
Neuroprotective Effects of Agri-Food By-Products Rich in Phenolic Compounds
 ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
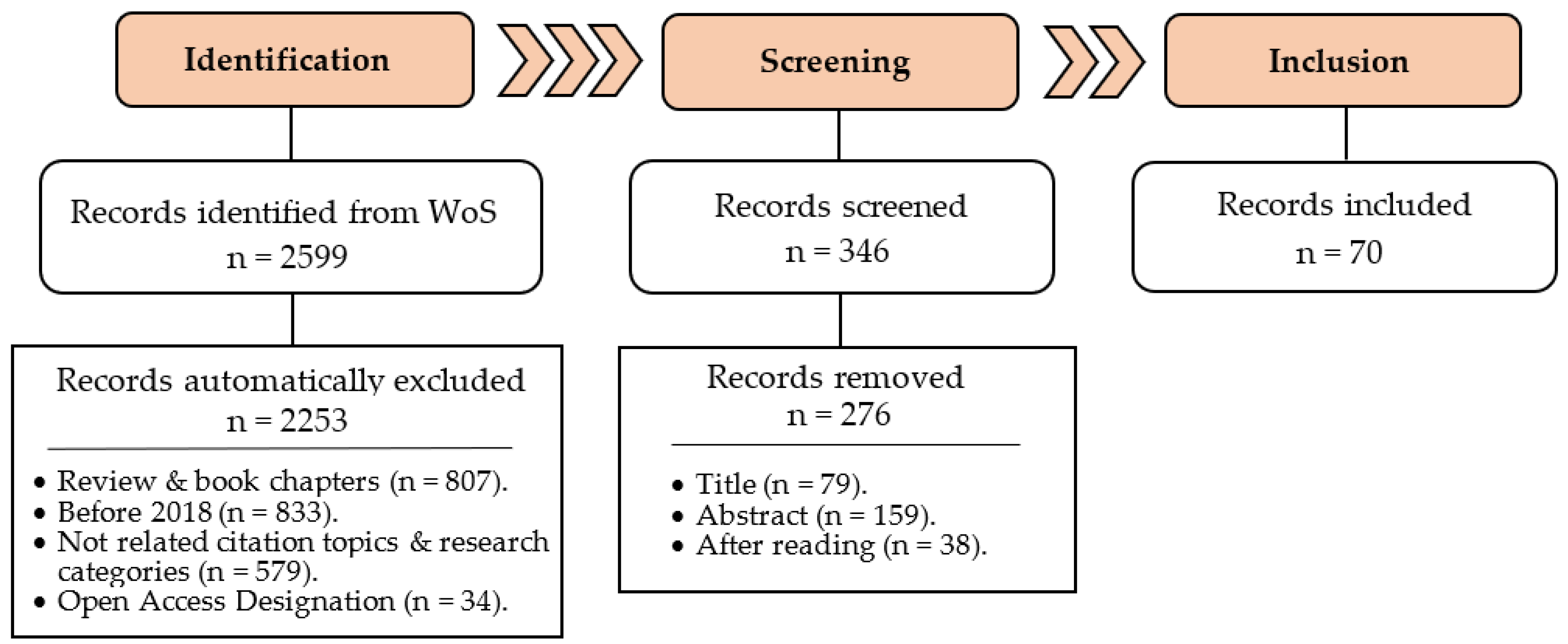
2. Materials and Methods
2.1. Search Strategy
2.2. Inclusion and Exclusion Criteria
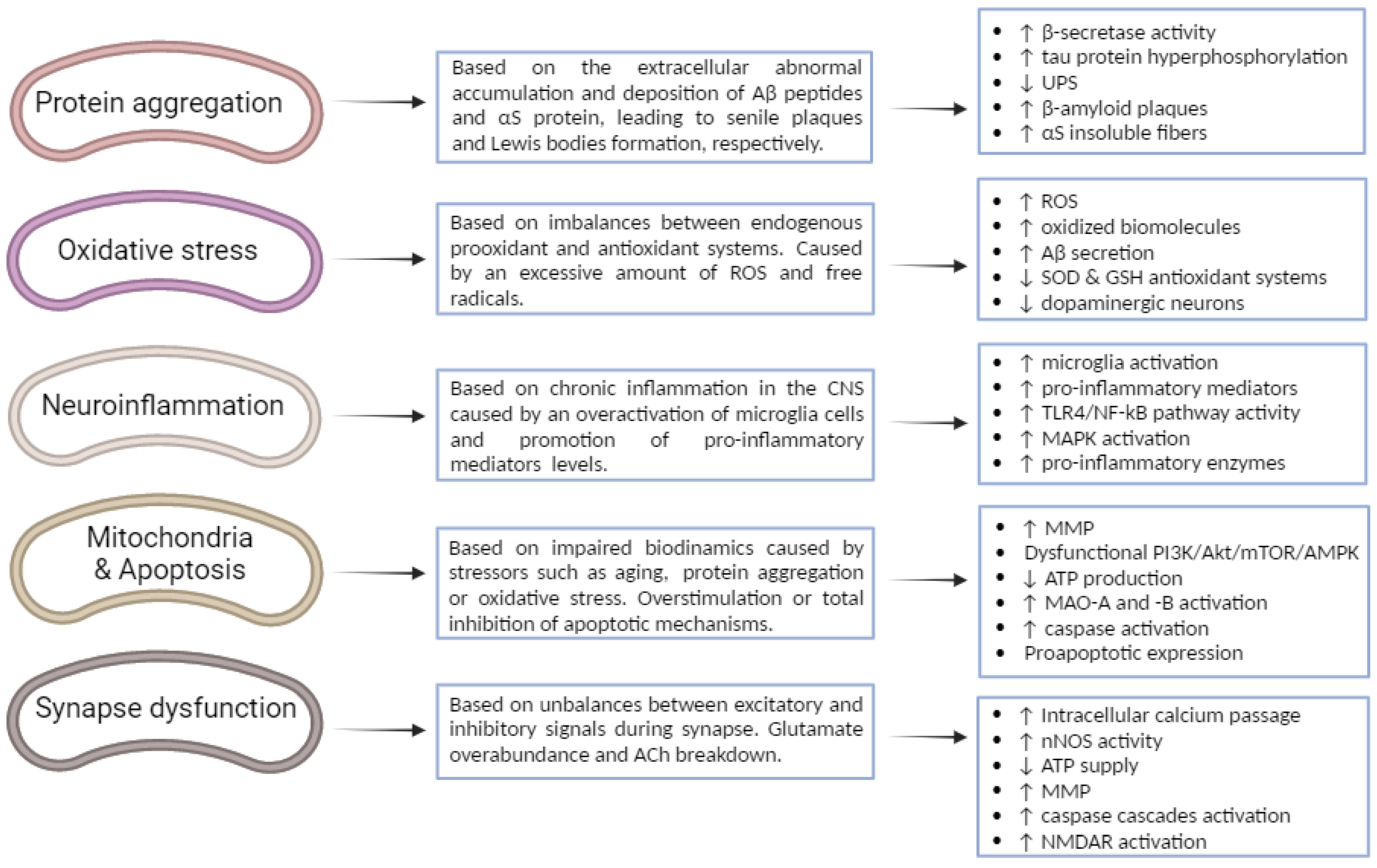
3. Pathogenesis and Main Biological Mechanisms of Neurodegeneration
3.1. Protein Misfolding, Aggregation and Deposition
3.2. Oxidative Stress
3.3. Neuroinflammation
3.4. Impaired Biodynamics and Mitochondrial Dysfunctions
3.5. Excitotoxicity—Glutamatergic and Cholinergic Neurotransmissions
4. Phenolic Compounds from Fruit and Vegetable By-Products: Neuroprotective Mechanisms for Potential Biological Targets
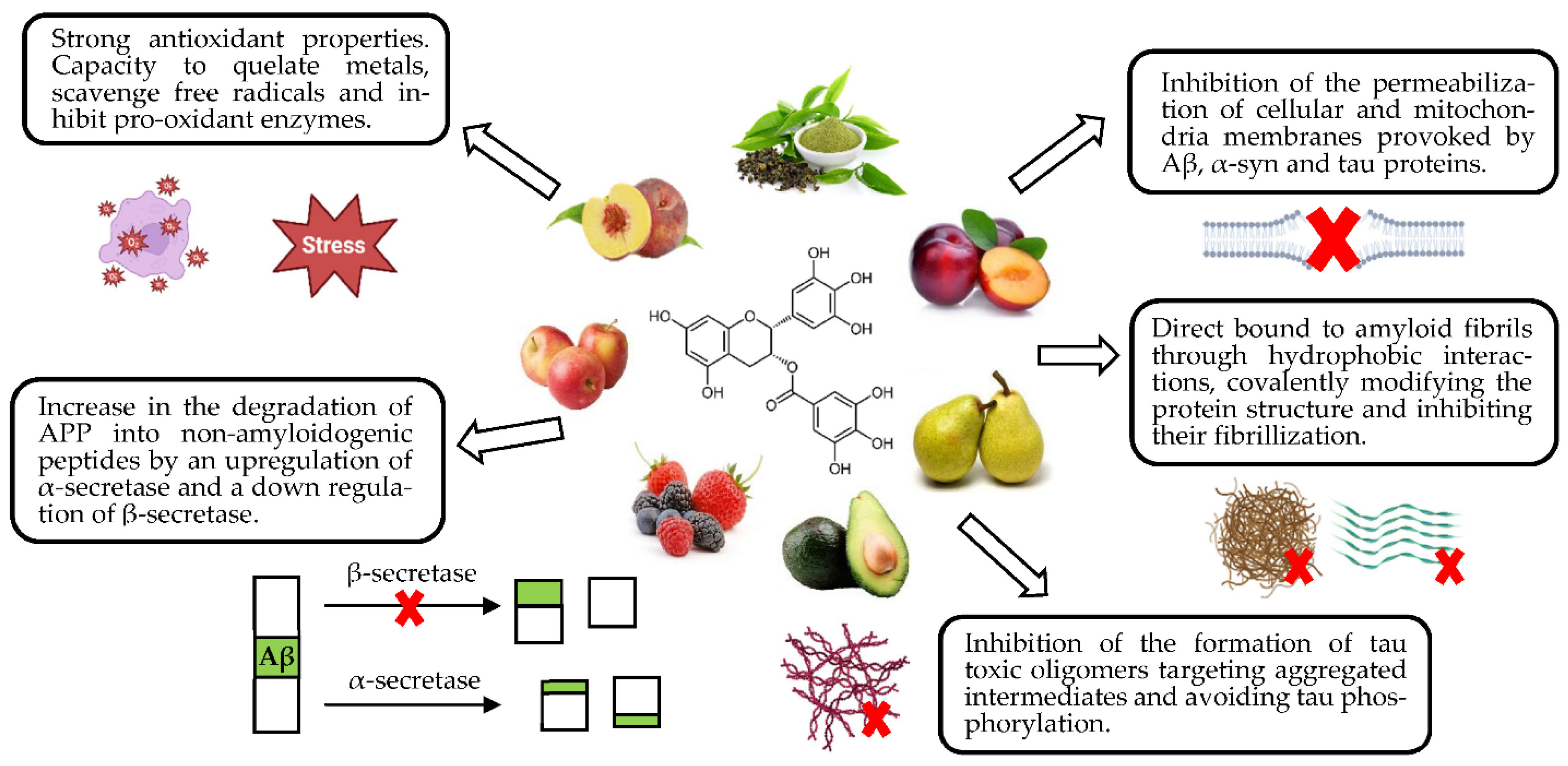
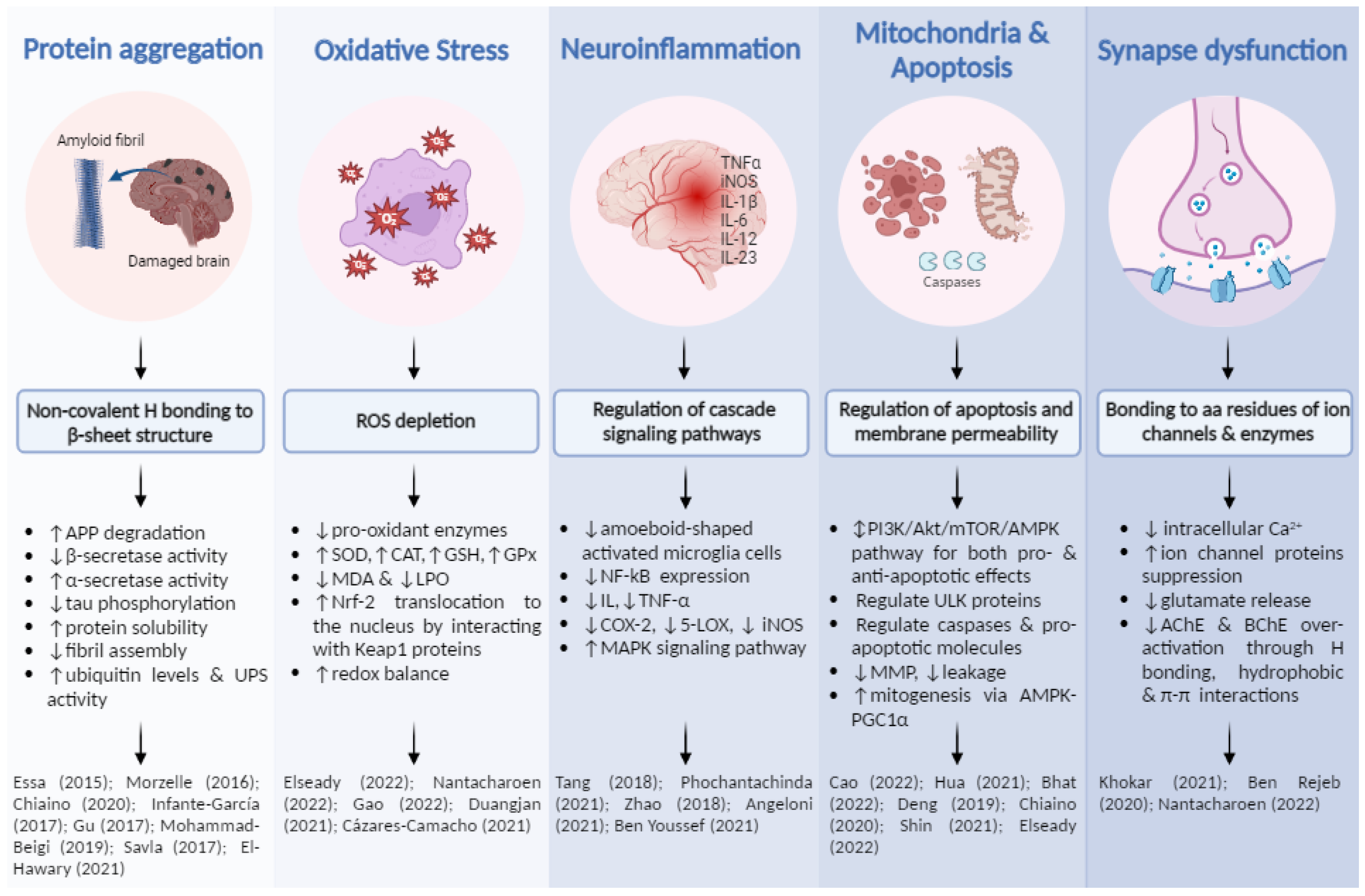
4.1. Neuroprotection against Abnormal Protein Deposition
4.2. Neuroprotection against Oxidative Stress
4.3. Neuroprotection against Neuroinflammation
4.4. Neuroprotection against Mitochondrial Dysfunctions
4.5. Neuroprotection against Impaired Glutamatergic and Cholinergic Neurotransmissions
5. Conclusions and Future Perspectives
- Digestion, absorption and metabolization of fruit, vegetables and other plants are crucial for recovering phenolic and other bioactive compounds from their matrices. Their bioavailability is considered one of the most limiting factors for plants to exert neuroprotection, since their effects are positively related to the amount consumed [1,156]. Low absorption rates and quick metabolism could limit their efficacy, but abusive consumption could lead polyphenols to show pro-oxidant activity.
- Secondary metabolites from phenolic compounds should be able to pass the BBB to reach the brain. Therefore, their effectiveness also depends on their permeability capacity across this barrier [12].
- Phenolic compounds are not the only compounds in plant matrices, so neuroprotection could be exerted by others too, and may be under synergistic or antagonistic interactions.
- Difficulties in carrying out in vivo studies in humans.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Silva, R.F.M.; Pogačnik, L. Polyphenols from food and natural products: Neuroprotection and safety. Antioxidants 2020, 9, 61. [Google Scholar] [CrossRef]
- Grodzicki, W.; Dziendzikowska, K. The role of selected bioactive compounds in the prevention of alzheimer’s disease. Antioxidants 2020, 9, 229. [Google Scholar] [CrossRef] [PubMed]
- Alzheimer’s Association 2022 Alzheimer’s Disease. Facts and Figures. Alzheimer’s Assoc. 2022. Available online: https://www.alz.org/media/documents/alzheimers-facts-and-figures.pdf (accessed on 2 January 2023).
- Alzheimer’s Disease International: Numbers of People with Dementia around the World. Dement. Stat. 2020. Available online: https://www.alzint.org/resource/numbers-of-people-with-dementia-worldwide/ (accessed on 2 January 2023).
- Gorzkiewicz, J.; Bartosz, G.; Sadowska-Bartosz, I. The potential effects of phytoestrogens: The role in neuroprotection. Molecules 2021, 26, 2954. [Google Scholar] [CrossRef]
- Mohd Sairazi, N.S.; Sirajudeen, K.N.S. Natural Products and Their Bioactive Compounds: Neuroprotective Potentials against Neurodegenerative Diseases. Evid.-Based Complement. Altern. Med. 2020, 2020, 6565396. [Google Scholar] [CrossRef]
- Understanding Parkinson’s, Statistics. 2019. Available online: https://www.parkinson.org/understanding-parkinsons/statistics (accessed on 5 December 2022).
- Ly, H.T.; Nguyen, T.T.H.; Le, V.M.; Lam, B.T.; Mai, T.T.T.; Dang, T.P.T. Therapeutic Potential of Polyscias fruticosa (L.) Harms Leaf Extract for Parkinson’s Disease Treatment by Drosophila melanogaster Model. Oxid. Med. Cell. Longev. 2022, 2022, 5262677. [Google Scholar] [CrossRef] [PubMed]
- Naoi, M.; Shamoto-Nagai, M.; Maruyama, W. Neuroprotection of multifunctional phytochemicals as novel therapeutic strategy for neurodegenerative disorders: Antiapoptotic and antiamyloidogenic activities by modulation of cellular signal pathways. Future Neurol. 2019, 14, FNL9. [Google Scholar] [CrossRef]
- Sun, C.; Zhang, S.; Ba, S.; Dang, J.; Ren, Q.; Zhu, Y.; Liu, K.; Jin, M. Eucommia ulmoides Olive Male Flower Extracts Ameliorate Alzheimer’s Disease-Like Pathology in Zebrafish via Regulating Autophagy, Acetylcholinesterase, and the Dopamine Transporter. Front. Mol. Neurosci. 2022, 15, 901953. [Google Scholar] [CrossRef]
- Katsnelson, A.; De Strooper, B.; Zoghbi, H.Y. Neurodegeneration: From cellular concepts to clinical applications. Sci. Transl. Med. 2016, 8, 36418. [Google Scholar] [CrossRef]
- Hornedo-Ortega, R.; Cerezo, A.B.; de Pablos, R.M.; Krisa, S.; Richard, T.; García-Parrilla, M.C.; Troncoso, A.M. Phenolic compounds characteristic of the mediterranean diet in mitigating microglia-mediated neuroinflammation. Front. Cell. Neurosci. 2018, 12, 373. [Google Scholar] [CrossRef]
- Pohl, F.; Lin, P.K.T. The potential use of plant natural products and plant extracts with antioxidant properties for the prevention/treatment of neurodegenerative diseases: In vitro, in vivo and clinical trials. Molecules 2018, 23, 3283. [Google Scholar] [CrossRef]
- Ma, Y.; Ma, B.; Shang, Y.; Yin, Q.; Wang, D.; Xu, S.; Hong, Y.; Hou, X.; Liu, X. Flavonoid-rich ethanol extract from the leaves of Diospyros kaki attenuates D-galactose-induced oxidative stress and neuroinflammation-mediated brain aging in mice. Oxid. Med. Cell. Longev. 2018, 2018, 8938207. [Google Scholar] [CrossRef] [PubMed]
- Ha, N.M.; Tran, S.H.; Shim, Y.H.; Kang, K. Caenorhabditis elegans as a powerful tool in natural product bioactivity research. Appl. Biol. Chem. 2022, 65, 18. [Google Scholar] [CrossRef]
- Fierascu, R.C.; Sieniawska, E.; Ortan, A.; Fierascu, I.; Xiao, J. Fruits By-Products—A Source of Valuable Active Principles. A Short Review. Front. Bioeng. Biotechnol. 2020, 8, 319. [Google Scholar] [CrossRef] [PubMed]
- Limanaqi, F.; Biagioni, F.; Mastroiacovo, F.; Polzella, M.; Lazzeri, G.; Fornai, F. Merging the multi-target effects of phytochemicals in neurodegeneration: From oxidative stress to protein aggregation and inflammation. Antioxidants 2020, 9, 22. [Google Scholar] [CrossRef]
- Figueira, I.; Garcia, G.; Pimpão, R.C.; Terrasso, A.P.; Costa, I.; Almeida, A.F.; Tavares, L.; Pais, T.F.; Pinto, P.; Ventura, M.R.; et al. Polyphenols journey through blood-brain barrier towards neuronal protection. Sci. Rep. 2017, 7, 11456. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Bhat, R. Suppression, disaggregation, and modulation of γ-Synuclein fibrillation pathway by green tea polyphenol EGCG. Protein Sci. 2019, 28, 382–402. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, L.C.; Mendiola, J.A.; Sánchez-camargo, A.D.P.; Álvarez-rivera, G.; Viganó, J.; Cifuentes, A.; Ibáñez, E.; Martínez, J. Selective extraction of piceatannol from passiflora edulis by-products: Application of hsps strategy and inhibition of neurodegenerative enzymes. Int. J. Mol. Sci. 2021, 22, 6248. [Google Scholar] [CrossRef]
- Maurya, H. Neuroprotective Potential of Swietenia macrophylla Seed Extract in Lead-induced Neurodegeneration in Albino Rats. Asian J. Biol. Sci. 2019, 12, 442–449. [Google Scholar] [CrossRef]
- Ortega-Arellano, H.F.; Jimenez-Del-Rio, M.; Velez-Pardo, C. Neuroprotective Effects of Methanolic Extract of Avocado Persea americana (var. Colinred) Peel on Paraquat-Induced Locomotor Impairment, Lipid Peroxidation and Shortage of Life Span in Transgenic knockdown Parkin Drosophila melanogaster. Neurochem. Res. 2019, 44, 1986–1998. [Google Scholar] [CrossRef]
- García-Villegas, A.; Rojas-García, A.; Villegas-Aguilar, M.D.C.; Fernández-Moreno, P.; Fernández-Ochoa, Á.; Cádiz-Gurrea, M.D.L.L.; Arráez-Román, D.; Segura-Carretero, A. Cosmeceutical Potential of Major Tropical and Subtropical Fruit By-Products for a Sustainable Revalorization. Antioxidants 2022, 11, 203. [Google Scholar] [CrossRef]
- Pasinetti, G. Role of grape seed polyphenols in Alzheimer’s disease neuropathology. Nutr. Diet. Suppl. 2010, 2, 97–103. [Google Scholar] [CrossRef]
- Agrawal, M. Molecular basis of chronic neurodegeneration. In Clinical Molecular Medicine: Principles and Practice; Academic Press: Cambridge, MA, USA, 2019; pp. 447–460. [Google Scholar] [CrossRef]
- Jellinger, K.A. General aspects of neurodegeneration. J. Neural Transm. Suppl. 2003, 65, 101–144. [Google Scholar] [CrossRef]
- Callens, M.; Kraskovskaya, N.; Derevtsova, K.; Annaert, W.; Bultynck, G.; Bezprozvanny, I.; Vervliet, T. The role of Bcl-2 proteins in modulating neuronal Ca2+ signaling in health and in Alzheimer’s disease. Biochim. Biophys. Acta-Mol. Cell Res. 2021, 1868, 118997. [Google Scholar] [CrossRef] [PubMed]
