
Bioactive Components of Human Milk and Their Impact on Child’s Health and Development, Literature Review
 , , , , , and
, , , , , and
Abstract
1. Introduction

2. Compounds of Mainly Immunological Importance
2.1. Antibodies
2.2. Lactoferrin, Lysozyme, Lactahedrin and Lactoperoxidase
2.3. Cytokines
3. Growth Factor Family
3.1. Epidermal Growth Factors
3.2. Heparin-Binding EGF-like Growth Factor (HB-EGF)
3.3. Vascular Endothelial Growth Factor (VEGF)
3.4. Erythropoietin (EPO)
3.5. Insulin-like Growth Factor (IGF)
3.6. Granulocyte Colony-Stimulating Growth Factor (G-CSF)
3.7. Hepatocyte Growth Factor (HGF)
3.8. Neurotrophic Factors (NFs)

| Growth Factor | Role and Ways of Applications | Sources |
|---|---|---|
| EGF | stimulating the development of epithelium, mainly intestines and limiting the pathogenic bacterial growth, can be supplemented in NEC to protect the intestines of a premature baby | [38,40,41,42,45] |
| HB-EGF | stimulating the proliferation, development, and regeneration of epithelial cells, could potentially be used in the perfusion disorders | [41,47] |
| VEGF | a strong factor in angiogenesis, considered for the prevention of ROP | [47,49,51] |
| EPO | stimulating erytropeiesis, used in the cases of lower RBC levels | [52,53,54] |
| IGF | stimulating the proliferation of stem cells, promoting the survival of cells, reducing or decreasing apoptosis, and the attenuation of inflammations, its high concentration in breast milk has a positive effect not only on the development of the newborn but also on BMI | [38,55,56,57] |
| G-CSF | responsible for proper inflammation response, the maintenance of hematopoiesis, and leukopoiesis, applied in states such as neutropenia or sepsis | [66,67] |
| HGF | responsible for proper hepar formation, as many others, the role of breastfeeding in supporting the final formation of the organs | [58,61] |
| NFs | impact on the nervous system’s final formation, breastfeeding supports proper nervous and mental development | [58,62,64,65] |
4. Hormones
4.1. Sex Steroid Hormones
4.2. Thyroxine and Thyroid-Stimulating Hormone
4.3. Glucocorticoids
4.4. Metabolic Hormones: Leptin, Adiponectin, Ghrelin, Insulin
4.5. Melatonin
| Hormone | Possible Effect on BM | Source |
|---|---|---|
| Estrogens (E1, E2, 23) | Positive correlation with infant developmental score. | [68,70] |
| Phytoestrogens | Impaired immune function and the reduction in weight and adipose tissue. | [68,72] |
| Progesterone | Negative correlation with infant developmental score and the reduction in infant weight. | [70] |
| Testosterone | Neuro- and immunomodulation and the increase of hemoglobin and hematocrit levels. | [74] |
| T4, T3 | The delay of clinical the recognition of hypothyroidism. | [76] |
| GCs (cortisol, cortiosne, corticosterone) | Impact on healthy growth, development, infant temperament, fear reactivity, the improvement of the intestinal microbiome and immune system, and lower levels of sIgA. | [78,79,81,82,83] |
| Leptin | Satiety signal, the impact of energy balance and body weight regulation, and lower body weight. | [88,89] |
| Adiponectin | The enhancement of insulin sensitivity, suppression of inflammation, and lower adiposity. | [93] |
| Ghrelin | Positive correlation with infant growth rate and impact on feeding behavior and body composition. | [89,91] |
| Insulin | Intermediate levels—lower infant weight High levels—higher body mass | [94,95] |
| Melatonin | Improved sleep duration and efficiency, circadian manner of physiological mechanisms, and the improvement of metabolic disturbances. | [80,96,97] |
5. Carbohydrates
6. Lipids
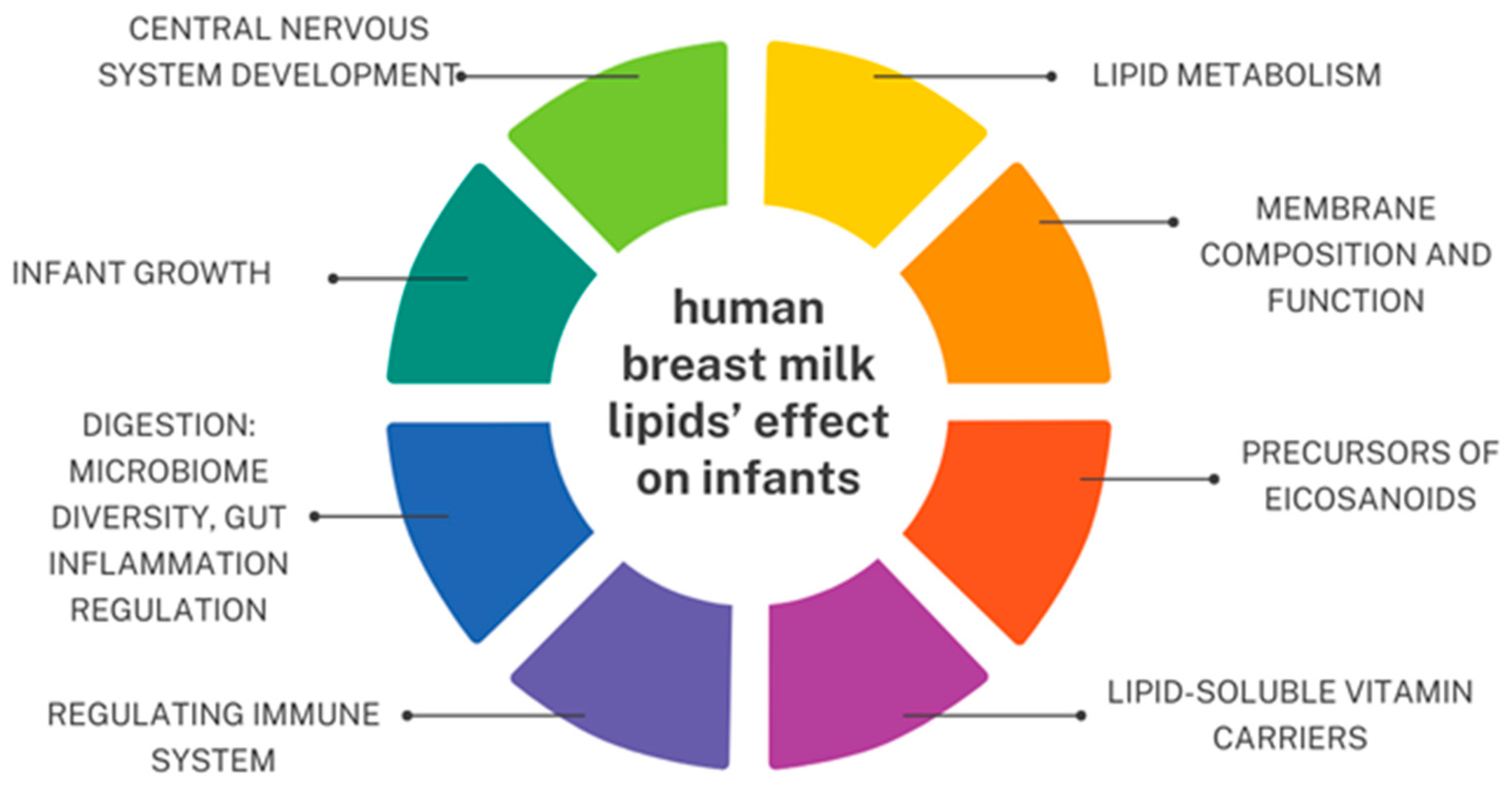
{kind=link}
{kind=link}
{kind=link}
{kind=link}
7. Minerals (Micronutrients)
7.1. Iron
7.2. Zinc

7.3. Calcium
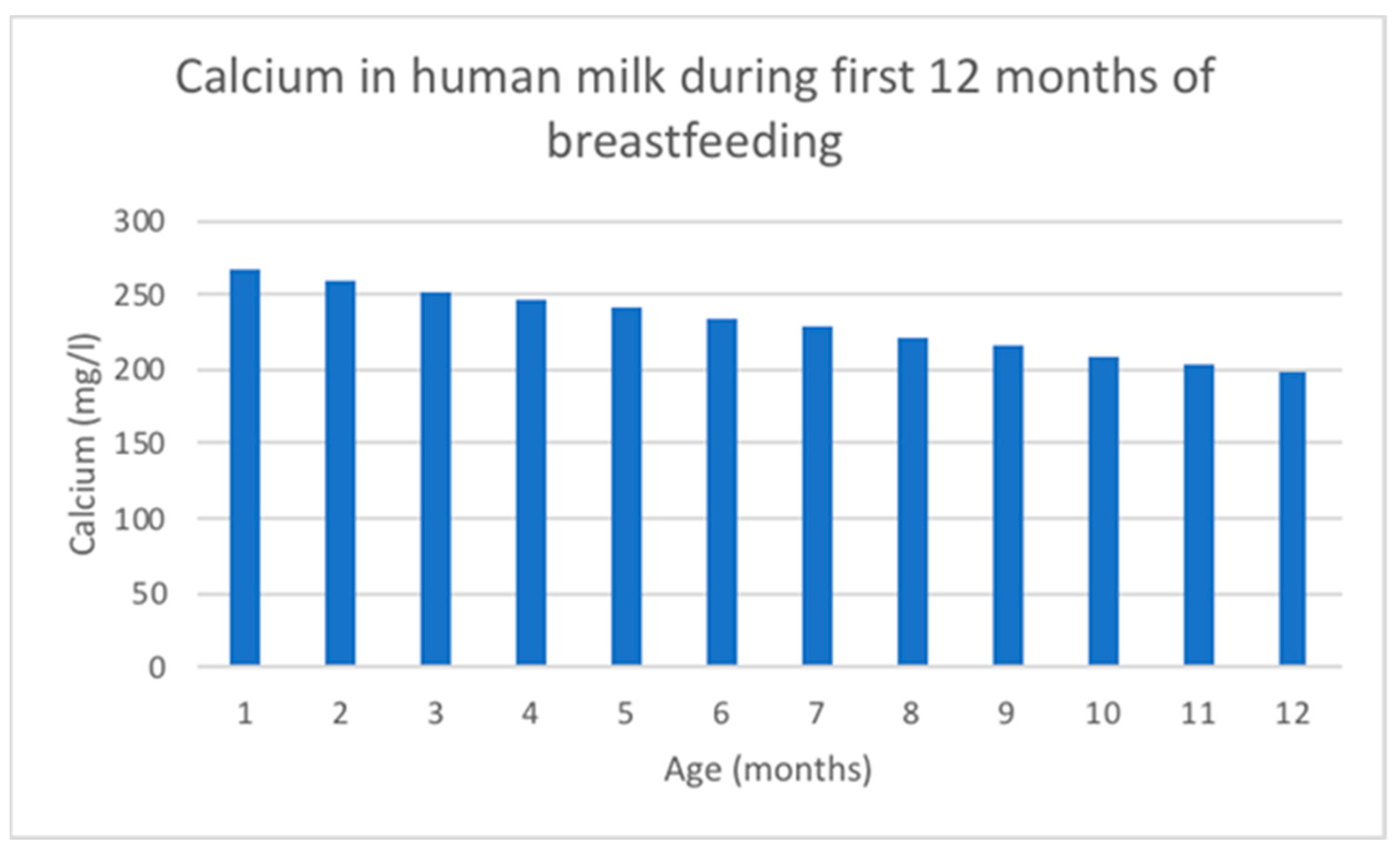
7.4. Copper
7.5. Phosphorus
7.6. Selenium
7.7. Magnesium
7.8. Iodine
7.9. Role of Micronutrients in Child’s Neurodevelopment
| Iron | Zinc | Calcium | Copper | Phosphorus | Selenium | Magnesium | Iodine | |
|---|---|---|---|---|---|---|---|---|
| Function | Metabolic processes, oxygen transportation, and myelination in CNS 2 | Cellular functions; immunity, growth, and development | Bone health and cell signaling | Cellular respiration and iron metabolism; myelination in CNS 2 | Bone mineralization and cell signaling, and forms cell membranes and nucleic acids | Antioxidant protection, thyroid function, immunity, and cholesterol metabolism | Bone structure and metabolic reactions | Growth, nervous system development, and thyroid hormone synthesis |
| Level in BM 1 | Highest in colostrum and decreases over lactation | Stages as follows: high in colostrum; then a rapid decline; plateau phase and further decline | Decreases over lactation (total calcium concentration; ionized form is stable) | Decreases over lactation | High in transitional milk and declines over lactation | High in colostrum and decreases over lactation | Varies between foremilk and hindmilk | Maximum levels in colostrum and decreases over lactation; no difference in values between fore- and hindmilk |
