
Impact of the FTO Gene Variation on Appetite and Fat Oxidation in Young Adults

 , ,
, ,  , , ,
, , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Design
2.2. Participants
2.3. Dietary Assessment
2.4. Physical Activity
2.5. Blood Samples
2.6. Body Composition
2.7. Appetite Sensation
2.8. Resting Metabolic Rate
2.9. Maximal Fat Oxidation and Cardiorespiratory Fitness
2.10. Statistical Analysis
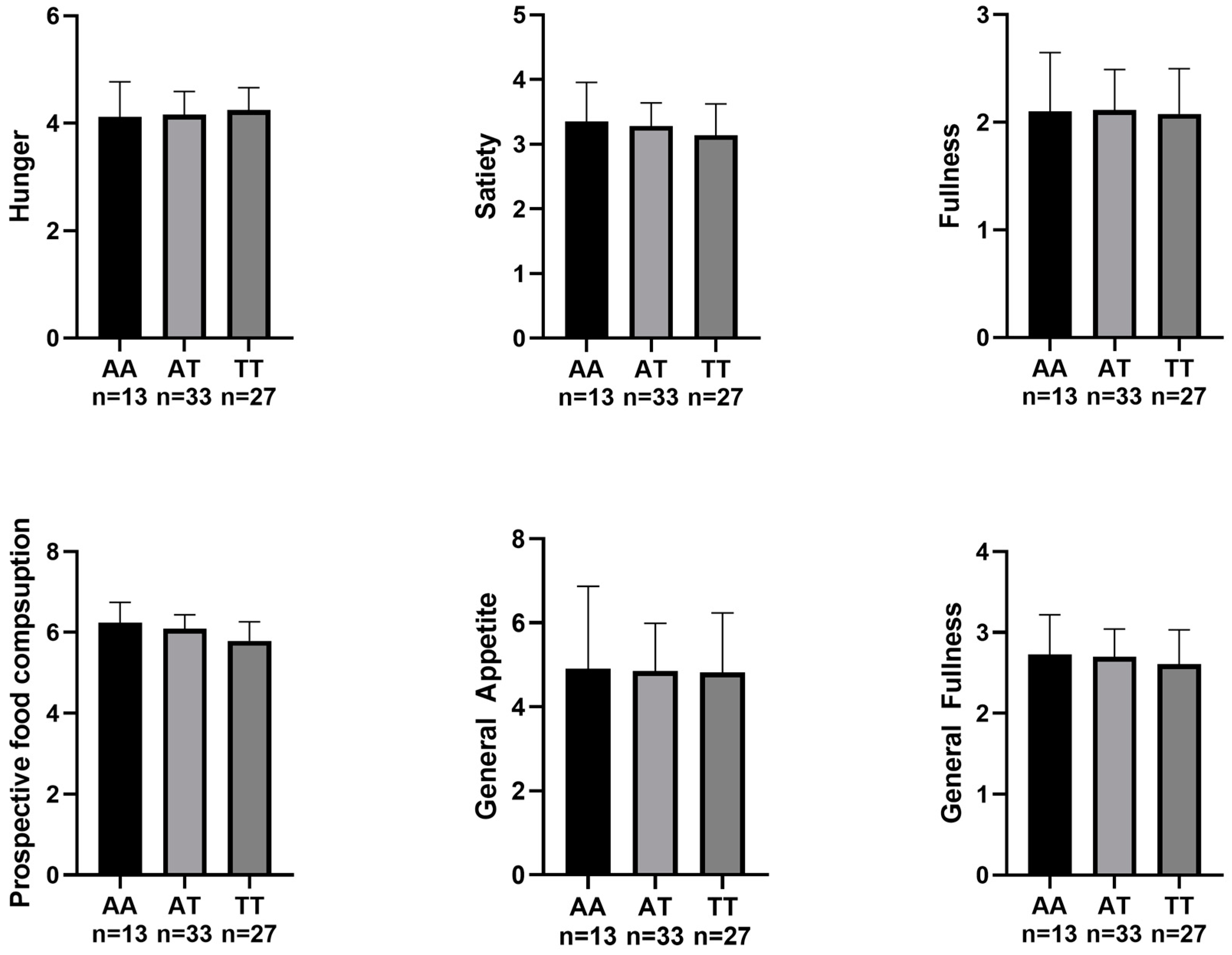
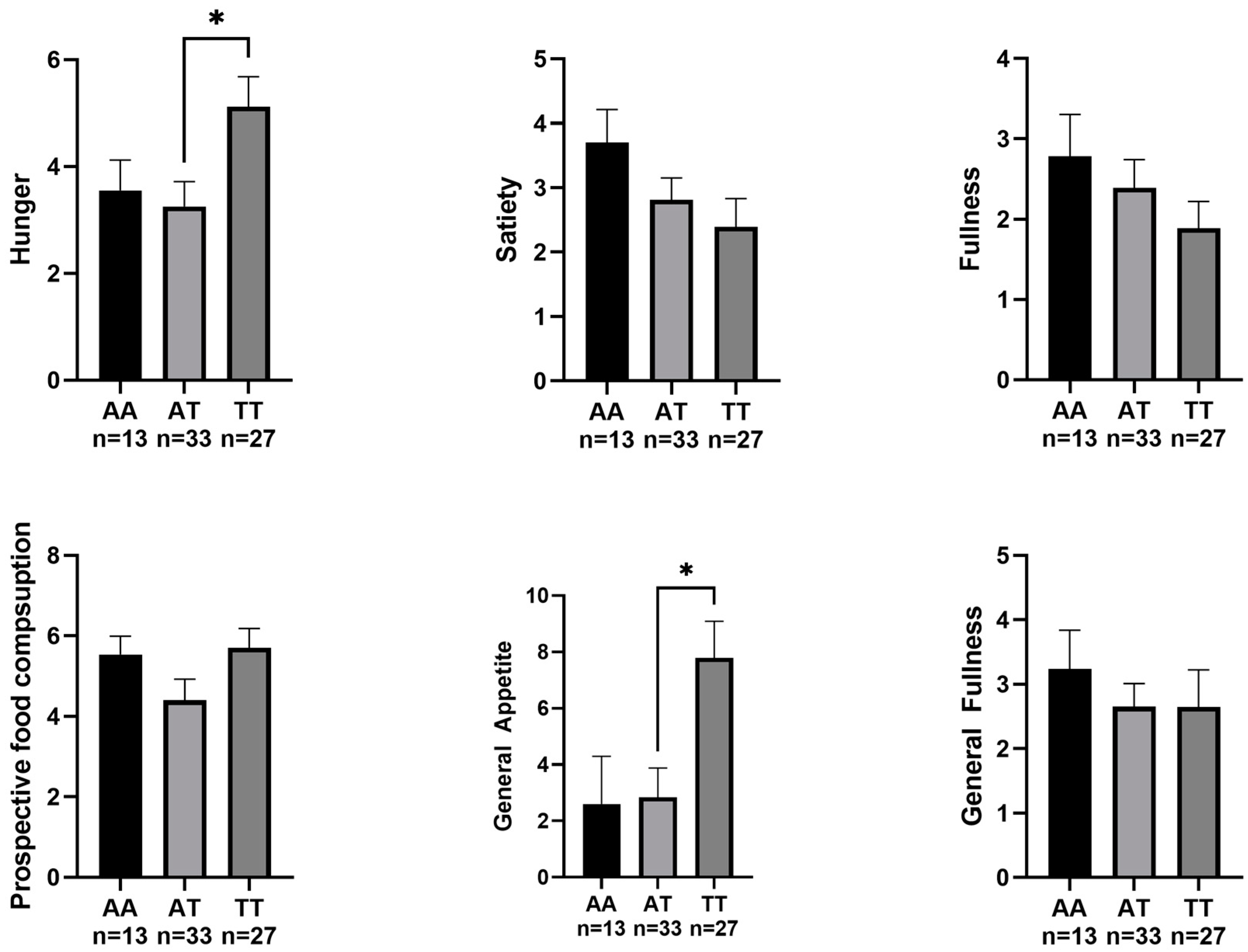
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prospective Studies Collaboration. Body-Mass Index and Cause-Specific Mortality in 900,000 Adults: Collaborative Analyses of 57 Prospective Studies. Lancet 2009, 373, 1083–1096. [Google Scholar] [CrossRef] [PubMed]
- Blüher, M. Obesity: Global Epidemiology and Pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Owen, J.B. Genetic Aspects of Body Composition. Nutrition 1999, 15, 609–613. [Google Scholar] [CrossRef] [PubMed]
- Scuteri, A.; Sanna, S.; Chen, W.-M.; Uda, M.; Albai, G.; Strait, J.; Najjar, S.; Nagaraja, R.; Orrú, M.; Usala, G.; et al. Genome-Wide Association Scan Shows Genetic Variants in the FTO Gene Are Associated with Obesity-Related Traits. PLoS Genet. 2007, 3, e115. [Google Scholar] [CrossRef]
- de Luis, D.A.; Aller, R.; Conde, R.; Izaola, O.; de la Fuente, B.; González Sagrado, M.; Primo, D.; Ruiz Mambrilla, M. Relation of the Rs9939609 Gene Variant in FTO with Cardiovascular Risk Factor and Serum Adipokine Levels in Morbid Obese Patients. Nutr. Hosp. 2012, 27, 1184–1189. [Google Scholar] [CrossRef] [PubMed]
- Karra, E.; O’Daly, O.G.; Choudhury, A.I.; Yousseif, A.; Millership, S.; Neary, M.T.; Scott, W.R.; Chandarana, K.; Manning, S.; Hess, M.E.; et al. A Link between FTO, Ghrelin, and Impaired Brain Food-Cue Responsivity. J. Clin. Investig. 2013, 123, 3539–3551. [Google Scholar] [CrossRef] [PubMed]
- McCaffery, J.M.; Papandonatos, G.D.; Peter, I.; Huggins, G.S.; Raynor, H.A.; Delahanty, L.M.; Cheskin, L.J.; Balasubramanyam, A.; Wagenknecht, L.E.; Wing, R.R. Obesity Susceptibility Loci and Dietary Intake in the Look AHEAD Trial. Am. J. Clin. Nutr. 2012, 95, 1477–1486. [Google Scholar] [CrossRef]
