
Monounsaturated Fatty Acids in Cardiovascular Disease: Intake, Individual Types, and Content in Adipose Tissue as a Biomarker of Endogenous Exposure
 , , , , , and
, , , , , and Highlights
- Monounsaturated fatty acids may influence several biological processes involved in atherosclerosis; however, recent studies have raised concern that they might increase the risk of cardiovascular disease.
- Individual types of monounsaturated fatty acids exert distinct biological effects and should be studied separately.
- Exposure to monounsaturated fatty acids is caused by both dietary intake and endogenous synthesis.
- The adipose tissue content of monounsaturated fatty acids represents the long-term exposure, but correlations vary depending on their specific type.
- Future studies should focus on individual monounsaturated fatty acids, their biological effects on the development of atherosclerotic cardiovascular disease, their food sources, and their dietary patterns, and preferably use the adipose tissue content of monounsaturated fatty acids as a measure of exposure.
Abstract
1. Introduction
2. Dietary Sources of MUFAs

{kind=link}
{kind=link}
| Name | Known Dietary Source |
|---|---|
| Oleic acid (18:1ω-9) | Vegetable oils (e.g., olive oil, rapeseed oil, sunflower oil), many plant and animal foods [22,24,32,33]. |
| Vaccenic acid (18:1ω-7) | Ruminant meat and dairy products [34] *, cod liver oil [33] |
| Palmitoleic acid (16:1ω-7) | Fatty fish (e.g., salmon) and fish oils (e.g., cod liver oil), nuts (e.g., macadamia nuts and their oil), sea buckhorn [33,35]. |
| Myristoleic acid (14:1ω-5) | Gap in literature. No comprehensive studies on food sources. |
| Gadoleic acid (20:1ω-9) | Cod liver oil and jojoba oil [33,36]. Otherwise, gap in literature. No comprehensive studies on food sources. |
| Gondoic acid (20:1ω-11) | Cod liver oil [33]. Otherwise, gap in literature. No comprehensive studies on food sources. |
| Erucic acid (22:1ω-9) | Rapeseed oil, jojoba oil and fish [33,36,37,38,39]. |
| Cetoleic acid (22:1ω-11) | Herring oil, cod liver oil, and jojoba oil [33,36,40]. |
3. Endogenous Pathway of MUFA Synthesis and Metabolism
4. The Content of MUFAs in Adipose Tissue and Correlations with MUFA Intake
5. MUFAs and the Risk of ASCVD
6. Conclusions and Perspectives
- (1)
- Investigation of the biological effects of individual types of MUFAs on the development of atherosclerosis. Preferably, this should be carried out in human studies, but animal studies may be of value.
- (2)
- Investigation of the importance of food sources for individual MUFAs and their associated risk of ASCVD. Preferably, this should be carried out in clinical trials, but food science studies investigating the MUFA effects and composition of different foods would be valuable, too.
- (3)
- Investigation of the associations between total and individual MUFA and ASCVD risk using complementary measures of exposure, such as estimated dietary intakes and content of MUFAs in blood or, preferably, adipose tissue. Future clinical trials or cohort studies in humans may benefit from using adipose tissue biopsies to indicate MUFA exposure, ideally obtaining several biopsies over time to reflect MUFA turnover and changes in dietary patterns.
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ASCVD | Atherosclerotic cardiovascular disease |
| FAs | Fatty acids |
| SFA | Saturated fatty acids |
| PUFAs | Polyunsaturated fatty acids |
| MUFAs | Monounsaturated fatty acids |
| SCD1 | Stearoyl-CoA desaturase 1 |
| ELOVL5 | Fatty acid elongase 5 |
| Δ6D | Δ-6-desaturase |
| FFQs | Food frequency questionnaires |
| LDL | Low-density lipoprotein |
| LCMUFAs | Long-chain MUFAs |
| BWHHS | British Women’s Heart and Healthy Study |
| SABRE | Southall and Brent Revisited |
References
- European Heart Network: Transforming European Food and Drink Policies for Cardiovascular Health 2017 [Version 1 29 September 2017]. Available online: https://ehnheart.org/wp-content/uploads/2023/05/Transforming-European-food-and-drink-policies_summary.pdf (accessed on 22 April 2025).
- Sacks, F.M.; Lichtenstein, A.H.; Wu, J.H.Y.; Appel, L.J.; Creager, M.A.; Kris-Etherton, P.M.; Miller, M.; Rimm, E.B.; Rudel, L.L.; Robinson, J.G.; et al. Dietary Fats and Cardiovascular Disease: A Presidential Advisory From the American Heart Association. Circulation 2017, 136, e1–e23. [Google Scholar] [CrossRef]
- Visseren, F.L.J.; Mach, F.; Smulders, Y.M.; Carballo, D.; Koskinas, K.C.; Bäck, M.; Benetos, A.; Biffi, A.; Boavida, J.M.; Capodanno, D.; et al. 2021 ESC Guidelines on cardiovascular disease prevention in clinical practice. Eur. Heart J. 2021, 42, 3227–3337. [Google Scholar] [CrossRef]
- Li, Y.; Hruby, A.; Bernstein, A.M.; Ley, S.H.; Wang, D.D.; Chiuve, S.E.; Sampson, L.; Rexrode, K.M.; Rimm, E.B.; Willett, W.C.; et al. Saturated Fats Compared with Unsaturated Fats and Sources of Carbohydrates in Relation to Risk of Coronary Heart Disease: A Prospective Cohort Study. J. Am. Coll. Cardiol. 2015, 66, 1538–1548. [Google Scholar] [CrossRef]