| The influence of maternal factors on the levels | No influence | No significant relationship | Maternal intake, geographic area, and anemia | No influence | No influence | Strongly influenced by mother’s diet | Not observed | Maternal intake and genetic factors |
| Comment | Both in the lipid form and small peptides; newborns have their own reserves | Both whey and fat fraction forms in BM; similar values between fore- and hindmilk; high bioavailability | - | Newborns have their own reserves | Relatively low concentration in BM protects the immature renal system of a newborn | The component of glutathione peroxidase in BM; newborns have their own reserves | A majority is bound to proteins in BM | A majority exists in the form of ionic iodide in BM |
| Recommended supplementation or extra intake for lactating mothers | Usually recommended, but not essential | Increase zinc intake by 50% | A total of 5 servings a day of calcium-rich foods | An amount of +400 μg to RDA 3 | Not needed | An amount of +15 μg to RDA 3 | Not needed | A total of 200 μg of potassium iodide a day |
| Source | [112,114,115,121] | [114,115,116,117,121] | [106,114,118,119,121] | [111,114,121] | [114,119,121] | [114,120] | [114,117] | [114,115,117] |
8. Vitamins
8.1. Riboflavin
8.2. Vitamin B6
8.3. Vitamin B12
8.4. Vitamin C
8.5. Vitamin A
8.6. Vitamin D
8.7. Vitamin E
8.8. Vitamin K
| Vitamin | Main Forms | Function | Recommended Dietary Allowance for Infants (0–6 msc.) | Risk of Inadequacy | Sources |
|---|---|---|---|---|---|
| Riboflavin | free riboflavin and conzymatic form FAD | energy metabolism, and part of co-enzymes FMN and FAD in redox reactions | 0.3 mg | - | [114,117,125] |
| Vitamin B6 | pyridoxal | neurological development, the synthesis of neurotransmitters, immune system regulation, and hemoglobin production | 0.1 mg | - | [111,114,117,125] |
| Vitamin B12 | methylcobalamin | cofactor in DNA synthesis and folate metabolism and crucial for maintaining neurological function and metabolism regulation | 0.4 mcg | the infants of vegan mothers are at risk of deficiency and might need supplementation | [114,115,117,123,125] |
| Vitamin C | ascorbic acid and dehydroascorbic acid | antioxidant, boosts white blood cell activity and antibody production, and promotes the synthesis of interferons | 40 mg | - | [114,115,117,126] |
| Vitamin A | retinyl esters | optimal vision, immune function, growth and psychomotor development, and the prevention of obesity and type 2 diabetes mellitus | 400 mcg RAE * | in developing countries deficient in vitamin A, mothers require supplementation | [114,115,117,121,124,127] |
| Vitamin D | ergocalciferol and cholecalciferol | proper bone mineralization and growth development, and blood pressure and glycemia regulation | 10 mcg; 400 IU | the concentrations in breast milk are insufficient to meet the daily dietary needs of an exclusively breastfed infant. Supplementation is necessary | [12,67,114,115,117,118,128,129,130] |
| Vitamin E | α-tocopherol | antioxidant and immune system regulation | 4 mg | - | [114,117,126] |
| Vitamin K | phylloquinone and menaquinone-4 | cofactor for the synthesis of coagulation factors II, VII, IX, X, and proteins C and S | 2.0 mcg | poor placenta traversal elevates the risk of vitamin K deficiency bleeding (VKDB). To prevent VKDB, a single, intramuscular dose of vitamin K1 at birth is recommended | [114,115,117,131,132] |
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Victora, C.G.; Bahl, R.; Barros, A.J.D.; França, G.V.A.; Horton, S.; Krasevec, J.; Murch, S.; Sankar, M.J.; Walker, N.; Rollins, N.C.; et al. Breastfeeding in the 21st Century: Epidemiology, Mechanisms, and Lifelong Effect. Lancet 2016, 387, 475–490. [Google Scholar] [CrossRef] [PubMed]
- Karamantziani, T.; Pouliakis, A.; Xanthos, T.; Ekmektzoglou, K.; Paliatsiou, S.; Sokou, R.; Iacovidou, N. The Effect of Oral Iron Supplementation/Fortification on the Gut Microbiota in Infancy: A Systematic Review and Meta-Analysis. Children 2024, 11, 231. [Google Scholar] [CrossRef] [PubMed]
- Rollins, N.C.; Bhandari, N.; Hajeebhoy, N.; Horton, S.; Lutter, C.K.; Martines, J.C.; Piwoz, E.G.; Richter, L.M.; Victora, C.G. Why Invest, and What It Will Take to Improve Breastfeeding Practices? Lancet 2016, 387, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.R.; Ling, P.R.; Blackburn, G.L. Review of Infant Feeding: Key Features of Breast Milk and Infant Formula. Nutrients 2016, 8, 279. [Google Scholar] [CrossRef]
- Phukan, D.; Ranjan, M.; Dwivedi, L.K. Impact of Timing of Breastfeeding Initiation on Neonatal Mortality in India. Int. Breastfeed. J. 2018, 13, 27. [Google Scholar] [CrossRef] [PubMed]