- Gong, B.; Radulovic, M.; Figueiredo-Pereira, M.E.; Cardozo, C. The ubiquitin-proteasome system: Potential therapeutic targets for alzheimer’s disease and spinal cord injury. Front. Mol. Neurosci. 2016, 9, 4. [Google Scholar] [CrossRef]
- Chitnis, T.; Weiner, H.L. CNS inflammation and neurodegeneration. J. Clin. Invest. 2017, 127, 3577–3587. [Google Scholar] [CrossRef]
- Essa, M.M.; Subash, S.; Akbar, M.; Al-Adawi, S.; Guillemin, G.J. Long-Term dietary supplementation of pomegranates, figs and dates alleviate neuroinflammation in a transgenic mouse model of alzheimer’s disease. PLoS ONE 2015, 10, e0120964. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, H.; Liu, J.; Chen, R.; Tang, Y.; Chen, H.; Gu, L.; Li, M.; Cao, S.; Qin, D.; et al. Inhibitory effect of lychee seed saponins on apoptosis induced by Aβ25-35 through regulation of the apoptotic and NF-κB pathways in PC12 cells. Nutrients 2017, 9, 337. [Google Scholar] [CrossRef]
- Lee, J.H.; Yang, D.S.; Goulbourne, C.N.; Im, E.; Stavrides, P.; Pensalfini, A.; Chan, H.; Bouchet-Marquis, C.; Bleiwas, C.; Berg, M.J.; et al. Faulty autolysosome acidification in Alzheimer’s disease mouse models induces autophagic build-up of Aβ in neurons, yielding senile plaques. Nat. Neurosci. 2022, 25, 688–701. [Google Scholar] [CrossRef]
- Fan, T.S.; Liu, S.C.H.; Wu, R.M. Alpha-synuclein and cognitive decline in parkinson disease. Life 2021, 11, 1239. [Google Scholar] [CrossRef] [PubMed]
- Mohammad-Beigi, H.; Aliakbari, F.; Sahin, C.; Lomax, C.; Tawfike, A.; Schafer, N.P.; Amiri-Nowdijeh, A.; Eskandari, H.; Møller, I.M.; Hosseini-Mazinani, M.; et al. Oleuropein derivatives from olive fruit extracts reduce—Synuclein fibrillation and oligomer toxicity. J. Biol. Chem. 2019, 294, 4215–4232. [Google Scholar] [CrossRef] [PubMed]
- Singh, E.; Devasahayam, G. Neurodegeneration by oxidative stress: A review on prospective use of small molecules for neuroprotection. Mol. Biol. Rep. 2020, 47, 3133–3140. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative stress: A key modulator in neurodegenerative diseases. Molecules 2019, 24, 1583. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhou, T.; Ziegler, A.C.; Dimitrion, P.; Zuo, L. Oxidative Stress in Neurodegenerative Diseases: From Molecular Mechanisms to Clinical Applications. Oxid. Med. Cell. Longev. 2017, 2017, 2525967. [Google Scholar] [CrossRef]
- Cenini, G.; Lloret, A.; Cascella, R. Oxidative stress in neurodegenerative diseases: From a mitochondrial point of view. Oxid. Med. Cell. Longev. 2019, 2019, 2105607. [Google Scholar] [CrossRef]
- Moreira, P.I. Mitochondrial dysfunction and oxidative stress in Alzheimer’s disease. Eur. Neurol. Rev. 2010, 5, 17–21. [Google Scholar] [CrossRef]
- Bonda, D.J.; Wang, X.; Perry, G.; Nunomura, A.; Tabaton, M.; Zhu, X.; Smith, M.A. Oxidative stress in Alzheimer disease: A possibility for prevention. Neuropharmacology 2010, 59, 290–294. [Google Scholar] [CrossRef]
- Chiaino, E.; Micucci, M.; Cosconati, S.; Novellino, E.; Budriesi, R.; Chiarini, A.; Frosini, M. Olive leaves and hibiscus flowers extracts-based preparation protect brain from oxidative stress-induced injury. Antioxidants 2020, 9, 806. [Google Scholar] [CrossRef]
- Sanchez-Martinez, J.D.; Bueno, M.; Alvarez-Rivera, G.; Tudela, J.; Ibanez, E.; Cifuentes, A. In vitro neuroprotective potential of terpenes from industrial orange juice by-products. Food Funct. 2021, 12, 302–314. [Google Scholar] [CrossRef]
- Quintero-Espinosa, D.A.; Ortega-Arellano, H.F.; Velez-Pardo, C.; Jimenez-Del-Rio, M. Phenolic-rich extract of avocado Persea americana (var. Colinred) peel blunts paraquat/maneb-induced apoptosis through blocking phosphorylation of LRRK2 kinase in human nerve-like cells. Environ. Toxicol. 2022, 37, 660–676. [Google Scholar] [CrossRef]
- Glass, C.K.; Saijo, K.; Winner, B.; Marchetto, M.C.; Gage, F.H. Mechanisms Underlying Inflammation in Neurodegeneration. Cell 2010, 140, 918–934. [Google Scholar] [CrossRef]
- Domínguez-Avila, J.A.; Salazar-López, N.J.; Montiel-Herrera, M.; Martínez-Martínez, A.; Villegas-Ochoa, M.A.; González-Aguilar, G.A. Phenolic compounds can induce systemic and central immunomodulation, which result in a neuroprotective effect. J. Food Biochem. 2022, 46, e14260. [Google Scholar] [CrossRef]
- Chen, W.W.; Zhang, X.; Huang, W.J. Role of neuroinflammation in neurodegenerative diseases (Review). Mol. Med. Rep. 2016, 13, 3391–3396. [Google Scholar] [CrossRef]
- Fiebich, B.L.; Batista, C.R.A.; Saliba, S.W.; Yousif, N.M.; de Oliveira, A.C.P. Role of microglia TLRs in neurodegeneration. Front. Cell. Neurosci. 2018, 12, 329. [Google Scholar] [CrossRef] [PubMed]
- Phochantachinda, S.; Chatchaisak, D.; Temviriyanukul, P.; Chansawang, A.; Pitchakarn, P.; Chantong, B. Ethanolic Fruit Extract of Emblica officinalis Suppresses Neuroinflammation in Microglia and Promotes Neurite Outgrowth in Neuro2a Cells. Evid.-Based Complement. Altern. Med. 2021, 2021, 6405987. [Google Scholar] [CrossRef]
- Spagnuolo, C.; Moccia, S.; Russo, G.L. Anti-inflammatory effects of flavonoids in neurodegenerative disorders. Eur. J. Med. Chem. 2018, 153, 105–115. [Google Scholar] [CrossRef]
- AlFadly, E.D.; Elzahhar, P.A.; Tramarin, A.; Elkazaz, S.; Shaltout, H.; Abu-Serie, M.M.; Janockova, J.; Soukup, O.; Ghareeb, D.A.; El-Yazbi, A.F.; et al. Tackling neuroinflammation and cholinergic deficit in Alzheimer’s disease: Multi-target inhibitors of cholinesterases, cyclooxygenase-2 and 15-lipoxygenase. Eur. J. Med. Chem. 2019, 167, 161–186. [Google Scholar] [CrossRef]
- Kempuraj, D.; Thangavel, R.; Natteru, P.A.; Selvakumar, G.P.; Saeed, D.; Zahoor, H.; Zaheer, S.; Iyer, S.S.; Zaheer, A. Neuroinflammation Induces Neurodegeneration. J. Neurol. Neurosurg. Spine 2016, 1, 1003. [Google Scholar] [PubMed]
- Vanhauwaert, R.; Bharat, V.; Wang, X. Surveillance and transportation of mitochondria in neurons. Curr. Opin. Neurobiol. 2019, 57, 87–93. [Google Scholar] [CrossRef]
- Stanga, S.; Caretto, A.; Boido, M.; Vercelli, A. Mitochondrial dysfunctions: A red thread across neurodegenerative diseases. Int. J. Mol. Sci. 2020, 21, 3719. [Google Scholar] [CrossRef]
- Naoi, M.; Wu, Y.; Shamoto-Nagai, M.; Maruyama, W. Mitochondria in Neuroprotection by Phytochemicals: Bioactive Polyphenols Modulate Mitochondrial Apoptosis System, Function and Structure. Int. J. Mol. Sci. 2019, 20, 2451. [Google Scholar] [CrossRef]
- Kung, H.-C.; Lin, K.-J.; Kung, C.; Lin, T. Oxidative Stress, Mitochondrial Dysfunction, and Neuroprotection of Polyphenols with Respect to Resveratrol in Parkinson’s Disease. Biomedicines 2021, 9, 918. [Google Scholar] [CrossRef] [PubMed]
- Abbaszadeh, F.; Fakhri, S.; Khan, H. Targeting apoptosis and autophagy following spinal cord injury: Therapeutic approaches to polyphenols and candidate phytochemicals. Pharmacol. Res. 2020, 160, 105069. [Google Scholar] [CrossRef]
- Cao, S.; Aman, Y.; Fang, E.F.; Tencomnao, T.P. Edulis Extract Protects Against Amyloid-β Toxicity in Alzheimer’s Disease Models Through Maintenance of Mitochondrial Homeostasis via the FOXO3/DAF-16 Pathway. Mol. Neurobiol. 2022, 59, 5612–5629. [Google Scholar] [CrossRef]
- Dutta, A.; Phukan, B.C.; Roy, R.; Mazumder, M.K.; Paul, R.; Choudhury, A.; Kumar, D.; Bhattacharya, P.; Nath, J.; Kumar, S.; et al. Garcinia morella extract confers dopaminergic neuroprotection by mitigating mitochondrial dysfunctions and inflammation in mouse model of Parkinson’s disease. Metab. Brain Dis. 2022, 37, 1887–1900. [Google Scholar] [CrossRef] [PubMed]
- Essa, M.M.; Subash, S.; Dhanalakshmi, C.; Manivasagam, T.; Al-Adawi, S.; Guillemin, G.J.; Thenmozhi, A.J. Dietary Supplementation of Walnut Partially Reverses 1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine Induced Neurodegeneration in a Mouse Model of Parkinson’s Disease. Neurochem. Res. 2015, 40, 1283–1293. [Google Scholar] [CrossRef]
- Iovino, L.; Tremblay, M.E.; Civiero, L. Glutamate-induced excitotoxicity in Parkinson’s disease: The role of glial cells. J. Pharmacol. Sci. 2020, 144, 151–164. [Google Scholar] [CrossRef]
- Du, X.; Wang, X.; Geng, M. Alzheimer’s disease hypothesis and related therapies. Transl. Neurodegener. 2018, 7, 1–7. [Google Scholar] [CrossRef]
- Schulz, M.; Gonzaga, L.V.; de Souza, V.; Farina, M.; Vitali, L.; Micke, G.A.; Costa, A.C.O.; Fett, R. Neuroprotective effect of juçara (Euterpe edulis Martius) fruits extracts against glutamate-induced oxytosis in HT22 hippocampal cells. Food Res. Int. 2019, 120, 114–123. [Google Scholar] [CrossRef]