- Andreasen, C.H.; Stender-Petersen, K.L.; Mogensen, M.S.; Torekov, S.S.; Wegner, L.; Andersen, G.; Nielsen, A.L.; Albrechtsen, A.; Borch-Johnsen, K.; Rasmussen, S.S.; et al. Low Physical Activity Accentuates the Effect of the FTO Rs9939609 Polymorphism on Body Fat Accumulation. Diabetes 2008, 57, 95–101. [Google Scholar] [CrossRef]
- Rauhio, A.; Uusi-Rasi, K.; Nikkari, S.T.; Kannus, P.; Sievänen, H.; Kunnas, T. Association of the FTO and ADRB2 Genes with Body Composition and Fat Distribution in Obese Women. Maturitas 2013, 76, 165–171. [Google Scholar] [CrossRef]
- Magno, F.; Guaraná, H.; Fonseca, A.C.; Cabello, G.; Carneiro, J.; Pedrosa, A.; Ximenes, A.; Rosado, E. Influence of FTO Rs9939609 Polymorphism on Appetite, Ghrelin, Leptin, IL6, TNFα Levels, and Food Intake of Women with Morbid Obesity. Diabetes Metab. Syndr. Obes. Targets Ther. 2018, 11, 199–207. [Google Scholar] [CrossRef]
- Goltz, F.R.; Thackray, A.E.; Varela-Mato, V.; King, J.A.; Dorling, J.L.; Dowejko, M.; Mastana, S.; Thompson, J.; Atkinson, G.; Stensel, D.J. Exploration of Associations between the FTO Rs9939609 Genotype, Fasting and Postprandial Appetite-Related Hormones and Perceived Appetite in Healthy Men and Women. Appetite 2019, 142, 104368. [Google Scholar] [CrossRef] [PubMed]
- Kowalska, I.; Adamska, A.; Malecki, M.T.; Karczewska-Kupczewska, M.; Nikolajuk, A.; Szopa, M.; Gorska, M.; Straczkowski, M. Impact of the FTO Gene Variation on Fat Oxidation and Its Potential Influence on Body Weight in Women with Polycystic Ovary Syndrome. Clin. Endocrinol. 2012, 77, 120–125. [Google Scholar] [CrossRef]
- Blauw, L.L.; Noordam, R.; Trompet, S.; Berbée, J.F.P.; Rosendaal, F.R.; van Heemst, D.; van Dijk, K.W.; Mook-Kanamori, D.O.; de Mutsert, R.; Rensen, P.C.N. Genetic Variation in the Obesity Gene FTO Is Not Associated with Decreased Fat Oxidation: The NEO Study. Int. J. Obes. 2017, 41, 1594–1600. [Google Scholar] [CrossRef]
- Kelley, D.E.; Mandarino, L.J. Fuel Selection in Human Skeletal Muscle in Insulin Resistance: A Reexamination. Diabetes 2000, 49, 677–683. [Google Scholar] [CrossRef]
- Montes-de-Oca-García, A.; Perez-Bey, A.; Corral-Pérez, J.; Velázquez-Díaz, D.; Opazo-Díaz, E.; Fernandez-Santos, J.R.; Rebollo-Ramos, M.; Amaro-Gahete, F.J.; Cuenca-García, M.; Ponce-González, J.-G. Maximal Fat Oxidation Capacity Is Associated with Cardiometabolic Risk Factors in Healthy Young Adults. Eur. J. Sport Sci. 2021, 21, 907–917. [Google Scholar] [CrossRef]
