
Health-Promoting Potential of Mandarin Pomace Extracts Enriched with Phenolic Compounds

 ,
,  , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemical Reagents
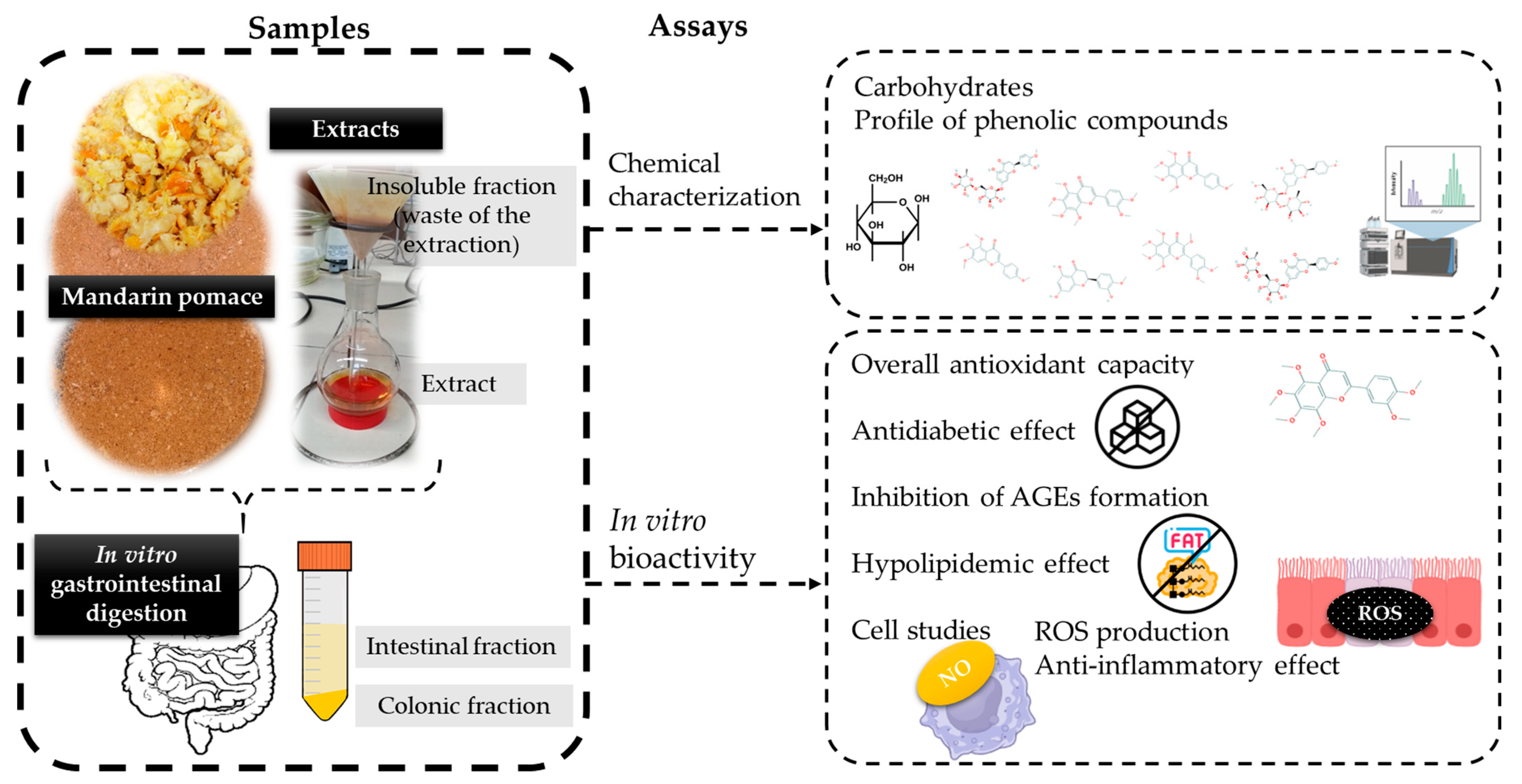
2.2. Samples
2.2.1. Raw Material
2.2.2. Mandarin Pomace Extracts
2.2.3. In Vitro Human Digests
2.3. Chemical Characterization of Dried Mandarin Pomace and Extracts
- Carbohydrates
- Profile of phenolic compounds
2.4. In Vitro Bioactivity
- Overall antioxidant capacity
- Antidiabetic effect
- Inhibition of AGE formation
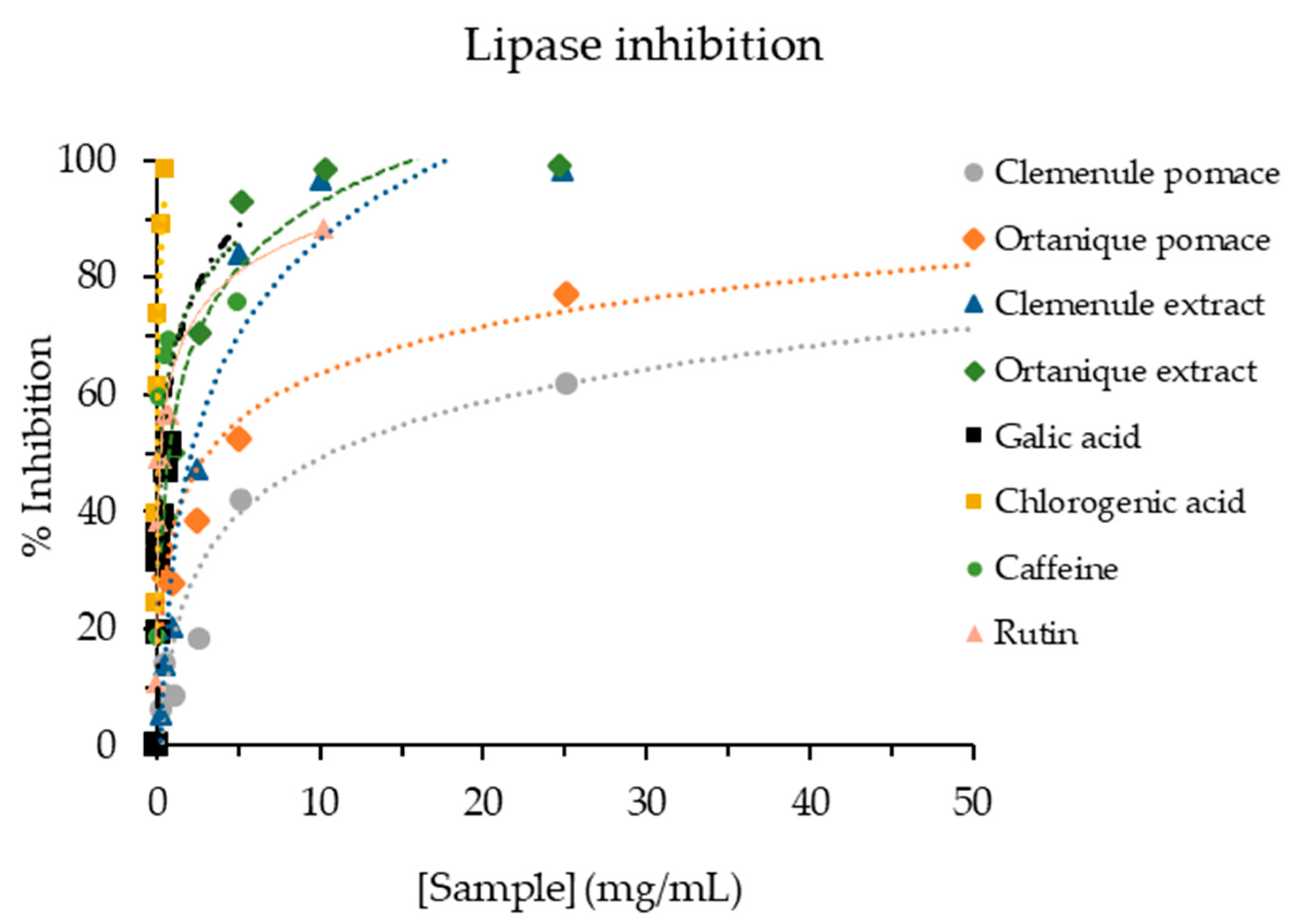
- Hypolipidemic effect
- Cell studiesNormal human subepithelial myofibroblast colonic cells (CCD-18Co) and mouse macrophages (RAW264.7) were used. Cells were grown using 75 cm2 cell culture flasks in complete cell culture medium (described in Section 2.1), incubating at 37 °C and 5% CO2 (100% relative humidity) for 24 h till cell confluence was achieved. The extract tests were performed in triplicate and in three different passages by preparing 10 mg/mL of