- Wen, L.; He, M.; Yin, C.; Jiang, Y.; Luo, D.; Yang, B. Phenolics in Citrus aurantium fruit identified by UHPLC-MS/MS and their bioactivities. LWT 2021, 147, 111671. [Google Scholar] [CrossRef]
- Liao, R.; Wood, T.R.; Nance, E. Nanotherapeutic modulation of excitotoxicity and oxidative stress in acute brain injury. Nanobiomedicine 2020, 7, 1–18. [Google Scholar] [CrossRef]
- Cheng, Y.J.; Lin, C.H.; Lane, H.Y. Involvement of cholinergic, adrenergic, and glutamatergic network modulation with cognitive dysfunction in alzheimer’s disease. Int. J. Mol. Sci. 2021, 22, 2283. [Google Scholar] [CrossRef] [PubMed]
- Olloquequi, J.; Cornejo-Córdova, E.; Verdaguer, E.; Soriano, F.X.; Binvignat, O.; Auladell, C.; Camins, A. Excitotoxicity in the pathogenesis of neurological and psychiatric disorders: Therapeutic implications. J. Psychopharmacol. 2018, 32, 265–275. [Google Scholar] [CrossRef]
- Kamda, J.L.D.; Boiangiu, R.S.; Brinza, I.; Djoumessi, L.B.K.; Rebe, R.N.; Kamleu, B.N.; Guedang, S.D.N.; Camdi, G.W.; Bouvourné, P.; Keugong, E.W.; et al. Neuroprotective Potential of Guiera senegalensis (Combretaceae) Leaf Hydroethanolic Extract against Cholinergic System Dysfunctions and Oxidative Stress in Scopolamine-Induced Cognitive Impairment in Zebrafish (Danio rerio). Plants 2022, 11, 1149. [Google Scholar] [CrossRef]
- Yadav, E.; Singh, D.; Debnath, B.; Rathee, P.; Yadav, P.; Verma, A. Molecular Docking and Cognitive Impairment Attenuating Effect of Phenolic Compound Rich Fraction of Trianthema portulacastrum in Scopolamine Induced Alzheimer’s Disease Like Condition. Neurochem. Res. 2019, 44, 1665–1677. [Google Scholar] [CrossRef]
- Temviriyanukul, P.; Sritalahareuthai, V.; Promyos, N.; Thangsiri, S.; Pruesapan, K.; Srinuanchai, W.; Nuchuchua, O.; Siriwan, D.; On-Nom, N.; Suttisansanee, U. The Effect of Sacred Lotus (Nelumbo nucifera) and Its Mixtures on Phenolic Profiles, Antioxidant Activities, and Inhibitions of the Key Enzymes Relevant to Alzheimer’s Disease. Molecules 2020, 25, 3713. [Google Scholar] [CrossRef]
- Fakhri, S.; Abbaszadeh, F.; Moradi, S.Z.; Cao, H.; Khan, H.; Xiao, J. Effects of Polyphenols on Oxidative Stress, Inflammation, and Interconnected Pathways during Spinal Cord Injury. Oxid. Med. Cell. Longev. 2022, 2022, 8100195. [Google Scholar] [CrossRef]
- Thomford, N.E.; Senthebane, D.A.; Rowe, A.; Munro, D.; Seele, P.; Maroyi, A.; Dzobo, K. Natural products for drug discovery in the 21st century: Innovations for novel drug discovery. Int. J. Mol. Sci. 2018, 19, 1578. [Google Scholar] [CrossRef]
- Teleanu, R.I.; Chircov, C.; Grumezescu, A.M.; Volceanov, A.; Teleanu, D.M. Antioxidant therapies for neuroprotection-a review. J. Clin. Med. 2019, 8, 1659. [Google Scholar] [CrossRef]
- Szwajgier, D.; Borowiec, K.; Pustelniak, K. The neuroprotective effects of phenolic acids: Molecular mechanism of action. Nutrients 2017, 9, 477. [Google Scholar] [CrossRef]
- Zhang, C.; Guan, J.; Zhang, J.; Yang, J.; Wang, X.; Peng, X. Protective effects of three structurally similar polyphenolic compounds against oxidative damage and their binding properties to human serum albumin. Food Chem. 2021, 349, 129118. [Google Scholar] [CrossRef]
- Cázares-Camacho, R.; Domínguez-Avila, J.A.; Astiazarán-García, H.; Montiel-Herrera, M.; González-Aguilar, G.A. Neuroprotective effects of mango cv. ‘Ataulfo’ peel and pulp against oxidative stress in streptozotocin-induced diabetic rats. J. Sci. Food Agric. 2021, 101, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Oboh, G.; Odubanjo, V.O.; Bello, F.; Ademosun, A.O.; Oyeleye, S.I.; Nwanna, E.E.; Ademiluyi, A.O. Aqueous extracts of avocado pear (Persea americana Mill.) leaves and seeds exhibit anti-cholinesterases and antioxidant activities in vitro. J. Basic Clin. Physiol. Pharmacol. 2016, 27, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Chhoud, R.; Montero, F.V.; Haj Romdhane, M.; Majdoub, H.; Duran Ogalla, R. Phytochemical and Bioactivities of Male Flower Buds of Fruit Trees from the Southern Tunisia: Polyphenols UPLC-MS Profiles and Antioxidant Enzymatic Potential in Human Plasma of Parkinson’s Disease Patients. Chem. Africa 2022, 5, 1337–1350. [Google Scholar] [CrossRef]
- da Silva, G.G.; Pimenta, L.P.S.; Melo, J.O.F.; Mendonça, H.D.O.P.; Augusti, R.; Takahashi, J.A. Phytochemicals of Avocado Residues as Potential Acetylcholinesterase Inhibitors, Antioxidants, and Neuroprotective Agents. Molecules 2022, 27, 1892. [Google Scholar] [CrossRef]
- Zhao, Y.; Zeng, Y.; Wu, A.; Yu, C.; Tang, Y.; Wang, X.; Xiong, R.; Chen, H.; Wu, J.; Qin, D. Lychee seed fraction inhibits Aβ(1-42)-induced neuroinflammation in BV-2 cells via NF-κB signaling pathway. Front. Pharmacol. 2018, 9, 380. [Google Scholar] [CrossRef]
- Tang, Y.; Xiong, R.; Wu, A.G.; Yu, C.L.; Zhao, Y.; Qiu, W.Q.; Wang, X.L.; Teng, J.F.; Liu, J.; Chen, H.X.; et al. Polyphenols derived from lychee seed suppress Aβ (1-42)-induced neuroinflammation. Int. J. Mol. Sci. 2018, 19, 2109. [Google Scholar] [CrossRef]
- Savla, P.; Das, G.; Mondal, P.; Gajbhiye, R.L.; Jaisankar, P.; Ghosh, S. Methanolic Extract of Papaya Leaves Shows Neuroprotective Effect. ChemistrySelect 2017, 2, 9454–9457. [Google Scholar] [CrossRef]
- Al-Radadi, N.S. Biogenic proficient synthesis of (Au-NPs) via aqueous extract of Red Dragon Pulp and seed oil: Characterization, antioxidant, cytotoxic properties, anti-diabetic anti-inflammatory, anti-Alzheimer and their anti-proliferative potential against cancer cell. Saudi J. Biol. Sci. 2022, 29, 2836–2855. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Rodríguez, G.; Ramón Vidal, D.; Martorell, P.; Plaza, M.; Marina, M.L. Composition of Nonextractable Polyphenols from Sweet Cherry Pomace Determined by DART-Orbitrap-HRMS and Their In Vitro and In Vivo Potential Antioxidant, Antiaging, and Neuroprotective Activities. J. Agric. Food Chem. 2022, 70, 7993–8009. [Google Scholar] [CrossRef]
- Debnath-Canning, M.; Unruh, S.; Vyas, P.; Daneshtalab, N.; Igamberdiev, A.U.; Weber, J.T. Fruits and leaves from wild blueberry plants contain diverse polyphenols and decrease neuroinflammatory responses in microglia. J. Funct. Foods 2020, 68, 103906. [Google Scholar] [CrossRef]
- El-Hawary, S.S.; Sayed, A.M.; Issa, M.Y.; Ebrahim, H.S.; Alaaeldin, R.; Elrehany, M.A.; Abd El-Kadder, E.M.; Abdelmohsen, U.R. Anti-Alzheimer chemical constituents of: Morus macroura Miq.: Chemical profiling, in silico and in vitro investigations. Food Funct. 2021, 12, 8078–8089. [Google Scholar] [CrossRef] [PubMed]
- Nantacharoen, W.; Baek, S.J.; Plaingam, W.; Charoenkiatkul, S.; Tencomnao, T.; Sukprasansap, M. Cleistocalyx nervosum var. paniala Berry Promotes Antioxidant Response and Suppresses Glutamate-Induced Cell Death via SIRT1/Nrf2 Survival Pathway in Hippocampal HT22 Neuronal Cells. Molecules 2022, 27, 5813. [Google Scholar] [CrossRef] [PubMed]
- Duangjan, C.; Rangsinth, P.; Zhang, S.; Gu, X.; Wink, M.; Tencomnao, T. Vitis Vinifera Leaf Extract Protects Against Glutamate-Induced Oxidative Toxicity in HT22 Hippocampal Neuronal Cells and Increases Stress Resistance Properties in Caenorhabditis Elegans. Front. Nutr. 2021, 8, 634100. [Google Scholar] [CrossRef] [PubMed]
- Khokar, R.; Hachani, K.; Hasan, M.; Othmani, F.; Essam, M.; Al Mamari, A.; UM, D.; Khan, S.A. Anti-Alzheimer potential of a waste by-product (peel) of Omani pomegranate fruits: Quantification of phenolic compounds, in-vitro antioxidant, anti-cholinesterase and in-silico studies. Biocatal. Agric. Biotechnol. 2021, 38, 102223. [Google Scholar] [CrossRef]
- Angeloni, S.; Freschi, M.; Marrazzo, P.; Hrelia, S.; Beghelli, D.; Juan-García, A.; Juan, C.; Caprioli, G.; Sagratini, G.; Angeloni, C. Antioxidant and Anti-Inflammatory Profiles of Spent Coffee Ground Extracts for the Treatment of Neurodegeneration. Oxid. Med. Cell. Longev. 2021, 2021, 6620913. [Google Scholar] [CrossRef] [PubMed]
- Savić, A.; Aradski, A.A.; Živković, J.; Šavikin, K.; Jarić, S.; Marin, P.D.; Duletić-Laušević, S. Phenolic composition, and antioxidant and antineurodegenerative potential of methanolic extracts of fruit peel and flesh of pear varieties from Serbia. Polish J. Food Nutr. Sci. 2021, 71, 225–236. [Google Scholar] [CrossRef]
- Olofinsan, K.A.; Salau, V.F.; Erukainure, O.L.; Islam, M.S. Harpephyllum caffrum fruit (wild plum) facilitates glucose uptake and modulates metabolic activities linked to neurodegeneration in isolated rat brain: An in vitro and in silico approach. J. Food Biochem. 2022, 46, e14177. [Google Scholar] [CrossRef]