- Corral-Pérez, J.; Velázquez-Díaz, D.; Perez-Bey, A.; Montes-de-Oca-García, A.; Fernandez-Santos, J.R.; Amaro-Gahete, F.J.; Jiménez-Pavón, D.; Casals, C.; Ponce-González, J.G. Accelerometer-Measured Physical Activity and Sedentary Time Are Associated with Maximal Fat Oxidation in Young Adults. Eur. J. Sport Sci. 2022, 22, 1595–1604. [Google Scholar] [CrossRef]
- Montes-de-Oca-García, A.; Perez-Bey, A.; Velázquez-Díaz, D.; Corral-Pérez, J.; Opazo-Díaz, E.; Rebollo-Ramos, M.; Gómez-Gallego, F.; Cuenca-García, M.; Casals, C.; Ponce-González, J.G. Influence of ACE Gene I/D Polymorphism on Cardiometabolic Risk, Maximal Fat Oxidation, Cardiorespiratory Fitness, Diet and Physical Activity in Young Adults. Int. J. Environ. Res. Public Health 2021, 18, 3443. [Google Scholar] [CrossRef] [PubMed]
- Rebollo-Ramos, M.; Velázquez-Díaz, D.; Corral-Pérez, J.; Barany-Ruiz, A.; Pérez-Bey, A.; Fernández-Ponce, C.; García-Cózar, F.J.; Ponce-González, J.G.; Cuenca-García, M. Capacidad Aeróbica, Dieta Mediterránea y Riesgo Cardiometabólico En Adultos. Endocrinol. Diabetes Nutr. 2020, 67, 113–121. [Google Scholar] [CrossRef]
- Montes-de-Oca-García, A.; Corral-Pérez, J.; Velázquez-Díaz, D.; Perez-Bey, A.; Rebollo-Ramos, M.; Marín-Galindo, A.; Gómez-Gallego, F.; Calderon-Dominguez, M.; Casals, C.; Ponce-González, J.G. Influence of Peroxisome Proliferator-Activated Receptor (PPAR)-Gamma Coactivator (PGC)-1 Alpha Gene Rs8192678 Polymorphism by Gender on Different Health-Related Parameters in Healthy Young Adults. Front. Physiol. 2022, 13, 885185. [Google Scholar] [CrossRef]
- Ward, D.S.; Evenson, K.R.; Vaughn, A.; Rodgers, A.B.; Troiano, R.P. Accelerometer Use in Physical Activity: Best Practices and Research Recommendations. Med. Sci. Sports Exerc. 2005, 37, S582–S588. [Google Scholar] [CrossRef]
- Marcotte, R.T.; Petrucci, G.J.; Cox, M.F.; Freedson, P.S.; Staudenmayer, J.W.; Sirard, J.R. Estimating Sedentary Time from a Hip- and Wrist-Worn Accelerometer. Med. Sci. Sports Exerc. 2020, 52, 225–232. [Google Scholar] [CrossRef]