solutions in PBS (10 mM; pH 7.4) and filtered with a porous medium of 0.22 µm. MTT assay was carried out to determine the cell viability of different concentrations of mandarin extracts [4] (see Supplementary Materials).
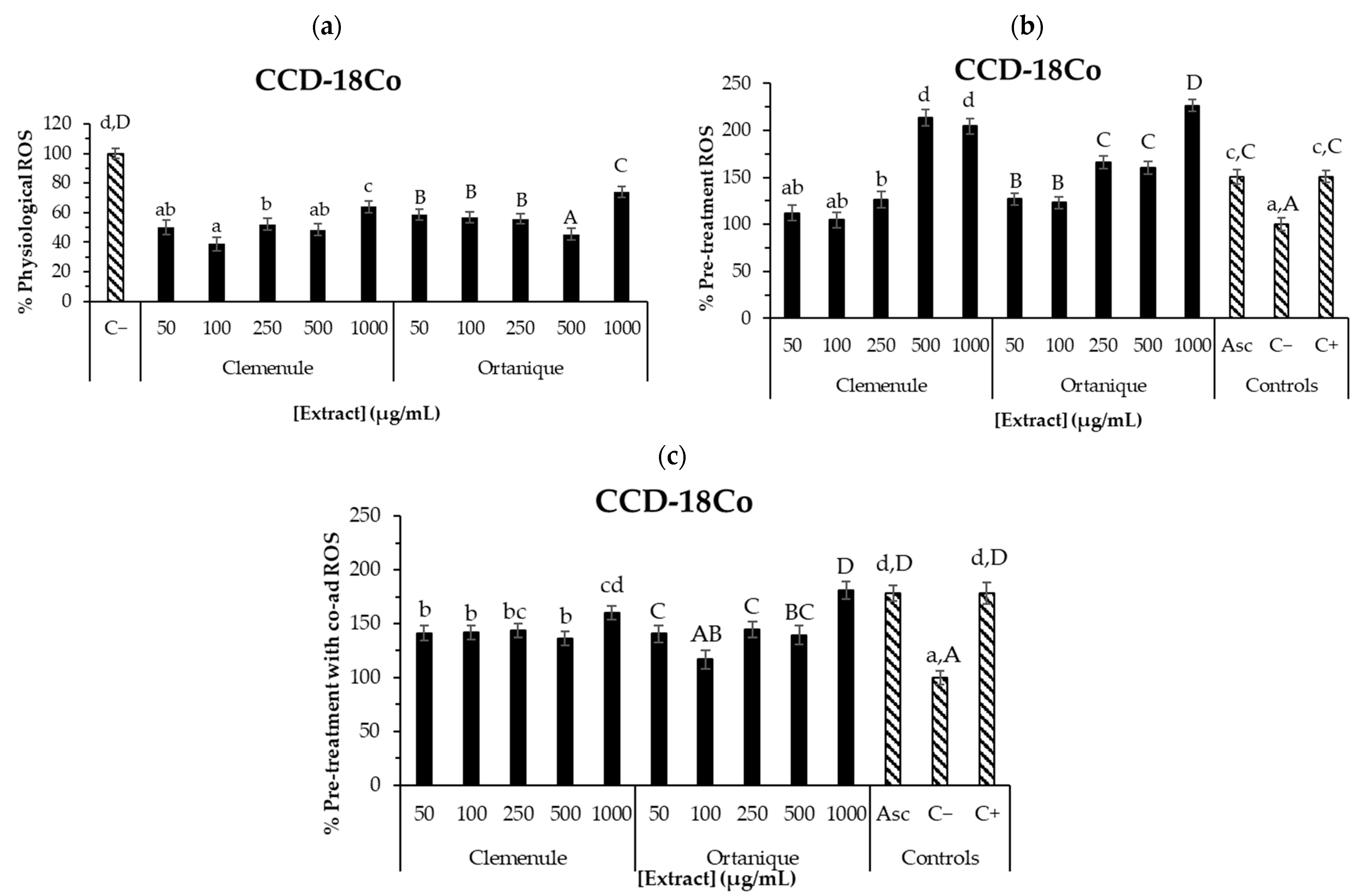
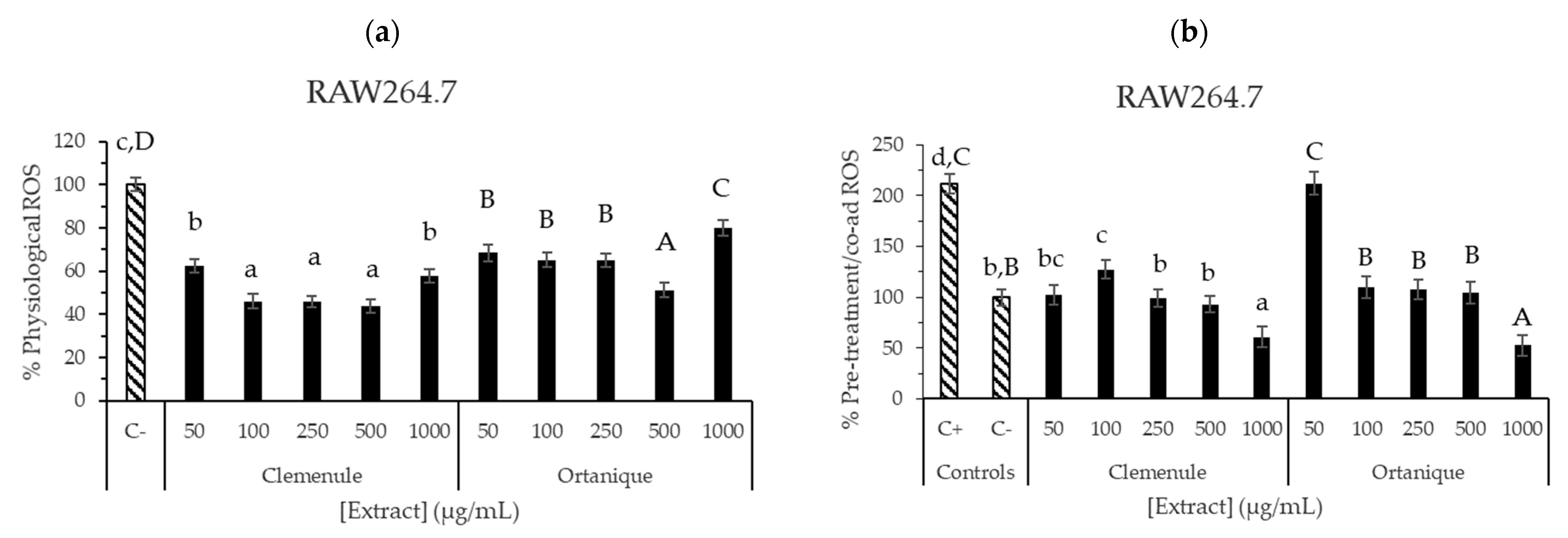
- Reactive Oxygen Species (ROS) production assayCCD-18Co and RAW 264.7 cell ROS production was measured [28] by seeding 10,000 CCD-18Co and 80,000 RAW264.7 cells/well. DCFH-DA fluorescent probe was employed to measure ROS within the cell by performing the following steps: add different concentrations of the extracts (1–1000 µg/mL) to each well; perform incubation for 24 h; add 2 µL of DCFH-DA to each well; perform incubation for 30 min; remove supernatants; perform one cell wash with PBS; and add the same extract solutions to each well and measure fluorescence in a microplate reader at λexcitation = 485 nm and λemission = 528 nm. Under oxidation-induced conditions, tert-butyl hydroperoxide (1 mM) was used as the oxidative