- Bień, A.; Kulesza-Brończyk, B.; Przestrzelska, M.; Iwanowicz-Palus, G.; Ćwiek, D. The Attitudes of Polish Women towards Breastfeeding Based on the Iowa Infant Feeding Attitude Scale (IIFAS). Nutrients 2021, 13, 4338. [Google Scholar] [CrossRef] [PubMed]
- Andrews, T. Modern Taboos and Moral Regulations: Mother’s Milk in the Symbolic Order. Sociology 2021, 56, 55–71. [Google Scholar] [CrossRef]
- Jackson, J.E.; Hallam, J.L. “It’s Quite a Taboo Subject”: An Investigation of Mother’s Experiences of Breastfeeding beyond Infancy and the Challenges They Face. Women Health 2021, 61, 572–580. [Google Scholar] [CrossRef]
- Theurich, M.A.; Davanzo, R.; Busck-Rasmussen, M.; Díaz-Gómez, N.M.; Brennan, C.; Kylberg, E.; Bærug, A.; McHugh, L.; Weikert, C.; Abraham, K.; et al. Breastfeeding Rates and Programs in Europe: A Survey of 11 National Breastfeeding Committees and Representatives. J. Pediatr. Gastroenterol. Nutr. 2019, 68, 400–407. [Google Scholar] [CrossRef]
- Segura-Pérez, S.; Hromi-Fiedler, A.; Adnew, M.; Nyhan, K.; Pérez-Escamilla, R. Impact of Breastfeeding Interventions among United States Minority Women on Breastfeeding Outcomes: A Systematic Review. Int. J. Equity Health 2021, 20, 72. [Google Scholar] [CrossRef]
- Palmeira, P.; Carneiro-Sampaio, M. Immunology of Breast Milk. Rev. Assoc. Med. Bras. 2016, 62, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Yi, D.Y. Components of Human Breast Milk: From Macronutrient to Microbiome and MicroRNA. Clin. Exp. Pediatr. 2020, 63, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Thai, J.D.; Gregory, K.E. Bioactive Factors in Human Breast Milk Attenuate Intestinal Inflammation during Early Life. Nutrients 2020, 12, 581. [Google Scholar] [CrossRef] [PubMed]
- Langel, S.N.; Wang, Q.; Vlasova, A.N.; Saif, L.J. Host Factors Affecting Generation of Immunity against Porcine Epidemic Diarrhea Virus in Pregnant and Lactating Swine and Passive Protection of Neonates. Pathogens 2020, 9, 130. [Google Scholar] [CrossRef] [PubMed]
- Selvamani, S.; Dailin, D.J.; Gupta, V.K.; Wahid, M.; Keat, H.C.; Natasya, K.H.; Malek, R.A.; Haque, S.; Sayyed, R.Z.; Abomoelak, B.; et al. An Insight into Probiotics Bio-Route: Translocation from the Mother’s Gut to the Mammary Gland. Appl. Sci. 2021, 11, 7247. [Google Scholar] [CrossRef]
- Kompaneets, I.Y.; Ermakov, E.A.; Sedykh, S.E.; Buneva, V.N.; Nevinsky, G.A. Secretory Immunoglobulin A from Human Milk Hydrolyzes MicroRNA. J. Dairy Sci. 2020, 103, 6782–6797. [Google Scholar] [CrossRef] [PubMed]
- Atyeo, C.; Alter, G. The Multifaceted Roles of Breast Milk Antibodies. Cell 2021, 184, 1486–1499. [Google Scholar] [CrossRef] [PubMed]
- Dimitrovska-Ivanova, M.; Zisovska, E. Impact of Breast Milk Secretory Immunoglobulin a on Infants Acute Gastroenteritis. Open Access Maced. J. Med. Sci. 2020, 8, 897–901. [Google Scholar] [CrossRef]
- Massi, M.N.; Febriani, A.D.B.; Hatta, M.; Karuniawati, A.; Rauf, S.; Wahyuni, S.; Hamid, F.; Alasiry, E.; Patellongi, I.; Permatasari, T.A.E. The Role of Exclusive Breastfeeding on SIgA and Lactoferrin Levels in Toddlers Suffering from Acute Respiratory Infection: A Cross-Sectional Study. Ann. Med. Surg. 2022, 77, 103644. [Google Scholar] [CrossRef] [PubMed]
- Janzon, A.; Goodrich, J.K.; Koren, O.; Waters, J.L.; Ley, R.E. Interactions between the Gut Microbiome and Mucosal Immunoglobulins A, M, and G in the Developing Infant Gut. mSystems 2019, 4. [Google Scholar] [CrossRef]
- Yi, D.Y.; Kim, S.Y. Human Breast Milk Composition and Function in Human Health: From Nutritional Components to Microbiome and MicroRNAs. Nutrients 2021, 13, 3094. [Google Scholar] [CrossRef] [PubMed]
- Venema, K. Intestinal Fermentation of Lactose and Prebiotic Lactose Derivatives, Including Human Milk Oligosaccharides. Int. Dairy J. 2012, 22, 123–140. [Google Scholar] [CrossRef]
- Yang, Z.; Jiang, R.; Chen, Q.; Wang, J.; Duan, Y.; Pang, X.; Jiang, S.; Bi, Y.; Zhang, H.; Lönnerdal, B.; et al. Concentration of Lactoferrin in Human Milk and Its Variation during Lactation in Different Chinese Populations. Nutrients 2018, 10, 1235. [Google Scholar] [CrossRef] [PubMed]
- Czosnykowska-Łukacka, M.; Orczyk-Pawiłowicz, M.; Broers, B.; Królak-Olejnik, B. Lactoferrin in Human Milk of Prolonged Lactation. Nutrients 2019, 11, 2350. [Google Scholar] [CrossRef] [PubMed]
- Parra-Saavedra, K.J.; Macias-Lamas, A.M.; Silva-Jara, J.M.; Solís-Pacheco, J.R.; Ortiz-Lazareno, P.C.; Aguilar-Uscanga, B.R. Human Lactoferrin from Breast Milk: Characterization by HPLC and Its in Vitro Antibiofilm Performance. J. Food Sci. Technol. 2022, 59, 4907–4914. [Google Scholar] [CrossRef] [PubMed]
- Minami, J.; Odamaki, T.; Hashikura, N.; Abe, F.; Xiao, J.Z. Lysozyme in Breast Milk Is a Selection Factor for Bifidobacterial Colonisation in the Infant Intestine. Benef. Microbes 2016, 7, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Wedekind, S.I.S.; Shenker, N.S. Antiviral Properties of Human Milk. Microorganisms 2021, 9, 715. [Google Scholar] [CrossRef] [PubMed]
