
Effect of Pre-, Pro-, and Synbiotics on Biomarkers of Systemic Inflammation in Children: A Scoping Review


Abstract

1. Introduction
- How many RCTs have assessed the effects of pre-, pro-, and synbiotics on inflammation in infants and children?
- What diseases have been studied?
- What pre-, pro-, and synbiotic preparations have been evaluated?
- What pro- and anti-inflammatory biomarkers have been evaluated?
- Is there sufficient evidence to support meta-analysis?
- What further research is required?
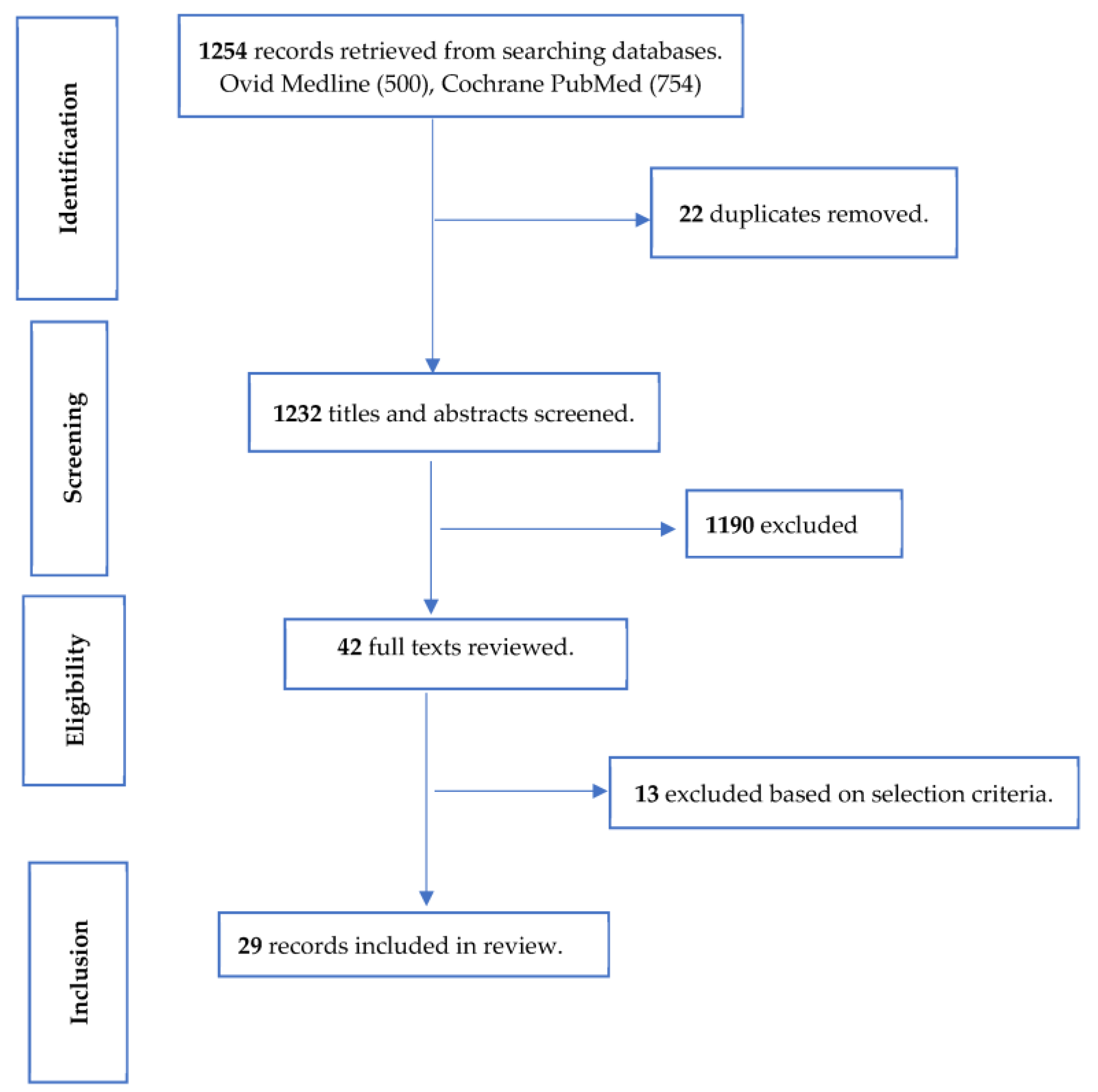
2. Materials and Methods
2.1. Search Strategy
2.2. Eligibility Criteria
- Population: infants and children aged 0–18 years
- Intervention: prebiotics; probiotics (single- or multi-strain); synbiotics
- Comparator: unexposed groups (routine treatment; control; no pre-, pro-, synbiotic)
- Outcomes: blood/serum concentration of inflammatory/pro-inflammatory biomarkers or anti-inflammatory markers
- Study design: RCTs
- Reviews, case studies, conference abstracts, observational studies, and papers that were not in English were excluded.