- Hua, H.; Zhang, W.; Li, J.; Li, J.; Liu, C.; Guo, Y.; Cheng, Y.; Pi, F.; Xie, Y.; Yao, W.; et al. Neuroprotection against cerebral ischemia/reperfusion by dietary phytochemical extracts from Tibetan turnip (Brassica rapa L.). J. Ethnopharmacol. 2021, 265, 113410. [Google Scholar] [CrossRef]
- Ben Rejeb, I.; Dhen, N.; Gargouri, M.; Boulila, A. Chemical Composition, Antioxidant Potential and Enzymes Inhibitory Properties of Globe Artichoke By-Products. Chem. Biodivers. 2020, 17, e2000073. [Google Scholar] [CrossRef]
- Ajiboye, B.O.; Ojo, O.A.; Okesola, M.A.; Akinyemi, A.J.; Talabi, J.Y.; Idowu, O.T.; Fadaka, A.O.; Boligon, A.A.; Anraku de Campos, M.M. In vitro antioxidant activities and inhibitory effects of phenolic extract of Senecio biafrae (Oliv and Hiern) against key enzymes linked with type II diabetes mellitus and Alzheimer’s disease. Food Sci. Nutr. 2018, 6, 1803–1810. [Google Scholar] [CrossRef]
- Gao, S.H.; Zhao, T.R.; Liu, Y.P.; Wang, Y.F.; Cheng, G.G.; Cao, J.X. Phenolic constituents, antioxidant activity and neuroprotective effects of ethanol extracts of fruits, leaves and flower buds from Vaccinium dunalianum Wight. Food Chem. 2022, 374, 131752. [Google Scholar] [CrossRef] [PubMed]
- Diniso, T.; Adeyemi, J.; Oriola, A.; Elufioye, T.; Gondwe, M.; Oyedeji, A. Polyphenolic Contents, Free Radical Scavenging and Cholinesterase Inhibitory Activities of Dalbergiella welwitschii Leaf Extracts. Plants 2022, 11, 2066. [Google Scholar] [CrossRef] [PubMed]
- Pratiksha, V.B.; Anand, T.; Mohan Manu, T.; Sharath Babu, G.R.; Mahantesh, M.P. Mucuna pruriens Seed Extract: A Possible Protective Agent Against Ochratoxin A Neurodegeneration. Rev. Bras. Farmacogn. 2022, 32, 395–409. [Google Scholar] [CrossRef]
- Lee, W.S.; Lee, H.J.; Yang, J.Y.; Shin, H.L.; Choi, S.W.; Kim, J.K.; Seo, W.D.; Kim, E.H. The Potential Neuroprotective Effects of Extracts from Oat Seedlings against Alzheimer’s Disease. Nutrients 2022, 14, 4103. [Google Scholar] [CrossRef]
- Woo, S.Y.; Yang, J.Y.; Lee, H.G.; Ahn, H.J.; Lee, Y.B.; Do, S.H.; Kim, J.Y.; Seo, W.D. Changes in metabolites with harvest times of seedlings of various Korean oat (Avena sativa L.) cultivars and their neuraminidase inhibitory effects. Food Chem. 2022, 373, 131429. [Google Scholar] [CrossRef]
- Ravi, S.K.; Narasingappa, R.B.; Joshi, C.G.; Girish, T.K.; Vincent, B. Neuroprotective effects of Cassia tora against paraquatinduced neurodegeneration: Relevance for Parkinson’s disease. Nat. Prod. Res. 2018, 32, 1476–1480. [Google Scholar] [CrossRef]
- Kim, Y.J.; Sohn, E.; Kim, J.H.; Na, M.K.; Jeong, S.J. Catechol-Type Flavonoids from the Branches of Elaeagnus glabra f. oxyphylla Exert Antioxidant Activity and an Inhibitory Effect on Amyloid-β Aggregation. Molecules 2020, 25, 4917. [Google Scholar] [CrossRef]
- Infante-Garcia, C.; Jose Ramos-Rodriguez, J.; Marin-Zambrana, Y.; Teresa Fernandez-Ponce, M.; Casas, L.; Mantell, C.; Garcia-Alloza, M. Mango leaf extract improves central pathology and cognitive impairment in a type 2 diabetes mouse model. Brain Pathol. 2017, 27, 499–507. [Google Scholar] [CrossRef]
- Wang, X.; Wu, J.; Yu, C.; Tang, Y.; Liu, J.; Chen, H.; Jin, B.; Mei, Q.; Cao, S.; Qin, D. Lychee seed saponins improve cognitive function and prevent neuronal injury via inhibiting neuronal apoptosis in a rat model of Alzheimer’s disease. Nutrients 2017, 9, 105. [Google Scholar] [CrossRef]
- Xiong, R.; Zhou, X.G.; Tang, Y.; Wu, J.M.; Sun, Y.S.; Teng, J.F.; Pan, R.; Law, B.Y.K.; Zhao, Y.; Qiu, W.Q.; et al. Lychee seed polyphenol protects the blood–brain barrier through inhibiting Aβ(25–35)-induced NLRP3 inflammasome activation via the AMPK/mTOR/ULK1-mediated autophagy in bEnd.3 cells and APP/PS1 mice. Phyther. Res. 2021, 35, 954–973. [Google Scholar] [CrossRef]
- Adedayo, B.C.; Oyeleye, S.I.; Okeke, B.M.; Oboh, G. Anti-cholinesterase and antioxidant properties of alkaloid and phenolic-rich extracts from pawpaw (Carica papaya) leaf: A comparative study. Flavour Fragr. J. 2021, 36, 47–54. [Google Scholar] [CrossRef]
- Phachonpai, W.; Tongun, T. Cognition enhancing effects of Clausena lansium (Lour.) peel extract attenuate chronic restraint stress-induced memory deficit in rats. Heliyon 2021, 7, e07003. [Google Scholar] [CrossRef]
- Prasad, K.N.; Xie, H.; Hao, J.; Yang, B.; Qiu, S.; Wei, X.; Chen, F.; Jiang, Y. Antioxidant and anticancer activities of 8-hydroxypsoralen isolated from wampee [Clausena lansium (Lour.) Skeels] peel. Food Chem. 2010, 118, 62–66. [Google Scholar] [CrossRef]
- Gu, P.S.; Moon, M.; Choi, J.G.; Oh, M.S. Mulberry fruit ameliorates Parkinson’s-disease-related pathology by reducing α-synuclein and ubiquitin levels in a 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine/probenecid model. J. Nutr. Biochem. 2017, 39, 15–21. [Google Scholar] [CrossRef]
- Zhang, W.; Han, F.; He, J.; Duan, C. HPLC-DAD-ESI-MS/MS analysis and antioxidant activities of nonanthocyanin phenolics in Mulberry (Morus alba L.). J. Food Sci. 2008, 73, C512–C518. [Google Scholar] [CrossRef]
- Shin, S.K.; Yoo, J.M.; Li, F.Y.; Baek, S.Y.; Kim, M.R. Mulberry fruit improves memory in scopolamine-treated mice: Role of cholinergic function, antioxidant system, and TrkB/Akt signaling. Nutr. Neurosci. 2021, 24, 940–950. [Google Scholar] [CrossRef]
- Wattanathorn, J.; Palachai, N.; Thukham-Mee, W.; Muchimapura, S. Memory-Enhancing Effect of a Phytosome Containing the Combined Extract of Mulberry Fruit and Ginger in an Animal Model of Ischemic Stroke with Metabolic Syndrome. Oxid. Med. Cell. Longev. 2020, 2020, 3096826. [Google Scholar] [CrossRef]
- Flanagan, E.; Cameron, D.; Sobhan, R.; Wong, C.; Pontifex, M.G.; Tosi, N.; Mena, P.; Del Rio, D.; Sami, S.; Narbad, A.; et al. Chronic Consumption of Cranberries (Vaccinium macrocarpon) for 12 Weeks Improves Episodic Memory and Regional Brain Perfusion in Healthy Older Adults: A Randomised, Placebo-Controlled, Parallel-Groups Feasibility Study. Front. Nutr. 2022, 9, 849902. [Google Scholar] [CrossRef]
- Abdou, H.M.; Wahby, M.M. Neuroprotection of Grape Seed Extract and Pyridoxine against Triton-Induced Neurotoxicity. Oxid. Med. Cell. Longev. 2016, 2016, 8679506. [Google Scholar] [CrossRef]
- Wang, J.; Ho, L.; Zhao, W.; Ono, K.; Rosensweig, C.; Chen, L.; Humala, N.; Teplow, D.B.; Pasinetti, G.M. Grape-derived polyphenolics prevent Aβ oligomerization and attenuate cognitive deterioration in a mouse model of Alzheimer’s disease. J. Neurosci. 2008, 28, 6388–6392. [Google Scholar] [CrossRef]
- Ben Youssef, S.; Brisson, G.; Doucet-Beaupré, H.; Castonguay, A.M.; Gora, C.; Amri, M.; Lévesque, M. Neuroprotective benefits of grape seed and skin extract in a mouse model of Parkinson’s disease. Nutr. Neurosci. 2021, 24, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Sheng, K.; Shui, S.; Yan, L.; Yu, J.; Hao, G.; Qu, H.; Liu, J.; Zhang, Y.; Liu, C.; Zheng, L. The beneficial effects of dietary grape supplementation on improving cognitive deficits in APP/PS1 double transgenic mice. J. Funct. Foods 2018, 49, 224–234. [Google Scholar] [CrossRef]
- Aravind, A.P.A.; Menon, L.N.; Rameshkumar, K.B. Structural diversity of secondary metabolites in Garcinia species. In Diversity of Garcinia Species Western Ghats: Pythochemical Perspect; Jawaharlal Nehru Tropical Botanic Garden and Research Institute: Kerala, India, 2016; pp. 19–75. [Google Scholar]
- Abou Baker, D.H.; Ibrahim, B.M.M.; Hassan, N.S.; Yousuf, A.F.; Gengaihi, S. El Exploiting Citrus aurantium seeds and their secondary metabolites in the management of Alzheimer disease. Toxicol. Reports 2020, 7, 723–729. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, S.; Kotani, Y.; Yamamoto, K.; Sawamoto, A.; Sugawara, K.; Sudo, M.; Ohkubo, Y.; Tamanaha, A.; Nakajima, M.; Furukawa, Y. The peel of Citrus kawachiensis (kawachi bankan) ameliorates microglial activation, tau hyper-phosphorylation, and suppression of neurogenesis in the hippocampus of senescence-accelerated mice. Biosci. Biotechnol. Biochem. 2018, 82, 869–878. [Google Scholar] [CrossRef]
- Okuyama, S.; Kanzaki, T.; Kotani, Y.; Katoh, M.; Sawamoto, A.; Nakajima, M.; Furukawa, Y. Continual treatment with the peels of citrus kawachiensis (kawachi bankan) protects against dopaminergic neuronal cell death in a lipopolysaccharide-induced model of parkinson’s disease. J. Nutr. Sci. Vitaminol. 2019, 65, 205–208. [Google Scholar] [CrossRef]
- Okuyama, S.; Shinoka, W.; Nakamura, K.; Kotani, M.; Sawamoto, A.; Sugawara, K.; Sudo, M.; Nakajima, M.; Furukawa, Y. Suppressive effects of the peel of Citrus kawachiensis (Kawachi Bankan) on astroglial activation, tau phosphorylation, and inhibition of neurogenesis in the hippocampus of type 2 diabetic db/db mice. Biosci. Biotechnol. Biochem. 2018, 82, 1384–1395. [Google Scholar] [CrossRef]