- Newburg, D.S.; Peterson, J.A.; Ruiz-Palacios, G.M.; Matson, D.O.; Morrow, A.L.; Shults, J.; De Lourdes Guerrero, M.; Chaturvedi, P.; Newburg, S.O.; Scallan, C.D.; et al. Role of Human-Milk Lactadherin in Protection against Symptomatic Rotavirus Infection. Lancet 1998, 351, 1160–1164. [Google Scholar] [CrossRef] [PubMed]
- Layman, D.K.; Lönnerdal, B.; Fernstrom, J.D. Applications for α-Lactalbumin in Human Nutrition. Nutr. Rev. 2018, 76, 444–460. [Google Scholar] [CrossRef] [PubMed]
- Lönnerdal, B.; Lien, E.L. Nutritional and Physiologic Significance of Alpha-Lactalbumin in Infants. Nutr. Rev. 2003, 61, 295–305. [Google Scholar] [CrossRef]
- Ng, T.B.; Cheung, R.C.F.; Wong, J.H.; Wang, Y.; Ip, D.T.M.; Wan, D.C.C.; Xia, J. Antiviral Activities of Whey Proteins. Appl. Microbiol. Biotechnol. 2015, 99, 6997–7008. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Dingess, K.A. The Functional Power of the Human Milk Proteome. Nutrients 2019, 11, 1834. [Google Scholar] [CrossRef]
- Kielbasa, A.; Gadzala-Kopciuch, R.; Buszewski, B. Cytokines-Biogenesis and Their Role in Human Breast Milk and Determination. Int. J. Mol. Sci. 2021, 22, 6238. [Google Scholar] [CrossRef] [PubMed]
- Ramiro-Cortijo, D.; Singh, P.; Liu, Y.; Medina-Morales, E.; Yakah, W.; Freedman, S.D.; Martin, C.R. Breast Milk Lipids and Fatty Acids in Regulating Neonatal Intestinal Development and Protecting against Intestinal Injury. Nutrients 2020, 12, 534. [Google Scholar] [CrossRef] [PubMed]
- Saso, A.; Blyuss, O.; Munblit, D.; Faal, A.; Moore, S.E.; Le Doare, K. Breast Milk Cytokines and Early Growth in Gambian Infants. Front. Pediatr. 2019, 6, 414. [Google Scholar] [CrossRef] [PubMed]
- Dawod, B.; Marshall, J.S. Cytokines and Soluble Receptors in Breast Milk as Enhancers of Oral Tolerance Development. Front. Immunol. 2019, 10, 432191. [Google Scholar] [CrossRef]
- Polat, A.; Tunc, T.; Erdem, G.; Yerebasmaz, N.; Tas, A.; Beken, S.; Basbozkurt, G.; Saldir, M.; Zenciroglu, A.; Yaman, H. Interleukin-8 and Its Receptors in Human Milk from Mothers of Full-Term and Premature Infants. Breastfeed. Med. 2016, 11, 247–251. [Google Scholar] [CrossRef]
- Sánchez, C.; Franco, L.; Regal, P.; Lamas, A.; Cepeda, A.; Fente, C. Breast Milk: A Source of Functional Compounds with Potential Application in Nutrition and Therapy. Nutrients 2021, 13, 1026. [Google Scholar] [CrossRef]
- Ballard, O.; Morrow, A.L. Human Milk Composition: Nutrients and Bioactive Factors. Pediatr. Clin. N. Am. 2013, 60, 49–74. [Google Scholar] [CrossRef] [PubMed]
- Knoop, K.A.; Coughlin, P.E.; Floyd, A.N.; Malick Ndao, I.; Hall-Moore, C.; Shaikh, N.; Gasparrini, A.J.; Rusconi, B.; Escobedo, M.; Good, M.; et al. Maternal Activation of the EGFR Prevents Translocation of Gut-Residing Pathogenic Escherichia coli in a Model of Late-Onset Neonatal Sepsis. Proc. Natl. Acad. Sci. USA 2020, 117, 7941–7949. [Google Scholar] [CrossRef]
- York, D.J.; Smazal, A.L.; Robinson, D.T.; De Plaen, I.G. Human Milk Growth Factors and Their Role in NEC Prevention: A Narrative Review. Nutrients 2021, 13, 3751. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, B.; Fituch, C.C.; Williams, C.S.; Hurst, N.M.; Schanler, R.J. Increased Epidermal Growth Factor Levels in Human Milk of Mothers with Extremely Premature Infants. Pediatr. Res. 2003, 54, 15–19. [Google Scholar] [CrossRef]
- Al-Nafussi, A.I.; Wright, N.A. The Effect of Epidermal Growth Factor (EGF) on Cell Proliferation of the Gastrointestinal Mucosa in Rodents. Virchows Arch. B 1982, 40, 63–69. [Google Scholar] [CrossRef]
- Basuroy, S.; Seth, A.; Elias, B.; Naren, A.P.; Rao, R. MAPK Interacts with Occludin and Mediates EGF-Induced Prevention of Tight Junction Disruption by Hydrogen Peroxide. Biochem. J. 2006, 393, 69–77. [Google Scholar] [CrossRef]
- Battersby, C.; Santhalingam, T.; Costeloe, K.; Modi, N. Incidence of Neonatal Necrotising Enterocolitis in High-Income Countries: A Systematic Review. Arch. Dis. Child. Fetal Neonatal Ed. 2018, 103, F182–F189. [Google Scholar] [CrossRef]
- Michalsky, M.P.; Lara-Marquez, M.; Chun, L.; Besner, G.E. Heparin-Binding EGF-like Growth Factor Is Present in Human Amniotic Fluid and Breast Milk. J. Pediatr. Surg. 2002, 37, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kobata, R.; Tsukahara, H.; Ohshima, Y.; Ohta, N.; Tokuriki, S.; Tamura, S.; Mayumi, M. High Levels of Growth Factors in Human Breast Milk. Early Hum. Dev. 2008, 84, 67–69. [Google Scholar] [CrossRef] [PubMed]
- Loui, A.; Eilers, E.; Strauss, E.; Pohl-Schickinger, A.; Obladen, M.; Koehne, P. Vascular Endothelial Growth Factor (VEGF) and Soluble VEGF Receptor 1 (SFlt-1) Levels in Early and Mature Human Milk from Mothers of Preterm versus Term Infants. J. Hum. Lact. 2012, 28, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N.; Adamis, A.P. Ten Years of Anti-Vascular Endothelial Growth Factor Therapy. Nat. Rev. Drug Discov. 2016, 15, 385–403. [Google Scholar] [CrossRef] [PubMed]