2.3. Data Extraction and Analysis
- Publication details: first author, publication year, year of study, study design, study population, study location, sample size, characteristics of study participants (health status, age), study arms and interventions evaluated (pre-, pro-, and/or synbiotic) and main study findings.
- Outcomes of interest: blood levels of pro-inflammatory or anti-inflammatory biomarkers.
3. Results
3.1. Health Status and Pre-, Pro-, and Synbiotic Interventions
3.2. Biomarkers of Systemic Inflammation
3.3. Effects of Pre-, Pro-, and Synbiotics on Biomarkers of Systemic Inflammation in Healthy Children
3.4. Effects of Pre-, Pro-, and Synbiotics on Biomarkers of Systemic Inflammation in Children with Diseases
3.5. Effects on Inflammatory Biomarkers according to Intervention Tested
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yu, D.; Meng, X.; de Vos, W.M.; Wu, H.; Fang, X.; Maiti, A.K. Implications of Gut Microbiota in Complex Human Diseases. Int. J. Mol. Sci. 2021, 22, 12661. [Google Scholar] [CrossRef] [PubMed]
- Inamura, K. Gut microbiota contributes towards immunomodulation against cancer: New frontiers in precision cancer therapeutics. Semin. Cancer Biol. 2021, 70, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Swanson, K.S.; Gibson, G.R.; Hutkins, R.; Reimer, R.A.; Reid, G.; Verbeke, K.; Scott, K.P.; Holscher, H.D.; Azad, M.B.; Delzenne, N.M.; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of synbiotics. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 687–701. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tran, D.Q.; Rhoads, J.M. Probiotics in Disease Prevention and Treatment. J. Clin. Pharmacol. 2018, 58, S164–S179. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, H.; Chen, T.; Shi, L.; Wang, D.; Tang, D. Regulatory role of short-chain fatty acids in inflammatory bowel disease. Cell Commun. Signal 2022, 20, 64. [Google Scholar] [CrossRef]
- Momo Kadia, B.; Otiti, M.I.; Ramsteijn, A.S.; Sow, D.; Faye, B.; Heffernan, C.; Hall, L.J.; Webster, J.P.; Walker, A.W.; Allen, S. Modulating the early-life gut microbiota using pro-, pre-, and synbiotics to improve gut health, child development, and growth. Nutr. Rev. 2023, 82, 244–247. [Google Scholar] [CrossRef]
- Milajerdi, A.; Mousavi, S.M.; Sadeghi, A.; Salari-Moghaddam, A.; Parohan, M.; Larijani, B.; Esmaillzadeh, A. The effect of probiotics on inflammatory biomarkers: A meta-analysis of randomized clinical trials. Eur. J. Nutr. 2020, 59, 633–649. [Google Scholar] [CrossRef]
- Ding, L.N.; Ding, W.Y.; Ning, J.; Wang, Y.; Yan, Y.; Wang, Z.B. Effects of Probiotic Supplementation on Inflammatory Markers and Glucose Homeostasis in Adults with Type 2 Diabetes Mellitus: A Systematic Review and Meta-Analysis. Front. Pharmacol. 2021, 12, 770861. [Google Scholar] [CrossRef]
- Kazemi, A.; Soltani, S.; Ghorabi, S.; Keshtkar, A.; Daneshzad, E.; Nasri, F.; Mazloomi, S.M. Effect of probiotic and synbiotic supplementation on inflammatory markers in health and disease status: A systematic review and meta-analysis of clinical trials. Clin. Nutr. 2020, 39, 789–819. [Google Scholar] [CrossRef] [PubMed]
- Noshadi, N.; Heidari, M.; Naemi Kermanshahi, M.; Zarezadeh, M.; Sanaie, S.; Ebrahimi-Mameghani, M. Effects of Probiotics Supplementation on CRP, IL-6, and Length of ICU Stay in Traumatic Brain Injuries and Multiple Trauma Patients: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Evid. Based Complement. Altern. Med. 2022, 2022, 4674000. [Google Scholar] [CrossRef] [PubMed]
- Nazari, M.; Faghfoori, Z.; As’Habi, A.; Arab, A.; Hajianfar, H. Probiotic consumption and inflammatory markers in athletes: A systematic review and meta-analysis. Int. J. Food Prop. 2020, 23, 1402–1415. [Google Scholar] [CrossRef]
- Tanaka, M.; Nakayama, J. Development of the gut microbiota in infancy and its impact on health in later life. Allergol. Int. 2017, 66, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Seyedsadjadi, N.; Grant, R. The Potential Benefit of Monitoring Oxidative Stress and Inflammation in the Prevention of Non-Communicable Diseases (NCDs). Antioxidants 2020, 10, 15. [Google Scholar] [CrossRef] [PubMed]
