
Vegetable Extracts as Therapeutic Agents: A Comprehensive Exploration of Anti-Allergic Effects
Abstract
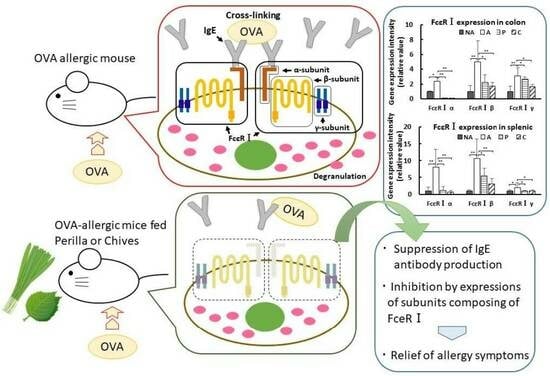
1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Determination of Total Polyphenols
2.3. Determination of Antioxidant Capacity
2.4. Laccase Treatment of Vegetable Extracts
2.5. Cell Model
2.6. Evaluation of Cell Viability of Vegetable Extracts on RBL-2H3 Cells
2.7. Degranulation Test via Hexosaminidase Release Examination
2.8. Intracellular Calcium Ion Evaluation
2.9. Animals and Experimental Diets
2.10. Study Design
2.11. Preparation of Fecal Samples
2.12. Determination of Antibody Concentrations in Blood and Feces
2.12.1. Fecal IgA
2.12.2. Total IgE in the Blood
2.12.3. Measurement of OVA-Specific IgE Antibody Titers in the Blood
2.13. RT-PCR
2.14. Statistical Analysis
3. Results
3.1. Total Polyphenol Content and Antioxidant Capacity of the 22 Vegetable Extracts
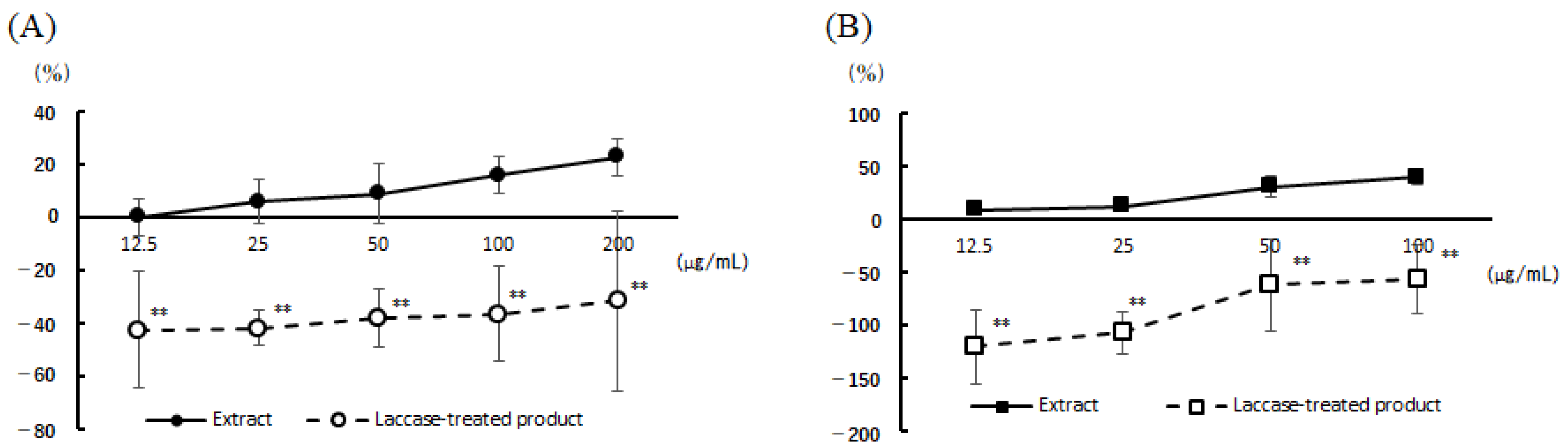
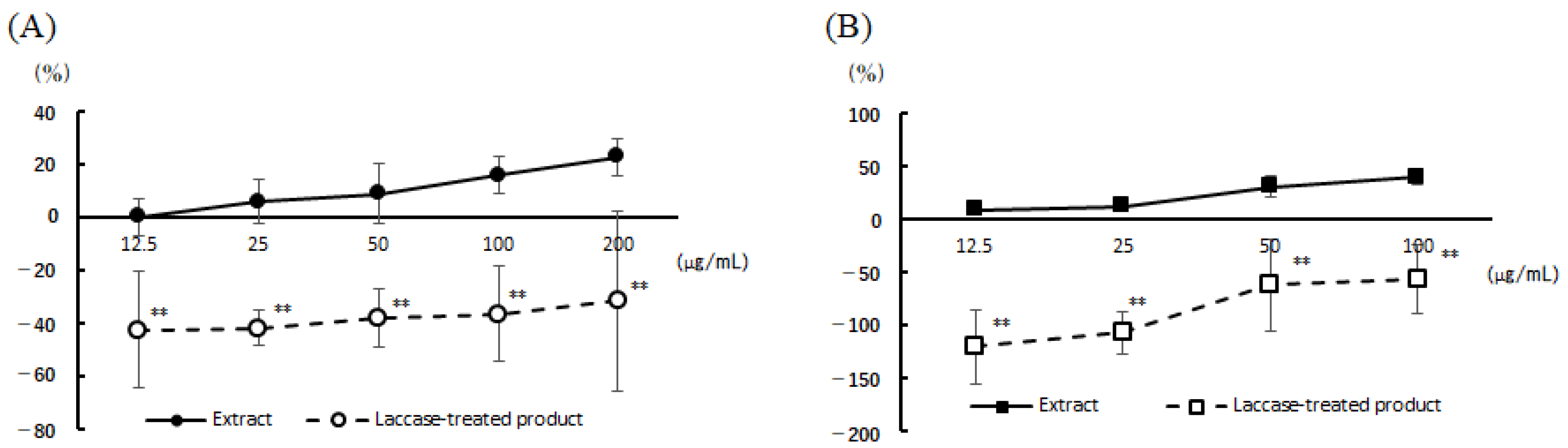
3.2. Cell Viability and Degranulation Inhibition Activity of the 22 Vegetable Extracts
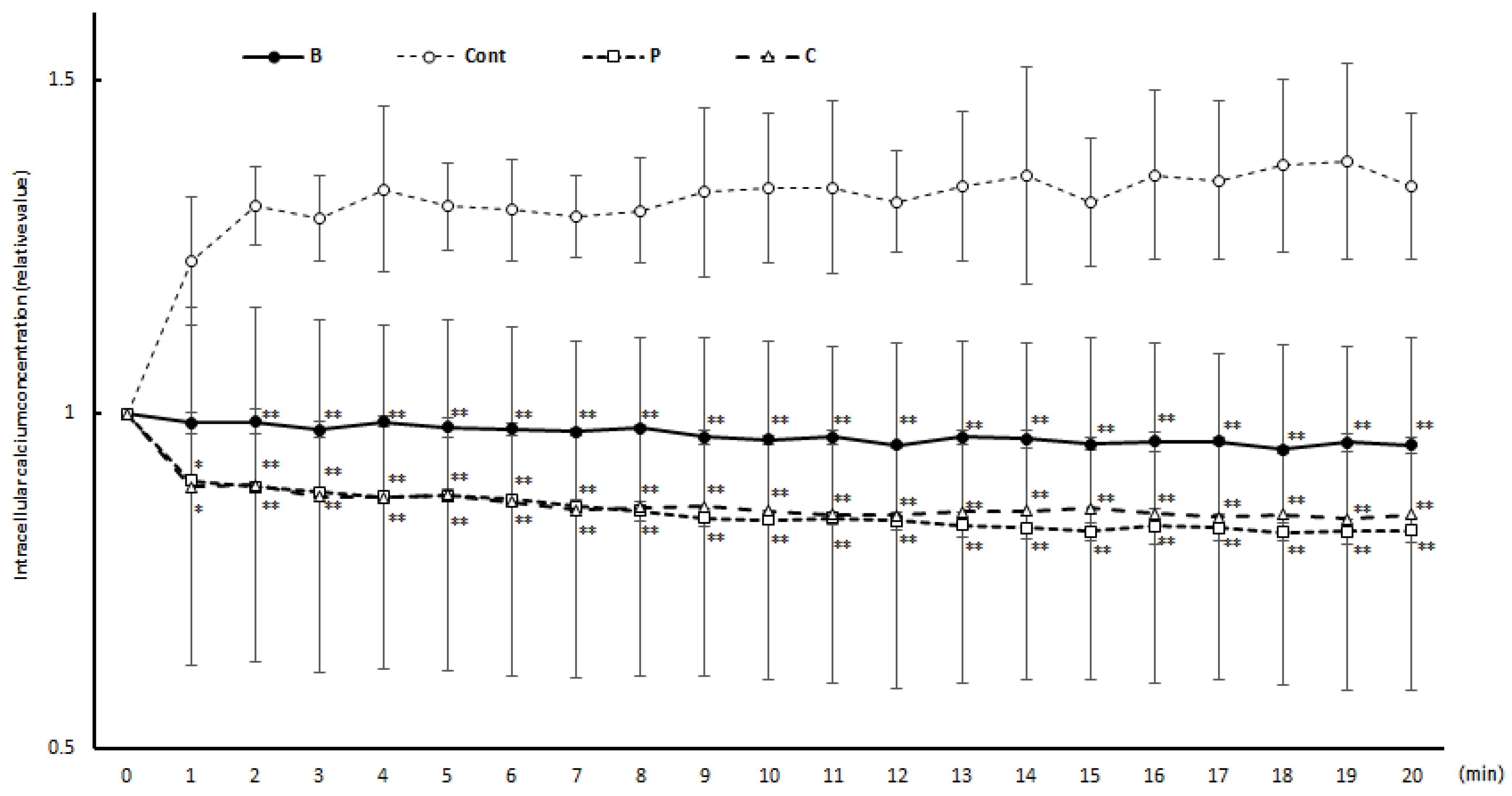
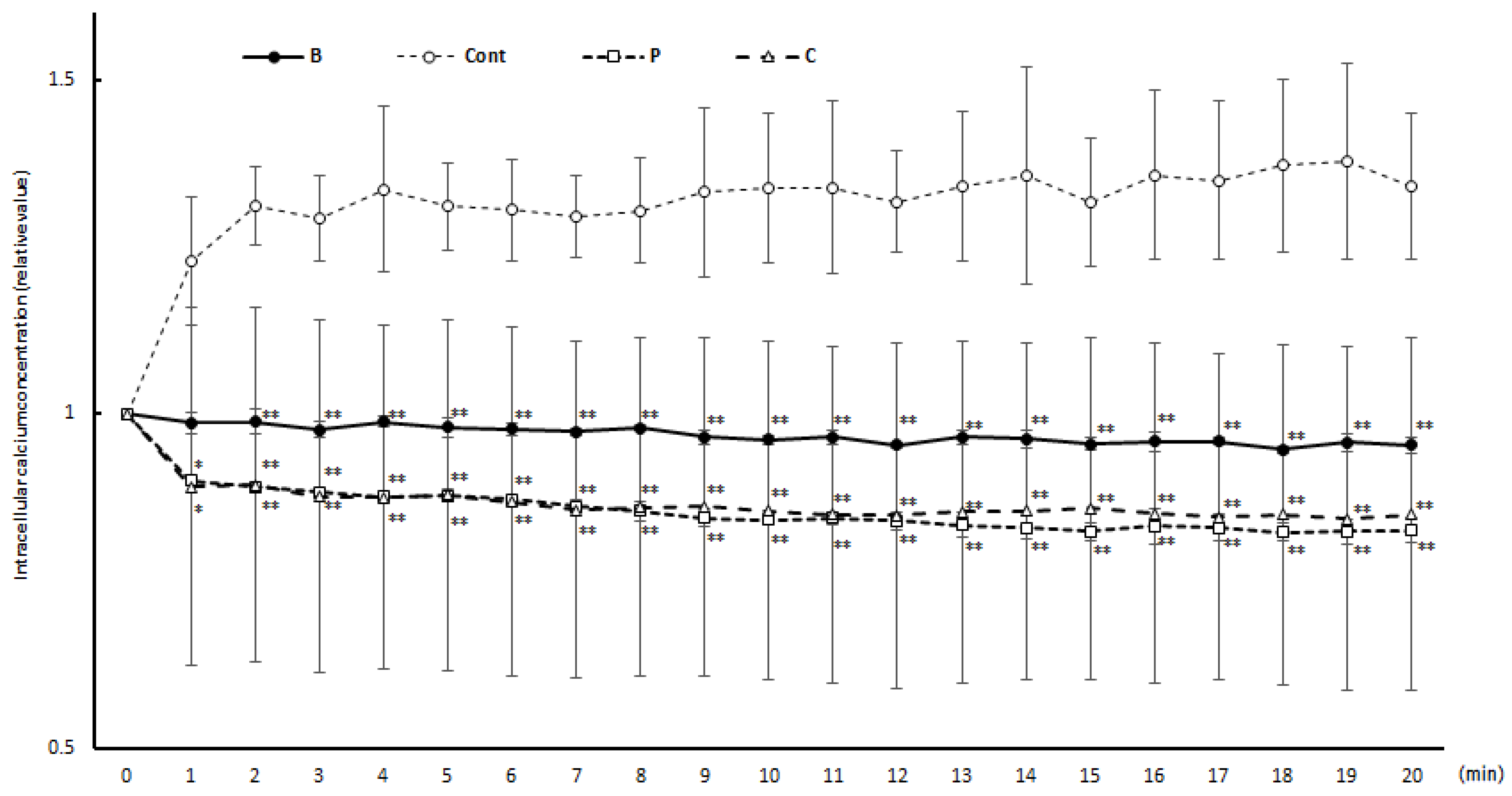
3.3. Intracellular Calcium Concentration Changes during Degranulation Reaction
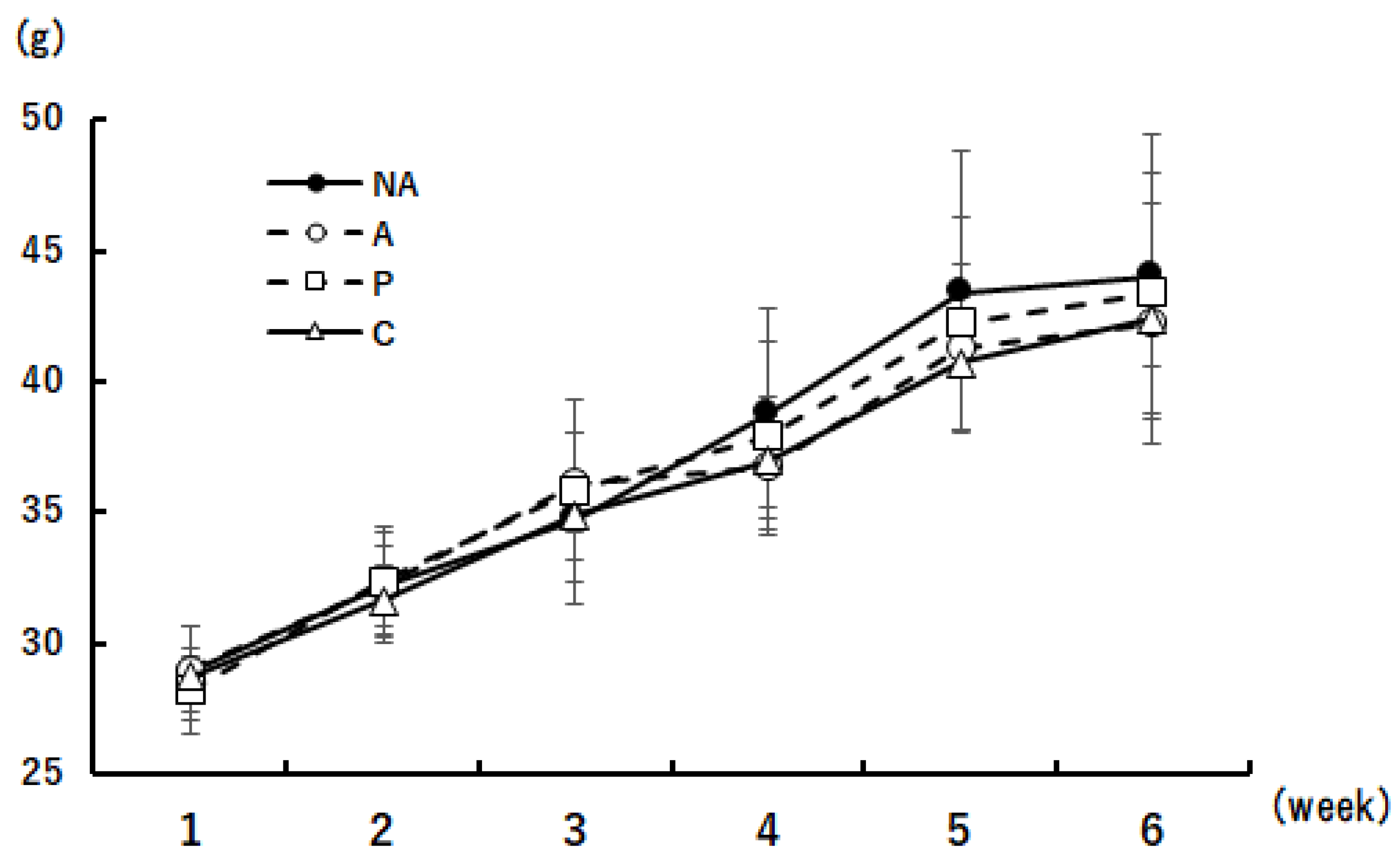
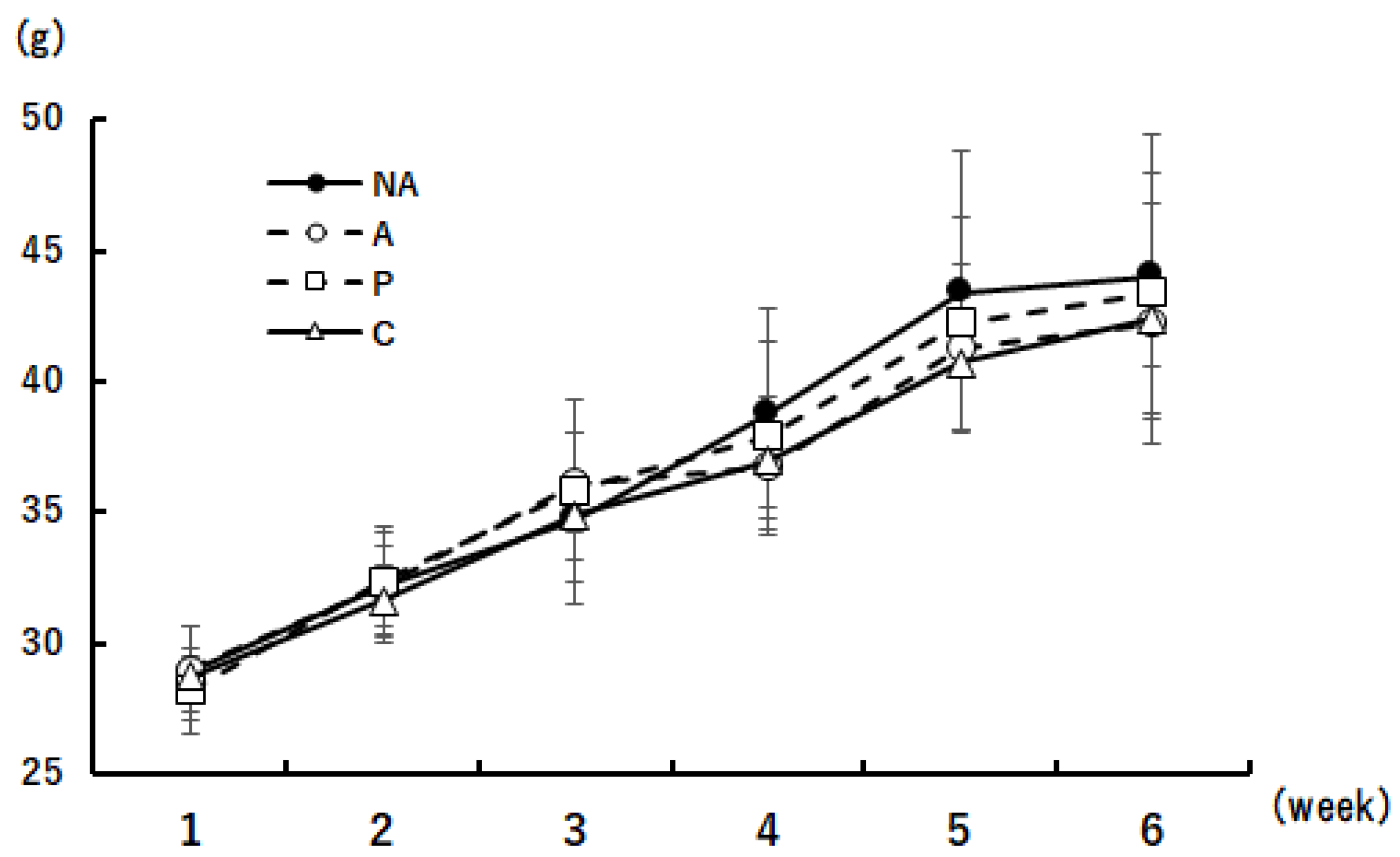
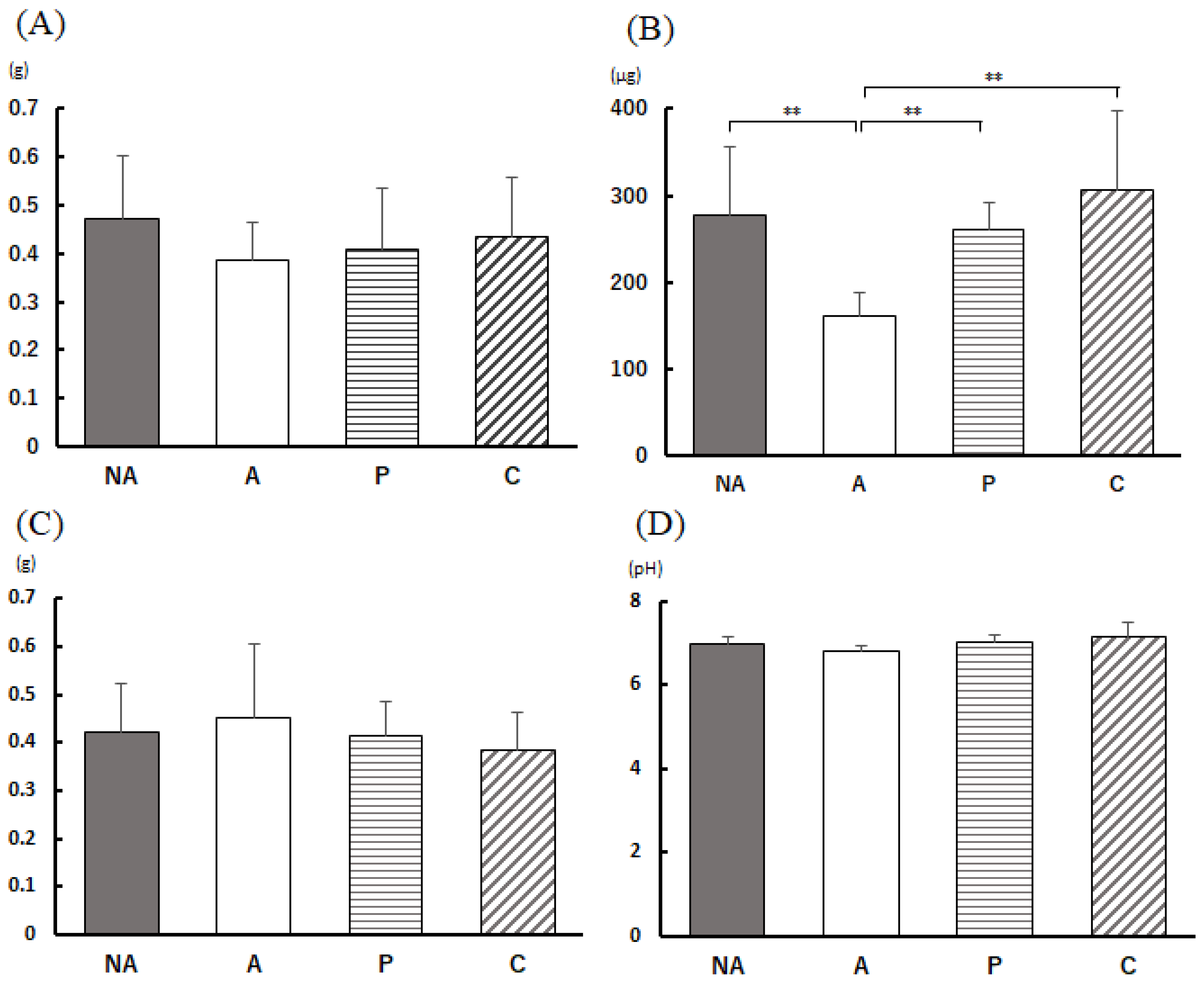
3.4. Weight Change in Mice during the Rearing Period
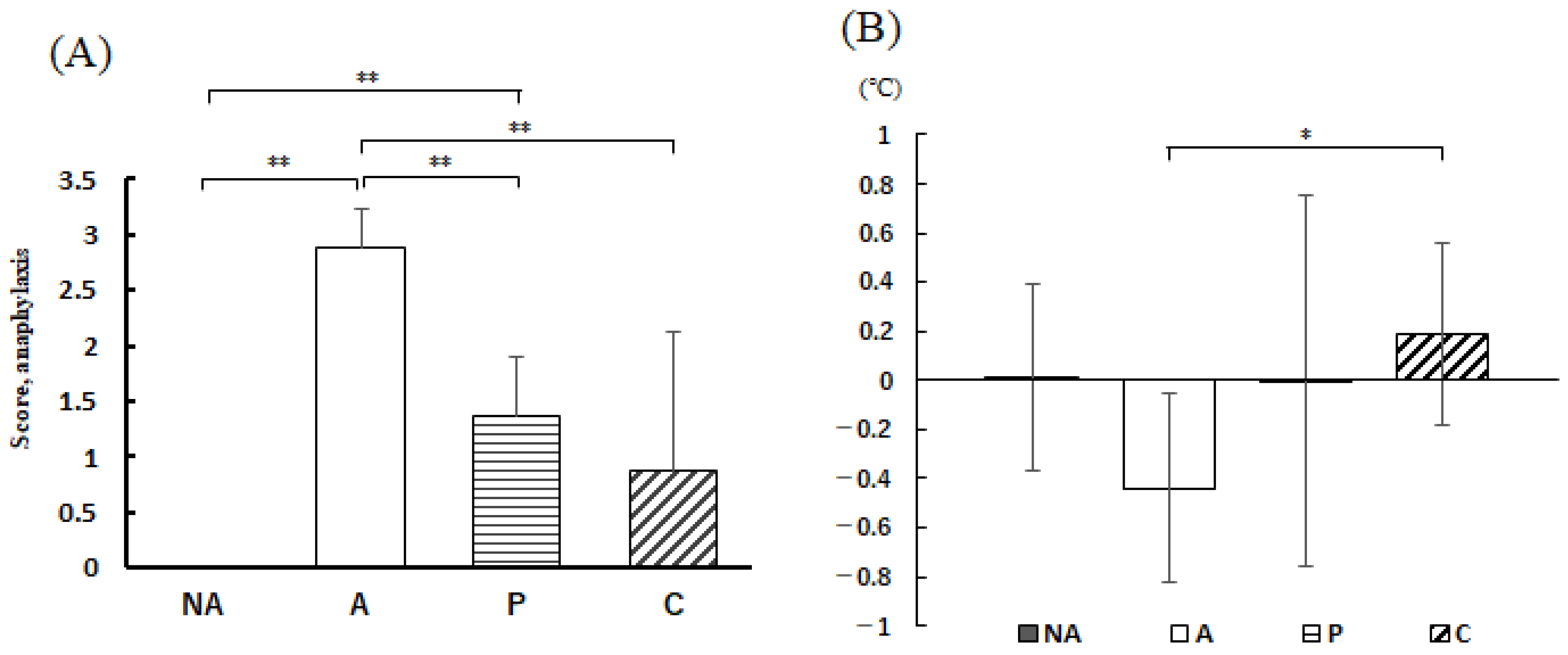
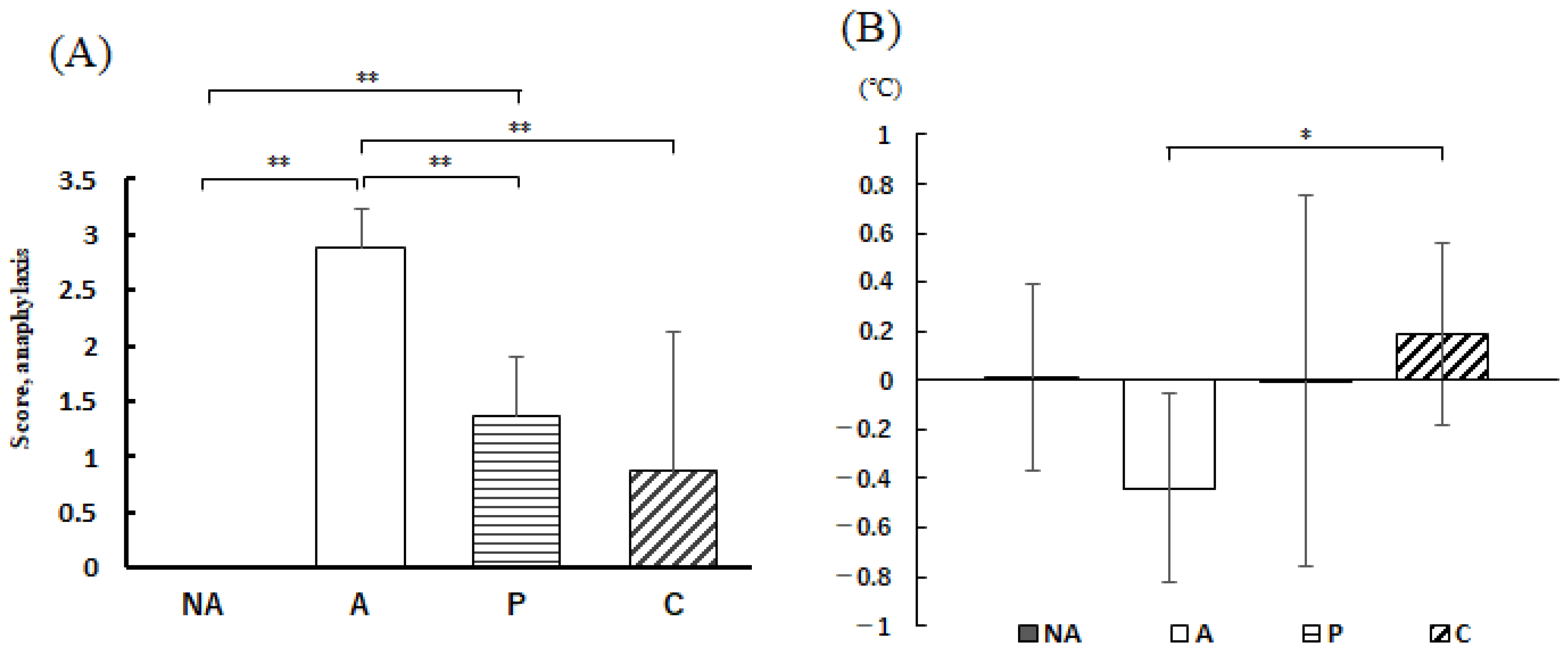
3.5. Anaphylactic Symptoms
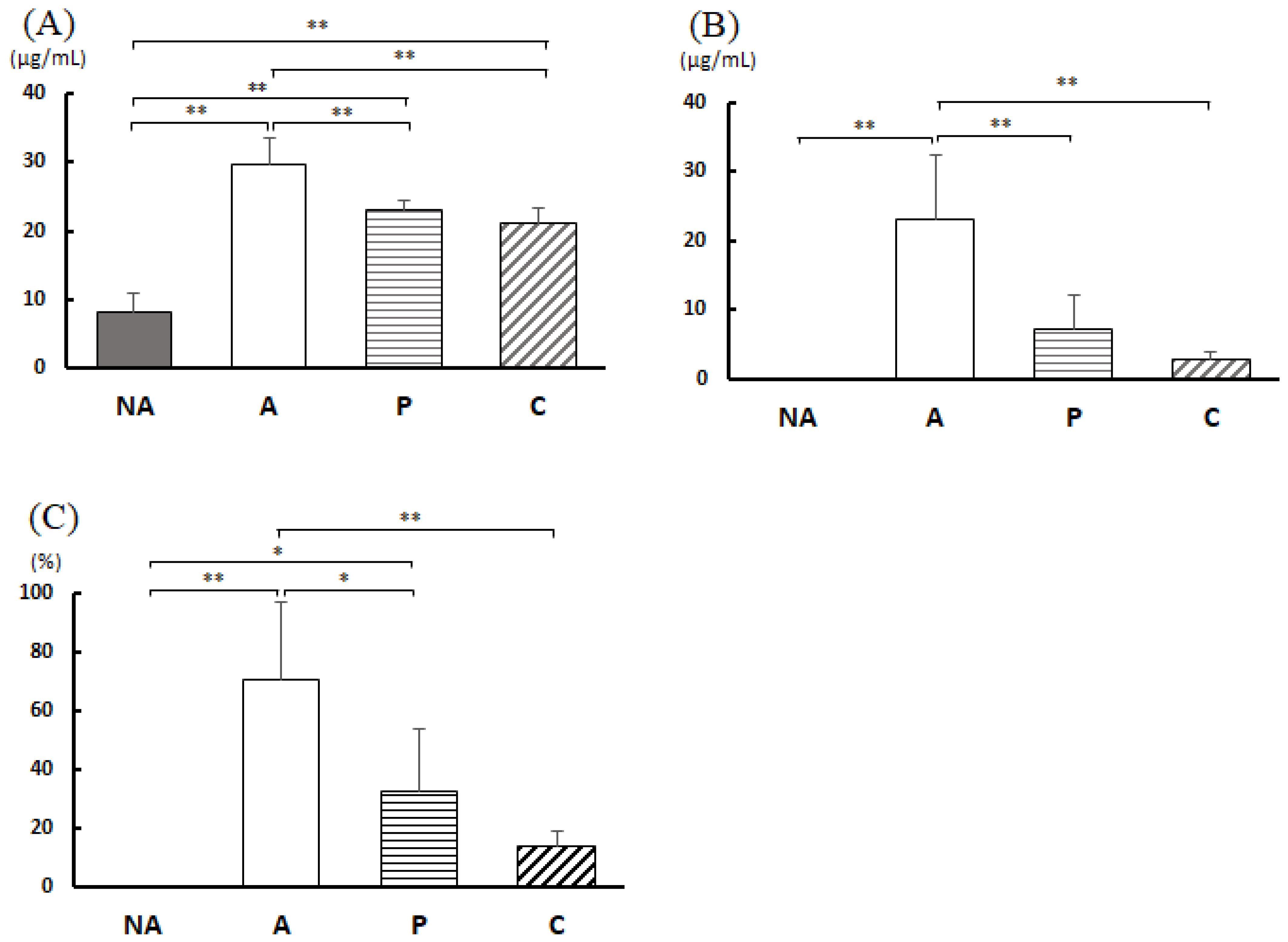
3.6. Analysis of Feces
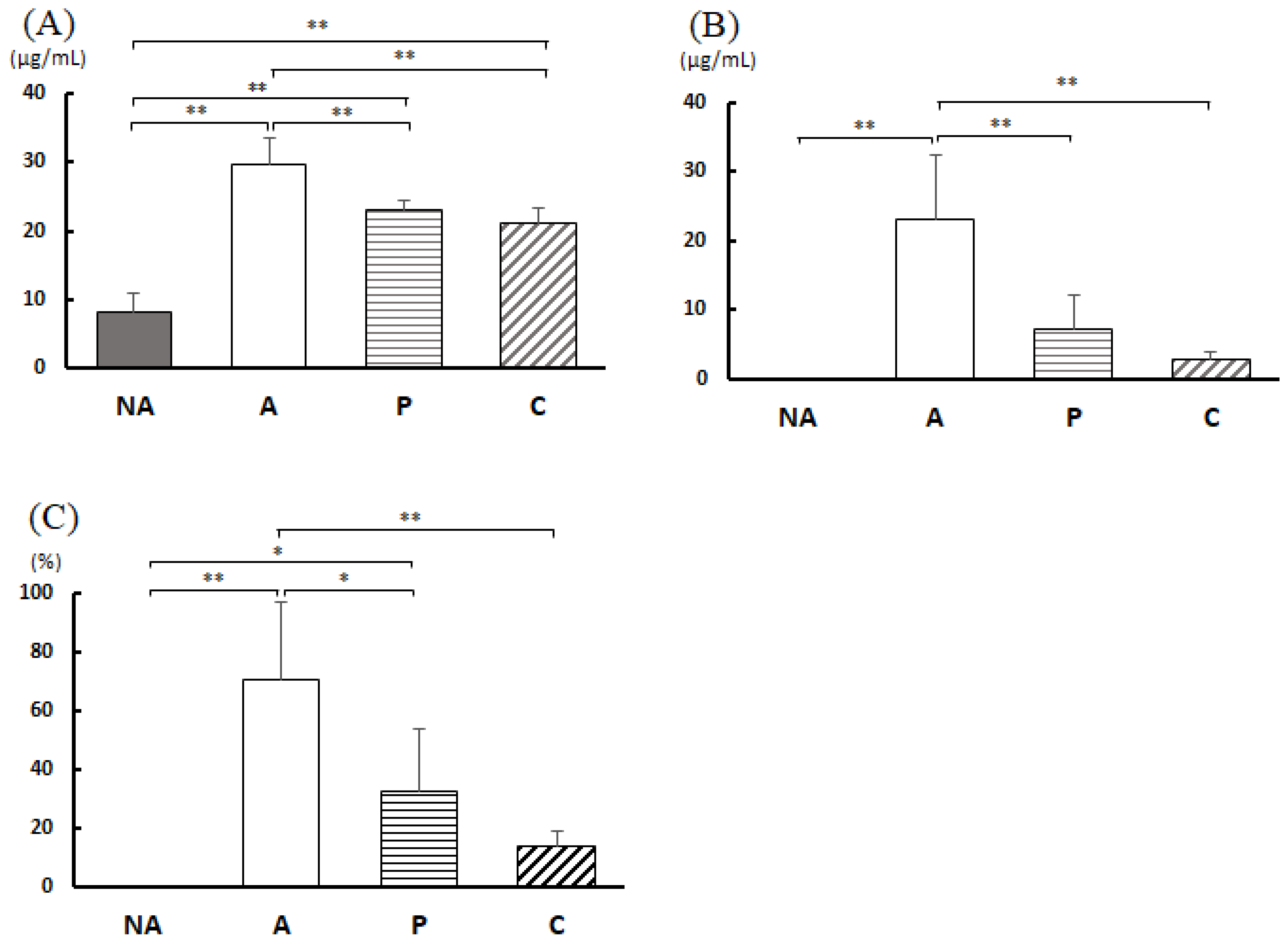
3.7. Blood Antibody Titer
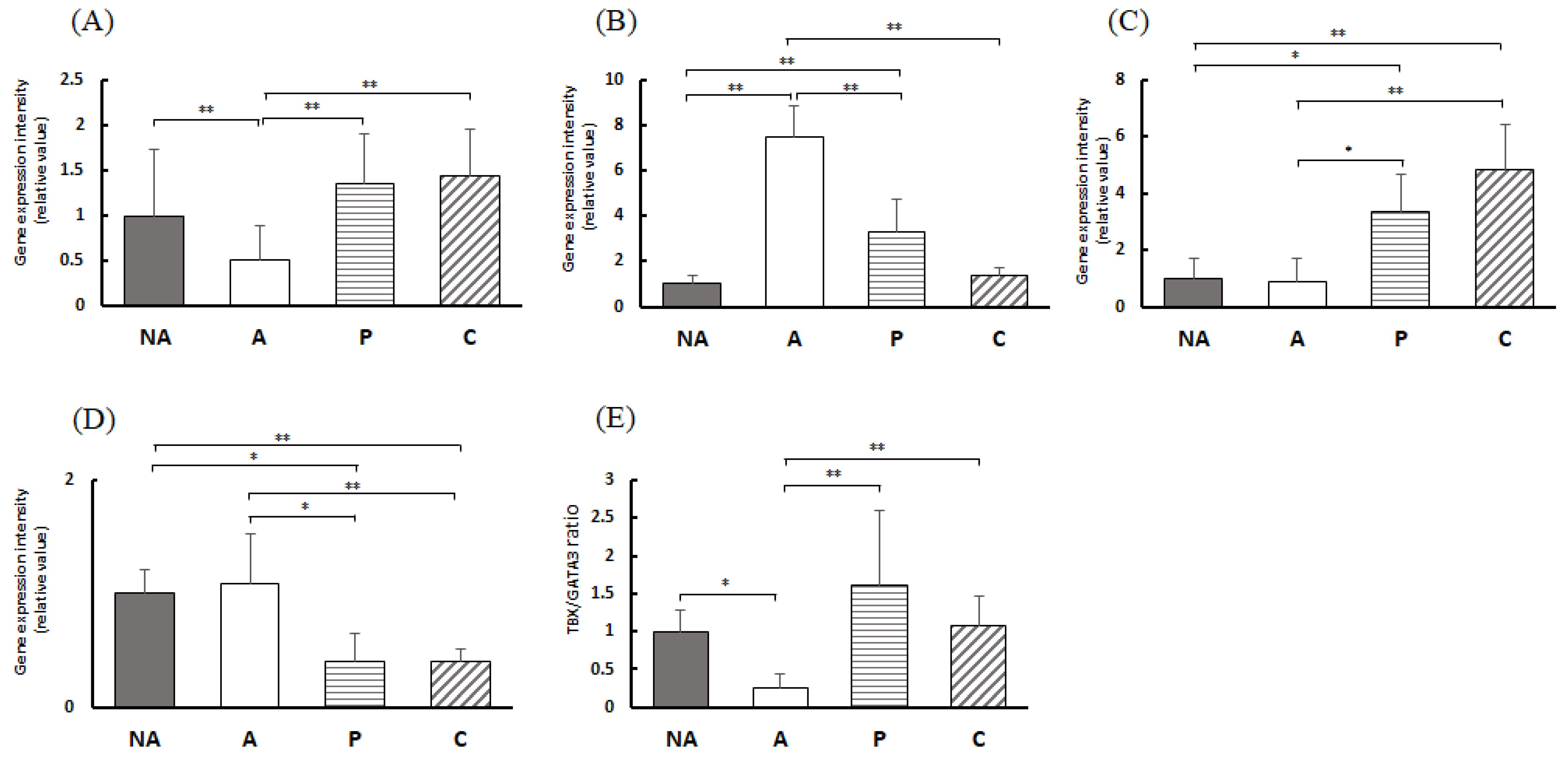
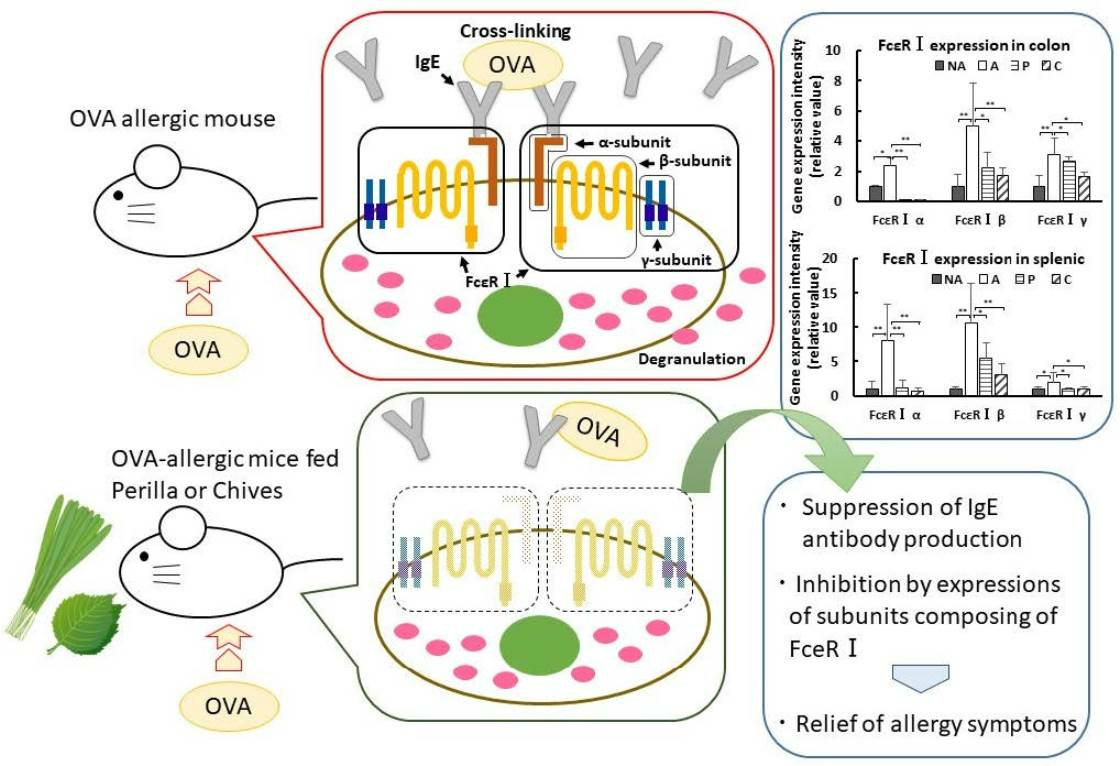
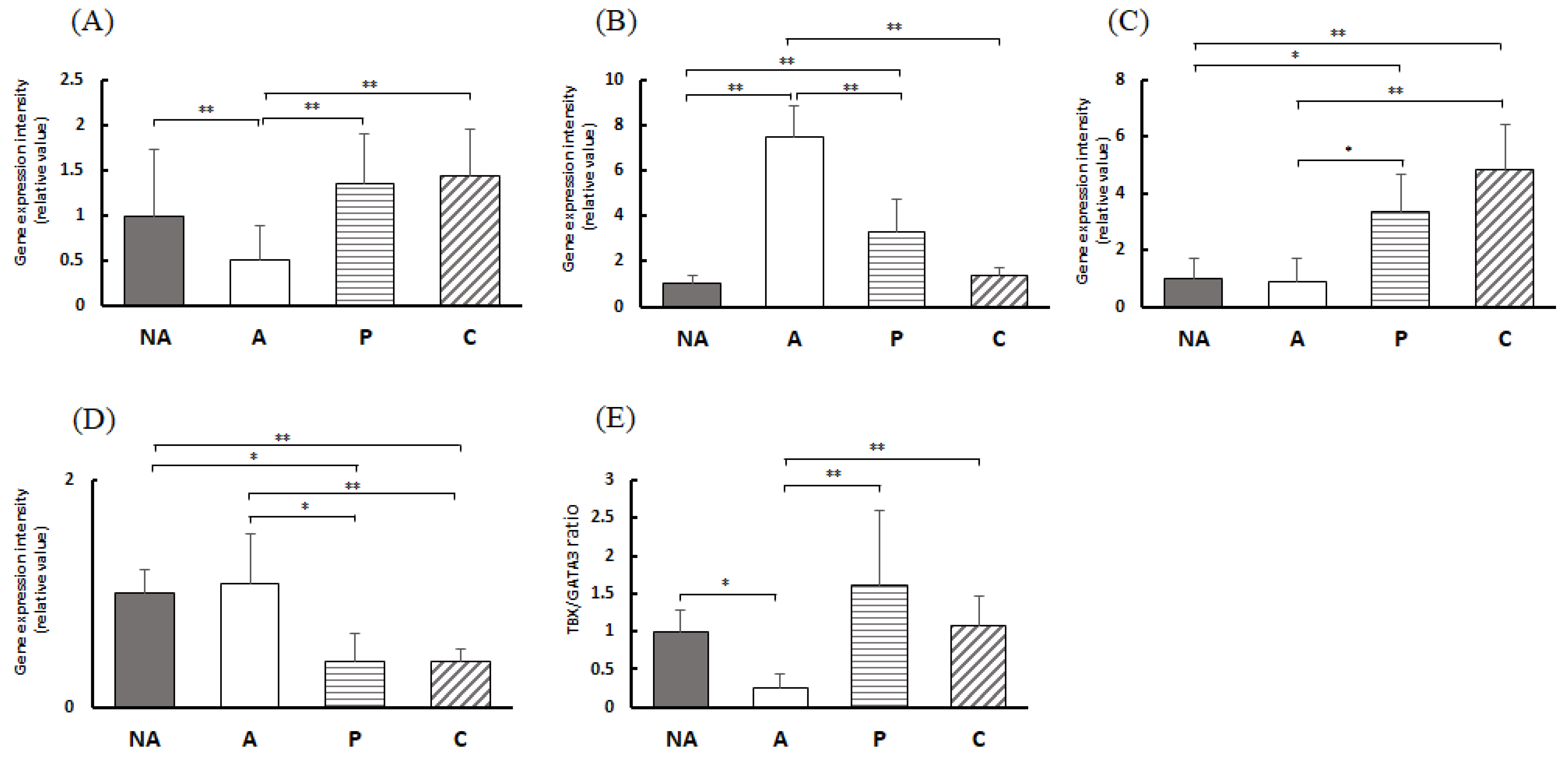
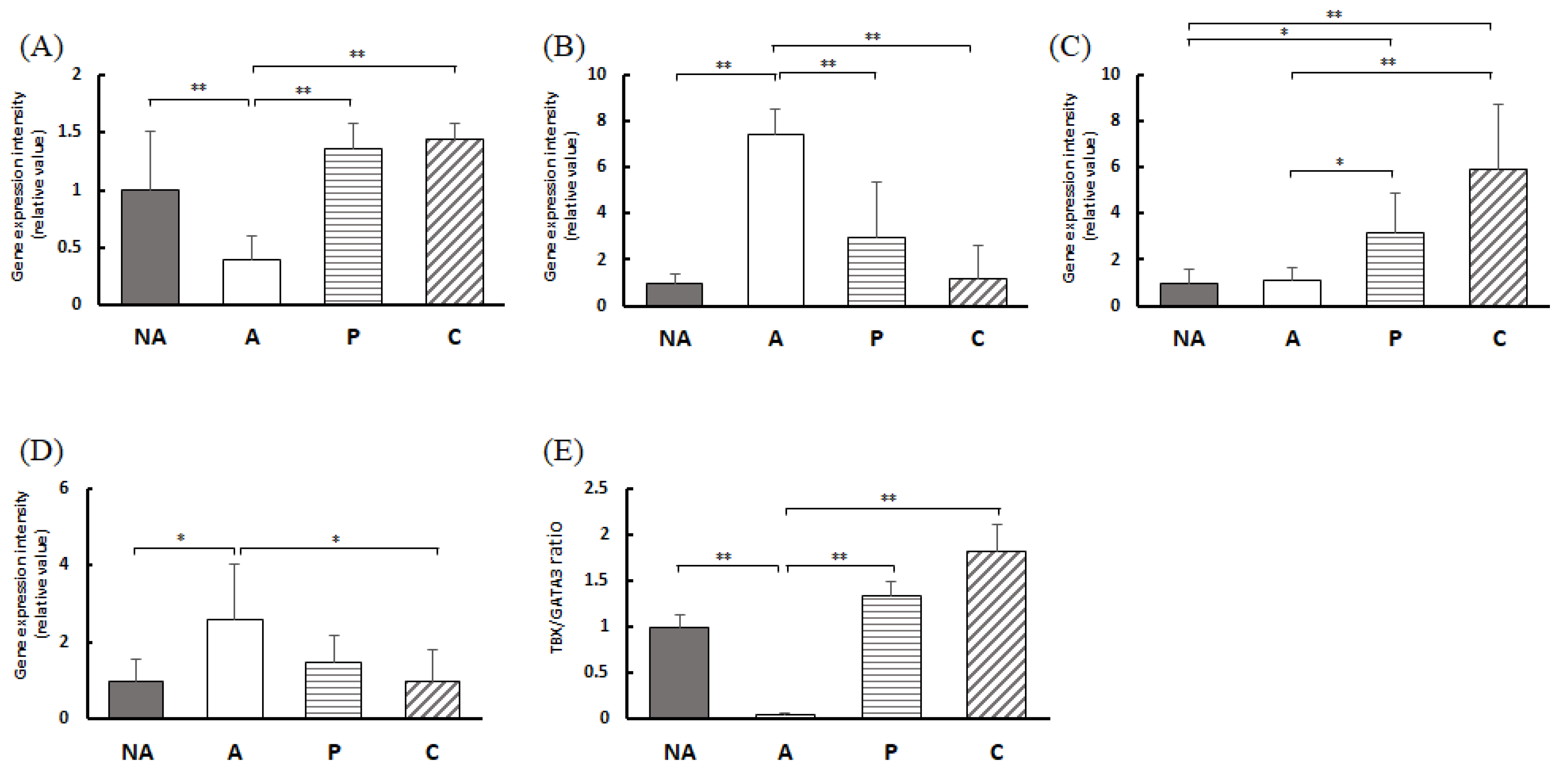
3.8. Gene Expression in the Colon
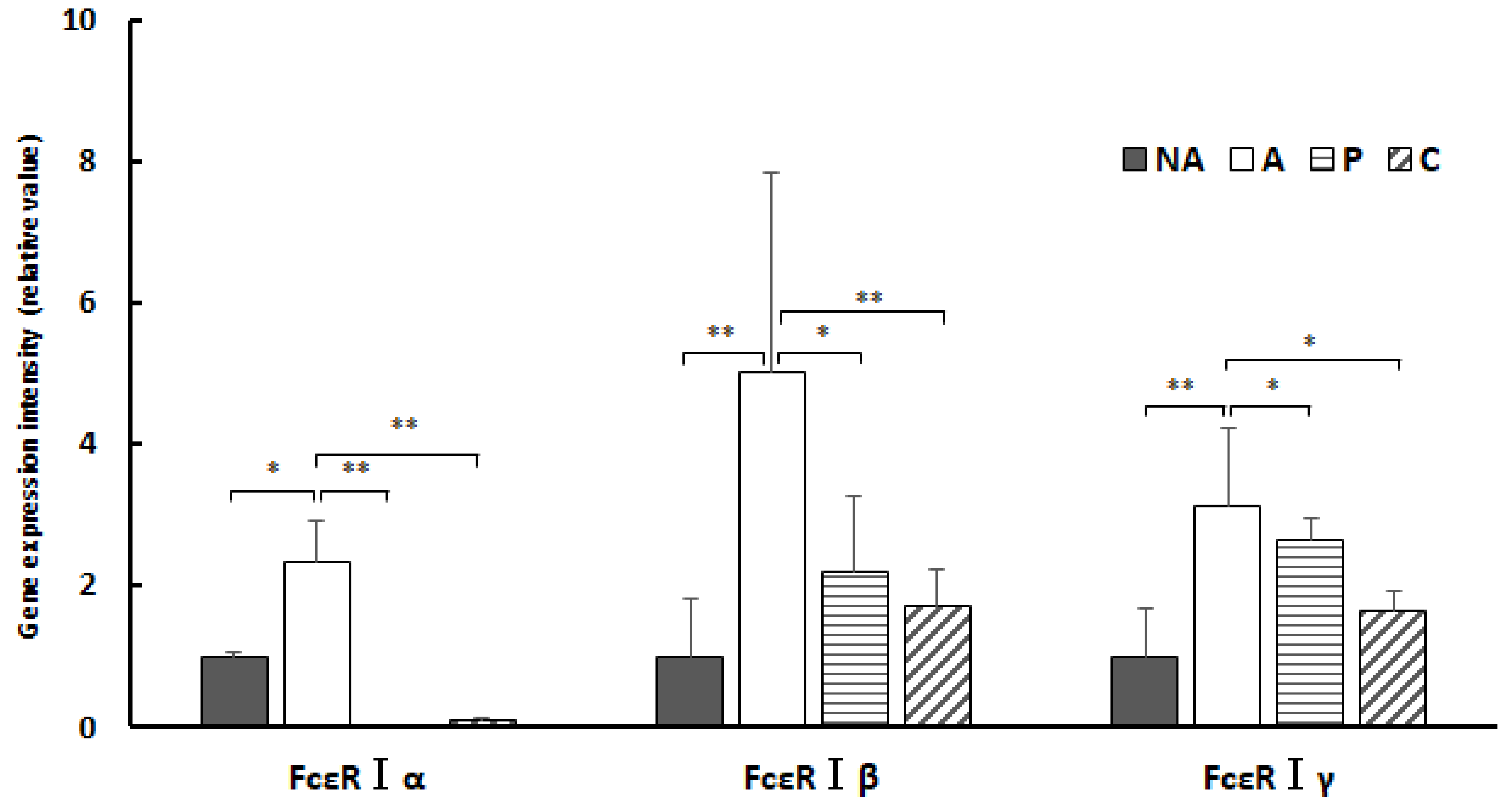
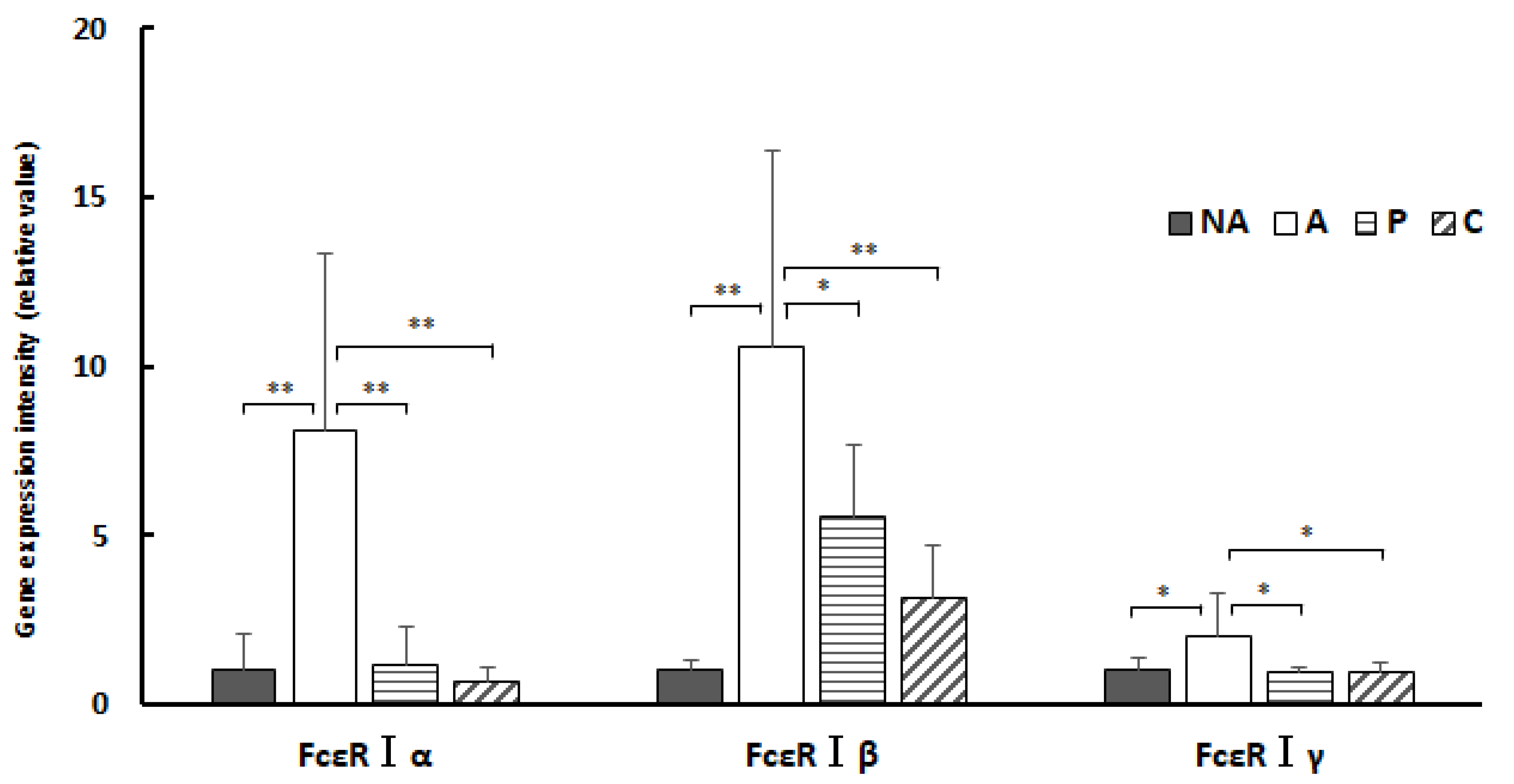
3.9. Gene Expression in the Spleen
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Martinis, M.; Sirufo, M.M.; Suppa, M.; Ginaldi, L. New Perspectives in Food Allergy. Int. J. Mol. Sci. 2020, 21, 1474. [Google Scholar] [CrossRef] [PubMed]