- Takeshita, K.; Satoh, M.; Ii, M.; Silver, M.; Limbourg, F.P.; Mukai, Y.; Rikitake, Y.; Radtke, F.; Gridley, T.; Losordo, D.W.; et al. Critical Role of Endothelial Notch1 Signaling in Postnatal Angiogenesis. Circ. Res. 2007, 100, 70–78. [Google Scholar] [CrossRef]
- DiBiasie, A.M. Evidence-Based Review of Retinopathy of Prematurity Prevention in VLBW and ELBW Infants. Neonatal Netw. 2006, 25, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Juul, S.E.; Zhao, Y.; Dame, J.B.; Du, Y.; Hutson, A.D.; Christensen, R.D. Origin and Fate of Erythropoietin in Human Milk. Pediatr. Res. 2000, 48, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Carbonell-Estrany, X.; Figueras-Aloy, J.; Alvarez, E. Erythropoietin and Prematurity—Where Do We Stand? J. Perinat. Med. 2005, 33, 277–286. [Google Scholar] [CrossRef]
- Saleh, M.I.; Nalbant, D.; Widness, J.A.; Veng-Pedersen, P. Population Pharmacodynamic Analysis of Erythropoiesis in Preterm Infants for Determining the Anemia Treatment Potential of Erythropoietin. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R772–R781. [Google Scholar] [CrossRef] [PubMed]
- Milsom, S.R.; Blum, W.F.; Gunn, A.J. Temporal Changes in Insulin-like Growth Factors I and II and in Insulin-like Growth Factor Binding Proteins 1, 2, and 3 in Human Milk. Horm. Res. 2008, 69, 307–311. [Google Scholar] [CrossRef]
- Elmlinger, M.W.; Hochhaus, F.; Loui, A.; Frommer, K.W.; Obladen, M.; Ranke, M.B. Insulin-like Growth Factors and Binding Proteins in Early Milk from Mothers of Preterm and Term Infants. Horm. Res. 2007, 68, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Galante, L.; Pundir, S.; Lagström, H.; Rautava, S.; Reynolds, C.M.; Milan, A.M.; Cameron-Smith, D.; Vickers, M.H. Growth Factor Concentrations in Human Milk Are Associated with Infant Weight and BMI from Birth to 5 Years. Front. Nutr. 2020, 7, 110. [Google Scholar] [CrossRef]
- Gila-Diaz, A.; Arribas, S.M.; Algara, A.; Martín-Cabrejas, M.A.; López de Pablo, Á.L.; Sáenz de Pipaón, M.; Ramiro-Cortijo, D. A Review of Bioactive Factors in Human Breastmilk: A Focus on Prematurity. Nutrients 2019, 11, 1307. [Google Scholar] [CrossRef]
- Lee, J.A.; Sauer, B.; Tuminski, W.; Cheong, J.; Fitz-Henley, J.; Mayers, M.; Ezuma-Igwe, C.; Arnold, C.; Hornik, C.P.; Clark, R.H.; et al. Effectiveness of Granulocyte Colony-Stimulating Factor in Hospitalized Infants with Neutropenia. Am. J. Perinatol. 2017, 34, 458–464. [Google Scholar] [CrossRef] [PubMed]
- Ramiro-Cortijo, D.; Herranz Carrillo, G.; Singh, P.; Rebollo-Hernanz, M.; Rodríguez-Rodríguez, P.; Ruvira, S.; Martín-Trueba, M.; Martin, C.R.; Arribas, S.M. Maternal and Neonatal Factors Modulating Breast Milk Cytokines in the First Month of Lactation. Antioxidants 2023, 12, 996. [Google Scholar] [CrossRef]
- Panopoulos, A.D.; Watowich, S.S. Granulocyte Colony-Stimulating Factor: Molecular Mechanisms of Action during Steady State and “emergency” Hematopoiesis. Cytokine 2008, 42, 277–288. [Google Scholar] [CrossRef]
- Ismail, A.M.; Babers, G.M.; El Rehany, M.A. Brain-Derived Neurotrophic Factor in Sera of Breastfed Epileptic Infants and in Breastmilk of Their Mothers. Breastfeed. Med. 2015, 10, 277–282. [Google Scholar] [CrossRef]
- Fichter, M.; Klotz, M.; Hirschberg, D.L.; Waldura, B.; Schofer, O.; Ehnert, S.; Schwarz, L.K.; Ginneken, C.V.; Schäfer, K.H. Breast Milk Contains Relevant Neurotrophic Factors and Cytokines for Enteric Nervous System Development. Mol. Nutr. Food Res. 2011, 55, 1592–1596. [Google Scholar] [CrossRef]
- Ratsika, A.; Codagnone, M.C.; O’mahony, S.; Stanton, C.; Cryan, J.F. Priming for Life: Early Life Nutrition and the Microbiota-Gut-Brain Axis. Nutrients 2021, 13, 423. [Google Scholar] [CrossRef]
- Sanchez, M.P.; Silos-Santiago, I.; Frisen, J.; He, B.; Lira, S.A.; Barbacid, M. Renal Agenesis and the Absence of Enteric Neurons in Mice Lacking GDNF. Nature 1996, 382, 70–73. [Google Scholar] [CrossRef]
- Calhoun, D.A.; Lunøe, M.; Du, Y.; Staba, S.L.; Christensen, R.D. Concentrations of Granulocyte Colony-Stimulating Factor in Human Milk after in Vitro Simulations of Digestion. Pediatr. Res. 1999, 46, 767. [Google Scholar] [CrossRef]
- Chang, S.W.; Lee, H.C. Vitamin D and Health—The Missing Vitamin in Humans. Pediatr. Neonatol. 2019, 60, 237–244. [Google Scholar] [CrossRef]
- Jin, X.; Perrella, S.L.; Lai, C.T.; Taylor, N.L.; Geddes, D.T. Oestrogens and Progesterone in Human Milk and Their Effects on Infant Health Outcomes: A Narrative Review. Food Chem. 2023, 424, 136375. [Google Scholar] [CrossRef]
- Fuentes, N.; Silveyra, P. Estrogen Receptor Signaling Mechanisms. Adv. Protein Chem. Struct. Biol. 2019, 116, 135–170. [Google Scholar] [CrossRef]
- Messripour, M.; Forooghi-Abary, A.; Dashti-Ardakani, F. Association between Sex Hormones in Human Breast Milk and Infant Growth and Development. Arch. Iran. Med. 2002, 5, 166–169. [Google Scholar]
- Landete, J.M.; Arqués, J.; Medina, M.; Gaya, P.; de Las Rivas, B.D.; Muñoz, R. Bioactivation of Phytoestrogens: Intestinal Bacteria and Health. Crit. Rev. Food Sci. Nutr. 2016, 56, 1826–1843. [Google Scholar] [CrossRef]