- Collinson, S.; Deans, A.; Padua-Zamora, A.; Gregorio, G.V.; Li, C.; Dans, L.F.; Allen, S.J. Probiotics for treating acute infectious diarrhoea. Cochrane Database Syst. Rev. 2020, 12, CD003048. [Google Scholar]
- Munn, Z.; Peters, M.D.J.; Stern, C.; Tufanaru, C.; McArthur, A.; Aromataris, E. Systematic review or scoping review? Guidance for authors when choosing between a systematic or scoping review approach. BMC Med. Res. Methodol. 2018, 18, 143. [Google Scholar] [CrossRef] [PubMed]
- Goehring, K.C.; Marriage, B.J.; Oliver, J.S.; Wilder, J.A.; Barrett, E.G.; Buck, R.H. Similar to Those Who Are Breastfed, Infants Fed a Formula Containing 2’-Fucosyllactose Have Lower Inflammatory Cytokines in a Randomized Controlled Trial. J. Nutr. 2016, 146, 2559–2566. [Google Scholar] [CrossRef]
- López-Velázquez, G.; Parra-Ortiz, M.; Mora Ide, L.; García-Torres, I.; Enríquez-Flores, S.; Alcántara-Ortigoza, M.A.; Angel, A.G.-D.; Velázquez-Aragón, J.; Ortiz-Hernández, R.; Cruz-Rubio, J.M.; et al. Effects of Fructans from Mexican Agave in Newborns Fed with Infant Formula: A Randomized Controlled Trial. Nutrients 2015, 7, 8939–8951. [Google Scholar] [CrossRef]
- Li, X.; Peng, Y.; Li, Z.; Christensen, B.; Heckmann, A.B.; Lagerqvist, C.; Stenlund, H.; Lönnerdal, B.; Hernell, O.; West, C.E. Serum cytokine patterns are modulated in infants fed formula with probiotics or milk fat globule membranes: A randomized controlled trial. PLoS ONE 2021, 16, e0251293. [Google Scholar] [CrossRef]
- Kusumo, P.D.; Bela, B.; Wibowo, H.; Munasir, Z.; Surono, I.S. Lactobacillus plantarum IS-10506 supplementation increases faecal sIgA and immune response in children younger than two years. Benef. Microbes 2019, 10, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Karlsson Videhult, F.; Andersson, Y.; Öhlund, I.; Stenlund, H.; Hernell, O.; West, C.E. Impact of probiotics during weaning on the metabolic and inflammatory profile: Follow-up at school age. Int. J. Food Sci. Nutr. 2015, 66, 686–691. [Google Scholar] [CrossRef] [PubMed]
- Larnkjær, A.; Michaelsen, K.F.; Rytter, M.J.H.; Mølgaard, C.; Laursen, R.P. Effect of probiotics on thymus size and markers of infection in late infancy: A randomized controlled trial. Pediatr. Res. 2021, 89, 563–568. [Google Scholar] [CrossRef] [PubMed]
- Drabińska, N.; Jarocka-Cyrta, E.; Złotkowska, D.; Abramowicz, P.; Krupa-Kozak, U. Daily oligofructose-enriched inulin intake impacts bone turnover markers but not the cytokine profile in pediatric patients with celiac disease on a gluten-free diet: Results of a randomised, placebo-controlled pilot study. Bone 2019, 122, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Aly, H.; Said, R.N.; Wali, I.E.; Elwakkad, A.; Soliman, Y.; Awad, A.R.; Shawky, M.A.; Alam, M.S.A.; Mohamed, M.A. Medically Graded Honey Supplementation Formula to Preterm Infants as a Prebiotic: A Randomized Controlled Trial. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 966–970. [Google Scholar] [CrossRef] [PubMed]
- Nicolucci, A.C.; Hume, M.P.; Martínez, I.; Mayengbam, S.; Walter, J.; Reimer, R.A. Prebiotics Reduce Body Fat and Alter Intestinal Microbiota in Children Who Are Overweight or with Obesity. Gastroenterology 2017, 153, 711–722. [Google Scholar] [CrossRef] [PubMed]
- D’Auria, E.; Panelli, S.; Lunardon, L.; Pajoro, M.; Paradiso, L.; Beretta, S.; Loretelli, C.; Tosi, D.; Perini, M.; Bedogni, G.; et al. Rice flour fermented with Lactobacillus paracasei CBA L74 in the treatment of atopic dermatitis in infants: A randomized, double- blind, placebo- controlled trial. Pharmacol. Res. 2021, 163, 105284. [Google Scholar] [CrossRef]
- Jeong, K.; Kim, M.; Jeon, S.A.; Kim, Y.H.; Lee, S. A randomized trial of Lactobacillus rhamnosus IDCC 3201 tyndallizate (RHT3201) for treating atopic dermatitis. Pediatr. Allergy Immunol. 2020, 31, 783–792. [Google Scholar] [CrossRef]