- Comberiati, P.; Costagliola, C.; D’Elios, S.; Peroni, D. Prevention of Food Allergy: The Significance of Early Introduction. Medicina 2019, 55, 323. [Google Scholar] [CrossRef]
- Gargano, D.; Appanna, R.; Santonicola, A.; De Bartolomeis, F.; Stellato, C.; Cianferoni, A.; Casolaro, V.; Iovino, P. Food Allergy and Intolerance: A Narrative Review on Nutritional Concerns. Nutrients 2021, 13, 1638. [Google Scholar] [CrossRef]
- Lyons, S.A.; Burney, P.G.J.; Ballmer-Weber, B.K.; Fernandez-Rivas, M.; Barreales, L.; Clausen, M.; Dubakiene, R.; Fernandez-Perez, C.; Fritsche, P.; Jedrzejczak-Czechowicz, M.; et al. Food Allergy in Adults: Substantial Variation in Prevalence and Causative Foods Across Europe. J. Allergy Clin. Immunol. Pract. 2019, 7, 1920–1928. [Google Scholar] [CrossRef] [PubMed]
- Freye, H.B.; Esch, R.E.; Litwin, C.M.; Sorkin, L. Anaphylaxis to the ingestion and inhalation of Tenebrio molitor (mealworm) and Zophobas morio (superworm). Allergy Asthma Proc. 1996, 17, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Gautreau, M.; Restuccia, M.; Senser, K.; Weisberg, S.N. Familial Anaphylaxis after Silkworm Ingestion. Prehosp. Emerg. Care 2017, 21, 83–85. [Google Scholar] [CrossRef]
- Pasquale, T.D.; Buonomo, A.; Illuminati, I.; D’Alò, S.; Pucci, S. Recurrent Anaphylaxis: A Case of IgE-Mediated Allergy to Carmine Red (E120). J. Investig Allergol. Clin. Immunol. 2015, 25, 440–441. [Google Scholar]
- De Gier, S.d.; Verhoeckx, K. Insect (food) allergy and allergens. Mol. Immunol. 2018, 100, 82–106. [Google Scholar] [CrossRef]
- Shimoda, H.; Tanaka, J.; Yamada, E.; Morikawa, T.; Kasajima, N.; Yoshikawa, M. Anti type I allergic property of Japanese butterbur extract and its mast cell degranulation inhibitory ingredients. J. Agric. Food Chem. 2006, 54, 2915–2920. [Google Scholar] [CrossRef]
- Duthie, G.G.; Gardner, P.T.; Kyle, J.A.M. Plant polyphenols: Are they the new magic bullet? Proc. Nutr. Soc. 2003, 62, 599–603. [Google Scholar] [CrossRef]