- Mehrabani, A.; Jalalzadeh, M.; Jannati, N.; Lotfi, K.; Arzhang, P.; Azadbakht, L. Association Between Monounsaturated Fatty Acid Intake and Risk of Total Stroke and Its Subtypes: A Systematic Review and Dose-Response Meta-analysis of Prospective Cohort Studies. Nutr. Rev. 2025, 83, 1035–1047. [Google Scholar] [CrossRef] [PubMed]
- Cheng, P.; Wang, J.; Shao, W. Monounsaturated Fatty Acid Intake and Stroke Risk: A Meta-analysis of Prospective Cohort Studies. J. Stroke Cerebrovasc. Dis. 2016, 25, 1326–1334. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.; Martin, N.; Jimoh, O.F.; Kirk, C.; Foster, E.; Abdelhamid, A.S. Reduction in saturated fat intake for cardiovascular disease. Cochrane Database Syst. Rev. 2020, 5, Cd011737. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.D.; Li, Y.; Chiuve, S.E.; Stampfer, M.J.; Manson, J.E.; Rimm, E.B.; Willett, W.C.; Hu, F.B. Association of Specific Dietary Fats with Total and Cause-Specific Mortality. JAMA Intern Med. 2016, 176, 1134–1145. [Google Scholar] [CrossRef]
- Arnesen, E.K.; Laake, I.; Veierød, M.B.; Retterstøl, K. Saturated fatty acids and total and CVD mortality in Norway: A prospective cohort study with up to 45 years of follow-up. Br. J. Nutr. 2024, 132, 466–478. [Google Scholar] [CrossRef]
- Guasch-Ferré, M.; Babio, N.; Martínez-González, M.A.; Corella, D.; Ros, E.; Martín-Peláez, S.; Estruch, R.; Arós, F.; Gómez-Gracia, E.; Fiol, M.; et al. Dietary fat intake and risk of cardiovascular disease and all-cause mortality in a population at high risk of cardiovascular disease. Am. J. Clin. Nutr. 2015, 102, 1563–1573. [Google Scholar] [CrossRef]
- Ho, F.K.; Gray, S.R.; Welsh, P.; Petermann-Rocha, F.; Foster, H.; Waddell, H.; Anderson, J.; Lyall, D.; Sattar, N.; Gill, J.M.R.; et al. Associations of fat and carbohydrate intake with cardiovascular disease and mortality: Prospective cohort study of UK Biobank participants. BMJ 2020, 368, m688. [Google Scholar] [CrossRef]
- Zhu, Y.; Bo, Y.; Liu, Y. Dietary total fat, fatty acids intake, and risk of cardiovascular disease: A dose-response meta-analysis of cohort studies. Lipids Health Dis. 2019, 18, 91. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zheng, Z.; Zhuang, L.; Wang, H.; Li, A.; Chen, L.; Liu, L. Dietary Macronutrient Intake and Cardiovascular Disease Risk and Mortality: A Systematic Review and Dose-Response Meta-Analysis of Prospective Cohort Studies. Nutrients 2024, 16, 152. [Google Scholar] [CrossRef] [PubMed]
- Estruch, R.; Ros, E.; Salas-Salvadó, J.; Covas, M.I.; Corella, D.; Arós, F.; Gómez-Gracia, E.; Ruiz-Gutiérrez, V.; Fiol, M.; Lapetra, J.; et al. Primary Prevention of Cardiovascular Disease with a Mediterranean Diet Supplemented with Extra-Virgin Olive Oil or Nuts. N. Engl. J. Med. 2018, 378, e34. [Google Scholar] [CrossRef] [PubMed]
- Praagman, J.; Beulens, J.W.; Alssema, M.; Zock, P.L.; Wanders, A.J.; Sluijs, I.; van der Schouw, Y.T. The association between dietary saturated fatty acids and ischemic heart disease depends on the type and source of fatty acid in the European Prospective Investigation into Cancer and Nutrition-Netherlands cohort. Am. J. Clin. Nutr. 2016, 103, 356–365. [Google Scholar] [CrossRef]
- Praagman, J.; de Jonge, E.A.; Kiefte-de Jong, J.C.; Beulens, J.W.; Sluijs, I.; Schoufour, J.D.; Hofman, A.; van der Schouw, Y.T.; Franco, O.H. Dietary Saturated Fatty Acids and Coronary Heart Disease Risk in a Dutch Middle-Aged and Elderly Population. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 2011–2018. [Google Scholar] [CrossRef]
- Jin, D.; Trichia, E.; Islam, N.; Lewington, S.; Lacey, B. Associations of circulating fatty acids with incident coronary heart disease: A prospective study of 89, 242 individuals in UK Biobank. BMC Cardiovasc. Disord. 2023, 23, 365. [Google Scholar] [CrossRef]
- Würtz, P.; Havulinna, A.S.; Soininen, P.; Tynkkynen, T.; Prieto-Merino, D.; Tillin, T.; Ghorbani, A.; Artati, A.; Wang, Q.; Tiainen, M.; et al. Metabolite profiling and cardiovascular event risk: A prospective study of 3 population-based cohorts. Circulation 2015, 131, 774–785. [Google Scholar] [CrossRef]
- Zhuang, P.; Liu, X.; Li, Y.; Wu, Y.; Li, H.; Wan, X.; Zhang, L.; Xu, C.; Jiao, J.; Zhang, Y. Circulating fatty acids, genetic risk, and incident coronary artery disease: A prospective, longitudinal cohort study. Sci. Adv. 2023, 9, eadf9037. [Google Scholar] [CrossRef]
- Borges, M.C.; Schmidt, A.F.; Jefferis, B.; Wannamethee, S.G.; Lawlor, D.A.; Kivimaki, M.; Kumari, M.; Gaunt, T.R.; Ben-Shlomo, Y.; Tillin, T.; et al. Circulating Fatty Acids and Risk of Coronary Heart Disease and Stroke: Individual Participant Data Meta-Analysis in Up to 16 126 Participants. J. Am. Heart Assoc. 2020, 9, e013131. [Google Scholar] [CrossRef]
- Mazidi, M.; Katsiki, N.; Shekoohi, N.; Banach, M. Monounsaturated Fatty Acid Levels May Not Affect Cardiovascular Events: Results from a Mendelian Randomization Analysis. Front. Nutr. 2020, 7, 123. [Google Scholar] [CrossRef]