- Okuyama, S.; Yamamoto, K.; Mori, H.; Sawamoto, A.; Amakura, Y.; Yoshimura, M.; Tamanaha, A.; Ohkubo, Y.; Sugawara, K.; Sudo, M.; et al. Neuroprotective effect of Citrus kawachiensis (Kawachi Bankan) peels, a rich source of naringin, against global cerebral ischemia/reperfusion injury in mice. Biosci. Biotechnol. Biochem. 2018, 82, 1216–1224. [Google Scholar] [CrossRef]
- Morzelle, M.C.; Salgado, J.M.; Telles, M.; Mourelle, D.; Bachiega, P.; Buck, H.S.; Viel, T.A. Neuroprotective effects of pomegranate peel extract after chronic infusion with amyloid-β peptide in mice. PLoS ONE 2016, 11, e0166123. [Google Scholar] [CrossRef]
- Pfohl, M.; DaSilva, N.A.; Marques, E.; Agudelo, J.; Liu, C.; Goedken, M.; Slitt, A.L.; Seeram, N.P.; Ma, H. Hepatoprotective and anti-inflammatory effects of a standardized pomegranate (Punica granatum) fruit extract in high fat diet-induced obese C57BL/6 mice. Int. J. Food Sci. Nutr. 2021, 72, 499–510. [Google Scholar] [CrossRef]
- Subash, S.; Essa, M.M.; Braidy, N.; Al-Jabri, A.; Vaishnav, R.; Al-Adawi, S.; Al-Asmi, A.; Guillemin, G.J. Consumption of fig fruits grown in Oman can improve memory, anxiety, and learning skills in a transgenic mice model of Alzheimer’s disease. Nutr. Neurosci. 2016, 19, 475–483. [Google Scholar] [CrossRef]
- Pujari, R.R.; Vyawahare, N.S.; Kagathara, V.G. Evaluation of antioxidant and neuroprotective effect of date palm (Phoenix dactylifera L.) against bilateral common carotid artery occlusion in rats. Indian J. Exp. Biol. 2011, 49, 627–633. [Google Scholar] [PubMed]
- Ichwan, M.; Walker, T.L.; Nicola, Z.; Ludwig-Müller, J.; Böttcher, C.; Overall, R.W.; Adusumilli, V.S.; Bulut, M.; Sykes, A.M.; Hübner, N.; et al. Apple Peel and Flesh Contain Pro-neurogenic Compounds. Stem Cell Rep. 2021, 16, 548–565. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yu, Z.; Xia, J.; Zhang, X.; Liu, K.; Sik, A.; Jin, M. Anti-Parkinson’s disease activity of phenolic acids from: Eucommia ulmoides Oliver leaf extracts and their autophagy activation mechanism. Food Funct. 2020, 11, 1425–1440. [Google Scholar] [CrossRef]
- Ravi, S.K.; Ramesh, B.N.; Mundugaru, R.; Vincent, B. Multiple pharmacological activities of Caesalpinia crista against aluminium-induced neurodegeneration in rats: Relevance for Alzheimer’s disease. Environ. Toxicol. Pharmacol. 2018, 58, 202–211. [Google Scholar] [CrossRef]
- Elseady, W.S.; Keshk, W.A.; Negm, W.A.; Elkhalawany, W.; Elhanafy, H.; Ibrahim, M.A.A.; Radwan, D.A. Saffron extract attenuates Sofosbuvir-induced retinal neurodegeneration in albino rat. Anat. Rec. 2022, 1–15. [Google Scholar] [CrossRef]
- Deng, X.; Zhang, S.; Wu, J.; Sun, X.; Shen, Z.; Dong, J.; Huang, J. Promotion of Mitochondrial Biogenesis via Activation of AMPK-PGC1ɑ Signaling Pathway by Ginger (Zingiber officinale Roscoe) Extract, and Its Major Active Component 6-Gingerol. J. Food Sci. 2019, 84, 2101–2111. [Google Scholar] [CrossRef] [PubMed]
- Henríquez, G.; Gomez, A.; Guerrero, E.; Narayan, M. Potential Role of Natural Polyphenols against Protein Aggregation Toxicity: In Vitro, In Vivo, and Clinical Studies. ACS Chem. Neurosci. 2020, 11, 2915–2934. [Google Scholar] [CrossRef]
- Bieschke, J. Natural Compounds May Open New Routes to Treatment of Amyloid Diseases. Neurotherapeutics 2013, 10, 429–439. [Google Scholar] [CrossRef] [PubMed]
- Elfiky, A.M.; Mahmoud, A.A.; Elreedy, H.A.; Ibrahim, K.S.; Ghazy, M.A. Quercetin stimulates the non-amyloidogenic pathway via activation of ADAM10 and ADAM17 gene expression in aluminum chloride-induced Alzheimer’s disease rat model. Life Sci. 2021, 285, 119964. [Google Scholar] [CrossRef] [PubMed]
- Kostomoiri, M.; Fragkouli, A.; Sagnou, M.; Skaltsounis, L.A.; Pelecanou, M.; Tsilibary, E.C.; Tzinia, A.K. Oleuropein, an anti-oxidant polyphenol constituent of olive promotes α-Secretase cleavage of the amyloid precursor protein (AβPP). Cell. Mol. Neurobiol. 2013, 33, 147–154. [Google Scholar] [CrossRef]
- Das, S.; Stark, L.; Musgrave, I.F.; Pukala, T.; Smid, S.D. Bioactive polyphenol interactions with β amyloid: A comparison of binding modelling, effects on fibril and aggregate formation and neuroprotective capacity. Food Funct. 2016, 7, 1138–1146. [Google Scholar] [CrossRef]
- Porat, Y.; Abramowitz, A.; Gazit, E. Inhibition of amyloid fibril formation by polyphenols: Structural similarity and aromatic interactions as a common inhibition mechanism. Chem. Biol. Drug Des. 2006, 67, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Dhouafli, Z.; Cuanalo-Contreras, K.; Hayouni, E.A.; Mays, C.E.; Soto, C.; Moreno-Gonzalez, I. Inhibition of protein misfolding and aggregation by natural phenolic compounds. Cell. Mol. Life Sci. 2018, 75, 3521–3538. [Google Scholar] [CrossRef] [PubMed]
- Hasanbašić, S.; Jahić, A.; Berbić, S.; Znidarič, M.T.; Zerovnik, E. Inhibition of protein aggregation by several antioxidants. Oxid. Med. Cell. Longev. 2018, 2018, 8613209. [Google Scholar] [CrossRef] [PubMed]
- Di Meo, F.; Valentino, A.; Petillo, O.; Peluso, G.; Filosa, S.; Crispi, S. Bioactive polyphenols and neuromodulation: Molecular mechanisms in neurodegeneration. Int. J. Mol. Sci. 2020, 21, 2564. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Awasthi, H.; Tota, S.; Hanif, K.; Nath, C.; Shukla, R. Protective effect of curcumin against intracerebral streptozotocin induced impairment in memory and cerebral blood flow. Life Sci. 2010, 86, 87–94. [Google Scholar] [CrossRef]
- Lim, H.J.; Lee, K.S.; Lee, S.; Park, J.H.; Choi, H.E.; Go, S.H.; Kwak, H.J.; Park, H.Y. 15d-PGJ2 stimulates HO-1 expression through p38 MAP kinase and Nrf-2 pathway in rat vascular smooth muscle cells. Toxicol. Appl. Pharmacol. 2007, 223, 20–27. [Google Scholar] [CrossRef]
- Bastianetto, S.; Ménard, C.; Quirion, R. Neuroprotective action of resveratrol. Biochim. Biophys. Acta-Mol. Basis Dis. 2015, 1852, 1195–1201. [Google Scholar] [CrossRef]
- Carregosa, D.; Carecho, R.; Figueira, I.; Santos, C.N. Low-Molecular Weight Metabolites from Polyphenols as Effectors for Attenuating Neuroinflammation. J. Agric. Food Chem. 2019, 68, 1790–1807. [Google Scholar] [CrossRef]
- Shih, R.H.; Wang, C.Y.; Yang, C.M. NF-kappaB signaling pathways in neurological inflammation: A mini review. Front. Mol. Neurosci. 2015, 8, 77. [Google Scholar] [CrossRef]
- Xie, Q.; Chen, Y.; Tan, H.; Liu, B.; Zheng, L.L.; Mu, Y. Targeting Autophagy with Natural Compounds in Cancer: A Renewed Perspective from Molecular Mechanisms to Targeted Therapy. Front. Pharmacol. 2021, 12, 748149. [Google Scholar] [CrossRef]
- Park, D.; Jeong, H.; Lee, M.N.; Koh, A.; Kwon, O.; Yang, Y.R.; Noh, J.; Suh, P.G.; Park, H.; Ryu, S.H. Resveratrol induces autophagy by directly inhibiting mTOR through ATP competition. Sci. Rep. 2016, 6, 21772. [Google Scholar] [CrossRef] [PubMed]
- Casamenti, F.; Stefani, M. Olive polyphenols: New promising agents to combat aging-associated neurodegeneration. Expert Rev. Neurother. 2017, 17, 345–358. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Teng, J.; Wei, B.; Huang, L.; Xia, N. Phenolic compounds, bioactivity, and bioaccessibility of ethanol extracts from passion fruit peel based on simulated gastrointestinal digestion. Food Chem. 2021, 356, 129682. [Google Scholar] [CrossRef]
- Fernandes, F.; Valentão, P.; Sousa, C.; Pereira, J.A.; Seabra, R.M.; Andrade, P.B. Chemical and antioxidative assessment of dietary turnip (Brassica rapa var. rapa L.). Food Chem. 2007, 105, 1003–1010. [Google Scholar] [CrossRef]
- Anantharaju, P.G.; Gowda, P.C.; Vimalambike, M.G.; Madhunapantula, S.V. An overview on the role of dietary phenolics for the treatment of cancers. Nutr. J. 2016, 15, 99. [Google Scholar] [CrossRef]
- Mikami, Y.; Yamazawa, T. Chlorogenic acid, a polyphenol in coffee, protects neurons against glutamate neurotoxicity. Life Sci. 2015, 139, 69–74. [Google Scholar] [CrossRef]
- Hung, Y.C.; Kuo, Y.H.; Hsieh, P.W.; Hsieh, T.Y.; Kuo, J.R.; Wang, S.J. Chlorogenic acid decreases glutamate release from rat cortical nerve terminals by p/q-type ca2+ channel suppression: A possible neuroprotective mechanism. Int. J. Mol. Sci. 2021, 22, 1447. [Google Scholar] [CrossRef]
- Wu, M.; Cai, J.; Fang, Z.; Li, S.; Huang, Z.; Tang, Z.; Luo, Q.; Chen, H. The Composition and Anti-Aging Activities of Polyphenol Extract from Phyllanthus emblica L. Fruit. Nutrients 2022, 14, 857. [Google Scholar] [CrossRef]