- Bramati, L.; Aquilano, F.; Pietta, P. Unfermented rooibos tea: Quantitative characterization of flavonoids by HPLC-UV and determination of the total antioxidant activity. J. Agric. Food Chem. 2003, 51, 7472–7474. [Google Scholar] [CrossRef]
- Singh, A.; Holvoet, S.; Mercenier, A. Dietary polyphenols in the prevention and treatment of allergic diseases. Clin. Exp. Allergy 2011, 41, 1346–1359. [Google Scholar] [CrossRef]
- Nishibe, S.; Yamaguchi, S.; Hasegawa, M.; Oba, K.; Fujikawa, T. Anti-obesity effect of Forsythia leaf extract containing polyphenolic compounds and its mechanism. J. Trad. Med. 2012, 29, 149–155. [Google Scholar]
- Lee, S.H.; Kim, B.J.; Choi, H.J.; Cho, S.W.; Shin, C.S.; Park, S.Y.; Lee, Y.S.; Lee, S.Y.; Kim, H.H.; Kim, G.S.; et al. (-)-Epigallocathechin-3-gallate, an AMPK activator, decreases ovariectomy-induced bone loss by suppression of bone resorption. Calcif. Tissue Int. 2012, 90, 404–410. [Google Scholar] [CrossRef]
- Savaskan, E.; Olivieri, G.; Meier, F.; Seifritz, E.; Wirz-Justice, A.; Müller-Spahn, F. Red wine ingredient resveratrol protects from beta-amyloid neurotoxicity. Gerontology 2003, 49, 380–383. [Google Scholar] [CrossRef] [PubMed]
- Shaik, Y.; Caraffa, A.; Ronconi, G.; Lessiani, G.; Conti, P. Impact of polyphenols on mast cells with special emphasis on the effect of quercetin and luteolin. Cent. Eur. J. Immunol. 2018, 43, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Granroth, B. Biosynthesis and decomposition of cysteine derivatives in onion and other AIlium species. Biosynth. Decompos. Cysteine Deriv. Onion Other AIlium Species 1970, 154, 77. [Google Scholar]
- Oki, T.; Masuda, M.; Kobayashi, M.; Nishiba, Y.; Furuta, S.; Suda, I.; Sato, T. Polymeric procyanidins as radical-scavenging components in red-hulled rice. J. Agric. Food Chem. 2002, 50, 7524–7529. [Google Scholar] [CrossRef] [PubMed]
- Li, X.M.; Schofield, B.H.; Huang, C.K.; Kleiner, G.I.; Sampson, H.A. A murine model of IgE-mediated cow’s milk hypersensitivity. J. Allergy Clin. Immunol. 1999, 103, 206–214. [Google Scholar] [CrossRef]
- Janusz, G.; Pawlik, A.; Świderska-Burek, U.; Polak, J.; Sulej, J.; Jarosz-Wilkołazka, A.; Paszczyński, A. Laccase Properties, Physiological Functions, and Evolution. Int. J. Mol. Sci. 2020, 21, 966. [Google Scholar] [CrossRef] [PubMed]