- Ivey, K.L.; Nguyen, X.T.; Li, R.; Furtado, J.; Cho, K.; Gaziano, J.M.; Hu, F.B.; Willett, W.C.; Wilson, P.W.; Djoussé, L. Association of dietary fatty acids with the risk of atherosclerotic cardiovascular disease in a prospective cohort of United States veterans. Am. J. Clin. Nutr. 2023, 118, 763–772. [Google Scholar] [CrossRef]
- Kris-Etherton, P.M. AHA science advisory: Monounsaturated fatty acids and risk of cardiovascular disease. J. Nutr. 1999, 129, 2280–2284. [Google Scholar] [CrossRef]
- Zock, P.L.; Blom, W.A.; Nettleton, J.A.; Hornstra, G. Progressing Insights into the Role of Dietary Fats in the Prevention of Cardiovascular Disease. Curr. Cardiol. Rep. 2016, 18, 111. [Google Scholar] [CrossRef] [PubMed]
- Christensen, J.J.; Arnesen, E.K.; Rundblad, A.; Telle-Hansen, V.H.; Narverud, I.; Blomhoff, R.; Bogsrud, M.P.; Retterstøl, K.; Ulven, S.M.; Holven, K.B. Dietary fat quality, plasma atherogenic lipoproteins, and atherosclerotic cardiovascular disease: An overview of the rationale for dietary recommendations for fat intake. Atherosclerosis 2024, 389, 117433. [Google Scholar] [CrossRef] [PubMed]
- Hodson, L.; Skeaff, C.M.; Fielding, B.A. Fatty acid composition of adipose tissue and blood in humans and its use as a biomarker of dietary intake. Prog. Lipid Res. 2008, 47, 348–380. [Google Scholar] [CrossRef]
- Coelho, M.; Oliveira, T.; Fernandes, R. Biochemistry of adipose tissue: An endocrine organ. Arch. Med. Sci. 2013, 9, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Tjønneland, A.; Overvad, K.; Thorling, E.; Ewertz, M. Adipose tissue fatty acids as biomarkers of dietary exposure in Danish men and women. Am. J. Clin. Nutr. 1993, 57, 629–633. [Google Scholar] [CrossRef]
- van Dijk, S.J.; Feskens, E.J.; Bos, M.B.; Hoelen, D.W.; Heijligenberg, R.; Bromhaar, M.G.; de Groot, L.C.P.G.M.; de Vries, J.H.M.; Müller, M.; Afman, L.A. A saturated fatty acid-rich diet induces an obesity-linked proinflammatory gene expression profile in adipose tissue of subjects at risk of metabolic syndrome. Am. J. Clin. Nutr. 2009, 90, 1656–1664. [Google Scholar] [CrossRef]
- Calder, P.C. Functional Roles of Fatty Acids and Their Effects on Human Health. JPEN J. Parenter Enteral. Nutr. 2015, 39, 18s–32s. [Google Scholar] [CrossRef]
- Kiani, A.K.; Medori, M.C.; Bonetti, G.; Aquilanti, B.; Velluti, V.; Matera, G.; Laconelli, A.; Stuppia, L.; Connelly, S.T.; Herbst, K.L.; et al. Modern vision of the Mediterranean diet. J. Prev. Med. Hyg. 2022, 63, E36–E43. [Google Scholar]
- Mennella, I.; Savarese, M.; Ferracane, R.; Sacchi, R.; Vitaglione, P. Oleic acid content of a meal promotes oleoylethanolamide response and reduces subsequent energy intake in humans. Food Funct. 2015, 6, 204–210. [Google Scholar] [CrossRef]
- Loftsson, T.; Gudmundsdóttir, T.K.; Fridriksdóttir, H.; Sigurdardóttir, A.M.; Thorkelsson, J.; Gudmundsson, G.; Hjaltason, B. Fatty acids from cod-liver oil as skin penetration enhancers. Pharmazie 1995, 50, 188–190. [Google Scholar] [PubMed]
- Turpeinen, A.M.; Mutanen, M.; Aro, A.; Salminen, I.; Basu, S.; Palmquist, D.L.; Griinari, J.M. Bioconversion of vaccenic acid to conjugated linoleic acid in humans. Am. J. Clin. Nutr. 2002, 76, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Frigolet, M.E.; Gutiérrez-Aguilar, R. The Role of the Novel Lipokine Palmitoleic Acid in Health and Disease. Adv. Nutr. 2017, 8, 173s–181s. [Google Scholar] [CrossRef] [PubMed]
- Awad, N.A.; Eliraq, M.; El-Bassel, E.H.; Ismail, A.S.M.; Abd El-Aziz, Y.S.G.; Gawish, M.S.; Zewail, R.M.Y.; Sami, R.; Khojah, E.; Hilary, U.; et al. Evaluation of the Effect of Elite Jojoba Lines on the Chemical Properties of their Seed Oil. Molecules 2022, 27, 3904. [Google Scholar] [CrossRef]
- Galanty, A.; Grudzińska, M.; Paździora, W.; Paśko, P. Erucic Acid-Both Sides of the Story: A Concise Review on Its Beneficial and Toxic Properties. Molecules 2023, 28, 1924. [Google Scholar] [CrossRef]
- Russo, M.; Yan, F.; Stier, A.; Klasen, L.; Honermeier, B. Erucic acid concentration of rapeseed (Brassica napus L.) oils on the German food retail market. Food Sci. Nutr. 2021, 9, 3664–3672. [Google Scholar] [CrossRef]
- Sissener, N.H.; Ørnsrud, R.; Sanden, M.; Frøyland, L.; Remø, S.; Lundebye, A.K. Erucic Acid (22:1n-9) in Fish Feed, Farmed, and Wild Fish and Seafood Products. Nutrients 2018, 10, 1443. [Google Scholar] [CrossRef]
- Aro, T.L.; Larmo, P.S.; Bäckman, C.H.; Kallio, H.P.; Tahvonen, R.L. Fatty acids and fat-soluble vitamins in salted herring (Clupea harengus) products. J. Agric. Food Chem. 2005, 53, 1482–1488. [Google Scholar] [CrossRef]
- Shi, F.; Chowdhury, R.; Sofianopoulou, E.; Koulman, A.; Sun, L.; Steur, M.; Aleksandrova, K.; Dahm, C.C.; Schulze, M.B.; van der Schouw, Y.T.; et al. Association of circulating fatty acids with cardiovascular disease risk: Analysis of individual-level data in three large prospective cohorts and updated meta-analysis. Eur. J. Prev. Cardiol. 2025, 32, 233–246. [Google Scholar] [CrossRef]
