Cancer incidence and mortality patterns in Europe: estimates for 40 countries in 2012. Eur. J. Cancer 2013, 49, 1374–1403. [Google Scholar] [CrossRef] [PubMed]
- Center, M.M.; Jemal, A.; Ward, E. International trends in colorectal cancer incidence rates. Cancer Epidemiol. Biomark. Prev. 2009, 18, 1688–1694. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.; Naishadham, D.; Jemal, A. Cancer statistics, 2012. CA Cancer J. Clin. 2012, 62, 283–298. [Google Scholar] [CrossRef] [PubMed]
- Murthy, N.S.; Mukherjee, S.; Ray, G.; Ray, A. Dietary factors and cancer chemoprevention: An overview of obesity-related malignancies. J. Postgrad. Med. 2009, 55, 45–54. [Google Scholar] [PubMed]
- Deryugina, E.I.; Quigley, J.P. Matrix metalloproteinases and tumor metastasis. Cancer Metastasis Rev. 2006, 25, 9–34. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Tang, F.; Zhang, B.; Zhao, Y.; Feng, J.; Rao, Z. Matrix metalloproteinase-9 overexpression is closely related to poor prognosis in patients with colon cancer. World J. Surg. Oncol. 2014, 12. [Google Scholar] [CrossRef] [PubMed]
- Magee, P.J.; Owusu-Apenten, R.; McCann, M.J.; Gill, C.I.; Rowland, I.R. Chickpea (Cicer arietinum) and other plant-derived protease inhibitor concentrates inhibit breast and prostate cancer cell proliferation in vitro. Nutr. Cancer 2012, 64, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, A.; Giri, A.P.; Harsulkar, A.M.; Gatehouse, J.A.; Gupta, V.S. A Kunitz trypsin inhibitor from chickpea (Cicer arietinum L.) that exerts anti-metabolic effect on podborer (Helicoverpa armigera) larvae. Plant Mol. Biol. 2005, 57, 359–374. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.W.; Lee, H.J. The roles of polyphenols in cancer chemoprevention. BioFactors 2006, 26, 105–121. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Saponins mg/g DW | Phenolics mg/g DW | Proteins mg/g DW | |
|---|---|---|---|
| UC | 3.6 ± 0.24 | 1.7 ± 0.16 | 297.4 ± 8.65 |
| C | 1.6 ± 0.24 | 0.7 ± 0.06 | 15.7 ± 4.82 |
| % of lost | 57.6 a | 60.9 a | 94.7 a |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lima, A.; Oliveira, J.; Saúde, F.; Mota, J.; Ferreira, R.B. Proteins in Soy Might Have a Higher Role in Cancer Prevention than Previously Expected: Soybean Protein Fractions Are More Effective MMP-9 Inhibitors Than Non-Protein Fractions, Even in Cooked Seeds. Nutrients 2017, 9, 201. https://doi.org/10.3390/nu9030201
Lima A, Oliveira J, Saúde F, Mota J, Ferreira RB. Proteins in Soy Might Have a Higher Role in Cancer Prevention than Previously Expected: Soybean Protein Fractions Are More Effective MMP-9 Inhibitors Than Non-Protein Fractions, Even in Cooked Seeds. Nutrients. 2017; 9(3):201. https://doi.org/10.3390/nu9030201
Chicago/Turabian StyleLima, Ana, Jennifer Oliveira, Filipe Saúde, Joana Mota, and Ricardo Boavida Ferreira. 2017. "Proteins in Soy Might Have a Higher Role in Cancer Prevention than Previously Expected: Soybean Protein Fractions Are More Effective MMP-9 Inhibitors Than Non-Protein Fractions, Even in Cooked Seeds" Nutrients 9, no. 3: 201. https://doi.org/10.3390/nu9030201
APA StyleLima, A., Oliveira, J., Saúde, F., Mota, J., & Ferreira, R. B. (2017). Proteins in Soy Might Have a Higher Role in Cancer Prevention than Previously Expected: Soybean Protein Fractions Are More Effective MMP-9 Inhibitors Than Non-Protein Fractions, Even in Cooked Seeds. Nutrients, 9(3), 201. https://doi.org/10.3390/nu9030201