- Prakoeswa, C.R.S.; Herwanto, N.; Prameswari, R.; Astari, L.; Sawitri, S.; Hidayati, A.N.; Indramaya, D.M.; Kusumowidagdo, E.R.; Surono, I.S. Lactobacillus plantarum IS-10506 supplementation reduced SCORAD in children with atopic dermatitis. Benef. Microbes 2017, 8, 833–840. [Google Scholar] [CrossRef]
- Han, Y.; Kim, B.; Ban, J.; Lee, J.; Kim, B.J.; Choi, B.S.; Hwang, S.; Ahn, K.; Kim, J. A randomized trial of Lactobacillus plantarum CJLP133 for the treatment of atopic dermatitis. Pediatr. Allergy Immunol. 2012, 23, 667–673. [Google Scholar] [CrossRef]
- Wang, I.J.; Wang, J.Y. Children with atopic dermatitis show clinical improvement after Lactobacillus exposure. Clin. Exp. Allergy 2015, 45, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Jerzynska, J.; Stelmach, W.; Balcerak, J.; Woicka-Kolejwa, K.; Rychlik, B.; Blauz, A.; Wachulec, M.; Stelmach, P.; Majak, P.; Stelmach, I. Effect of Lactobacillus rhamnosus GG and vitamin D supplementation on the immunologic effectiveness of grass-specific sublingual immunotherapy in children with allergy. Allergy Asthma Proc. 2016, 37, 324–334. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.Y.; Fu, L.S.; Lin, H.K.; Shen, C.Y.; Chen, Y.J. Evaluation of the effect of Lactobacillus paracasei (HF.A00232) in children (6–13 years old) with perennial allergic rhinitis: A 12-week, double-blind, randomized, placebo-controlled study. Pediatr. Neonatol. 2014, 55, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.L.; Hale, J.; Wiltschut, J.; Lehmann, H.; Dunstan, J.A.; Prescott, S.L. Effects of probiotic supplementation for the first 6 months of life on allergen- and vaccine-specific immune responses. Clin. Exp. Allergy 2006, 36, 1227–1235. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.S.; Jan, R.L.; Lin, Y.L.; Chen, H.H.; Wang, J.Y. Randomized placebo-controlled trial of lactobacillus on asthmatic children with allergic rhinitis. Pediatr. Pulmonol. 2010, 45, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.F.; Chie, W.C.; Wang, I.J. Efficacy of Lactobacillus Administration in School-Age Children with Asthma: A Randomized, Placebo-Controlled Trial. Nutrients 2018, 10, 1678. [Google Scholar] [CrossRef] [PubMed]
- Olivares, M.; Castillejo, G.; Varea, V.; Sanz, Y. Double-blind, randomised, placebo-controlled intervention trial to evaluate the effects of Bifidobacterium longum CECT 7347 in children with newly diagnosed coeliac disease. Br. J. Nutr. 2014, 112, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Klemenak, M.; Dolinšek, J.; Langerholc, T.; Di Gioia, D.; Mičetić-Turk, D. Administration of Bifidobacterium breve Decreases the Production of TNF-α in Children with Celiac Disease. Dig. Dis. Sci. 2015, 60, 3386–3392. [Google Scholar] [CrossRef]
- Fortes, P.M.; Teles Filho, R.V.; Azevêdo, L.H.S.; Queiroz, V.C.J.; Costa, P. Inflammatory cytokines and lipid profile in children and adolescents with nephrotic syndrome receiving L. plantarum: A randomized, controlled feasibility trial. Rev. Assoc. Medica Bras. 2020, 66, 1487–1492. [Google Scholar] [CrossRef]
- Fujii, T.; Ohtsuka, Y.; Lee, T.; Kudo, T.; Shoji, H.; Sato, H.; Nagata, S.; Shimizu, T.; Yamashiro, Y. Bifidobacterium breve enhances transforming growth factor beta1 signaling by regulating Smad7 expression in preterm infants. J. Pediatr. Gastroenterol. Nutr. 2006, 43, 83–88. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, L.; Yang, Z.; Chen, F.; Zhang, Y. Effects of probiotics on ghrelin and lungs in children with acute lung injury: A double-blind randomized, controlled trial. Pediatr. Pulmonol. 2018, 53, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Ellis, C.L.; Bokulich, N.A.; Kalanetra, K.M.; Mirmiran, M.; Elumalai, J.; Haapanen, L.; Schegg, T.; Rutledge, J.C.; Raff, G.; Mills, D.A.; et al. Probiotic administration in congenital heart disease: A pilot study. J. Perinatol. 2013, 33, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, Z.; Xu, Z.; Wang, Z.; Feng, J. Probiotics prevent Hirschsprung’s disease-associated enterocolitis: A prospective multicenter randomized controlled trial. Int. J. Color. Dis. 2015, 30, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Shukla, A.; Gaur, P.; Aggarwal, A. Effect of probiotics on clinical and immune parameters in enthesitis-related arthritis category of juvenile idiopathic arthritis. Clin. Exp. Immunol. 2016, 185, 301–308. [Google Scholar] [CrossRef]