- Steur, M.; Johnson, L.; Sharp, S.J.; Imamura, F.; Sluijs, I.; Key, T.J.; Wood, A.; Chowdhury, R.; Guevara, M.; Jakobsen, M.U.; et al. Dietary Fatty Acids, Macronutrient Substitutions, Food Sources and Incidence of Coronary Heart Disease: Findings From the EPIC-CVD Case-Cohort Study Across Nine European Countries. J. Am. Heart Assoc. 2021, 10, e019814. [Google Scholar] [CrossRef]
- Tjønneland, A.; Olsen, A.; Boll, K.; Stripp, C.; Christensen, J.; Engholm, G.; Overvad, K. Study design, exposure variables, and socioeconomic determinants of participation in Diet, Cancer and Health: A population-based prospective cohort study of 57, 053 men and women in Denmark. Scand. J. Public Health 2007, 35, 432–441. [Google Scholar] [CrossRef]
- Tjønneland, A.; Overvad, K.; Haraldsdóttir, J.; Bang, S.; Ewertz, M.; Jensen, O.M. Validation of a semiquantitative food frequency questionnaire developed in Denmark. Int. J. Epidemiol. 1991, 20, 906–912. [Google Scholar] [CrossRef]
- Bork, C.; Dahm, C.C.; Calder, P.C.; Lundbye-Christensen, S.; Olsen, A.; Overvad, K.; Schmidt, E.B. Adipose tissue content of ω-6 polyunsaturated fatty acids and all-cause mortality: A Danish prospective cohort study. Am. J. Clin. Nutr. 2025, 121, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Lasota, A.N.; Grønholdt, M.M.; Bork, C.S.; Lundbye-Christensen, S.; Overvad, K.; Schmidt, E.B. Marine n-3 Fatty Acids and the Risk of Peripheral Arterial Disease. J. Am. Coll. Cardiol. 2018, 72, 1576–1584. [Google Scholar] [CrossRef] [PubMed]
- Yee, J.K.; Mao, C.S.; Hummel, H.S.; Lim, S.; Sugano, S.; Rehan, V.K.; Xiao, G.; Lee, W.N.P. Compartmentalization of stearoyl-coenzyme A desaturase 1 activity in HepG2 cells. J. Lipid Res. 2008, 49, 2124–2134. [Google Scholar] [CrossRef] [PubMed]
- Borges, M.C.; Haycock, P.; Zheng, J.; Hemani, G.; Howe, L.J.; Schmidt, A.F.; Staley, J.R.; Lumbers, R.T.; Henry, A.; Lemaitre, R.N.; et al. The impact of fatty acids biosynthesis on the risk of cardiovascular diseases in Europeans and East Asians: A Mendelian randomization study. Hum. Mol. Genet. 2022, 31, 4034–4054. [Google Scholar] [CrossRef]
- Sun, Q.; Xing, X.; Wang, H.; Wan, K.; Fan, R.; Liu, C.; Wang, Y.; Wu, W.; Wang, Y.; Wang, R. SCD1 is the critical signaling hub to mediate metabolic diseases: Mechanism and the development of its inhibitors. Biomed. Pharmacother. 2024, 170, 115586. [Google Scholar] [CrossRef]
- Vessby, B.; Gustafsson, I.B.; Tengblad, S.; Berglund, L. Indices of fatty acid desaturase activity in healthy human subjects: Effects of different types of dietary fat. Br. J. Nutr. 2013, 110, 871–879. [Google Scholar] [CrossRef]
- Eissing, L.; Scherer, T.; Tödter, K.; Knippschild, U.; Greve, J.W.; Buurman, W.A.; Pinnschmidt, H.O.; Rensen, S.S.; Wolf, A.M.; Bartelt, A.; et al. De novo lipogenesis in human fat and liver is linked to ChREBP-β and metabolic health. Nat. Commun. 2013, 4, 1528. [Google Scholar] [CrossRef]
- Green, C.D.; Ozguden-Akkoc, C.G.; Wang, Y.; Jump, D.B.; Olson, L.K. Role of fatty acid elongases in determination of de novo synthesized monounsaturated fatty acid species. J. Lipid Res. 2010, 51, 1871–1877. [Google Scholar] [CrossRef]
- Ohno, Y.; Suto, S.; Yamanaka, M.; Mizutani, Y.; Mitsutake, S.; Igarashi, Y.; Sassa, T.; Kihara, A. ELOVL1 production of C24 acyl-CoAs is linked to C24 sphingolipid synthesis. Proc. Natl. Acad. Sci. USA 2010, 107, 18439–18444. [Google Scholar] [CrossRef]
- Zadravec, D.; Brolinson, A.; Fisher, R.M.; Carneheim, C.; Csikasz, R.I.; Bertrand-Michel, J.; Borén, J.; Guilluo, H.; Rudling, M.; Jacobsson, A. Ablation of the very-long-chain fatty acid elongase ELOVL3 in mice leads to constrained lipid storage and resistance to diet-induced obesity. FASEB J. 2010, 24, 4366–4377. [Google Scholar]
- Centenera, M.M.; Scott, J.S.; Machiels, J.; Nassar, Z.D.; Miller, D.C.; Zinonos, I.; Dehairs, J.; Burvenich, I.J.G.; Zadra, G.; Chetta, P.M.; et al. ELOVL5 Is a Critical and Targetable Fatty Acid Elongase in Prostate Cancer. Cancer Res. 2021, 81, 1704–1718. [Google Scholar] [CrossRef]
- Østbye, T.K.; Berge, G.M.; Nilsson, A.; Romarheim, O.H.; Bou, M.; Ruyter, B. The long-chain monounsaturated cetoleic acid improves the efficiency of the n-3 fatty acid metabolic pathway in Atlantic salmon and human HepG2 cells. Br. J. Nutr. 2019, 122, 755–768. [Google Scholar] [CrossRef]
- Tobin, D.; Midtbø, L.K.; Mildenberger, J.; Svensen, H.; Stoknes, I. The effect of fish oil rich in cetoleic acid on the omega-3 index and skin quality. Prostaglandins Leukot. Essent. Fat. Acids 2024, 201, 102616. [Google Scholar] [CrossRef]
- Park, H.G.; Engel, M.G.; Vogt-Lowell, K.; Lawrence, P.; Kothapalli, K.S.; Brenna, J.T. The role of fatty acid desaturase (FADS) genes in oleic acid metabolism: FADS1 Δ7 desaturates 11–20:1 to 7,11–20:2. Prostaglandins Leukot. Essent. Fat. Acids 2018, 128, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Beynen, A.C.; Hermus, R.J.; Hautvast, J.G. A mathematical relationship between the fatty acid composition of the diet and that of the adipose tissue in man. Am. J. Clin. Nutr. 1980, 33, 81–85. [Google Scholar] [CrossRef] [PubMed]