agent. Afterwards, cell viability was assessed (20 µL of MTT reagent in each well), following the described procedure in the Supplementary Materials, to correct ROS values with cell viability:
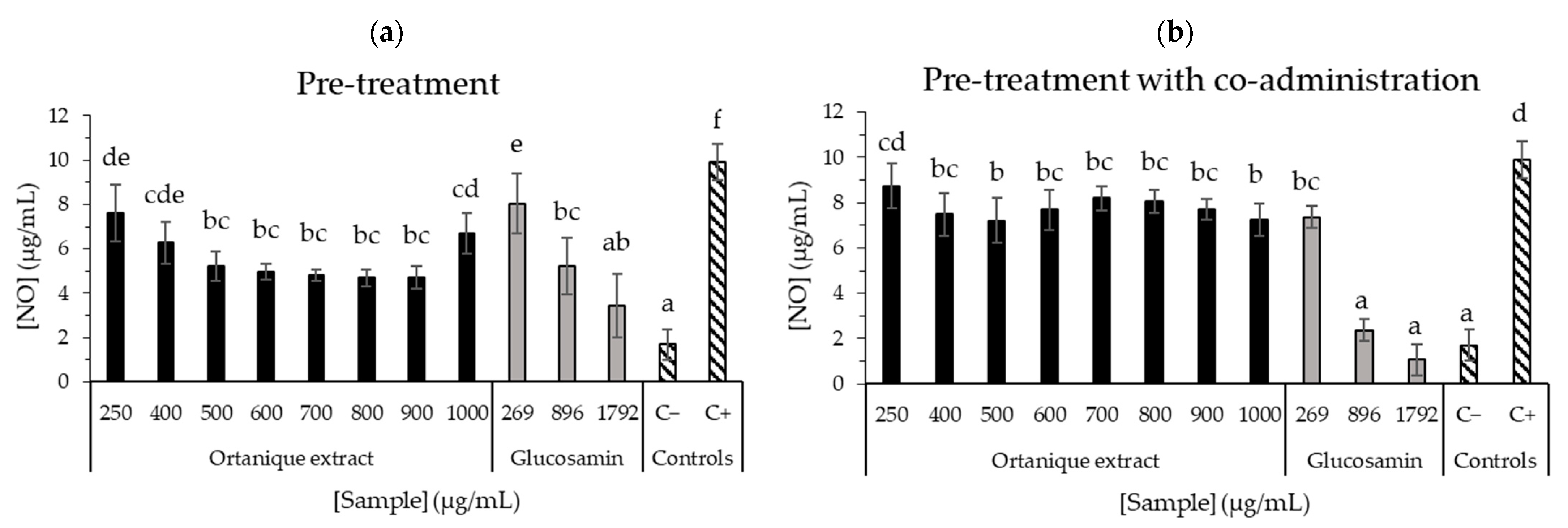
- Anti-inflammatory effectNitric oxide (NO) production was measured in RAW264.7 mouse macrophages under stimulating conditions (lipopolysaccharide, LPS) according to Fernández-Fernández et al. [20]. The macrophages on each well were treated with concentrations of mandarin extracts (250–1000 µg/mL) prepared in a medium without FBS and incubated for 24 h, followed by a transfer of cells’ supernatants to another 96-well plate. Then, Griess reagent was added to each well, incubation was performed for 15 min at room temperature, and measurement was taken at 550 nm in a microplate reader. In parallel, the standard curve (sodium nitrite, 0–10 µg/mL) was also measured. Positive and negative controls were also measured for a medium without FBS but with LPS, and a medium without FBS, respectively.