- Mira, O.; Kim, S.Y.; Park, S.; Kim, K.N.; Kim, S.H. Phytochemicals in Chinese chive (Allium tuberosum) induce the skeletal muscle cell proliferation via PI3K/Akt/mTOR and smad pathways in C2C12 Cells. Int. J. Mol. Sci. 2021, 22, 2296. [Google Scholar]
- Asif, M. Phytochemical study of polyphenols in Perilla Frutescens as an antioxidant. Avicenna. J. Phytomed. 2012, 2, 169–178. [Google Scholar]
- Makabe-Kobayashi, Y.; Hori, Y.; Adachi, T.; Ishigaki-Suzuki, S.; Kikuchi, Y.; Kagaya, Y.; Shirato, K.; Nagy, A.; Ujike, A.; Takai, T.; et al. The control effect of histamine on body temperature and respiratory function in IgE-dependent systemic anaphylaxis. J. Allergy Clin. Immunol. 2002, 110, 298–303. [Google Scholar] [CrossRef]
- France, M.M.; Turner, J.R. The mucosal barrier at a glance. J. Cell Sci. 2017, 130, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.; Pamp, S.J.; Hill, J.A.; Surana, N.A.; Edelman, S.M.; Troy, E.B.; Reading, N.C.; Villablanca, E.J.; Wang, S.; Mora, J.R.; et al. Gut immune maturation depends on colonization with a host-specific microbiota. Cell 2012, 149, 1578–1593. [Google Scholar] [CrossRef] [PubMed]
- Corthésy, B. Multi-faceted functions of secretory IgA at mucosal surfaces. Front. Immunol. 2013, 4, 185. [Google Scholar] [CrossRef] [PubMed]
- Strait, R.T.; Mahler, A.; Hogan, S.; Khodoun, M.; Shibuya, A.; Finkelman, F.D. Ingested allergens must be absorbed systemically to induce systemic anaphylaxis. J. Allergy Clin. Immunol. 2011, 127, 982–989. [Google Scholar] [CrossRef]
- El Ansari, Y.S.; Kanagaratham, C.; Burton, O.T.; Santos, J.V.; Hollister, B.A.; Lewis, O.L.; Renz, H.; Oettgen, H.C. Allergen-Specific IgA Antibodies Block IgE-Mediated Activation of Mast Cells and Basophils. Front. Immunol. 2022, 13, 881655. [Google Scholar] [CrossRef]
- Zhu, X.L.; Zhu, J.F. CD4 T Helper Cell Subsets and Related Human Immunological Disorders. Int. J. Mol. Sci. 2020, 21, 8011–8036. [Google Scholar] [CrossRef]