- London, S.J.; Sacks, F.M.; Caesar, J.; Stampfer, M.J.; Siguel, E.; Willett, W.C. Fatty acid composition of subcutaneous adipose tissue and diet in postmenopausal US women. Am. J. Clin. Nutr. 1991, 54, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Craig, L.C.A.; Thies, F.; Brittenden, J.; Kyle, J.; McNeill, G. Relative validity of fatty acid intakes from an FFQ compared with subcutaneous adipose tissue fatty acids. Proc. Nutr. Soc. 2009, 68, E92. [Google Scholar] [CrossRef]
- Malcom, G.T.; Bhattacharyya, A.K.; Velez-Duran, M.; Guzman, M.A.; Oalmann, M.C.; Strong, J.P. Fatty acid composition of adipose tissue in humans: Differences between subcutaneous sites. Am. J. Clin. Nutr. 1989, 50, 288–291. [Google Scholar] [CrossRef] [PubMed]
- Pezeshkian, M.; Noori, M.; Najjarpour-Jabbari, H.; Abolfathi, A.; Darabi, M.; Darabi, M.; Shaaker, M.; Shahmohammadi, G. Fatty acid composition of epicardial and subcutaneous human adipose tissue. Metab. Syndr. Relat. Disord. 2009, 7, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Petrus, P.; Edholm, D.; Rosqvist, F.; Dahlman, I.; Sundbom, M.; Arner, P.; Rydén, M.; Risérus, U. Depot-specific differences in fatty acid composition and distinct associations with lipogenic gene expression in abdominal adipose tissue of obese women. Int. J. Obes. 2017, 41, 1295–1298. [Google Scholar] [CrossRef] [PubMed]
- Hjelmgaard, K.; Eschen, R.B.; Schmidt, E.B.; Andreasen, J.J.; Lundbye-Christensen, S. Fatty Acid Composition in Various Types of Cardiac Adipose Tissues and Its Relation to the Fatty Acid Content of Atrial Tissue. Nutrients 2018, 10, 1506. [Google Scholar] [CrossRef]
- Baker, A.R.; Silva, N.F.; Quinn, D.W.; Harte, A.L.; Pagano, D.; Bonser, R.S.; Kumar, S.; McTernan, P.G. Human epicardial adipose tissue expresses a pathogenic profile of adipocytokines in patients with cardiovascular disease. Cardiovasc. Diabetol. 2006, 5, 1. [Google Scholar] [CrossRef]
- Tavendale, R.; Lee, A.J.; Smith, W.C.; Tunstall-Pedoe, H. Adipose tissue fatty acids in Scottish men and women: Results from the Scottish Heart Health Study. Atherosclerosis 1992, 94, 161–169. [Google Scholar] [CrossRef]
- Marks, K.A.; Kitson, A.P.; Stark, K.D. Hepatic and plasma sex differences in saturated and monounsaturated fatty acids are associated with differences in expression of elongase 6, but not stearoyl-CoA desaturase in Sprague-Dawley rats. Genes Nutr. 2013, 8, 317–327. [Google Scholar] [CrossRef]
- Romanski, S.A.; Nelson, R.M.; Jensen, M.D. Meal fatty acid uptake in human adipose tissue: Technical and experimental design issues. Am. J. Physiol. Endocrinol. Metab. 2000, 279, E447–E454. [Google Scholar] [CrossRef]
- Palacios-Marin, I.; Serra, D.; Jimenez-Chillarón, J.; Herrero, L.; Todorčević, M. Adipose Tissue Dynamics: Cellular and Lipid Turnover in Health and Disease. Nutrients 2023, 15, 3968. [Google Scholar] [CrossRef]
- Field, C.J.; Angel, A.; Clandinin, M.T. Relationship of diet to the fatty acid composition of human adipose tissue structural and stored lipids. Am. J. Clin. Nutr. 1985, 42, 1206–1220. [Google Scholar] [CrossRef]
- Libby, P.; Buring, J.E.; Badimon, L.; Hansson, G.K.; Deanfield, J.; Bittencourt, M.S.; Tokgözoğlu, L.; Lewis, E.F. Atherosclerosis. Nat. Rev. Dis. Primers 2019, 5, 56. [Google Scholar] [CrossRef]
- Strong, J.P.; Malcom, G.T.; McMahan, C.A.; Tracy, R.E.; Newman, W.P., 3rd; Herderick, E.E.; Cornhill, J.F. Prevalence and extent of atherosclerosis in adolescents and young adults: Implications for prevention from the Pathobiological Determinants of Atherosclerosis in Youth Study. JAMA 1999, 281, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Kurushima, H.; Hayashi, K.; Toyota, Y.; Kambe, M.; Kajiyama, G. Comparison of hypocholesterolemic effects induced by dietary linoleic acid and oleic acid in hamsters. Atherosclerosis 1995, 114, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Thomson, A.M.; Bottema, C.D.; Roach, P.D. Polyunsaturated fatty acids downregulate the low density lipoprotein receptor of human HepG2 cells. J. Nutr. Biochem. 2002, 13, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Cagen, L.M.; Fungwe, T.V.; Wilcox, H.G.; Heimberg, M. Biphasic effect of oleic acid on hepatic cholesterogenesis. Biochem. Biophys. Res. Commun. 1992, 183, 21–26. [Google Scholar] [CrossRef]
- Salam, W.H.; Wilcox, H.G.; Cagen, L.M.; Heimberg, M. Stimulation of hepatic cholesterol biosynthesis by fatty acids. Effects of oleate on cytoplasmic acetoacetyl-CoA thiolase, acetoacetyl-CoA synthetase and hydroxymethylglutaryl-CoA synthase. Biochem. J. 1989, 258, 563–568. [Google Scholar] [CrossRef]