2.5. Statistical Analysis
3. Results and Discussion
3.1. Composition on Carbohydrates
3.2. Phenolic Profile of Mandarin Pomace Extracts
3.3. Health-Promoting Potential
3.3.1. Antioxidant Effect
3.3.2. Antidiabetic and Anti-Glycative Effect
3.3.3. Hypolipidemic Effect of Mandarin Pomace and Their Respective Extracts
3.3.4. Anti-Inflammatory Effect of Mandarin Pomace Extracts
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Núñez-Gómez, V.; González-Barrio, R.; Periago, M.J. Interaction between Dietary Fibre and Bioactive Compounds in Plant By-Products: Impact on Bioaccessibility and Bioavailability. Antioxidants 2023, 12, 976. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Fernández, A.M.; Dellacassa, E.; Medrano-Fernandez, A.; Del Castillo, M.D. Citrus Waste Recovery for Sustainable Nutrition and Health. In Food Wastes and By-Products: Nutraceutical and Health Potential; Campos-Vega, R., Oomah, B.D., Vergara-Castañeda, H.A., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2020; pp. 193–211. ISBN 9781119534105. [Google Scholar]
- Sorrenti, V.; Burò, I.; Consoli, V.; Vanella, L. Recent Advances in Health Benefits of Bioactive Compounds from Food Wastes and By-Products: Biochemical Aspects. Int. J. Mol. Sci. 2023, 24, 2019. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Fernández, A.M.; Dellacassa, E.; Nardin, T.; Larcher, R.; Gámbaro, A.; Medrano-Fernandez, A.; del Castillo, M.D. In Vitro Bioaccessibility of Bioactive Compounds from Citrus Pomaces and Orange Pomace Biscuits. Molecules 2021, 26, 3480. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Ranjit, A.; Sharma, K.; Prasad, P.; Shang, X.; Gowda, K.G.M.; Keum, Y.-S. Bioactive Compounds of Citrus Fruits: A Review of Composition and Health Benefits of Carotenoids, Flavonoids, Limonoids, and Terpenes. Antioxidants 2022, 11, 239. [Google Scholar] [CrossRef] [PubMed]
- Costa, C.; Tsatsakis, A.; Mamoulakis, C.; Teodoro, M.; Briguglio, G.; Caruso, E.; Tsoukalas, D.; Margina, D.; Dardiotis, E.; Kouretas, D.; et al. Current Evidence on the Effect of Dietary Polyphenols Intake on Chronic Diseases. Food Chem. Toxicol. 2017, 110, 286–299. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-T.; Chu, H.-L.; Chyau, C.-C.; Chu, C.-C.; Duh, P.-D. Protective Effects of Sweet Orange (Citrus Sinensis) Peel and Their Bioactive Compounds on Oxidative Stress. Food Chem. 2012, 135, 2119–2127. [Google Scholar] [CrossRef] [PubMed]
- dos Santos Dias, M.M.; Duarte Martino, H.S.; Noratto, G.; Roque-Andrade, A.; Stringheta, P.C.; Talcott, S.; Mota Ramos, A.; Mertens-Talcott, S.U. Anti-Inflammatory Activity of Polyphenolics from Açai (Euterpe oleracea Martius) in Intestinal Myofibroblasts CCD-18Co Cells. Food Funct. 2015, 6, 3249–3256. [Google Scholar] [CrossRef] [PubMed]
- Thomas-Valdés, S.; Theoduloz, C.; Jiménez-aspee, F.; Burgos-edwards, A.; Schmeda-hirschmann, G. Changes in Polyphenol Composition and Bioactivity of the Native Chilean White Strawberry (Fragaria chiloensis spp. chiloensis f. chiloensis) after In Vitro Gastrointestinal Digestion. Food Res. Int. 2018, 105, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, J.; Bains, Y.; Guha, S.; Kahn, A.; Hall, D.; Bose, N.; Gugliucci, A.; Kapahi, P. The Role of Advanced Glycation End Products in Aging and Metabolic Diseases: Bridging Association and Causality. Cell Metab. 2018, 28, 337–352. [Google Scholar] [CrossRef] [PubMed]
- Gugliucci, A. Formation of Fructose-Mediated Advanced Glycation End Products and Their Roles in Metabolic and Inflammatory Diseases. Adv. Nutr. 2017, 8, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Sergent, T.; Vanderstraeten, J.; Winand, J.; Beguin, P.; Schneider, Y.J. Phenolic Compounds and Plant Extracts as Potential Natural Anti-Obesity Substances. Food Chem. 2012, 135, 68–73. [Google Scholar] [CrossRef]
- Lu, K.; Yip, Y.M. Therapeutic Potential of Bioactive Flavonoids from Citrus Fruit Peels toward Obesity and Diabetes Mellitus. Future Pharmacol. 2023, 3, 14–37. [Google Scholar] [CrossRef]
- Stafussa, A.P.; Maciel, G.M.; Bortolini, D.G.; Maroldi, W.V.; Ribeiro, V.R.; Fachi, M.M.; Pontarolo, R.; Bach, F.; Pedro, A.C.; Haminiuk, C.W.I. Bioactivity and Bioaccessibility of Phenolic Compounds from Brazilian Fruit Purees. Future Foods 2021, 4, 100066. [Google Scholar] [CrossRef]
- Li, C.X.; Wang, F.R.; Zhang, B.; Deng, Z.Y.; Li, H.Y. Stability and Antioxidant Activity of Phenolic Compounds during In Vitro Digestion. J. Food Sci. 2023, 88, 696–716. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.Q.; Chen, J.B.; Zhao, C.N.; Liu, X.J.; Chen, Y.Y.; Liang, J.J.; Cao, J.P.; Wang, Y.; Sun, C. De Advances in Extraction and Purification of Citrus Flavonoids. Food Front. 2023, 4, 750–781. [Google Scholar] [CrossRef]