- Migueles, J.H.; Cadenas-Sanchez, C.; Ekelund, U.; Delisle Nyström, C.; Mora-Gonzalez, J.; Löf, M.; Labayen, I.; Ruiz, J.R.; Ortega, F.B. Accelerometer Data Collection and Processing Criteria to Assess Physical Activity and Other Outcomes: A Systematic Review and Practical Considerations. Sport. Med. 2017, 47, 1821–1845. [Google Scholar] [CrossRef]
- Choi, L.; Liu, Z.; Matthews, C.E.; Buchowski, M.S. Validation of Accelerometer Wear and Nonwear Time Classification Algorithm. Med. Sci. Sports Exerc. 2011, 43, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Kozey-Keadle, S.; Libertine, A.; Lyden, K.; Staudenmayer, J.; Freedson, P.S. Validation of Wearable Monitors for Assessing Sedentary Behavior. Med. Sci. Sports Exerc. 2011, 43, 1561–1567. [Google Scholar] [CrossRef]
- Knopp, J.L.; Holder-Pearson, L.; Chase, J.G. Insulin Units and Conversion Factors: A Story of Truth, Boots, and Faster Half-Truths. J. Diabetes Sci. Technol. 2019, 13, 597–600. [Google Scholar] [CrossRef] [PubMed]
- Cornier, M.-A.; Grunwald, G.K.; Johnson, S.L.; Bessesen, D.H. Effects of Short-Term Overfeeding on Hunger, Satiety, and Energy Intake in Thin and Reduced-Obese Individuals. Appetite 2004, 43, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Frayn, K.N. Calculation of Substrate Oxidation Rates in Vivo from Gaseous Exchange. J. Appl. Physiol. 1983, 55, 628–634. [Google Scholar] [CrossRef] [PubMed]
- Achten, J.; Gleeson, M.; Jeukendrup, A.E. Determination of the Exercise Intensity That Elicits Maximal Fat Oxidation. Med. Sci. Sports Exerc. 2002, 34, 92–97. [Google Scholar] [CrossRef]
- Frayling, T.M.; Timpson, N.J.; Weedon, M.N.; Zeggini, E.; Freathy, R.M.; Lindgren, C.M.; Perry, J.R.B.; Elliott, K.S.; Lango, H.; Rayner, N.W.; et al. A Common Variant in the FTO Gene Is Associated with Body Mass Index and Predisposes to Childhood and Adult Obesity. Science 2007, 316, 889–894. [Google Scholar] [CrossRef]
- Dorling, J.L.; Clayton, D.J.; Jones, J.; Carter, W.G.; Thackray, A.E.; King, J.A.; Pucci, A.; Batterham, R.L.; Stensel, D.J. A Randomized Crossover Trial Assessing the Effects of Acute Exercise on Appetite, Circulating Ghrelin Concentrations, and Butyrylcholinesterase Activity in Normal-Weight Males with Variants of the Obesity-Linked FTO Rs9939609 Polymorphism. Am. J. Clin. Nutr. 2019, 110, 1055–1066. [Google Scholar] [CrossRef]