- Angurana, S.K.; Bansal, A.; Singhi, S.; Aggarwal, R.; Jayashree, M.; Salaria, M.; Mangat, N.K. Evaluation of Effect of Probiotics on Cytokine Levels in Critically Ill Children with Severe Sepsis: A Double-Blind, Placebo-Controlled Trial. Crit. Care Med. 2018, 46, 1656–1664. [Google Scholar] [CrossRef]
- de Freitas, M.B.; Moreira, E.A.M.; Oliveira, D.L.; Tomio, C.; da Rosa, J.S.; Moreno, Y.M.F.; Barbosa, E.; Ludwig Neto, N.; Buccigrossi, V.; Guarino, A.; et al. Effect of synbiotic supplementation in children and adolescents with cystic fibrosis: A randomized controlled clinical trial. Eur. J. Clin. Nutr. 2018, 72, 736–743. [Google Scholar] [CrossRef]
- Oozeer, R.; van Limpt, K.; Ludwig, T.; Ben Amor, K.; Martin, R.; Wind, R.D.; Boehm, G.; Knol, J. Intestinal microbiology in early life: Specific prebiotics can have similar functionalities as human-milk oligosaccharides. Am. J. Clin. Nutr. 2013, 98, 561S–571S. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Search # | Search Term(s) and Combinations |
|---|---|
| 1 | (probiotic * or prebiotic * or synbiotic *): ti, ab, kw |
| 2 | (infant * or neonat * or newborn * or p?ediatric or child *): ti, ab, kw |
| 3 | (‘Erythrocyte sedimentation rate’ or ESR or interleukin * or IL or ‘tumo?r necrosis factor’ or TNF or cytokine * or Orosomucoid or ORM or ‘alpha 1 acid glycoprotein’ or AGP or CRP or ‘C reaction protein’ or GlycA or ‘systemic inflammation’): ti, ab, kw |
| 4 | (Randomi?ed controlled trial): ti, ab, kw |
| 5 | (Review or protocol): ti, ab, kw |
| 6 | 1 AND 2 AND 3 AND 4 NOT 5 |
| Author, Year | Study Design and Population 1 | Intervention(s) | Biomarker(s) | Significant Intervention Effects | ||
|---|---|---|---|---|---|---|
| Anti-Inflammatory | Pro-Inflammatory | Overall Effect 2 | ||||
| Prebiotic studies | ||||||
| Goehring et al., 2016 USA [18] | Sub-study; 3-arm RCT; 113 term newborns | Prebiotic: Infant formulas containing 2.4 g total oligosaccharides/L (control: GOS only; experimental formulas: GOS + 0.2 or 1.0 g 2′-fucosyllactose/L [2′FL/L]) from 5 days to 4 months | IFN-α2; IFN-γ; IL-1α; IL-1β; IL-6; IFN-γ–induced protein 10 (IP-10); TNF-α; IL-1 receptor antagonist (IL-1ra); regulated upon activation, normal T cell expressed and secreted (RANTES); IL-10; lymphocyte count; PBMC phenotyping and stimulation ex vivo; measured at 6 weeks of age | Lower IL-1α, IL-1ra, IL-1β, IL-6 and TNF-α (p ≤ 0.05) in both 2′FL/L groups. Lower TNF-α, IFN-γ and IL-6 in 0.2 g 2′FL/L group in culture supernatants of RSV-stimulated PBMCs ex-vivo (p ≤ 0.05). Greater percentage of sub-G0/G1 CD56+ cells in 0.2 g 2′FL/L group (p ≤ 0.05). | - | ↓ |
| Probiotic studies—single strain | ||||||
| Li et al., 2021 China [20] | 3-arm RCT: 495 infants aged 21 ± 7 days randomised to formula with F19, milk fat globule membrane or standard formula | Probiotic: L. paracasei subsp. paracasei strain F19 108 cfu/L from age 21 ± 7 days until 4 months | IL-2, IL-6, IL-17A, IFN-γ, TNF-α; TGF-β1, IL-4 measured at 4 months of age | Lower IFN-γ (p = 0.008) | Higher IL-2 (p = 0.024) | ↑/↓ |
| Kusumo et al., 2019 Indonesia [21] | 4-arm RCT: 38 children aged 12–24 months randomised to placebo, probiotic, zinc, and probiotic + zinc | Probiotic: L. plantarum IS-10506 of ‘dadih’ origin 2.3 × 1010 cfu/g/d for 90 days | TNF-α and TGF-β1 pre-/post-treatment ratio measured at baseline and end of study | - | - | → |
| Karlsson et al., 2015 Sweden [22] | 2-arm RCT: 120 children aged 4–13 months | Probiotic: L. paracasei subsp. paracasei F19 (LF19; deposition number LMG P-17806) 108 cfu daily from age 4 to 13 months | HsCRP at 8–9 years of age | - | - | → |
| Probiotic studies—multi-strain | ||||||
| Larnkjaer et al., 2021 Denmark [23] | 2-arm RCT: 186 children expected to start daycare at age 8–14 months | Probiotic: L. rhamnosus LGG® plus B. animalis subsp. lactis, BB-12® each 109 cfu daily from up to 12 weeks before starting daycare for 6 months | CRP measured at 6 months | - | - | → |