- Fernández-Fernández, A.M.; Iriondo-DeHond, A.; Nardin, T.; Larcher, R.; Dellacassa, E.; Medrano-Fernandez, A.; del Castillo, M.D. In Vitro Bioaccessibility of Extractable Compounds from Tannat Grape Skin Possessing Health Promoting Properties with Potential to Reduce the Risk of Diabetes. Foods 2020, 9, 1575. [Google Scholar] [CrossRef] [PubMed]
- Hollebeeck, S.; Borlon, F.; Schneider, Y.-J.; Larondelle, Y.; Rogez, H. Development of a Standardised Human In Vitro Digestion Protocol Based on Macronutrient Digestion Using Response Surface Methodology. Food Chem. 2013, 138, 1936–1944. [Google Scholar] [CrossRef] [PubMed]
- Masuko, T.; Minami, A.; Iwasaki, N.; Majima, T.; Nishimura, S.I.; Lee, Y.C. Carbohydrate Analysis by a Phenol-Sulfuric Acid Method in Microplate Format. Anal. Biochem. 2005, 339, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Fernández, A.M.; Iriondo-DeHond, A.; Dellacassa, E.; Medrano-Fernandez, A.; del Castillo, M.D. Assessment of Antioxidant, Antidiabetic, Antiobesity, and Anti-Inflammatory Properties of a Tannat Winemaking by-Product. Eur. Food Res. Technol. 2019, 245, 1539–1551. [Google Scholar] [CrossRef]
- Olt, V.; Báez, J.; Curbelo, R.; Boido, E.; Amarillo, M.; Gámbaro, A.; Alborés, S.; Gerez García, N.; Cesio, M.V.; Heinzen, H.; et al. Tannat Grape Pomace as an Ingredient for Potential Functional Biscuits: Bioactive Compound Identification, In Vitro Bioactivity, Food Safety, and Sensory Evaluation. Front. Nutr. 2023, 10, 1241105. [Google Scholar] [CrossRef] [PubMed]
- Slinkard, K.; Singleton, V.L. Total Phenol Analysis: Automation and Comparison with Manual Methods. Am. J. Enol. Vitic. 1977, 28, 49–55. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant Activity Applying an Improved ABTS Radical Cation Decolorization Assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and Validation of an Improved Oxygen Radical Absorbance Capacity Assay Using Fluorescein as the Fluorescent Probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef] [PubMed]
- Dávalos, A.; Bartolomé, B.; Gómez-Cordovés, C. Antioxidant Properties of Commercial Grape Juices and Vinegars. Food Chem. 2005, 93, 325–330. [Google Scholar] [CrossRef]
- Li, K.; Yao, F.; Du, J.; Deng, X.; Li, C. Persimmon Tannin Decreased the Glycemic Response through Decreasing the Digestibility of Starch and Inhibiting α-Amylase, α-Glucosidase, and Intestinal Glucose Uptake. J. Agric. Food Chem. 2018, 66, 1629–1637. [Google Scholar] [CrossRef] [PubMed]
- Starowicz, M.; Zieliński, H. Inhibition of Advanced Glycation End-Product Formation by High Antioxidant-Leveled Spices Commonly Used in European Cuisine. Antioxidants 2019, 8, 100. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Fernández, A.M.; Dellacassa, E.; Nardin, T.; Larcher, R.; Ibañez, C.; Terán, D.; Gámbaro, A.; Medrano-Fernandez, A.; del Castillo, M.D. Tannat Grape Skin: A Feasible Ingredient for the Formulation of Snacks with Potential for Reducing the Risk of Diabetes. Nutrients 2022, 14, 419. [Google Scholar] [CrossRef] [PubMed]
- Peña-Vázquez, G.I.; Dominguez-Fernández, M.T.; Camacho-Zamora, B.D.; Hernandez-Salazar, M.; Urías-Orona, V.; De Peña, M.P.; de la Garza, A.L. In Vitro Simulated Gastrointestinal Digestion Impacts Bioaccessibility and Bioactivity of Sweet Orange (Citrus Sinensis) Phenolic Compounds. J. Funct. Foods 2022, 88, 104891. [Google Scholar] [CrossRef]
- Cilla, A.; Rodrigo, M.J.; De Ancos, B.; Sánchez-Moreno, C.; Cano, M.P.; Zacariás, L.; Barberá, R.; Alegriá, A. Impact of High-Pressure Processing on the Stability and Bioaccessibility of Bioactive Compounds in Clementine Mandarin Juice and Its Cytoprotective Effect on Caco-2 Cells. Food Funct. 2020, 11, 8951–8962. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, Y.; Zhu, T.; Yang, S.; Cao, J.; Li, X.; Wang, L.S.; Sun, C. Beneficial Regulatory Effects of Polymethoxyflavone—Rich Fraction from Ougan (Citrus reticulata Cv. Suavissima) Fruit on Gut Microbiota and Identification of Its Intestinal Metabolites in Mice. Antioxidants 2020, 9, 831. [Google Scholar] [CrossRef] [PubMed]
- Denaro, M.; Smeriglio, A.; Trombetta, D. Antioxidant and Anti-Inflammatory Activity of Citrus Flavanones Mix and Its Stability after in Vitro Simulated Digestion. Antioxidants 2021, 10, 140. [Google Scholar] [CrossRef] [PubMed]
- Mosele, J.I.; Macià, A.; Romero, M.-P.; Motilva, M.-J.; Rubió, L. Application of in Vitro Gastrointestinal Digestion and Colonic Fermentation Models to Pomegranate Products (Juice, Pulp and Peel Extract) to Study the Stability and Catabolism of Phenolic Compounds. J. Funct. Foods 2015, 14, 529–540. [Google Scholar] [CrossRef]