- Yang, Z.H.; Miyahara, H.; Iwasaki, Y.; Takeo, J.; Katayama, M. Dietary supplementation with long-chain monounsaturated fatty acids attenuates obesity-related metabolic dysfunction and increases expression of PPAR gamma in adipose tissue in type 2 diabetic KK-Ay mice. Nutr. Metab. 2013, 10, 16. [Google Scholar] [CrossRef]
- Mensink, R.P.; Katan, M.B. Effect of dietary fatty acids on serum lipids and lipoproteins. A meta-analysis of 27 trials. Arter. Thromb. 1992, 12, 911–919. [Google Scholar]
- Mensink, R.P.; Zock, P.L.; Kester, A.D.; Katan, M.B. Effects of dietary fatty acids and carbohydrates on the ratio of serum total to HDL cholesterol and on serum lipids and apolipoproteins: A meta-analysis of 60 controlled trials. Am. J. Clin. Nutr. 2003, 77, 1146–1155. [Google Scholar] [CrossRef]
- Bonanome, A.; Pagnan, A.; Biffanti, S.; Opportuno, A.; Sorgato, F.; Dorella, M.; Maiorino, M.; Ursini, F. Effect of dietary monounsaturated and polyunsaturated fatty acids on the susceptibility of plasma low density lipoproteins to oxidative modification. Arter. Thromb. 1992, 12, 529–533. [Google Scholar] [CrossRef]
- Yaqoob, P. Monounsaturated fatty acids and immune function. Eur. J. Clin. Nutr. 2002, 56, S9–S13. [Google Scholar] [CrossRef]
- Bermudez, B.; Lopez, S.; Ortega, A.; Varela, L.M.; Pacheco, Y.M.; Abia, R.; Muriana, F.J.G. Oleic acid in olive oil: From a metabolic framework toward a clinical perspective. Curr. Pharm. Des. 2011, 17, 831–843. [Google Scholar] [CrossRef]
- Pérez-Martínez, P.; García-Ríos, A.; Delgado-Lista, J.; Pérez-Jiménez, F.; López-Miranda, J. Mediterranean diet rich in olive oil and obesity, metabolic syndrome and diabetes mellitus. Curr. Pharm. Des. 2011, 17, 769–777. [Google Scholar] [CrossRef]
- Rehman, K.; Haider, K.; Jabeen, K.; Akash, M.S.H. Current perspectives of oleic acid: Regulation of molecular pathways in mitochondrial and endothelial functioning against insulin resistance and diabetes. Rev. Endocr. Metab. Disord. 2020, 21, 631–643. [Google Scholar] [CrossRef]
- Yang, Z.H.; Nill, K.; Takechi-Haraya, Y.; Playford, M.P.; Nguyen, D.; Yu, Z.X.; Pryor, M.; Tang, J.; Rojulpote, K.V.; Mehta, N.N.; et al. Differential Effect of Dietary Supplementation with a Soybean Oil Enriched in Oleic Acid versus Linoleic Acid on Plasma Lipids and Atherosclerosis in LDLR-Deficient Mice. Int. J. Mol. Sci. 2022, 23, 8385. [Google Scholar] [CrossRef]
- Rosqvist, F.; Niinistö, S. Fats and oils—A scoping review for Nordic Nutrition Recommendations 2023. Food Nutr. Res. 2024, 68. [Google Scholar] [CrossRef]
- Revelou, P.K.; Xagoraris, M.; Alexandropoulou, A.; Kanakis, C.D.; Papadopoulos, G.K.; Pappas, C.S.; Tarantilis, P.A. Chemometric Study of Fatty Acid Composition of Virgin Olive Oil from Four Widespread Greek Cultivars. Molecules 2021, 26, 4151. [Google Scholar] [CrossRef]
- Cao, H.; Gerhold, K.; Mayers, J.R.; Wiest, M.M.; Watkins, S.M.; Hotamisligil, G.S. Identification of a lipokine, a lipid hormone linking adipose tissue to systemic metabolism. Cell 2008, 134, 933–944. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.H.; Pryor, M.; Noguchi, A.; Sampson, M.; Johnson, B.; Pryor, M.; Donkor, K.; Amar, M.; Remaley, A.T. Dietary Palmitoleic Acid Attenuates Atherosclerosis Progression and Hyperlipidemia in Low-Density Lipoprotein Receptor-Deficient Mice. Mol. Nutr. Food Res. 2019, 63, e1900120. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Li, H.; Xu, H.; Halim, V.; Zhang, W.; Wang, H.; Ong, K.T.; Woo, S.L.; Walzem, R.L.; Mashek, D.G.; et al. Palmitoleate induces hepatic steatosis but suppresses liver inflammatory response in mice. PLoS ONE 2012, 7, e39286. [Google Scholar] [CrossRef] [PubMed]
- Souza, C.O.; Teixeira, A.A.; Biondo, L.A.; Silveira, L.S.; Calder, P.C.; Rosa Neto, J.C. Palmitoleic acid reduces the inflammation in LPS-stimulated macrophages by inhibition of NFκB, independently of PPARs. Clin. Exp. Pharmacol. Physiol. 2017, 44, 566–575. [Google Scholar] [CrossRef] [PubMed]
- de Souza, C.O.; Vannice, G.K.; Rosa Neto, J.C.; Calder, P.C. Is Palmitoleic Acid a Plausible Nonpharmacological Strategy to Prevent or Control Chronic Metabolic and Inflammatory Disorders? Mol. Nutr. Food Res. 2018, 62, 1700504. [Google Scholar] [CrossRef] [PubMed]
- de Souza, C.O.; Valenzuela, C.A.; Baker, E.J.; Miles, E.A.; Rosa Neto, J.C.; Calder, P.C. Palmitoleic Acid has Stronger Anti-Inflammatory Potential in Human Endothelial Cells Compared to Oleic and Palmitic Acids. Mol. Nutr. Food Res. 2018, 62, e1800322. [Google Scholar] [CrossRef] [PubMed]
- Curb, J.D.; Wergowske, G.; Dobbs, J.C.; Abbott, R.D.; Huang, B. Serum lipid effects of a high-monounsaturated fat diet based on macadamia nuts. Arch. Intern. Med. 2000, 160, 1154–1158. [Google Scholar] [CrossRef]
- Garg, M.L.; Blake, R.J.; Wills, R.B. Macadamia nut consumption lowers plasma total and LDL cholesterol levels in hypercholesterolemic men. J. Nutr. 2003, 133, 1060–1063. [Google Scholar] [CrossRef]