- Sekiya, T.; Kashiwagi, I.; Yoshida, R.; Fukaya, T.; Morita, R.; Kimura, A.; Ichinose, H.; Metzger, D.; Chambon, P.; Yoshimura, A. Nr4a receptors are essential for thymic regulatory T cell development and immune homeostasis. Nat. Immunol. 2013, 14, 230–237. [Google Scholar] [CrossRef]
- Turcanu, V.; Maleki, S.J.; Lack, G. Characterization of lymphocyte responses to peanuts in normal children, peanut-allergic children, and allergic children who acquired tolerance to peanuts. J. Clin. Investig. 2003, 111, 1065–1072. [Google Scholar] [CrossRef]
- Gap Ryol Lee, G.R. The Balance of Th17 versus Treg Cells in Autoimmunity. Int. J. Mol. Sci. 2018, 19, 730. [Google Scholar]
- Vultaggio, A.; Virgili, G.; Gaeta, F.; Romano, A.; Maggi, E.; Matucci, A. High serum β-lactams specific/total IgE ratio is associated with immediate reactions to β-lactams antibiotics. PLoS ONE 2015, 10, e0121857. [Google Scholar] [CrossRef]
- Yuka Nagata, Y.; Suzuki, R. FcεRI: A Master Regulator of Mast Cell Functions. Cells 2022, 11, 622. [Google Scholar] [CrossRef]
- Li, Y.; Leung, P.S.C.; Gershwin, M.E.; Song, J. New Mechanistic Advances in FcεRI-Mast Cell-Mediated Allergic Signaling. Clin. Rev. Allergy Immunol. 2022, 63, 431–446. [Google Scholar] [CrossRef] [PubMed]
- Sutton, B.J.; Davies, A.M. Structure and dynamics of IgE-receptor interactions: FcεRI and CD23/FcεRII. Immunol Rev. 2015, 268, 222–235. [Google Scholar] [CrossRef] [PubMed]
- Kinet, J.P. The high-affinity IgE receptor (Fc epsilon RI): From physiology to pathology. Annu. Rev. Immunol. 1999, 17, 931–972. [Google Scholar] [CrossRef]
- Park, Y.H.; Kim, H.W.; Kim, H.S.; Nam, S.T.; Lee, D.; Lee, M.B.; Min, K.Y.; Koo, J.; Kim, S.J.; Kim, Y.M.; et al. An Anti-Cancer Drug Candidate CYC116 Suppresses Type I Hypersensitive Immune Responses through the Inhibition of Fyn Kinase in Mast Cells. Biomol. Ther. 2019, 27, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Lecce, M.; Molfetta, R.; Nadia Domenica Milito, N.D.; Santoni, A.; Paolini, R. FcεRI Signaling in the Modulation of Allergic Response: Role of Mast Cell-Derived Exosomes. Int. J. Mol. Sci. 2020, 21, 5464. [Google Scholar] [CrossRef]
- Dispenza, M.C.; Bochner, B.S.; MacGlashan, D.W., Jr. Targeting the FcεRI Pathway as a Potential Strategy to Prevent Food-Induced Anaphylaxis. Front. Immunol. 2020, 11, 614402. [Google Scholar] [CrossRef] [PubMed]