| Synbiotic/probiotic studies | ||||||
| López-Velázquez et al., 2015 Mexico [19] | 5-arm RCT: 600 infants aged 20 ± 7 days | Prebiotics: Infant formula with fructans obtained from Agave tequilana var Weber: Metlin® and Metlos®. Probiotic: L. rhamnosus (LR) 0.3 × 107 cfu. Group 1: Synbiotic (LR + Metlin® + Metlos®); Group 2: Synbiotic (LR + Metlin®); Group 3: Synbiotic (LR + Metlos®); Group 4: Probiotic (LR; Group 5: infant formula only; daily to age 3 months | CRP measured at baseline and 3 months | - | - | → |
| Author, Year, Country | Study Design and Population | Intervention(s) | Biomarker(s) | Significant Intervention Effects | |||
|---|---|---|---|---|---|---|---|
| Anti-Inflammatory | Pro-Inflammatory | Overall Effect 1 | Clinical Effect 2 | ||||
| Atopic/allergic disorders: atopic dermatitis | |||||||
| D’Auria et al., 2021 Italy [27] | 2-arm RCT: 58 children aged 6–36 months with atopic dermatitis | Probiotic: 8g rice-dried powder containing heat-killed L. paracasei CBA L74 powder daily for 12 weeks | G-CSF, GM-CSF, IFN-γ, IL-1β, IL-2, IL-5, IL-6, IL-7, IL-8, IL-12, IL-17A, MCP-1, MIP-1β, TNF-α, IL-4, IL-10, and IL-13 measured at 12 weeks | - | - | → | Yes |
| Jeong et al., 2020 Korea [28] | 2-arm RCT: 66 children aged 1–12 years with atopic dermatitis | Probiotic: tyndallized L. rhamnosus (RHT3201) 1010 cfu daily for 12 weeks | Eosinophil cationic protein (ECP), CRP, IL-31, TNF-α, chemokines; IL-4, IL-10 measured at 6 and 12 weeks | Improved ECP level at week 12 (p = 0.022) | - | ↓ | Yes |
| Prakoeswa et al., 2017 Indonesia [29] | 2-arm RCT: 22 children aged 0–14 years with mild and moderate atopic dermatitis | Probiotic: microencapsulated L. plantarum IS-10506 1010 cfu/day twice daily for 12 weeks | IFN-γ, IL-17; IL-4, forkhead box P3 (Foxp3+)/IL-10 ratio; CD4+ expression of IL-4, IFN-γ, Foxp3+/IL-10 ratio, and IL-17 measured at 12 weeks | IFN-γ decreased more in probiotic group (p = 0.006). CD4+ expression of IFN-γ fell in probiotic but not control group. Increased Foxp3+/IL-10 ratio in probiotic compared with control group (p = 0.001). | IL-4 decreased more in probiotic group (p < 0.001) | ↑/↓ | Yes |
| Han et al., 2012 Korea [30] | 2-arm RCT: 79 children aged 12 months to 13 years with atopic dermatitis | Probiotic: L. plantarum CJLP133 0.5 × 1010 cfu twice daily for 12 weeks | TNF-α, IFN-γ, and IL-4 measured at 2 and 14 weeks | IFN-γ decreased from baseline (p < 0.001) and was lower in the probiotic than placebo group at 14 weeks (p < 0.05) | IL-4 decreased from baseline (p = 0.049) | ↑/↓ | Yes |
| Wang et al., 2015 Taiwan [31] | 4-arm RCT: 220 children aged 1–18 years with moderate-to-severe atopic dermatitis | Probiotics: L. paracasei (LP; 2 × 109 cfu) or L. fermentum (LF; 2 × 109 cfu) or both (4 × 109 cfu) daily for 3 months | IFN-γ, TNF-α, and TGF-β, IL-4 measured at baseline and 3 months | Increased IL-4 from baseline (p = 0.04) | - | ↓ | Yes |
| Atopic/allergic disorders: allergic rhinitis | |||||||
| Jerzynska et al., 2016 Poland [32] | 3-arm RCT: 100 children aged 5–12 years with allergic rhinitis and sensitivity to grass pollen receiving sublingual immunotherapy | Probiotic: L. rhamnosus GG (Dicoflor 30) 3 billion live cultures daily for 5 months | CD4+CD25+Foxp3+ (forkhead box P3) cells, Toll-like receptor (TLR) 4, IL-1, IL-6, TNF-α, TLR activation, IL-10, and TGF β-1 in cell supernatants of grass-stimulated PBMCs measured at baseline and 5 months | Increased forkhead box P3 cell induction at 12 months (p < 0.05) | - | ↓ | Yes |
| Lin et al., 2014 Taiwan [33] | 2-arm RCT: 60 children aged 6–13 years with perennial allergic rhinitis receiving levocetirizine (antihistamine) | Probiotic: L. paracasei (LP) strain HF. A00232 5 × 109 cfu/day for 8 weeks | IFN-γ, IL-4, IL-10, and TGF-β measured at baseline and weeks 8 and 12 | - | - | → | Yes |
| Atopic/allergic disorders: other | |||||||