- Ye, X.; Cao, D.; Zhao, X.; Song, F.; Huang, Q.; Fan, G.; Wu, F. Chemical Fingerprint and Metabolic Profile Analysis of Citrus Reticulate “Chachi” Decoction by HPLC-PDA-IT-MS n and HPLC-Quadrupole-Orbitrap-MS Method. J. Chromatogr. B 2014, 970, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Mejía, E.; Sacristán, I.; Rosales-Conrado, N.; León-González, M.E.; Madrid, Y. Valorization of Citrus Reticulata Blanco Peels to Produce Enriched Wheat Bread: Phenolic Bioaccessibility and Antioxidant Potential. Antioxidants 2023, 12, 1742. [Google Scholar] [CrossRef] [PubMed]
- Aiello, F.; Caputo, P.; Oliviero Rossi, C.; Restuccia, D.; Spizzirri, U.G. Formulation of Antioxidant Gummies Based on Gelatin Enriched with Citrus Fruit Peels Extract. Foods 2024, 13, 320. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Qian, J.; Cao, J.; Wang, D.; Liu, C.; Yang, R.; Li, X.; Sun, C. Antioxidant Capacity, Anticancer Ability and Flavonoids Composition of 35 Citrus (Citrus reticulata Blanco) Varieties. Molecules 2017, 22, 1114. [Google Scholar] [CrossRef] [PubMed]
- Ozdemirli, N.; Kamiloglu, S. Influence of Industrial Blanching, Cutting, and Freezing Treatments on in Vitro Gastrointestinal Digestion Stability of Orange (Citrus sinensis L.) and Lemon (Citrus limon L.) Peel Polyphenols. J. Sci. Food Agric. 2023, 104, 2165–2173. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Kou, G.; Chen, Q.; Li, Y.; Zhou, Z. Protection and Delivery of Mandarin (Citrus Reticulata Blanco) Peel Extracts by Encapsulation of Whey Protein Concentrate Nanoparticles. LWT—Food Sci. Technol. 2019, 99, 24–33. [Google Scholar] [CrossRef]
- Inada, K.O.P.; Revorêdo Silva, T.B.; Lobo, L.A.; Cavalcante Pilotto Domingues, R.M.; Perrone, D.; Monteiro, M. Bioaccessibility of Phenolic Compounds of Jaboticaba (Plinia jaboticaba) Peel and Seed after Simulated Gastrointestinal Digestion and Gut Microbiota Fermentation. J. Funct. Foods 2020, 67, 103851. [Google Scholar] [CrossRef]
- Nieto, J.A.; Fernández-Jalao, I.; Siles-Sánchez, M.d.l.N.; Santoyo, S.; Jaime, L. Implication of the Polymeric Phenolic Fraction and Matrix Effect on the Antioxidant Activity, Bioaccessibility, and Bioavailability of Grape Stem Extracts. Molecules 2023, 28, 2461. [Google Scholar] [CrossRef]
- Yang, C.; Han, Y.; Tian, X.; Sajid, M.; Mehmood, S.; Wang, H.; Li, H. Phenolic Composition of Grape Pomace and Its Metabolism. Crit. Rev. Food Sci. Nutr. 2022, 64, 4865–4881. [Google Scholar] [CrossRef] [PubMed]
- Reguengo, L.M.; Salgaço, M.K.; Sivieri, K.; Maróstica Júnior, M.R. Agro-Industrial by-Products: Valuable Sources of Bioactive Compounds. Food Res. Int. 2022, 152, 110871. [Google Scholar] [CrossRef] [PubMed]
- Munir, H.; Yaqoob, S.; Awan, K.A.; Imtiaz, A.; Naveed, H.; Ahmad, N.; Naeem, M.; Sultan, W.; Ma, Y. Unveiling the Chemistry of Citrus Peel: Insights into Nutraceutical Potential and Therapeutic Applications. Foods 2024, 13, 1681. [Google Scholar] [CrossRef] [PubMed]
- Gironés-Vilaplana, A.; Villaño, D.; Moreno, D.A.; García-Viguera, C. New Isotonic Drinks with Antioxidant and Biological Capacities from Berries (Maqui, Açaí and Blackthorn) and Lemon Juice. Int. J. Food Sci. Nutr. 2013, 64, 897–906. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, S. Functional Foods against Metabolic Syndrome (Obesity, Diabetes, Hypertension and Dyslipidemia) and Cardiovasular Disease. Trends Food Sci. Technol. 2014, 35, 114–128. [Google Scholar] [CrossRef]
- Lordan, S.; Smyth, T.J.; Soler-Vila, A.; Stanton, C.; Paul Ross, R. The α-Amylase and α-Glucosidase Inhibitory Effects of Irish Seaweed Extracts. Food Chem. 2013, 141, 2170–2176. [Google Scholar] [CrossRef] [PubMed]
- Tundis, R.; Bonesi, M.; Sicari, V.; Pellicanò, T.M.; Tenuta, M.C.; Leporini, M.; Menichini, F.; Loizzo, M.R. Poncirus trifoliata (L.) Raf.: Chemical Composition, Antioxidant Properties and Hypoglycaemic Activity via the Inhibition of α-Amylase and α-Glucosidase Enzymes. J. Funct. Foods 2016, 25, 477–485. [Google Scholar] [CrossRef]
- Loizzo, M.R.; Leporini, M.; Sicari, V.; Falco, T.; Pellicanò, T.M.; Tundis, R. Rosa Investigating the In Vitro Hypoglycaemic and Antioxidant Properties of Citrus × Clementina Hort. Juice. Eur. Food Res. Technol. 2018, 244, 523–534. [Google Scholar] [CrossRef]
- Menichini, F.; Tundis, R.; Loizzo, M.R.; Bonesi, M.; Liu, B.; Jones, P.; Persaud, S.J.; Mastellone, V.; Lombardi, P.; Houghton, P.J.; et al. C. medica Cv Diamante Peel Chemical Composition and Influence on Glucose Homeostasis and Metabolic Parameters. Food Chem. 2011, 124, 1083–1089. [Google Scholar] [CrossRef]
- Ha, T.T.; Mai, T.N.P.; Tran, T.T.; Nguyen, N.H.K.; Le, T.D.; Nguyen, V.M. Antioxidant Activity and Inhibitory Efficacy of Citrus Grandis Peel Extract against Carbohydrate Digestive Enzymes In Vitro. Food Sci. Technol. 2022, 42, e109721. [Google Scholar] [CrossRef]