- Stefan, N.; Kantartzis, K.; Celebi, N.; Staiger, H.; Machann, J.; Schick, F.; Cegan, A.; Elcnerova, M.; Schleicher, E.; Fritsche, A.; et al. Circulating palmitoleate strongly and independently predicts insulin sensitivity in humans. Diabetes Care 2010, 33, 405–407. [Google Scholar] [CrossRef]
- Wang, L.; Folsom, A.R.; Zheng, Z.J.; Pankow, J.S.; Eckfeldt, J.H. Plasma fatty acid composition and incidence of diabetes in middle-aged adults: The Atherosclerosis Risk in Communities (ARIC) Study. Am. J. Clin. Nutr. 2003, 78, 91–98. [Google Scholar] [CrossRef]
- Cetin, E.; Pedersen, B.; Porter, L.M.; Adler, G.K.; Burak, M.F. Protocol for a randomized placebo-controlled clinical trial using pure palmitoleic acid to ameliorate insulin resistance and lipogenesis in overweight and obese subjects with prediabetes. Front. Endocrinol. 2023, 14, 1306528. [Google Scholar] [CrossRef]
- Djoussé, L.; Matthan, N.R.; Lichtenstein, A.H.; Gaziano, J.M. Red blood cell membrane concentration of cis-palmitoleic and cis-vaccenic acids and risk of coronary heart disease. Am. J. Cardiol. 2012, 110, 539–544. [Google Scholar] [CrossRef]
- Wang, Y.; Jacome-Sosa, M.M.; Vine, D.F.; Proctor, S.D. Beneficial effects of vaccenic acid on postprandial lipid metabolism and dyslipidemia: Impact of natural trans-fats to improve CVD risk. Lipid Technol. 2010, 22, 103–106. [Google Scholar] [CrossRef]
- Gebauer, S.K.; Destaillats, F.; Dionisi, F.; Krauss, R.M.; Baer, D.J. Vaccenic acid and trans fatty acid isomers from partially hydrogenated oil both adversely affect LDL cholesterol: A double-blind, randomized controlled trial. Am. J. Clin. Nutr. 2015, 102, 1339–1346. [Google Scholar] [CrossRef]
- Abbasi, A.; Mostafaie, A.; Bahrami, G.; Mansouri, K.; Sisakhtnejad, S. The Effect of Cis and Trans Vaccenic Acids on Expression of ICAM-1 And VCAM-1 in Human Microvascular Endothelial Cells (HMEC). J. Rep. Pharm. Sci. 2015, 4, 65–74. [Google Scholar] [CrossRef]
- Valenzuela, C.A.; Baker, E.J.; De Souza, C.O.; Miles, E.A.; Calder, P.C. Differential Effects of Ruminant and Industrial 18-Carbon trans-Monounsaturated Fatty Acids (trans Vaccenic and Elaidic) on the Inflammatory Responses of an Endothelial Cell Line. Molecules 2021, 26, 5834. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.; Brandt, E. Association Between Dietary Fat Content and Serum Lipoprotein(a) Level. J. Clin. Lipidol. 2022, 16, e62. [Google Scholar] [CrossRef]
- Wulff, A.B.; Nordestgaard, B.G. Residual cardiovascular risk beyond low-density lipoprotein cholesterol: Inflammation, remnant cholesterol, and lipoprotein(a). Eur. Heart J. 2025, ehaf274. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.E.; Vedel-Krogh, S.; Kamstrup, P.R.; Nordestgaard, B.G. Lipoprotein(a) Cardiovascular Risk Explained by LDL Cholesterol, Non-HDL Cholesterol, ApoB, or hsCRP Is Minimal. J. Am. Coll Cardiol. 2025, 85, 2046–2051. [Google Scholar] [CrossRef]
- Yang, Z.H.; Gordon, S.M.; Sviridov, D.; Wang, S.; Danner, R.L.; Pryor, M.; Vaisman, B.; Shichijo, Y.; Doisaki, N.; Remaley, A.T. Dietary supplementation with long-chain monounsaturated fatty acid isomers decreases atherosclerosis and alters lipoprotein proteomes in LDLr(-/-) mice. Atherosclerosis 2017, 262, 31–38. [Google Scholar] [CrossRef]
- Yang, Z.H.; Amar, M.; Sorokin, A.V.; Troendle, J.; Courville, A.B.; Sampson, M.; Playford, M.P.; Yang, S.; Stagliano, M.; Ling, C.; et al. Supplementation with saury oil, a fish oil high in omega-11 monounsaturated fatty acids, improves plasma lipids in healthy subjects. J. Clin. Lipidol. 2020, 14, 53–65. [Google Scholar] [CrossRef]
- Takahashi, A.; Ishizaki, M.; Kimira, Y.; Egashira, Y.; Hirai, S. Erucic Acid-Rich Yellow Mustard Oil Improves Insulin Resistance in KK-A(y) Mice. Molecules 2021, 26, 546. [Google Scholar] [CrossRef]
- Rimmen, E.V.; Mjøs, S.A.; Søfteland, E.; Gudbrandsen, O.A. Intake of herring oil, but not of anchovy oil, resulted in a lower serum cholesterol concentration in male Zucker Diabetic Sprague Dawley rats. Br. J. Nutr. 2024, 132, 547–557. [Google Scholar] [CrossRef]
- Mjaatveit, M.; Oldernes, H.; Gudbrandsen, O.A. Effects of diets containing fish oils or fish oil concentrates with high cetoleic acid content on the circulating cholesterol concentration in rodents. A systematic review and meta-analysis. Br. J. Nutr. 2024, 131, 606–621. [Google Scholar] [CrossRef]
- Zong, G.; Li, Y.; Sampson, L.; Dougherty, L.W.; Willett, W.C.; Wanders, A.J.; Alssema, M.; Zock, P.L.; Hu, F.B.; Sun, Q. Monounsaturated fats from plant and animal sources in relation to risk of coronary heart disease among US men and women. Am. J. Clin. Nutr. 2018, 107, 445–453. [Google Scholar] [CrossRef]
- Lawlor, D.A.; Ebrahim, S.; Davey Smith, G. Socioeconomic position in childhood and adulthood and insulin resistance: Cross sectional survey using data from British women’s heart and health study. BMJ 2002, 325, 805. [Google Scholar] [CrossRef]
- Lennon, L.T.; Ramsay, S.E.; Papacosta, O.; Shaper, A.G.; Wannamethee, S.G.; Whincup, P.H. Cohort Profile Update: The British Regional Heart Study 1978–2014: 35 years follow-up of cardiovascular disease and ageing. Int. J. Epidemiol. 2015, 44, 826–826g. [Google Scholar] [CrossRef]