| Taylor et al., 2006 Australia [34] | 2-arm RCT; 118 newborns of women with a history of allergic disease and positive allergen skin prick test | Probiotic: L. acidophilus LAVRI-A1 3 × 109 daily for first 6 months | IL-5, IL-6, TNF-α, IL-10, IL-13, and TGF- β measured at 6 months | Reduced IL-5 production in response to polyclonal stimulation (p = 0.044); reduced TNF-α responsiveness to house dust mite allergens (p = 0.046) | Reduced TGF-β production in response to polyclonal stimulation (p = 0.015); reduced IL-10 responses to tetanus toxoid vaccine antigen (p = 0.03) and house dust mite allergens (p = 0.014) | ↑/↓ | N/a |
| Chen et al., 2010 Taiwan [35] | 2-arm RCT: 105 children aged 6–12 years with asthma and allergic rhinitis | Probiotic: L. gasseri PM-A0005 (A5) 2 × 109 cells twice daily for 8 weeks | TNF-α, IFN-γ, IL-12p40, IL-10, and IL-13 produced by PBMCs stimulated with PHA, Dermatophagoides pteronyssinus (Der p), or Der p supplemented with L. gasseri A5 measured at baseline and 8 weeks | Decreased TNF-α in PBMCs in medium alone and stimulated with Der p. (p < 0.05). Decreased IFN-γ, IL-12 in PBMCs stimulated with PHA or Der p (p < 0.05). | Decreased IL-13 in medium alone and stimulated with PHA or Der p (p < 0.05) | ↑/↓ | Yes |
| Huang et al., 2018 Taiwan [36] | 4-arm RCT: 160 children aged 6–18 years with asthma | Probiotic: L. paracasei GMNL-133 (BCRC 910520, CCTCC M2011331), L. fermentum GM-090 (BCRC 910259, CCTCC M204055), or both for 3 months (frequency and dose not specified) | IFN-γ, TNF α, and IL-4 measured at baseline and 3 months | → | → | → | Yes |
| Autoimmune disorders: coeliac disease | |||||||
| Drabińska et al., 2019 Poland [24] | 2-arm RCT: 34 children aged 4–17 years with coeliac disease | Prebiotic: 10 g of oligofructose-enriched inulin (Synergy 1) daily for three months | IL-1β, IL-1ra, IL-6, IL-8, IL-12p70, TNF-α, and IL-10 measured at baseline and 3 months | → | → | → | N/a |
| Olivares et al., 2014 Spain [37] | 2-arm RCT: 33 children aged 2–17 years with newly diagnosed coeliac disease | Probiotic: B. longum CECT 7347 10⁹ cfu daily for 3 months | Lymphocyte subsets, TNF-α, IF-γ, IL-10, IL-13, and TGF-β measured at baseline and 3 months | Fall in CD3+ (p = 0.013) and HLA-DR+ T lymphocytes (p = 0.029) from baseline; decreased CD3+ T lymphocytes (p = 0.004) compared with controls | - | ↓ | Yes |
| Klemenak et al., 2015 Slovenia [38] | 3-arm RCT: 49 children with coeliac disease compared to 18 healthy controls | Probiotic: B. breve BR03 and B632 2 × 109 cfu daily for three months | TNF-α and IL-10 measured at baseline and 3 months | Decreased TNF-α levels from baseline (p < 0.05) | - | ↓ | N/a |
| Autoimmune disorders: Other | |||||||
| Fortes et al., 2020 Brazil [39] | 2-arm RCT: 4 children aged 2–17 years with compensated or partially compensated nephrotic syndrome and dyslipidaemia | Probiotic: L. plantarum Lp-G18 2.5 × 109 cfu daily for 12 weeks | TNF-α and IL-10 measured at baseline and during and at the end of the study | - | - | → | No |
| Shukla et al., 2016 India [44] | 2-arm RCT: 46 children aged 13–19 years with active enthesitis-related juvenile inflammatory arthritis | Probiotic: (S. thermophilus, B. breve, B. longum, B. infantis, L. acidophilus, L. plantarum, L. paracasei and L. delbrueckii (VSL#3) capsules 112·5 billion cells twice daily for 12 weeks | Blood Th1, Th2, Th17 and Treg cells, ESR, CRP, IFN-γ, IL-6, IL-17, TNF-α, IL−4, and IL-10 measured at baseline and 12 weeks | Fall in IL-6 from baseline (p = 0·007) | Increased Th2 cells (p < 0.05) and IL-10 in placebo group (p = 0·013) from baseline | ↓/→ | No |
| Preterm infants | |||||||
| Aly et al., 2017 Egypt [25] | 4-arm RCT: 40 newborns with gestational age ≤ 34 weeks and age > 3 days | Prebiotic: different doses of medically graded bee honey daily for 14 days | CD4 and CD8 cells measured at baseline and 7 and 14 days | - | - | → | Yes |
| Fujii et al., 2006 Japan [40] | 2-arm RCT: 19 preterm newborns | Probiotic: B. breve M-16V 109 cells twice daily from several hours after birth until discharge | IL-4, IL-5, IL-6, IFN-γ, TGF-β1, and TNF-α measured at baseline and 14 and 28 days | Increased TGF-β1 from baseline on days 14 (p = 0.026) and 28 (p = 0.029); increased TGF-β1 on day 28 (p = 0.005) | - | ↓ | No |