- Vo, T.S. Natural products targeting FcεRI receptor for anti-allergic therapeutics. J. Food Biochem. 2020, 44, e13335. [Google Scholar] [CrossRef] [PubMed]
- Gaihre, Y.G.; Iwamoto, A.; Oogai, S.; Hamajima, H.; Tsuge, K.; Nagata, Y.; Yanagita, T. Perilla pomace obtained from four different varieties have different levels and types of polyphenols and anti-allergic activity. Cytotechnology 2022, 74, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Han, S.Y.; Bae, J.Y.; Park, S.H.; Kim, Y.H.; Park, J.H.Y.; Kang, Y.H. Resveratrol inhibits IgE-mediated basophilic mast cell degranulation and passive cutaneous anaphylaxis in mice. J. Nutr. 2013, 143, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Shim, S.Y.; Park, J.R.; Byun, D.S. 6-Methoxyluteolin from Chrysanthemum zawadskii var. latilobum suppresses histamine release and calcium influx via down-regulation of FcεRI α chain expression. J. Microbiol. Biotechnol. 2012, 22, 622–627. [Google Scholar] [CrossRef]
- Tamura, S.; Yoshihira, K.; Kawano, T.; Murakami, N. Inhibitor for FcεRI expression on mast cell from Verbasucum thapsus L. Bioorg. Med. Chem. Lett. 2018, 28, 3342–3345. [Google Scholar] [CrossRef]
- Tamura, S.; Yoshihira, K.; Tokumaru, M.; Zisheng, X.; Murakami, N. Inhibitors for expression of IgE receptor on human mast cell from Puerariae Flos. Bioorg. Med. Chem. Lett. 2010, 20, 3872–3875. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feed Mixing List | AIN93G | Perilla Diet | Chive Diet |
|---|---|---|---|
| Milk casein | 20.00 | 20.00 | 20.00 |
| L-Cystine | 0.30 | 0.30 | 0.30 |
| Cornstarch | 63.1986 | 61.1986 | 61.1986 |
| Vegetable extract | 0.0000 | 2.0000 | 2.0000 |
| Edible oil | 7.00 | 7.00 | 7.00 |
| Cellulose powder | 5.00 | 5.00 | 5.00 |
| AIG-93 mineral mix | 3.50 | 3.50 | 3.50 |
| Contains AIG-93 vitamins (choline deuterate) | 1.00 | 1.00 | 1.00 |
| Tertiary butylhydroquinone | 0.00140 | 0.00140 | 0.00140 |
| Total | 100.000 | 100.000 | 100.000 |
| Score | Symptoms |
|---|---|
| 0 | No symptoms |
| 1 | Scratching and rubbing around the nose and head |