- Marmot, M.; Brunner, E. Cohort Profile: The Whitehall II study. Int. J. Epidemiol. 2005, 34, 251–256. [Google Scholar] [CrossRef]
- Tillin, T.; Forouhi, N.G.; McKeigue, P.M.; Chaturvedi, N. Southall And Brent REvisited: Cohort profile of SABRE, a UK population-based comparison of cardiovascular disease and diabetes in people of European, Indian Asian and African Caribbean origins. Int. J. Epidemiol. 2012, 41, 33–42. [Google Scholar] [CrossRef]
- Bainton, D.; Miller, N.E.; Bolton, C.H.; Yarnell, J.W.; Sweetnam, P.M.; Baker, I.A.; Lewis, B.; Elwood, P.C. Plasma triglyceride and high density lipoprotein cholesterol as predictors of ischaemic heart disease in British men. The Caerphilly and Speedwell Collaborative Heart Disease Studies. Br. Heart J. 1992, 68, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Menon, U.; Gentry-Maharaj, A.; Ryan, A.; Sharma, A.; Burnell, M.; Hallett, R.; Lewis, S.; Lopez, A.; Godfrey, K.; Oram, D.; et al. Recruitment to multicentre trials--lessons from UKCTOCS: Descriptive study. BMJ 2008, 337, a2079. [Google Scholar] [CrossRef] [PubMed]
- Vartiainen, E.; Laatikainen, T.; Peltonen, M.; Juolevi, A.; Männistö, S.; Sundvall, J.; Jousilahti, P.; Salomaa, V.; Valsta, L.; Puska, P. Thirty-five-year trends in cardiovascular risk factors in Finland. Int. J. Epidemiol. 2010, 39, 504–518. [Google Scholar] [CrossRef] [PubMed]
- Baylin, A.; Kabagambe, E.K.; Siles, X.; Campos, H. Adipose tissue biomarkers of fatty acid intake123. Am. J. Clin. Nutr. 2002, 76, 750–757. [Google Scholar] [CrossRef]
- Luan, D.; Wang, D.; Campos, H.; Baylin, A. Adipose tissue palmitoleic acid is inversely associated with nonfatal acute myocardial infarction in Costa Rican adults. Nutr. Metab. Cardiovasc. Dis. 2018, 28, 973–979. [Google Scholar] [CrossRef]
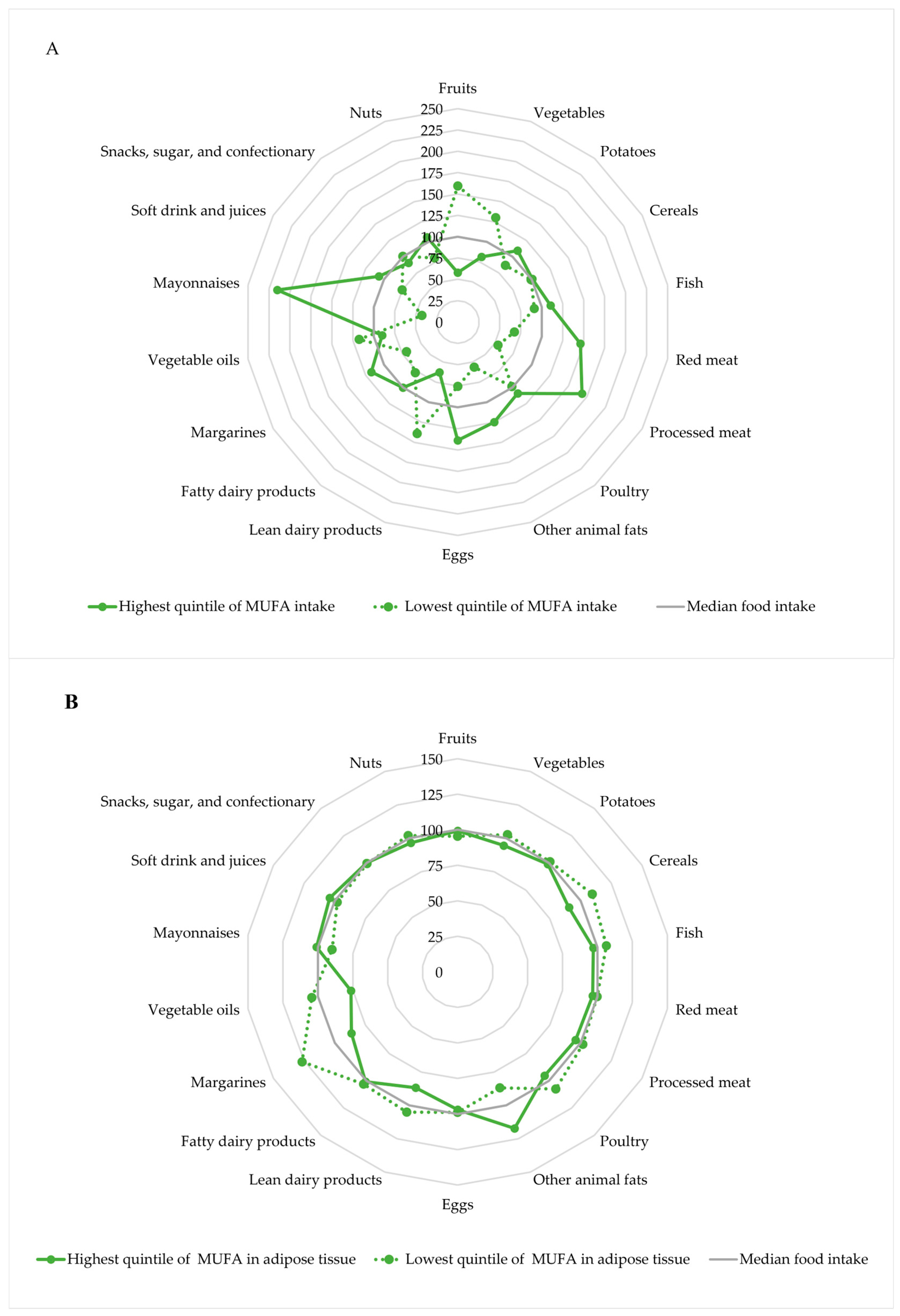

| Dietary Source | MUFA Intake in g/d |
|---|---|
| Fatty dairy products | 5.3 (1.2, 14.1) |
| Red Meat | 4.8 (1.9, 10.2) |
| Margarines | 2.8 (0.3, 9.1) |
| Processed meat | 2.4 (0.5, 7.8) |
| Lean dairy products | 1.2 (0.2, 3.2) |
| Eggs | 1.1 (0.3, 3.3) |
| Vegetable oils | 1.1 (0.5, 7.1) |
| Fish | 1.0 (0.2, 3.3) |
| Mayonnaise and similar products | 0.8 (0.1, 6.4) |
| Snacks | 0.7 (0.1, 3.7) |
| Poultry | 0.6 (0.1, 1.7) |
| Nuts | 0.2 (0.0, 1.0) |
| Vegetables | 0.1 (0.0, 0.5) |
| Fruit | 0.1 (0.0, 0.2) |
| Other animal fat | 0.1 (0.0, 0.4) |
| MUFA | Daily Intake of MUFA (% of Total Fat Intake) | Adipose Tissue Content of MUFA (% of Total Fatty Acids) |
|---|---|---|
| Total MUFA | 27.46 (15.02, 49.28) | 54.22 (48.84, 59.35) |
| Oleic acid (18:1ω-9) | 23.02 (12.30, 42.49) | 44.02 (40.67, 47.15) |
| Vaccenic acid (18:1ω-7) | 1.41 (0.19, 5.93) | 1.99 (1.54, 2.77) |
| Palmitoleic acid (16:1ω-7) | 1.39 (0.76, 2.47) | 6.55 (4.15, 9.49) |
| Myristoleic acid (14:1ω-5) | 0.37 (0.16, 0.71) | 0.42 (0.24, 0.66) |
| Gadoleic (20:1ω-9) and Gondoic acid (20:1ω-11) | 0.72 (0.34, 1.37) | 0.99 (0.72, 1.41) |
| Erucic acid (22:1ω-9) | 0.01 (0.00, 0.05) | 0.05 (0.02, 0.10) |
| Absolute Dietary Intake of MUFAs | Dietary Intake of MUFAs as Proportion of Total Dietary FA Intake | |
|---|---|---|
| Total MUFAs | −0.08 | 0.16 |
| Oleic acid (18:1ω-9) | 0.04 | 0.22 |
| Vaccenic acid (18:1ω-7) | −0.09 | −0.06 |
| Palmitoleic acid (16:1ω-7) | −0.10 | 0.14 |
| Myristoleic acid (14:1ω-5) | 0.14 | 0.43 |
| Gadoleic (20:1ω-9) and Gondoic acid (20:1ω-11) | 0.26 | 0.34 |
| Erucic acid (22:1ω-9) | 0.10 | 0.10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pedersen, J.; Hedegaard, B.S.; Schmidt, E.B.; Dahm, C.C.; Holven, K.B.; Retterstøl, K.; Calder, P.C.; Bork, C. Monounsaturated Fatty Acids in Cardiovascular Disease: Intake, Individual Types, and Content in Adipose Tissue as a Biomarker of Endogenous Exposure. Nutrients 2025, 17, 2509. https://doi.org/10.3390/nu17152509
Pedersen J, Hedegaard BS, Schmidt EB, Dahm CC, Holven KB, Retterstøl K, Calder PC, Bork C. Monounsaturated Fatty Acids in Cardiovascular Disease: Intake, Individual Types, and Content in Adipose Tissue as a Biomarker of Endogenous Exposure. Nutrients. 2025; 17(15):2509. https://doi.org/10.3390/nu17152509
Chicago/Turabian StylePedersen, Jonas, Berit Storgaard Hedegaard, Erik Berg Schmidt, Christina C. Dahm, Kirsten B. Holven, Kjetil Retterstøl, Philip C. Calder, and Christian Bork. 2025. "Monounsaturated Fatty Acids in Cardiovascular Disease: Intake, Individual Types, and Content in Adipose Tissue as a Biomarker of Endogenous Exposure" Nutrients 17, no. 15: 2509. https://doi.org/10.3390/nu17152509
APA StylePedersen, J., Hedegaard, B. S., Schmidt, E. B., Dahm, C. C., Holven, K. B., Retterstøl, K., Calder, P. C., & Bork, C. (2025). Monounsaturated Fatty Acids in Cardiovascular Disease: Intake, Individual Types, and Content in Adipose Tissue as a Biomarker of Endogenous Exposure. Nutrients, 17(15), 2509. https://doi.org/10.3390/nu17152509