- den Hoed, M.; Westerterp-Plantenga, M.S.; Bouwman, F.G.; Mariman, E.C.; Westerterp, K.R. Postprandial Responses in Hunger and Satiety Are Associated with the Rs9939609 Single Nucleotide Polymorphism in FTO. Am. J. Clin. Nutr. 2009, 90, 1426–1432. [Google Scholar] [CrossRef] [PubMed]
- Rosenkilde, M.; Nordby, P.; Nielsen, L.B.; Stallknecht, B.M.; Helge, J.W. Fat Oxidation at Rest Predicts Peak Fat Oxidation during Exercise and Metabolic Phenotype in Overweight Men. Int. J. Obes. 2010, 34, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.L.; Hattersley, J.; Frost, G.S.; Chambers, E.S.; Wallis, G.A. Maximal Fat Oxidation during Exercise Is Positively Associated with 24-Hour Fat Oxidation and Insulin Sensitivity in Young, Healthy Men. J. Appl. Physiol. 2015, 118, 1415–1422. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Men (n = 46) | Women (n = 27) | p | |
|---|---|---|---|
| Men, No. (%) | |||
| Age (years) | 22.24 ± 3.67 | 23.18 ± 5.06 | 0.359 |
| Energy intake (kcal/day) | 2848.00 ± 1070.62 | 2233.32 ± 604.43 | 0.007 |
| Light physical activity (min/day) | 1911.48 ± 377.10 | 2104.48 ± 519.16 | 0.084 |
| Moderate physical activity (min/day) | 301.71 ± 96.70 | 332.72 ± 143.48 | 0.295 |
| Vigorous physical activity (min/day) | 38.33 ± 37.29 | 30.08 ± 27.01 | 0.407 |
| Physical activity expenditure (kcal/day) | 327.72 ± 132.67 | 289.56 ± 117.37 | 0.239 |
| Plasma triglycerides (mg/dL) | 71.72 ± 24.92 | 66.83 ± 24.58 | 0.414 |
| Plasma glucose (mg/dL) | 103.44 ± 9.64 | 97.03 ± 9.93 | 0.008 |
| Plasma insulin (mUI/L) | 23.56 ± 22.76 | 22.64 ± 19.37 | 0.860 |
| Plasma ghrelin (pg/mL) | 15.66 ± 9.31 | 25.27 ± 20.94 | 0.009 |
| Plasma leptin (ng/mL) | 1.99 ± 2.86 | 7.36 ± 4.93 | <0.001 |
| Plasma GIP (pg/mL) | 65.98 ± 36.88 | 66.58 ± 29.38 | 0.942 |
| Plasma GLP-1 (pg/mL) | 8.45 ± 5.75 | 11.49 ± 9.16 | 0.085 |
| Plasma PYY (pg/mL) | 88.92 ± 45.48 | 94.44 ± 45.36 | 0.663 |
| Height (cm) | 176.74 ± 6.13 | 164.62 ± 6.81 | <0.001 |
| Body mass (kg) | 78.75 ± 72.27 | 72.2 ± 16.98 | 0.089 |
| BMI (kg/m2) | 25.16 ± 4.17 | 26.86 ± 7.39 | 0.210 |
| Fat mass (kg) | 15.84 ± 9.26 | 23.37 ± 12.89 | 0.005 |