| 2 | Puffiness around the eyes and mouth, pilar erecti, reduced activity, and/or decreased activity with increased respiratory rate |
| 3 | Wheezing, labored respiration, and cyanosis around the mouth and the tail |
| 4 | No activity after prodding or tremor and convulsion |
| 5 | Death |
| Vegetable (Scientific Name) | Total Polyphenol Content (mg/g Chlorogenic Acid Equivalent) | Antioxidant Capacity (Mg/G L-Ascorbic Acid Equivalent) |
|---|---|---|
| Tomato (Solanum lycopersicum) | 4.35 ± 0.11 | 1.13 ± 0.02 |
| Bell pepper (Capsicum annuum ‘Grossum’) | 17.97 ± 5.79 | 1.41 ± 0.00 |
| Green pepper (Capsicum annuum ‘Grossum’) | 19.14 ± 0.66 | 1.45 ± 0.00 |
| Pumpkin (Cucurbita maxima) | 4.11 ± 0.05 | 1.06 ± 0.01 |
| Eggplant (Solanum melongena) | 15.10 ± 1.10 | 1.43 ± 0.01 |
| Bitter melon (Momordica charantia) | 20.85 ± 0.30 | 1.45 ± 0.00 |
| Snap pea (Pisum sativum) | 5.00 ± 0.14 | 1.24 ± 0.01 |
| Broccoli (Brassica oleracea var. italic) | 22.69 ± 0.28 | 1.41 ± 0.00 |
| Asparagus (Asparagus officinalis) | 18.56 ± 0.38 | 1.41 ± 0.00 |
| Welsh onion (Allium fistulosum) | 5.22 ± 0.06 | 1.12 ± 0.01 |
| Komatsuna (Brassica rapa var. perviridis) | 32.52 ± 0.24 | 1.41 ± 0.00 |
| Spinach (Spinacia oleracea) | 17.56 ± 0.18 | 1.41 ± 0.00 |
| Garland chrysanthemum (Glebionis coronaria) | 32.38 ± 1.46 | 1.39 ± 0.00 |
| Cabbage (Brassica oleracea var. capitate) | 12.84 ± 0.14 | 1.45 ± 0.00 |
| Perilla (Perilla frutescens var. crispa f. viridis) | 115.94 ± 7.84 | 1.39 ± 0.00 |
| Chives (Allium tuberosum) | 9.51 ± 0.17 | 1.40 ± 0.01 |
| Carrot (Daucus carota subsp. sativus) | 3.87 ± 0.07 | 0.77 ± 0.03 |
| Onion (Allium cepa) | 4.55 ± 0.11 | 0.76 ± 0.02 |
| Garlic (Allium sativum) | 3.27 ± 0.01 | 0.63 ± 0.01 |
| Myoga (Zingiber mioga) | 5.62 ± 0.27 | 1.92 ± 0.01 |
| Daikon (Raphanus sativus var. hortensis) | 5.61 ± 0.08 | 1.45 ± 0.00 |
| Ginger (Zingiber officinale) | 12.63 ± 0.87 | 1.44 ± 0.00 |
| Vegetable (Scientific Name) | Cell Adaptation Concentration (µg/mL) | Maximum Inhibition (%) (Concentration of Vegetable Extracts Added) 1 |
|---|---|---|
| Tomato (Solanum lycopersicum) | ≤250 | 0 |