- Cederroth, C.R.; Vinciguerra, M.; Kühne, F.; Madani, R.; Doerge, D.R.; Visser, T.J.; Foti, M.; Rohner-Jeanrenaud, F.; Vassalli, J.D.; Nef, S. A Phytoestrogen-Rich Diet Increases Energy Expenditure and Decreases Adiposity in Mice. Environ. Health Perspect. 2007, 115, 1467–1473. [Google Scholar] [CrossRef]
- Knight, C.H. An Endocrine Hypothesis to Explain Obesity-Related Lactation Insufficiency in Breastfeeding Mothers. J. Dairy Res. 2020, 87, 78–81. [Google Scholar] [CrossRef]
- Vass, R.A.; Bell, E.F.; Roghair, R.D.; Kiss, G.; Funke, S.; Bokor, S.; Molnár, D.; Miseta, A.; Bódis, J.; Kovács, K.; et al. Insulin, Testosterone, and Albumin in Term and Preterm Breast Milk, Donor Milk, and Infant Formula. Nutrients 2023, 15, 1476. [Google Scholar] [CrossRef]
- Vass, R.A.; Kiss, G.; Bell, E.F.; Miseta, A.; Bódis, J.; Funke, S.; Bokor, S.; Molnár, D.; Kósa, B.; Kiss, A.A.; et al. Thyroxine and Thyroid-Stimulating Hormone in Own Mother’s Milk, Donor Milk, and Infant Formula. Life 2022, 12, 584. [Google Scholar] [CrossRef]
- Sack, J.; Amado, O.; Lunenfeld, B. Thyroxine Concentration in Human Milk. J. Clin. Endocrinol. Metab. 1977, 45, 171–173. [Google Scholar] [CrossRef]
- Mizuta, H.; Amino, N.; Ichihara, K.; Harada, T.; Nose, O.; Tanizawa, O.; Miyai, K. Thyroid Hormones in Human Milk and Their Influence on Thyroid Function of Breast-Fed Babies. Pediatr. Res. 1983, 17, 468–471. [Google Scholar] [CrossRef]
- Zielinska-Pukos, M.A.; Bryś, J.; Kucharz, N.; Chrobak, A.; Wesolowska, A.; Grabowicz-Chądrzyńska, I.; Hamulka, J. Factors Influencing Cortisol Concentrations in Breastmilk and Its Associations with Breastmilk Composition and Infant Development in the First Six Months of Lactation. Int. J. Environ. Res. Public Health 2022, 19, 14809. [Google Scholar] [CrossRef]
- Vacaru, S.V.; Brett, B.E.; Eckermann, H.; de Weerth, C. Determinants of Maternal Breast Milk Cortisol Increase: Examining Dispositional and Situational Factors. Psychoneuroendocrinology 2023, 158, 106385. [Google Scholar] [CrossRef]
- Italianer, M.F.; Naninck, E.F.G.; Roelants, J.A.; van der Horst, G.T.J.; Reiss, I.K.M.; van Goudoever, J.B.; Joosten, K.F.M.; Chaves, I.; Vermeulen, M.J. Circadian Variation in Human Milk Composition, a Systematic Review. Nutrients 2020, 12, 2328. [Google Scholar] [CrossRef]
- Grey, K.R.; Davis, E.P.; Sandman, C.A.; Glynn, L.M. Human Milk Cortisol Is Associated with Infant Temperament. Psychoneuroendocrinology 2013, 38, 1178–1185. [Google Scholar] [CrossRef]
- Nolvi, S.; Uusitupa, H.M.; Bridgett, D.J.; Pesonen, H.; Aatsinki, A.K.; Kataja, E.L.; Korja, R.; Karlsson, H.; Karlsson, L. Human Milk Cortisol Concentration Predicts Experimentally Induced Infant Fear Reactivity: Moderation by Infant Sex. Dev. Sci. 2018, 21, e12625. [Google Scholar] [CrossRef]
- Anyim, R.; Li, S.; Armstrong, D.; Spathis, R.; Wander, K. Associations between Milk Cortisol and Activity of the Immune System of Milk. Am. J. Hum. Biol. 2023, 35, e23960. [Google Scholar] [CrossRef]
- De Graaf, C.; Blom, W.A.M.; Smeets, P.A.M.; Stafleu, A.; Hendriks, H.F.J. Biomarkers of Satiation and Satiety. Am. J. Clin. Nutr. 2004, 79, 946–961. [Google Scholar] [CrossRef]
- Smith-Kirwin, S.M.; O’Connor, D.M.; Johnston, J.; de Lancy, E.; Hassink, S.G.; Funanage, V.L. Leptin Expression in Human Mammary Epithelial Cells and Breast Milk. J. Clin. Endocrinol. Metab. 1998, 83, 1810. [Google Scholar] [CrossRef]
- Bonnet, M.; Delavaud, C.; Laud, K.; Gourdou, I.; Leroux, C.; Djiane, J.; Chilliard, Y. Mammary Leptin Synthesis, Milk Leptin and Their Putative Physiological Roles. Reprod. Nutr. Dev. 2002, 42, 399–413. [Google Scholar] [CrossRef]
- Bielicki, J.; Huch, R.; von Mandach, U. Time-Course of Leptin Levels in Term and Preterm Human Milk. Eur. J. Endocrinol. 2004, 151, 271–276. [Google Scholar] [CrossRef]
- Savino, F.; Liguori, S.A.; Fissore, M.F.; Oggero, R. Breast Milk Hormones and Their Protective Effect on Obesity. Int. J. Pediatr. Endocrinol. 2009, 2009, 327505. [Google Scholar] [CrossRef]
- Savino, F.; Fissore, M.F.; Liguori, S.A.; Oggero, R. Can Hormones Contained in Mothers’ Milk Account for the Beneficial Effect of Breast-Feeding on Obesity in Children? Clin. Endocrinol. 2009, 71, 757–765. [Google Scholar] [CrossRef]
- Mazzocchi, A.; Giannì, M.L.; Morniroli, D.; Leone, L.; Roggero, P.; Agostoni, C.; De Cosmi, V.; Mosca, F. Hormones in Breast Milk and Effect on Infants’ Growth: A Systematic Review. Nutrients 2019, 11, 1845. [Google Scholar] [CrossRef]
- Weyer, C.; Funahashi, T.; Tanaka, S.; Hotta, K.; Matsuzawa, Y.; Pratley, R.E.; Tataranni, P.A. Hypoadiponectinemia in Obesity and Type 2 Diabetes: Close Association with Insulin Resistance and Hyperinsulinemia. J. Clin. Endocrinol. Metab. 2001, 86, 1930–1935. [Google Scholar] [CrossRef]
- Suwaydi, M.A.; Gridneva, Z.; Perrella, S.L.; Wlodek, M.E.; Lai, C.T.; Geddes, D.T. Human Milk Metabolic Hormones: Analytical Methods and Current Understanding. Int. J. Mol. Sci. 2021, 22, 8708. [Google Scholar] [CrossRef]