- Benayad, O.; Bouhrim, M.; Tiji, S.; Kharchoufa, L.; Addi, M.; Drouet, S.; Hano, C.; Lorenzo, J.M.; Bendaha, H.; Bnouham, M.; et al. Phytochemical Profile, α-Glucosidase, and α-Amylase Inhibition Potential and Toxicity Evaluation of Extracts from Citrus aurantium (L) Peel, a Valuable by-Product from Northeastern Morocco. Biomolecules 2021, 11, 1555. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Warren, F.J.; Gidley, M.J. Natural Products for Glycaemic Control: Polyphenols as Inhibitors of Alpha-Amylase. Trends Food Sci. Technol. 2019, 91, 262–273. [Google Scholar] [CrossRef]
- Byun, K.; Yoo, Y.; Son, M.; Lee, J.; Jeong, G.-B.; Mok Park, Y.; Hosseini Salekdeh, G.; Lee, B. Advanced Glycation End-Products Produced Systemically and by Macrophages: A Common Contributor to Inflammation and Degenerative Diseases. Pharmacol. Ther. 2017, 177, 44–55. [Google Scholar] [CrossRef]
- Huang, R.; Zhang, Y.; Shen, S.; Zhi, Z.; Cheng, H.; Chen, S.; Ye, X. Antioxidant and Pancreatic Lipase Inhibitory Effects of Flavonoids from Different Citrus Peel Extracts: An in Vitro Study. Food Chem. 2020, 326, 126785. [Google Scholar] [CrossRef] [PubMed]
- Swapna Rekha, S.; Pradeepkiran, J.A.; Bhaskar, M. Bioflavonoid Hesperidin Possesses the Anti-Hyperglycemic and Hypolipidemic Property in STZ Induced Diabetic Myocardial Infarction (DMI) in Male Wister Rats. J. Nutr. Intermed. Metab. 2019, 15, 58–64. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Ashour, M.B.; Abdel-Moneim, A.; Ahmed, O.M. Hesperidin and Naringin Attenuate Hyperglycemia-Mediated Oxidative Stress and Proinflammatory Cytokine Production in High Fat Fed/Streptozotocin-Induced Type 2 Diabetic Rats. J. Diabetes Complicat. 2012, 26, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, H.; Butt, M.S.; Iqbal, M.J. Citrus Peel Extract and Powder Attenuate Hypercholesterolemia and Hyperglycemia Using Rodent Experimental Modeling. Asian Pac. J. Trop. Biomed. 2017, 7, 870–880. [Google Scholar] [CrossRef]
- Lee, Y.-S.; Cha, B.-Y.; Saito, K.; Choi, S.-S.; Wang, X.X.; Choi, B.-K.; Yonezawa, T.; Teruya, T.; Nagai, K.; Woo, J.-T. Effects of a Citrus depressa Hayata (Shiikuwasa) Extract on Obesity in High-Fat Diet-Induced Obese Mice. Eur. J. Integr. Med. 2010, 18, 648–654. [Google Scholar] [CrossRef] [PubMed]
- Teng, J.; Li, Y.; Yu, W.; Zhao, Y.; Hu, X.; Tao, N.P.; Wang, M. Naringenin, a Common Flavanone, Inhibits the Formation of AGEs in Bread and Attenuates AGEs-Induced Oxidative Stress and Inflammation in RAW264.7 Cells. Food Chem. 2018, 269, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, V.M.; Moala, T.; Caria, C.R.e.P.; Moura, C.S.; Amaya-Farfan, J.; Gambero, A.; Macedo, G.A.; Macedo, J.A. Biotransformed Citrus Extract as a Source of Anti-Inflammatory Polyphenols: Effects in Macrophages and Adipocytes. Food Res. Int. 2017, 97, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.-C.; Kuo, C.-T. Hesperidin, Nobiletin, and Tangeretin Are Collectively Responsible for the Anti-Neuroinflammatory Capacity of Tangerine Peel (Citri reticulatae Pericarpium). FOOD Chem. Toxicol. 2014, 71, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, V.M.; Alves Macedo, G.; Macedo, J.A. Citrus Bioactive Phenolics: Role in the Obesity Treatment. LWT—Food Sci. Technol. 2014, 59, 1205–1212. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Carbohydrates | Sucrose | Glucose | Fructose |
|---|---|---|---|---|
| Clemenule | 35.65 ± 1.71 b | 4.39 ± 1.50 a | 9.34 ± 1.26 a | 15.28 ± 0.71 a |
| Ortanique | 34.57 ± 5.68 b | 5.10 ± 0.81 a | 11.15 ± 0.99 a | 15.95 ± 1.27 a |
| Clemenule extract | 65.77 ± 5.56 d | 34.22 ± 2.49 c | 7.36 ± 3.45 a | 35.66 ± 7.01 b |
| Ortanique extract | 53.06 ± 4.68 c | 24.27 ± 1.99 b | 16.99 ± 1.44 b | 37.68 ± 4.55 b |
| Clemenule insoluble fraction | 6.89 ± 0.53 a | - | - | - |
| Ortanique insoluble fraction | 6.88 ± 0.57 a | - | - | - |
| Sample | Sucrose | Glucose | Fructose |
|---|---|---|---|
| Clemenule pomace | 3.80 ± 0.35 a | 3.70 ± 1.65 a | 10.42 ± 0.61 a |
| Ortanique pomace | 3.55 ± 0.80 a | 3.83 ± 0.30 a | 10.17 ± 0.38 a |
| Clemenule extract | 8.44 ± 1.12 b | 3.81 ± 0.85 a | 15.27 ± 0.65 b |
| Ortanique extract | 10.16 ± 1.82 b | 5.46 ± 2.09 a | 17.29 ± 3.39 b |
| Positive ESI | |||||
| Compound 1 | Clemenule extract 2 | Ortanique extract 2 | RT (min) | [M+H]+ (m/z) | Fragments (m/z) |
| Rhoifolin/Isorhoifolin | 0.000999 | 0.001663 | 11.0 | 579.1708 | 271.0595 |
| Isosinensetin/Sinensetin/Tangeretin 1 | 0.016582 | 0.026516 | 16.2 | 373.1282 | 343.0806, 153.0181 |
| Isosinensetin/Sinensetin/Tangeretin 2 | 0.039204 | 0.081296 | 16.7 | 373.1282 | 343.0806, 153.0181 |