| Bell pepper (Capsicum annuum ‘Grossum’) | ≤250 | 0 |
| Green pepper (Capsicum annuum ‘Grossum’) | ≤63 | 0 |
| Pumpkin (Cucurbita maxima) | ≤250 | 0 |
| Eggplant (Solanum melongena) | ≤250 | 0 |
| Bitter melon (Momordica charantia) | ≤16 | 0 |
| Snap pea (Pisum sativum) | ≤16 | 0 |
| Broccoli (Brassica oleracea var. italic) | ≤250 | 0 |
| Asparagus (Asparagus officinalis) | ≤250 | 0 |
| Welsh onion (Allium fistulosum) | ≤31 | 0 |
| Komatsuna (Brassica rapa var. perviridis) | ≤63 | 0 |
| Spinach (Spinacia oleracea) | ≤250 | 0 |
| Garland chrysanthemum (Glebionis coronaria) | ≤16 | 27 ± 4 (8 µg/mL) |
| Cabbage (Brassica oleracea var. capitate) | ≤63 | 0 |
| Perilla (Perilla frutescens var. crispa f. viridis) | ≤500 | 44 ± 4 (500 µg/mL) |
| Chives (Allium tuberosum) | ≤250 | 49 ± 4 (250 µg/mL) |
| Carrot (Daucus carota subsp. sativus) | ≤31 | 0 |
| Onion (Allium cepa) | ≤63 | 0 |
| Garlic (Allium sativum) | ≤16 | 0 |
| Myoga (Zingiber mioga) | ≤63 | 19 ± 3 (8 µg/mL) |
| Daikon (Raphanus sativus var. hortensis) | ≤125 | 0 |
| Ginger (Zingiber officinale) | ≤250 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takemoto, K.; Ganlin, T.; Iji, M.; Narukawa, T.; Koyama, T.; Hao, L.; Watanabe, H. Vegetable Extracts as Therapeutic Agents: A Comprehensive Exploration of Anti-Allergic Effects. Nutrients 2024, 16, 693. https://doi.org/10.3390/nu16050693
Takemoto K, Ganlin T, Iji M, Narukawa T, Koyama T, Hao L, Watanabe H. Vegetable Extracts as Therapeutic Agents: A Comprehensive Exploration of Anti-Allergic Effects. Nutrients. 2024; 16(5):693. https://doi.org/10.3390/nu16050693
Chicago/Turabian StyleTakemoto, Kazuhito, Tian Ganlin, Masaki Iji, Takahiro Narukawa, Tomohisa Koyama, Luo Hao, and Hiroyuki Watanabe. 2024. "Vegetable Extracts as Therapeutic Agents: A Comprehensive Exploration of Anti-Allergic Effects" Nutrients 16, no. 5: 693. https://doi.org/10.3390/nu16050693
APA StyleTakemoto, K., Ganlin, T., Iji, M., Narukawa, T., Koyama, T., Hao, L., & Watanabe, H. (2024). Vegetable Extracts as Therapeutic Agents: A Comprehensive Exploration of Anti-Allergic Effects. Nutrients, 16(5), 693. https://doi.org/10.3390/nu16050693