- Fernández-Ochoa, Á.; de la Luz Cádiz-Gurrea, M.; Fernández-Moreno, P.; Rojas-García, A.; Arráez-Román, D.; Segura-Carretero, A. Recent Analytical Approaches for the Study of Bioavailability and Metabolism of Bioactive Phenolic Compounds. Molecules 2022, 27, 777. [Google Scholar] [CrossRef] [PubMed]
- Murakami, A. Dose-dependent functionality and toxicity of green tea polyphenols in experimental rodents. Arch. Biochem. Biophys. 2014, 557, 3–10. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | By-Product | Main Compounds | Biological Target | Mechanisms of Action | Neuroprotective Effects | Citation |
|---|---|---|---|---|---|---|
| Avocado (Persea americana) | Seed + Peel | Caffeic acid, (epi)catechin, rutin, B- type procyanidins. | AChE inhibition. | Inhibition of AChE activity up to 65%. | Anti-AChE. | [78] |
| Seed + Leaf | Polyphenols. | AChE and BChE inhibition. | Remarkable enzymatic inhibition of AChE and BChE, avoiding acetylcholine breakdown and increasing communication between nerve cells. | Anti-AChE/-BChE. | [76] | |
| Passion Fruit (Passiflora edulis) | Pulp + Seed | Phenolic acids, flavonoids, stilbenes. Piceatannol. | Downregulation of enzymes. | AChE IC50 29.42 µg/mL and LOX IC50 27.682 µg/mL. | Anti-inflammatory and anti-AChE. | [20] |
| Lychee (Litchi chinensis) | Seed | Saponins. | Apoptosis and MMP. | Suppression of apoptosis in Aβ-induced cells through upregulation of Bcl-2 proteins. Decrease in caspase-3 mRNA expression and nuclear translocation of NF-kB. Improvement in mitochondrial function (MMP decrease). | Anti-apoptotic and anti-MMP. | [31] |
| A- and B-type procyanidins, rutin, quercetin, saponins. | Suppression of brain inflammation induced by amyloid aggregation. | Modulation of NF- κB signaling pathway. Reduce of mRNA and protein expressions of IL-1, TNF-α, COX-2 AND iNOS. Inhibition of cell apoptosis in microglial cells overactivated by Aβ deposition. | Anti-inflammatory and anti-apoptotic. | [79] | ||
| Catechin and procyanidin A2. | Suppression of brain inflammation induced by amyloid aggregation. | Inhibition of the neuroinflammatory effect induced by amyloid aggregation and deposition by modulation of NF-kB signaling pathway. | Anti-inflammatory. | [80] | ||
| Papaya (Carica papaya) | Leaf | Carpaine enriched (alkaloid). | Cytotoxicity and inhibition of Aβ42 aggregation. | Stabilization of intracellular microtubules and formation of hydrophobic bind to Aβ42 sequence, thus inhibiting Aβ42 peptide aggregation. | Anti-amyloidogenic and cell structural stabilizer. | [81] |
| Red Dragon Fruit (Hylocereus polyrhizus) | Pulp + Seed | Phenolic and organic acids, flavonoids and carotenoids. | AChE and BChE inhibition. | Inhibition of 69.11 ± 1.12% against AChE and 64. 78 ± 0.71% against BChE. | Anti-AChE/-BChE. | [82] |
| Sweet cherry (Prunus avium) | Pulp | Organic and phenolic acids, (epi)catechin, quercetin, luteolin and procyanidin B2. | AChE inhibition and antioxidant potential. | Inhibition of AChE activity and high antioxidant capacity. | Anti-AChE and antioxidant. | [83] |
| Blueberry (Vaccinium carymbosum) | Pulp + Leaf | Anthocyanins (Delphinidin and cyanidin). | Neuroinflammatory responses in microglia. | Antioxidant activity, reduction in inflammatory mediator levels. | Antioxidant and anti-inflammatory. | [84] |
| King white mulberry (Morus macroura) | Pulp + Leaf | Phenolic acids, flavonoids, terpenes, sterols, stilbenes and anthocyanins. | Protein accumulation and AChE inhibition. | Inhibition of the abnormal aggregation of Aβ1-42 through inhibition of β-secretase (in silico), and downregulation of AChE and MAO. | Anti-secretase and anti-AChE. | [85] |
| Indian gooseberry (Emblica officinalis) | Pulp | Gallic, ellagic, ascorbic and mucic acids, tannins, quercetin and rutin. | Neuroinflammation. | Reduced release of proinflammatory cytokines IL-6 and TNF-α from the microglia activation. Growth of neurite length. Induction of mRNA expression of neuronal markers such as MAP2. Decrease in NO production. | Anti-inflammatory. | [48] |
| North-Thai Berry (Cleistocalyx nervosum var. paniala) | Pulp | Flavonoids and resveratrol | Excitotoxicity. Apoptosis and oxidative stress. | Improvement in cell morphology and survival after glutamate-induced toxicity. Inhibition of apoptosis suppressing cleaved caspase-3 levels. Suppression of intracellular ROS levels and upregulation of mRNA/gene expression of SOD, CAT, GPX1. Stimulation of Nrf2 activity. | Anti-excitotoxic and antioxidant. | [86] |
| Grape (Vitis vinifera) | Leaf | Resveratrol, gallic acid, apigenin, catechin, quercetin. | Antioxidant potential and excitotoxicity. | Low glutamate-induced cell death. Inhibition of ROS accumulation and promotion of CAT, SOD, GST and GPx enzymes. | Antioxidant and anti-excitotoxic. | [87] |
| Orange (Citrus sinensis) | Entire fruit | Mono and sesquiterpenes. | Enzyme inhibition, inflammation and oxidation. | AChE, BChE and LOX inhibition capacity. Significant anti-cholinergic compounds expected to have fewer side effects than synthetics. | Anti-AChE/-BChE and anti-inflammatory. | [42] |
| Pomegranate (Punica granatum) | Peel | Caffeic, chlorogenic, coumaric and gallic acids, catechin, quercetin, delphinidin and cyanidin. | AChE inhibition and in silico studies. | AChE and β-secretase inhibition due to hydrogen bond formation. | Anti-AChE and anti-secretase. | [88] |
| Pistachio, fig and date | Flower buds | Chrysoeriol, epicatechin, feruloyl-malic, caffeoylquinic and coumarylquinic acids, quercetin, isorhamnetin, etc. | Endogenous antioxidant enzymes in plasma from PD humans. | Significant antioxidant enzymatic properties expressed by the increase in plasma SOD and GR activity. Positive correlations between TPC and GR, and TFC and GR. Demonstration of synergistic effect of phenolics. | Antioxidant. | [77] |
| Coffee (Coffea) | Waste | Gallic, loganic, caffeoylquinic acids, rutin, naringin, quercetin, quercitrin, catechin. | Mitochondrial dysfunction and neuroinflammation. | Increase in cell viability and intensification of mitochondrial respiration. Reduction in iNOS, COX-2 and cytokines expression maintaining NF-kB proteins from TLR4/NF-kB pathway at control levels. | Pro-mitochondrial respiration and anti-inflammatory. | [89] |
| Pear (Pyrus communis) | Pulp + Peel | Arbutin, rutin, chlorogenic acid, quercitrin and procyanidins B1 and B2. | Antioxidant potential and AChE inhibition. | Significant antioxidant activity. Inhibition of AChE. | Antioxidant and anti-AChE. | [90] |
| Olive (Olea europaea) | Entire fruit | Oleuropein. | Protein accumulation. | Inhibition of nucleation, elongation, fibrillation and oligomerization of αSn. | Anti-aggregation. | [34] |
| Wild plum (Harpephyllum caffrum) | Pulp | Nobiletin, puerarin, phellamurin, tetragalloyl glucose, quercetin glucoside, etc. | Antioxidant potential, LPO and AChE inhib. (in silico). | Increase in GSH, SOD, and CAT levels. Reduction in MDA concentration and AChE activity. | Antioxidant and anti-AChE. | [91] |
| Wild turnip (Brassica rapa) | Root | Kaempferol and isorhamnetin, phenolic and organic acids. | Mitochondrial apoptosis. | Avoidance of autophagy promoting phosphorylation of PI3K, Akt and mTOR and activating PI3K/Akt/mTOR pathway on HT-22 cells. Inhibition of ROS and restauration of mitochondrial expression. | Anti-autophagic and antioxidant. | [92] |
| Artichoke (Cynara scolymus) | Leaf, Bract and Stem | Caffeoylquinic, feruloylquinic acids, luteolin derivatives. | Antioxidant potential and AChE inhibition. | Radical scavenging and antioxidant potential. Inhibition of AChE and BChE. | Antioxidant, anti-AChE and anti-BChE. | [93] |
| Black Pigweed (Trianthema portulacastrum) | Leaf | Protocatechuic, caffeic, chlorogenic, and ferulic acids. | Antioxidant potential and AChE inhibition (in silico). | Inhibition of AChE activity, as well as high antioxidant activity. Chlorogenic acid showed most significant binding affinity towards AChE. | Antioxidant and anti-AChE. | [68] |
| Senecio biafrae (Senecio biafrae) | Leaf | Gallic, chlorogenic, caffeic acids, kaempferol, rutin, quercetin. | Antioxidant potential and AChE inhibition. | NO radical scavenging, ferric chelating capacity and antioxidant potential. Inhibition of AChE and BChE. | Antioxidant, anti-AChE and anti-BChE. | [94] |
| Sacred lotus (Nelumbo nucifera) | Seed, stalk, leaf, petal and stamen | Gallic, ferulic, p-coumaric acids, naringenin, quercetin, luteolin, kaempferol, cyanidin, etc. | Antioxidant potential, AChE inhibition and protein accumulation. | High TPC and antioxidant values. Ability to inhibit enzymes such as AChE, BChE and BACE-1, avoiding cytotoxic Aβ peptide aggregation. | Antiamyloidogenic, antioxidant and anti-AChE/-BChE. | [69] |