| Nobiletin | 0.097034 | 0.169946 | 17.3 | 403.1387 | 373.091, 183.0288 |
| Heptamethoxyflavone | 0.089591 | 0.019377 | 17.6 | 433.1493 | 403.1019, 418.1251 |
| Tetramethylscutellarein | 0.040104 | 0.143404 | 17.7 | 343.1176 | 313.0701, 153.0180 |
| Isosinensetin/Sinensetin/Tangeretin 3 | 0.044997 | 0.134339 | 18.1 | 373.1282 | 343.0806, 153.0181 |
| Negative ESI | |||||
| Compound 1 | Clemenule extract 2 | Ortanique extract 2 | RT (min) | [M-H]− (m/z) | Fragments (m/z) |
| Nariturin-4-glucoside/Naringin glucoside | 0.000068 | 0.000353 | 9.6 | 741.2248 | 271.0639, 151.0035 |
| Rutin | 0.004279 | 0.005631 | 10.3 | 609.1461 | 301.0350, 271.0257 |
| Eriocitrin/Neoeriocitrin 1 | 0.001618 | n.d. | 10.6 | 595.1668 | 287.0580, 151.0034 |
| Naringin/Narirutin | 0.008252 | 0.026907 | 11.2 | 579.1719 | 271.0637, 151.0035 |
| Diosmin isomer 1 | 0.000730 | 0.001610 | 11.3 | 607.1668 | 299.0580, 284.0338 |
| Eriocitrin/Neoeriocitrin 2 | 0.000162 | 0.000044 | 11.3 | 595.1668 | 287.0580, 151.0034 |
| Diosmin isomer 2 | 0.000072 | 0.000175 | 11.4 | 607.1668 | 299.0580, 284.0338 |
| Hesperidin/ Neohesperidin | 0.037499 | 0.027973 | 11.6 | 609.1825 | 301.0739, 151.0035 |
| Poncirin/Isosakuranetin-7-O-rutinoside | 0.000177 | 0.002077 | 13.1 | 593.1876 | 285.0763 |
| TIC | 1,001,915,855 | 1,050,566,219 |
| Intestinal Fraction | Colonic Fraction | ||||||
|---|---|---|---|---|---|---|---|
| Compound | [M+H]+ | O | C | EO | EC | O | C |
| Isosinensetin/Sinensetin/Tangeretin 1 | 373 | 18,176 | 8145 | 122,121 | 9009 | 10,265 | 7488 |
| Isosinensetin/Sinensetin/Tangeretin 2 | 373 | 531 | 122,276 | 898 | 117 | ||
| Nobiletin | 403 | 662 | 110,244 | 2187 | 488,478 | 238 | 254 |
| Heptamethoxyflavone | 433 | 4625 | 28,971 | 7865 | 7245 | 933 | 3474 |
| Isosinensetin/Sinensetin/Tangeretin 3 | 373 | 807 | 220 | 22,460 | 899 | 22,481 | 341 |
| Total area | 24,932 | 148,111 | 154,633 | 627,907 | 34,815 | 11,674 | |
| Samples | Folin Reaction (mg GAE/g Sample) | ABTS (µmol TE/g Sample) | ORAC-FL (µmol TE/g Sample) | |||
|---|---|---|---|---|---|---|
| Samples | Digests | Samples | Digests | Samples | Digests | |
| Clemenule | 17.73 ± 0.75 d | 9.98 ± 0.51 bc | 108.4 ± 2.1 c | 90.1 ± 1.5 b | 284.6 ± 14.4 cd | 135.5 ± 22.2 a |
| Clemenule extract | 26.86 ± 3.47 f | 9.74 ± 0.76 bc | 310.6 ± 9.6 f | 83.2 ± 2.6 b | 346.7 ± 34.7 d | 142.8 ± 38.1 ab |
| Clemenule insoluble fraction | 6.93 ± 0.20 a | - | 51.1 ± 1.3 a | - | 587.4 ± 61.1 f | - |
| Ortanique | 16.72 ± 0.35 d | 11.00 ± 0.65 c | 120.8 ± 4.1 d | 92.0 ± 2.0 b | 288.2 ± 24.0 cd | 232.5 ± 26.7 bc |
| Ortanique extract | 22.29 ± 3.19 e | 9.20 ± 0.58 abc | 271.7 ± 13.0 e | 91.7 ± 10.3 b | 269.0 ± 43.3 cd | 891.6 ± 123.8 g |
| Ortanique insoluble fraction | 7.75 ± 0.31 ab | - | 63.2 ± 1.7 a | - | 483.4 ± 24.8 e | - |
| Samples | IC50 (mg/mL) | |||||
|---|---|---|---|---|---|---|
| α-Glucosidase | α-Amylase | AGE Inhibition | ||||
| Samples | Digests | Samples | Digests | Samples | Digests | |
| Clemenule | 4.92 ± 0.27 d | 3.97 ± 0.97 c,d | 70.19 ± 11.16 c | 58.04 ± 2.09 b,c | 3.25 ± 0.47 b | 31.23 ± 2.57 c |
| Clemenule extract | 2.08 ± 0.08 a,b | 13.50 ± 0.37 f | 16.23 ± 0.19 a | 28.79 ± 1.33 a | 1.00 ± 0.05 a | 16.50 ± 1.38 ab |
| Ortanique | 3.42 ± 0.64 b,c | 4.93 ± 0.41 d | 50.07 ± 2.42 b | 105.68 ± 16.03 d | 5.37 ± 0.20 c | 18.60 ± 3.91 b |
| Ortanique extract | 1.72 ± 0.09 a | 11.07 ± 1.11 e | 19.15 ± 0.18 a | 69.64 ± 1.22 c | 1.19 ± 0.08 a | 12.77 ± 0.29 a |
| Standards (µg/mL) | AC = 4.0 ± 0.3 | AC = 34.1 ± 0.8 | AG = 46.5 ± 2.8 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Fernández, A.M.; Dellacassa, E.; Curbelo, R.; Nardin, T.; Larcher, R.; Medrano-Fernandez, A.; del Castillo, M.D. Health-Promoting Potential of Mandarin Pomace Extracts Enriched with Phenolic Compounds. Nutrients 2024, 16, 2370. https://doi.org/10.3390/nu16142370
Fernández-Fernández AM, Dellacassa E, Curbelo R, Nardin T, Larcher R, Medrano-Fernandez A, del Castillo MD. Health-Promoting Potential of Mandarin Pomace Extracts Enriched with Phenolic Compounds. Nutrients. 2024; 16(14):2370. https://doi.org/10.3390/nu16142370
Chicago/Turabian StyleFernández-Fernández, Adriana Maite, Eduardo Dellacassa, Romina Curbelo, Tiziana Nardin, Roberto Larcher, Alejandra Medrano-Fernandez, and María Dolores del Castillo. 2024. "Health-Promoting Potential of Mandarin Pomace Extracts Enriched with Phenolic Compounds" Nutrients 16, no. 14: 2370. https://doi.org/10.3390/nu16142370
APA StyleFernández-Fernández, A. M., Dellacassa, E., Curbelo, R., Nardin, T., Larcher, R., Medrano-Fernandez, A., & del Castillo, M. D. (2024). Health-Promoting Potential of Mandarin Pomace Extracts Enriched with Phenolic Compounds. Nutrients, 16(14), 2370. https://doi.org/10.3390/nu16142370