| Fat mass (%) | 18.95 ± 7.36 | 30.28 ± 9.54 | <0.001 |
| Lean mass (kg) | 59.46 ± 6.49 | 46.04 ± 4.86 | <0.001 |
| Lean mass (%) | 76.56 ± 6.55 | 65.54 ± 8.49 | <0.001 |
| Resting metabolic rate (kcal/day) | 2013.29 ± 276.76 | 1580.07 ± 235.81 | <0.001 |
| Energy expenditure (kcal/day) | 2321.49 ± 333.76 | 1838.60 ± 308.04 | <0.001 |
| Energy balance (kcal/day) | 513.39 ± 1173.05 | 394.72 ± 703.76 | 0.630 |
| VO2max (mL/min) | 3483.73 ± 530.92 | 2287.89 ± 419.47 | <0.001 |
| Vo2max (mL/kg/min) | 45.51 ± 10.55 | 33.72 ± 10.36 | <0.001 |
| AA (n = 13) | AT (n = 33) | TT (n = 27) | p | |
|---|---|---|---|---|
| Men, No. (%) | 9 (69.2) | 22 (64.7) | 15 (55.6) | 0.647 |
| Age (years) | 22.54 ± 4.48 | 22.15 ± 3.42 | 23.19 ± 5.07 | 0.642 |
| Energy intake (kcal/day) | 2474.00 ± 589.60 | 2412.00 ± 646.30 | 2941.00 ± 1323.00 | 0.086 |
| Light physical activity (min/day) | 1946.00 ± 444.60 | 1971.00 ± 415.00 | 1977.00 ± 470.7 | 0.978 |
| Moderate physical activity (min/day) | 328.30 ± 84.46 | 315.40 ± 104.10 | 305.40 ± 144.90 | 0.856 |
| Vigorous physical activity (min/day) | 9.22 ± 7.71 * | 28.07 ± 22.07 | 33.38 ± 30.88 | 0.055 |
| Physical activity expenditure (kcal/day) | 365.30 ± 140.10 | 297.60 ± 121.80 | 307.60 ± 127.20 | 0.292 |
| Plasma triglycerides (mg/dL) | 76.39 ± 33.81 | 69.44 ± 24.37 | 67.26 ± 20.21 | 0.551 |
| Plasma glucose (mg/dL) | 99.54 ± 7.77 | 103.44 ± 10.49 | 98.66 ± 10.42 | 0.161 |
| Plasma insulin (mUI/L) | 16.66 ± 11.56 | 23.39 ± 21.48 | 26.14 ± 24.67 | 0.428 |
| Plasma ghrelin (ng/mL) | 21.88 ± 17.59 | 21.15 ± 18.22 | 15.92 ± 9.61 | 0.352 |
| Plasma leptin (ng/mL) | 5.31 ± 5.22 | 3.23 ± 3.10 | 4.12 ± 4.02 | 0.392 |
| Plasma GIP (pg/mL) | 74.36 ± 34.09 | 66.16 ± 6.91 | 62.35 ± 25.96 | 0.584 |
| Plasma GLP-1 (pg/mL) | 10.93 ± 7.15 | 9.39 ± 8.33 | 9.31 ± 6.20 | 0.798 |
| Plasma PYY (pg/mL) | 75.75 ± 33.17 | 96.16 ± 45.10 | 90.98 ± 50.58 | 0.478 |
| Height (cm) | 175.00 ± 9.34 | 3.23 ± 3.92 | 4.12 ± 4.23 | 0.177 |
| Body mass (kg) | 81.66 ± 15.66 | 75.15 ± 16.42 | 75.16 ± 15.34 | 0.412 |
| BMI (kg/m2) | 26.77 ± 5.40 | 25.00 ± 4.29 | 25.36 ± 4.95 | 0.518 |
| Fat mass (kg) | 19.86 ± 10.82 | 17.16 ± 10.31 | 18.53 ± 10.28 | 0.330 |
| Fat mass (%) | 23.25 ± 9.90 | 21.70 ± 9.05 | 22.73 ± 8.87 | 0.842 |