| Que Zui Tea (Vaccinium dunalianum) | Flower, fruit and leaf | Chlorogenic. caffeic, p-coumaroyl and feruloyl quinic acids, kaempferol, eriodyctiol, etc. | Antioxidant potential. | High intracellular ROS scavenging effect. High levels of CAT, GSH and SOD. Downregulation of MDA and apoptosis. Elevated expression of Nrf2. | Antioxidant and antiapoptotic. | [95] |
| African blackwood (Dalbergiella welwitschii) | Leaf | Proanthocyanidins and flavonoids. | Antioxidant potential and AChE inhibition. | High antioxidant activity and radical scavenging capacity. Ability to inhibit AChE and BChE activity. | Antioxidant and anti-AChE/-BChE. | [96] |
| Velvet bean (Mucuna pruriens) | Seed | Isoflavanones and alkaloids. | Antioxidant potential, AChE and neurotransmission. | Good antioxidant activity, reduction in MAO A, B and AChE activities. | Antioxidant and anti-AChE. | [97] |
| Oat (Avena sativa) | Seedling | Feruloylquinic acid, isoorientin, vitexin, avenacosides A and B, diosgenoside, etc. | LPO oxidative stress induction and protein aggregation. | Possible inhibition of LPS-induced ROS in microglia. Significant reduction in Aβ 42 and Aβ 40, reduction in APP levels. | Antioxidant and anti-amyloidogenic. | [98,99] |
| Sickle Senna (Cassia tora) | Leaf | Gentistic, ferulic, p-coumaric, vanillic and gallic acids. | Antioxidant potential, DNA damage and apoptosis. | Prevention of ROS generation, LPO, DNA damage, apoptosis and neuronal cell death. | Antioxidant and antiapoptotic. | [100] |
| Elaeagnus glabra (Eleagnus glabra) | Branch | 4-hydroxybenzoic, vanillic acids, (epi)(gallo)catechin, procyanidin B type, kaempferol, etc. | Protein aggregation and antioxidant potential. | Strong inhibitory effect on Aβ aggregation. Remarkable radical scavenging activity. | Anti-amyloidogenic and antioxidant. | [101] |
| Plant Specie | By-Product | Main Compounds | Biological Target | Mechanisms of Action | Neuroprotective Effect | Citation |
|---|---|---|---|---|---|---|
| Avocado (Persea americana) | Seed and peel | Caffeic acid, catechin, rutin, procyanidin and flavonoids. | Drosophila melanogaster survival and movement. | Increase in the number of flies capable of flying above the limit established. | Anti-AChE. | [78] |
| Peel | B-type procyanidins, flavanols monomers and chlorogenic acids. | Antioxidant potential in Drosophila melanogaster. | Increase in the life span and locomotor activity, decrease in LPO, prevention of induced locomotor impairment. | Antioxidant. | [22] | |
| Mango (Mangifera indica) | Leaf | Iriflophenones, mangiferin, quercetin, galloyls, etc. | Neuroinflammation and protein accumulation in diabetes mice. | Decrease in the inflammatory response and microglia burden, reduction in tau hyperphosphorylation in cortex and hippocampus. | Anti-inflammatory and antifibrillization. | [102] |
| Peel and pulp | Gallic, chlorogenic, vanillic acids, mangiferin, etc. | Antioxidant potential and LPO in rats (ex vivo). | Increased expression and activity of SOD and GST. Prevention in LPO upregulation. | Antioxidant. | [75] | |
| Passion Fruit (Passiflora edulis) | Pericarp | Polyphenols, triterpenoids, glycosides, carotenoids, aromatic oils, etc. | Maintenance of mitochondrial homeostasis in Caenorhabditis elegans. | Stimulation of mitophagy through activating protein ULK1, enhancement of the cholinergic neuronal resistance and glutamatergic neurons protection. | Pro-phagocytic and anti-excitotoxic. | [57] |
| Lychee (Litchi chinensis) | Seed | Saponins. | Apoptosis in rats. | Suppression of apoptosis in Aβ-induced cells through upregulation of Bcl-2 proteins and decrease in caspase-3 mRNA expression. | Anti-apoptotic. | [103] |
| Rutin, quercetin, catechin and Proanthocyanidins. | BBB dysfunction. | Improvement in spatial learning and memory function. Inhibition of caspase-1 and IL-1 expression, and inhibition inflammasome activation through AMPK/mTOR/ULK. | Pro-autophagocytic and anti-inflammatory. | [104] | ||
| Papaya (Carica papaya) | Leaf | Flavonols, tannins, alkaloids. | AChE inhibition and antioxidant potential in Wistar rats (ex vivo). | Inhibition of AChE and BChE. High radical scavenging capacity. | Anti-AChE and antioxidant. | [105] |
| Wampee (Clausena lansium) | Peel | Coumarins and others. | AChE inhibition, LPO and antioxidant enzymes evaluation in Wistar rats. | Reduced AChE activity and decreased oxidative status restoring SOD, CAT, and GPx activities. Low LPO. | Anti-AChE and antioxidant. | [106,107] |
| Sweet cherry (Prunus avium) | Pulp | Organic and phenolic acids, (epi) catechin, quercetin, luteolin and procyanidin B2. | Antioxidant potential in Caenorhabditis elegans. | High antioxidant in vivo capacity. | Antioxidant. | [83] |
| Mulberry (Morus alba) | Entire fruit | Procatechuic, chlorogenic and caffeolyquinic acids, taxifolin, rutin, quercetin, etc. | Protein accumulation and proteolytic systems dysfunctions in parkinsonian mice. | Protection against induced loss of dopaminergic neurons through reduction in α-synuclein and ubiquitin upregulation levels. | Anti-aggregation and UPS promoter. | [108,109] |
| Phenolic acids and flavonoids. | Antioxidant potential. Mitochondria and apoptosis in mice (ex vivo). | Improvement in glutathione level. Prevention of mitochondria membrane depolarization. Suppression of pro-apoptotic factors such as cytochrome c. Elevated expression of Akt peptide and Nrf2 translocation. | Antioxidant, antiapoptotic and anti-MMP. | [110] | ||
| Mulberry and ginger | Fruit andrhizomes | Flavonoids, gingerol, quercetin, cyanidin, ferulic and gallic acids, etc. | Antioxidant enzymes, AChE inhibition and neuroinflammation on Wistar rats (ex vivo). | Cognitive enhancing effect and mitigation of reduction in neuron density. Increased levels of SOD, CAT and GSH, and decreased levels of MDA and AChE activity. Suppression of IL-6 expression. | Antioxidant, anti-AChE and anti-inflammatory. | [111] |
| Cranberry (Vaccinium macrocarpon) | Entire fruit | Anthocyanins, procyanidins A and B, hydroxycinnamic acids and flavonols. | Cognitive health examination of healthy older adults. | Improvement in episodic memory performance and neural functioning maybe through increased regional perfusion in several brain areas. | Memory promoter. | [112] |
| Grape (Vitis vinifera) | Seed | Proanthocyanidins and resveratrol. | Antioxidant potential on albino rats. | Increase in levels of GSH, GPX, SOD and CAT activities. Low MDA levels. Inhibition of LPO. Downregulation of iNOS mRNA expression. | Antioxidant. | [113] |
| Proanthocyanidins and resveratrol. | Protein accumulation in a mouse model of AD. | Blockage of Aβ fibril formation interfering with (pre)protofibril formation and oligomerization through β-sheet structure. Reduction in tau aggregations and promotion of α-secretase activity. | Anti-fibrillization, anti-aggregation and anti-secretase. | [24,114] | ||
| Seed and peel | Rutin, gallic acid, (epi)catechins, quercetin, vanillin, resveratrol. | Neuroinflammation in mice. | Protective effect against neurotoxicity. Mitigation of caspases-3 and ROS production. Decrease in NF-kB p65 subunit liberation and translocation. Improvement in motor deficits preventing DA neurons. | Anti-apoptotic, antioxidant and anti-inflammatory. | [115] | |
| Peel | Gallic, caffeic, ferulic acids, quercetin, rutin, (epi)catechin, ECG, resveratrol, etc. | Protein aggregation, neuroinflammation and antioxidant potential. | Improved spatial learning and memory ability. Reduced levels of BACE-1, β cleavage of APP and inflammatory cytokines. Increased SOD and decreased MDA. | Anti-amyloidogenic, antioxidant and anti-inflammatory. | [116] | |
| Leaf | Resveratrol, gallic acid, apigenin, catechin, quercetin. | Promotion of endogenous antioxidant systems in C. elegans. | Stress resistance and improvement in survival rate. Reduced intracellular ROS levels. Significant increase in DAF-16 nuclear translocation. Promotion of sod-3 and gst-4 gene expression. | Antioxidant. | [87] | |
| Indian gamboge (Garcinia morella) | Peel | Morellin, mangostin, cambogic acid, garcinol, etc. | Neuroinflammation and mitochondria dysfunction in parkinsonian albino mice. | Prevention of the loss of nigral dopaminergic neurons. Alleviation of inflammatory marker nNOS and recovery of mitochondrial complex II inhibition. | Anti-inflammatory and mitochondrial alleviator. | [58,117] |