| Overweight/obesity | |||||||
| Nicolucci et al., 2017 Canada [26] | 2-arm RCT: 42 overweight and obese children aged 7–12 years | Prebiotic: oligofructose-enriched inulin (Synergy1) 8 g/day for 16 weeks | CRP, IFNγ, IL-1β, IL-4, IL-6, IL-8, IL-10, IL-33, monocyte chemoattractant protein-1, TNF-α, and lipopolysaccharide measured at baseline and 16 weeks | Decreased IL-6 from baseline compared with controls (p = 0.01) | - | ↓ | Yes |
| Other diseases; severe illness | |||||||
| Wang et al., 2018 China [41] | 2-arm RCT: 80 children aged ≤14 years with acute lung injury | Probiotic: Eosinophil Lactobacillus 5 × 106 cfu 3 times daily for 10 days | TNF-α and IL-6 measured at baseline and 10 days | Decreased TNF-α (p = 0.0005) and IL-6 (p = 0.0004) | - | ↓ | Yes |
| Angurana et al., 2018 India [45] | 2-arm RCT: 100 children aged 3 months to 12 years with severe sepsis | Probiotic: VSL#3 (L. paracasei DSM 24734, L. plantarum DSM 24730, L. acidophilus DSM 24735, L. delbrueckii subsp. bulgaricus DSM 24734, B. longum DSM 24736, B. infantis DSM 24737, B. breve DSM 24732, S. thermophilus DSM 24731; Danisco-Dupont USA, Madison, WI); 450 billion bacteria twice daily for 7 days | IL-12p70, IL-6, IL-17, TNF-α, IL-10, and TGF -β1 measured on days 1 and 7 | Decreased IL-6 (p = 0.001), IL-12p70 (p = 0.001); IL-17 (p = 0.01), TNF-α (p = 0.01] and increased IL-10 (p = 0.02), TGF-β1 (p = 0.01) in probiotic vs. controls at day 7. From baseline, fall in IL-6 (p = 0.001), IL-12p70 (p= 0.01], IL-17 (p = 0.01), TNF-α (p = 0.001); increase in IL-10 (p = 0.001) and TGF-β1 (p = 0.001) | - | ↓ | Yes |
| Other diseases | |||||||
| de Freitas et al., 2018 Brazil [46] | 3-arm RCT: children with cystic fibrosis received synbiotic (n = 22, mean (SD) age: 9.6 ± 2.8 years) vs. healthy controls (n = 17, mean age: 8.6 ± 3.0 years) | Synbiotic: L. paracasei, L. rhamnosus, L. acidophilus, B. lactis 108–109 cfu daily of each strain and FOS (5.5 g/day) (Lactofos®) for 90 days | High sensitivity CRP, IL-1β, IL-6, IL-8, IL-12, TNF-α, myeloperoxidase, nitric oxide metabolites (NOx), and IL-10 measured at baseline and 90 days | Fall from baseline in NOx (p = 0.030) | - | ↓ | No |
| Wang et al., 2015 China [43] | 2-arm RCT: 60 children < 18 years with Hirschsprung’s disease at risk of enterocolitis | Probiotic: Bifidobacterium, L. acidophilus, and E. faecalis; (BIFICO; strains not specified) >108 cfu daily for 4 weeks | T lymphocyte subsets, TNF-α, IFN-γ, IL-6, IL-10 unclear when measured | Increased CD4+ cells (p = 0.048) and CD4+/CD8+ ratio (p = 0.005) compared with controls; decreased TNF-α p < 0.01), IFN-γ (p = 0.029), IL-6 (p = 0.015); increased IL-10 (p = 0.011) compared with controls | - | ↓ | Yes |
| Ellis et al., 2013 USA [42] | 2-arm RCT: 16 term infants with congenital heart disease at risk of necrotising enterocolitis | Probiotic: B. longum ssp. infantis ATCC 15,697 4.2 × 109 cfu twice two daily for 8 weeks or until death or discharge if sooner | IFN-γ, IL-1β, IL-8, TNF-α, and IL-10 measured weekly | - | Increased IFN-γ (p = 0.007) and IL-1β (p = 0.04) | ↑ | N/a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kadia, B.M.; Allen, S.J. Effect of Pre-, Pro-, and Synbiotics on Biomarkers of Systemic Inflammation in Children: A Scoping Review. Nutrients 2024, 16, 336. https://doi.org/10.3390/nu16030336
Kadia BM, Allen SJ. Effect of Pre-, Pro-, and Synbiotics on Biomarkers of Systemic Inflammation in Children: A Scoping Review. Nutrients. 2024; 16(3):336. https://doi.org/10.3390/nu16030336
Chicago/Turabian StyleKadia, Benjamin Momo, and Stephen John Allen. 2024. "Effect of Pre-, Pro-, and Synbiotics on Biomarkers of Systemic Inflammation in Children: A Scoping Review" Nutrients 16, no. 3: 336. https://doi.org/10.3390/nu16030336
APA StyleKadia, B. M., & Allen, S. J. (2024). Effect of Pre-, Pro-, and Synbiotics on Biomarkers of Systemic Inflammation in Children: A Scoping Review. Nutrients, 16(3), 336. https://doi.org/10.3390/nu16030336