- Newburg, D.S.; Woo, J.G.; Morrow, A.L. Characteristics and Potential Functions of Human Milk Adiponectin. J. Pediatr. 2010, 156, S41–S46. [Google Scholar] [CrossRef]
- Fields, D.A.; George, B.; Williams, M.; Whitaker, K.; Allison, D.B.; Teague, A.; Demerath, E.W. Associations between Human Breast Milk Hormones and Adipocytokines and Infant Growth and Body Composition in the First 6 Months of Life. Pediatr. Obes. 2017, 12 (Suppl. 1), 78–85. [Google Scholar] [CrossRef]
- Chan, D.; Goruk, S.; Becker, A.B.; Subbarao, P.; Mandhane, P.J.; Turvey, S.E.; Lefebvre, D.; Sears, M.R.; Field, C.J.; Azad, M.B. Adiponectin, Leptin and Insulin in Breast Milk: Associations with Maternal Characteristics and Infant Body Composition in the First Year of Life. Int. J. Obes. 2018, 42, 36–43. [Google Scholar] [CrossRef]
- Qin, Y.; Shi, W.; Zhuang, J.; Liu, Y.; Tang, L.; Bu, J.; Sun, J.; Bei, F. Variations in Melatonin Levels in Preterm and Term Human Breast Milk during the First Month after Delivery. Sci. Rep. 2019, 9, 17984. [Google Scholar] [CrossRef]
- Gombert, M.; Codoñer-Franch, P. Melatonin in Early Nutrition: Long-Term Effects on Cardiovascular System. Int. J. Mol. Sci. 2021, 22, 6809. [Google Scholar] [CrossRef]
- Picciano, M.F. Nutrient Composition of Human Milk. Pediatr. Clin. N. Am. 2001, 48, 53–67. [Google Scholar] [CrossRef]
- Zhang, P.; Zhu, Y.; Li, Z.; Zhang, W.; Mu, W. Recent Advances on Lacto- N-Neotetraose, a Commercially Added Human Milk Oligosaccharide in Infant Formula. J. Agric. Food Chem. 2022, 70, 4534–4547. [Google Scholar] [CrossRef]
- Ojo-okunola, A.; Cacciatore, S.; Nicol, M.P.; du Toit, E. The Determinants of the Human Milk Metabolome and Its Role in Infant Health. Metabolites 2020, 10, 77. [Google Scholar] [CrossRef]
- Gridneva, Z.; Rea, A.; Tie, W.J.; Lai, C.T.; Kugananthan, S.; Ward, L.C.; Murray, K.; Hartmann, P.E.; Geddes, D.T. Carbohydrates in Human Milk and Body Composition of Term Infants during the First 12 Months of Lactation. Nutrients 2019, 11, 1472. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Akkerman, R.; Kong, C.; Walvoort, M.T.C.; de Vos, P. More than Sugar in the Milk: Human Milk Oligosaccharides as Essential Bioactive Molecules in Breast Milk and Current Insight in Beneficial Effects. Crit. Rev. Food Sci. Nutr. 2021, 61, 1184–1200. [Google Scholar] [CrossRef] [PubMed]
- Kiely, L.J.; Busca, K.; Lane, J.A.; Van Sinderen, D.; Hickey, R.M. Molecular Strategies for the Utilisation of Human Milk Oligosaccharides by Infant Gut-Associated Bacteria. FEMS Microbiol. Rev. 2023, 47, fuad056. [Google Scholar] [CrossRef] [PubMed]
- Walsh, C.; Lane, J.A.; van Sinderen, D.; Hickey, R.M. Human Milk Oligosaccharides: Shaping the Infant Gut Microbiota and Supporting Health. J. Funct. Foods 2020, 72, 104074. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Li, T.; Xie, J.; Zhang, D.; Pi, C.; Zhou, L.; Yang, W. Gold Standard for Nutrition: A Review of Human Milk Oligosaccharide and Its Effects on Infant Gut Microbiota. Microb. Cell Factories 2021, 20, 108. [Google Scholar] [CrossRef] [PubMed]
- Bzikowska-Jura, A.; Czerwonogrodzka-Senczyna, A.; Olędzka, G.; Szostak-Węgierek, D.; Weker, H.; Wesołowska, A. Maternal Nutrition and Body Composition During Breastfeeding: Association with Human Milk Composition. Nutrients 2018, 10, 1379. [Google Scholar] [CrossRef] [PubMed]
- Koletzko, B. Human Milk Lipids. Ann. Nutr. Metab. 2016, 69, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Cheng, J.; Sun, L.; Li, X.; Liu, L.; Ma, Q.; Ma, C.; Jia, Z.; Zhang, L.; Zhang, Q.; et al. Stabilization of Human Milk Fat Analog Emulsions Using Milk Fat Globule Membrane Material–Coated Lipid Droplets: Structural and Physical Properties. LWT 2022, 171, 114154. [Google Scholar] [CrossRef]
- Bobiński, R.; Bobińska, J. Fatty Acids of Human Milk—A Review. Int. J. Vitam. Nutr. Res. 2020, 92, 280–291. [Google Scholar] [CrossRef]
- Schipper, L.; van Dijk, G.; van der Beek, E.M. Milk Lipid Composition and Structure; The Relevance for Infant Brain Development. OCL 2020, 27, 5. [Google Scholar] [CrossRef]
- Lockyer, F.; McCann, S.; Moore, S.E. Breast Milk Micronutrients and Infant Neurodevelopmental Outcomes: A Systematic Review. Nutrients 2021, 13, 3848. [Google Scholar] [CrossRef] [PubMed]
- Georgieff, M.K.; Ramel, S.E.; Cusick, S.E. Nutritional Influences on Brain Development. Acta Paediatr. 2018, 107, 1310–1321. [Google Scholar] [CrossRef] [PubMed]
- Cusick, S.E.; Georgieff, M.K. The Role of Nutrition in Brain Development: The Golden Opportunity of the “First 1000 Days”. J. Pediatr. 2016, 175, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Dror, D.K.; Allen, L.H. Overview of Nutrients in Human Milk. Adv. Nutr. 2018, 9, 278S–294S. [Google Scholar] [CrossRef] [PubMed]
- Drugs and Lactation Database (LactMed®); National Institute of Child Health and Human Development: Bethesda, MD, USA, 2006.