| Sour orange (Citrus aurantium) | Seed | Limonoids and hesperidin. | AChE inhibition and protein aggregation in Wistar rats. | Reduced AChE, tau protein and Aβ levels. | Anti-AChE and anti-aggregation. | [118] |
| Kawachi bankan (Citrus kawachiensis) | Peel | Naringin, narirutin, auraptene, flavones. | Neuroinflammation, antioxidant potential, avoidance of dopaminergic neurons and protein dysfunctions in mice. | Suppression of microglial activation and blocking morphological changes (ameboid shaped) in the activated microglia. Suppression of tau hyperphosphorylation and neuronal death. Increase in total GSH. | Anti-inflammatory, anti-hyperphosphorylation and antioxidant. | [119,120,121,122] |
| Pomegranate (Punica granatum) | Peel | Anthocyanins, ellagic acid, flavanols (catechins and gallocatechins), punicalagin. | Protein accumulation, AChE inhibition and antioxidant potential in mice. | Reduction in amyloid plaque density, increase in neurotrophin BDNF and reduction in AChE activity. Decrease in LPO and in the concentration of pro-inflammatory cytokine TNF-α. | Anti-aggregation antioxidant, anti-AChE and anti-inflammatory. | [123] |
| Fruit | Gallic acid, galloyl hexosides and ellagic acid derivatives. | Neuroinflammation. | Suppression of the gene expression of IL and inflammatory mediators (interferons and Tnfα). | Anti-inflammatory. | [124] | |
| Figs (Ficus carica) | Pulp | Proanthocyanidins. | Memory, anxiety and learning in mice. Protein accumulation. | Enhancement of memory, spatial learning and motor coordination. Reduction in anxiety-related behavior through inhibition of the fibrillization of Aβ. | Anti-fibrillization. | [125] |
| Pomegranates, figs and dates | Pulp | Ferulic, sinapic, coumaric acids, luteolin, quercetin, catechin, epicatechin. | Neuroinflammation and protein accumulation in mice. | Suppression of Aβ levels. Delays in the formation of senile plaques. Reduction in pro-inflammatory ILs and TNF-α expression. Improvement in ATP formation. | Antiamyloidogenic and anti-inflammatory. | [30,126] |
| Olive and hibiscus (Hibiscus sabdariffa) | Leaf and Flower | Elenolic acid, oleuropein and hibiscus acid. | Apoptosis in Wistar rats (in silico). | Increase in cell viability under oxidative stress conditions. Reduction in mitochondria membrane potential loss, and reversion of caspases to basal levels. Reduced apoptosis mediated by oxidative stress. | Anti-MMP and antiapoptotic. | [41] |
| Apple (Malus domestica) | Pulp + Peel | Quercetin and dihydroxybenzoic acid. | Neural precursor cells in mice. | Promotion of cellular survival and neuronal differentiation. Presence of different pro-neurogenic compounds. Increase in neural precursor cell proliferation and neurogenesis. Inducement of endogenous antioxidants. | Pro-neurogenic and antioxidant. | [127] |
| Persimmon (Diospyros kaki) | Leaf | Myricetin, kaempferol, quercetin, hyperoside, astragalin and vitexin. | Neuroinflammation, antioxidant enzymes and apoptosis. | Decreased number of activated microglia and astrocytes reducing specific markers expression. Attenuation of NF-kB nuclear translocation. Improvement in SOD, GSH and CAT activity, and in PI3K and p-Akt levels. | Anti-inflammatory, antioxidant and anti-apoptotic. | [14] |
| Walnut (Juglans regia) | Kernel | Flavonoids, phenolic acids (ellagic), etc. | LPO, endogenous enzymes and mitochondrial dysfunctions in mice. | Reduction in LPO processes. activities. Improvement in SOD, CAT, GSH and GPx activity. Enhancement of dopamine levels inhibiting MAO enzymes. Restauration of mitochondrial complex-I activity. | Antioxidant and mitochondrial alleviator. | [59] |
| Black Pigweed (Trianthema portulacastrum) | Leaf | Protocatechuic, caffeic, chlorogenic, and ferulic acids. | Antioxidant potential and AChE inhibition in albino mice (ex vivo). | Reduction in TL and TRC. Significant increase in enzymatic antioxidants as well as hippocampal ACh levels. | Antioxidant and anti-AChE. | [68] |
| Hardy rubber-tree (Eucommia ulmoides) | Flower | All types of flavonoids. Anthocyanins and chalcones. | Protein accumulation and apoptosis. AChE inhibition (ex vivo). | Increase exercise capacity. Low number of Aβ plaques. Inhibition of AChE. Reduction in apoptotic cells. Regulate the expression of autophagy-related genes (ulk). | Anti-aAnti-autophagocytic. | [10] |
| Leaf | Chlorogenic acid, quercetin, rutin, isoquercitrin, astragalin, etc. | Apoptosis in zebrafishes. | Inhibition of vasculature loss. Reduction in apoptotic cells. Relief of locomotor impairments. Upregulation of transcript levels of genes such as ulk2 and ulk1b. | Pro-autophagocytic. | [128] | |
| Velvet bean (Mucuna pruriens) | Seed | Isoflavanones and alkaloids. | Antioxidant potential and apoptosis in albino mice. | Increase in neurotransmitter levels. Promotion of cell survival by increasing Akt activity and repairing OTA damage, mediating in neuronal apoptosis through PI3K/Akt signaling pathway. | Pro-synaptic and antiapoptotic. | [97] |
| Oat (Avena sativa) | Seedling | Feruloylquinic acid, isoorientin, vitexin, avenacosides A and B, diosgenoside, etc. | LPO, protein aggregation and neuroinflammation in mice (ex vivo). | Possible inhibition of LPS-induced ROS in microglia. Drastic reduction in numbers and size of Aβ plaques, and significant drop in BACE1 expression. Decrease in neuroinflammatory cells. Impact on MAPK signaling. | Anti-amyloidogenic, antioxidant and anti-inflammatory. | [98,99] |
| Caesalpinia crista (Caesalpinia crista) | Leaf | Gallic, coumaric and ferulic acids, rutin and genistin. | AChE inhibition, neuroinflammation, endogenous antioxidant system and LPO in rats. | AChE activity inhibition, interference at mRNA levels for the downregulation of pro-inflammatory cytokines and increase in NTF. Restoration of CAT, GSH and GST levels. Decrease in MDA levels. | Anti-AChE, anti-inflammatory and antioxidant. | [129] |
| Saffron (Crocus sativus) | Stigma | Carotenoids (crocin) and others. | Neuroinflammation and apoptosis in albino rats. | Decrease in caspase-3, COX-2 and GFAP expression, and MDA, TNF-α and IL6 levels. | Anti-apoptotic, anti-inflammatory and antioxidant. | [130] |
| Ginger (Zingiber officinale) | Rhizome | 6-gingerol (phenolic). | Mitochondrial biogenesis in mice. | Mitochondrial biogenesis promotion, increase in mitochondrial mass, mtDNA copy number and ATP production. Regulation of AMPK-PGC1α pathway. | Mitochondrial alleviator. | [131] |
| Guiera senegalensis (Guiera senegalensis) | Leaf | Quercetin, glycosides, catechin, apigenin, epigallocatechin, caffeic, chlorogenic, cinnamic and ellagic acids. | Endogenous antioxidant system, LPO and AChE inhibition in Danio rerio. | Enhancement of antioxidant defense by preventing decrease in SOD, CAT, GPX and GSH, and suppressing the increase in MDA and protein peroxidation. AChE inhibition. | Antioxidant and anti-AChE. | [67] |
| Big-leaf mahogany (Swietenia macrophylla) | Seed | Alkaloids, terpenoids, tannins and flavonoids. | Downregulation of enzymes and cytokines in albino rats. Protein deposition. | Diminish inflammatory cascades reducing the neuronal effect of TNF-α. Significant improvement in GSH, SOD, CAT, MDA, NO and AChE levels. Avoidance of beta-amyloid depositions. | Anti-inflammatory, anti-AChE and anti-amyloidogenic. | [21] |
| Ming aralia (Polyscias fruticose) | Leaf | Proanthocyanidins, saponins, flavonoids, etc. | Parkinson on dUCH-knockdown D. melanogaster. | Improvements in flies’ mobility. Decrease in the degeneration of dopaminergic neurons. | Anti-PD. | [8] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rojas-García, A.; Fernández-Ochoa, Á.; Cádiz-Gurrea, M.d.l.L.; Arráez-Román, D.; Segura-Carretero, A. Neuroprotective Effects of Agri-Food By-Products Rich in Phenolic Compounds. Nutrients 2023, 15, 449. https://doi.org/10.3390/nu15020449
Rojas-García A, Fernández-Ochoa Á, Cádiz-Gurrea MdlL, Arráez-Román D, Segura-Carretero A. Neuroprotective Effects of Agri-Food By-Products Rich in Phenolic Compounds. Nutrients. 2023; 15(2):449. https://doi.org/10.3390/nu15020449
Chicago/Turabian StyleRojas-García, Alejandro, Álvaro Fernández-Ochoa, María de la Luz Cádiz-Gurrea, David Arráez-Román, and Antonio Segura-Carretero. 2023. "Neuroprotective Effects of Agri-Food By-Products Rich in Phenolic Compounds" Nutrients 15, no. 2: 449. https://doi.org/10.3390/nu15020449
APA StyleRojas-García, A., Fernández-Ochoa, Á., Cádiz-Gurrea, M. d. l. L., Arráez-Román, D., & Segura-Carretero, A. (2023). Neuroprotective Effects of Agri-Food By-Products Rich in Phenolic Compounds. Nutrients, 15(2), 449. https://doi.org/10.3390/nu15020449