| Lean mass (kg) | 58.40 ± 9.04 | 54.65 ± 9.36 | 52.12 ± 7.46 | 0.104 |
| Lean mass (%) | 72.45 ± 9.06 | 73.68 ± 8.13 | 71.67 ± 9.09 | 0.454 |
| Resting metabolic rate (kcal/day) | 1978.00 ± 352.30 | 1801.00 ± 357.80 | 1840.00 ± 292.90 | 0.275 |
| Energy expenditure (kcal/day) | 2346.00 ± 467.90 | 2063.00 ± 370.30 | 2125.00 ± 378.30 | 0.102 |
| Energy balance (kcal/day) | 127.80 ± 782.40 | 317.00 ± 708.70 | 816.00 ± 253.80 | 0.066 |
| VO2max (mL/min) | 3189.00 ± 680.00 | 2969.00 ± 674.60 | 3015.00 ± 904.40 | 0.680 |
| Vo2max (mL/kg/min) | 39.92 ± 8.97 | 40.59 ± 10.64 | 42.00 ± 14.63 | 0.849 |
| AA (n = 13) | AT (n = 33) | TT (n = 27) | p | |
|---|---|---|---|---|
| MFO (g/min) | 0.38 ± 0.15 | 0.33 ± 0.15 * | 0.42 ± 0.15 | 0.031 |
| MFO/VO2max (mg/mL/min) | 0.12 ± 0.04 | 0.11 ± 0.04 ** | 0.14 ± 0.04 | 0.004 |
| MFO/lean mass (mg/kg/min) | 6.57 ± 2.37 | 6.25 ± 2.86 * | 8.21 ± 2.76 | 0.011 |
| Fatmax (%) | 40.98 ± 8.81 | 40.25 ± 7.31 | 42.07 ± 6.88 | 0.642 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ponce-Gonzalez, J.G.; Martínez-Ávila, Á.; Velázquez-Díaz, D.; Perez-Bey, A.; Gómez-Gallego, F.; Marín-Galindo, A.; Corral-Pérez, J.; Casals, C. Impact of the FTO Gene Variation on Appetite and Fat Oxidation in Young Adults. Nutrients 2023, 15, 2037. https://doi.org/10.3390/nu15092037
Ponce-Gonzalez JG, Martínez-Ávila Á, Velázquez-Díaz D, Perez-Bey A, Gómez-Gallego F, Marín-Galindo A, Corral-Pérez J, Casals C. Impact of the FTO Gene Variation on Appetite and Fat Oxidation in Young Adults. Nutrients. 2023; 15(9):2037. https://doi.org/10.3390/nu15092037
Chicago/Turabian StylePonce-Gonzalez, Jesús G., Ángel Martínez-Ávila, Daniel Velázquez-Díaz, Alejandro Perez-Bey, Félix Gómez-Gallego, Alberto Marín-Galindo, Juan Corral-Pérez, and Cristina Casals. 2023. "Impact of the FTO Gene Variation on Appetite and Fat Oxidation in Young Adults" Nutrients 15, no. 9: 2037. https://doi.org/10.3390/nu15092037
APA StylePonce-Gonzalez, J. G., Martínez-Ávila, Á., Velázquez-Díaz, D., Perez-Bey, A., Gómez-Gallego, F., Marín-Galindo, A., Corral-Pérez, J., & Casals, C. (2023). Impact of the FTO Gene Variation on Appetite and Fat Oxidation in Young Adults. Nutrients, 15(9), 2037. https://doi.org/10.3390/nu15092037