- Savarino, G.; Corsello, A.; Corsello, G. Macronutrient Balance and Micronutrient Amounts through Growth and Development. Ital. J. Pediatr. 2021, 47, 109. [Google Scholar] [CrossRef] [PubMed]
- Hampel, D.; Dror, D.K.; Allen, L.H. Micronutrients in Human Milk: Analytical Methods. Adv. Nutr. 2018, 9, 313S–331S. [Google Scholar] [CrossRef] [PubMed]
- Rios-Leyvraz, M.; Yao, Q. Calcium, Zinc, and Vitamin D in Breast Milk: A Systematic Review and Meta-Analysis. Int. Breastfeed. J. 2023, 18, 27. [Google Scholar] [CrossRef] [PubMed]
- Bzikowska-Jura, A.; Wesołowska, A.; Sobieraj, P.; Michalska-Kacymirow, M.; Bulska, E.; Starcevic, I. Maternal Diet during Breastfeeding in Correlation to Calcium and Phosphorus Concentrations in Human Milk. J. Hum. Nutr. Diet. 2023, 36, 798–809. [Google Scholar] [CrossRef] [PubMed]
- Skröder, H.M.; Hamadani, J.D.; Tofail, F.; Persson, L.Å.; Vahter, M.E.; Kippler, M.J. Selenium Status in Pregnancy Influences Children’s Cognitive Function at 1.5 Years of Age. Clin. Nutr. 2015, 34, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Segura, S.A.; Ansótegui, J.A.; Marta Díaz-Gómez, N. The Importance of Maternal Nutrition during Breastfeeding: Do Breastfeeding Mothers Need Nutritional Supplements? An. Pediatría 2016, 84, 347.e1–347.e7. [Google Scholar] [CrossRef]
- Boylan, L.M.; Hart, S.; Porter, K.B.; Driskell, J.A. Vitamin B-6 Content of Breast Milk and Neonatal Behavioral Functioning. J. Am. Diet. Assoc. 2002, 102, 1433–1438. [Google Scholar] [CrossRef] [PubMed]
- Dror, D.K.; Allen, L.H. Vitamin B-12 in Human Milk: A Systematic Review. Adv. Nutr. 2018, 9, 358S–366S. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Ren, X.; Yang, Z.; Lai, J. Vitamin A Concentration in Human Milk: A Meta-Analysis. Nutrients 2022, 14, 4844. [Google Scholar] [CrossRef] [PubMed]
- Institute of Medicine. Dietary Reference Intakes for Thiamin, Riboflavin, Niacin, Vitamin B6, Folate, Vitamin B12, Pantothenic Acid, Biotin, and Choline; National Academies Press: Washington, DC, USA, 1998. [Google Scholar] [CrossRef]
- Institute of Medicine. Dietary Reference Intakes for Vitamin C, Vitamin E, Selenium, and Carotenoids; National Academies Press: Washington, DC, USA, 2000. [Google Scholar] [CrossRef]
- De Souza Mesquita, L.M.; Mennitti, L.V.; De Rosso, V.V.; Pisani, L.P. The Role of Vitamin A and Its Pro-Vitamin Carotenoids in Fetal and Neonatal Programming: Gaps in Knowledge and Metabolic Pathways. Nutr. Rev. 2021, 79, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Stoica, A.B.; Mărginean, C. The Impact of Vitamin D Deficiency on Infants’ Health. Nutrients 2023, 15, 4379. [Google Scholar] [CrossRef] [PubMed]
- Del Valle, H.B.; Yaktine, A.L.; Taylor, C.L.; Ross, A.C. Dietary Reference Intakes for Calcium and Vitamin D; National Academies Press: Washington, DC, USA, 2011. [Google Scholar] [CrossRef]
- Kazemain, E.; Ansari, S.; Davoodi, S.H.; Patterson, W.B.; Shakerinava, P.; Wagner, C.L.; Amouzegar, A. The Effect of Maternal Vitamin D Supplementation on Vitamin D Status of Exclusively Breastfeeding Mothers and Their Nursing Infants: A Systematic Review and Meta-Analysis of Randomized Clinical Trials. Adv. Nutr. 2022, 13, 568–585. [Google Scholar] [CrossRef]
- Araki, S.; Shirahata, A. Vitamin K Deficiency Bleeding in Infancy. Nutrients 2020, 12, 780. [Google Scholar] [CrossRef]
- Institute of Medicine. Dietary Reference Intakes for Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc; National Academies Press: Washington, DC, USA, 2001. [Google Scholar] [CrossRef]
| Cytokine | Possible Effect on BM | Source |
|---|---|---|
| IL-2 | regulates the growth and differentiation of the T lymphocytes and NK cells which are transported to the BM | [33] |
| IL-6 | has both pro-inflammatory and anti-inflammatory properties, stimulates mammary gland epithelial cells to increase sIgA antibody transport, and stimulates the neonatal body’s own antibody production by inducing follicular T helper cells in Peyer’s patches | [14,33,36] |
| IL-8 | leukocytes recruitment in the maternal organism and flow into the milk by chemotactic properties | [33,37] |
| IL-10 | stimulates the expansion of B lymphocytes and the regulation of the immune response in the infant gut area, and promotes the formation of a physiological microbiota | [13,34,36] |
| TGF-β1 TGF-β2 | immunoregulation by inhibiting naïve T cells from differentiation into Th1 and Th2 and maintaining the differentiation of Treg | [13,33,38] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szyller, H.; Antosz, K.; Batko, J.; Mytych, A.; Dziedziak, M.; Wrześniewska, M.; Braksator, J.; Pytrus, T. Bioactive Components of Human Milk and Their Impact on Child’s Health and Development, Literature Review. Nutrients 2024, 16, 1487. https://doi.org/10.3390/nu16101487
Szyller H, Antosz K, Batko J, Mytych A, Dziedziak M, Wrześniewska M, Braksator J, Pytrus T. Bioactive Components of Human Milk and Their Impact on Child’s Health and Development, Literature Review. Nutrients. 2024; 16(10):1487. https://doi.org/10.3390/nu16101487
Chicago/Turabian StyleSzyller, Hubert, Katarzyna Antosz, Joanna Batko, Agata Mytych, Marta Dziedziak, Martyna Wrześniewska, Joanna Braksator, and Tomasz Pytrus. 2024. "Bioactive Components of Human Milk and Their Impact on Child’s Health and Development, Literature Review" Nutrients 16, no. 10: 1487. https://doi.org/10.3390/nu16101487
APA StyleSzyller, H., Antosz, K., Batko, J., Mytych, A., Dziedziak, M., Wrześniewska, M., Braksator, J., & Pytrus, T. (2024). Bioactive Components of Human Milk and Their Impact on Child’s Health and Development, Literature Review. Nutrients, 16(10), 1487. https://doi.org/10.3390/nu16101487