
Fermentation of Abelmoschus manihot Extract with Halophilic Bacillus licheniformis CP6 Results in Enhanced Anti-Inflammatory Activities
 , and
, and
Abstract
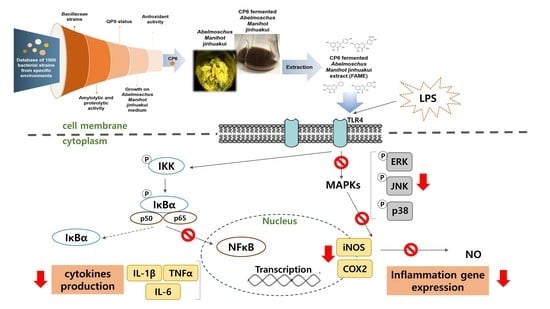
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Microbial Fermentation of Abelmoschus manihot
2.3. Antioxidant Activity Measurement
2.4. Preparation of A. manihot Fermentation Extract
2.5. Measurement of Phenolic and Flavonoid Contents
2.6. Cell Culture
2.7. Cell Viability
2.8. Flow Cytometric Analysis of NO Production
2.9. IL-1β, IL-6, and TNF-α Production
2.10. Western Blotting
2.11. Statistical Analysis
3. Results
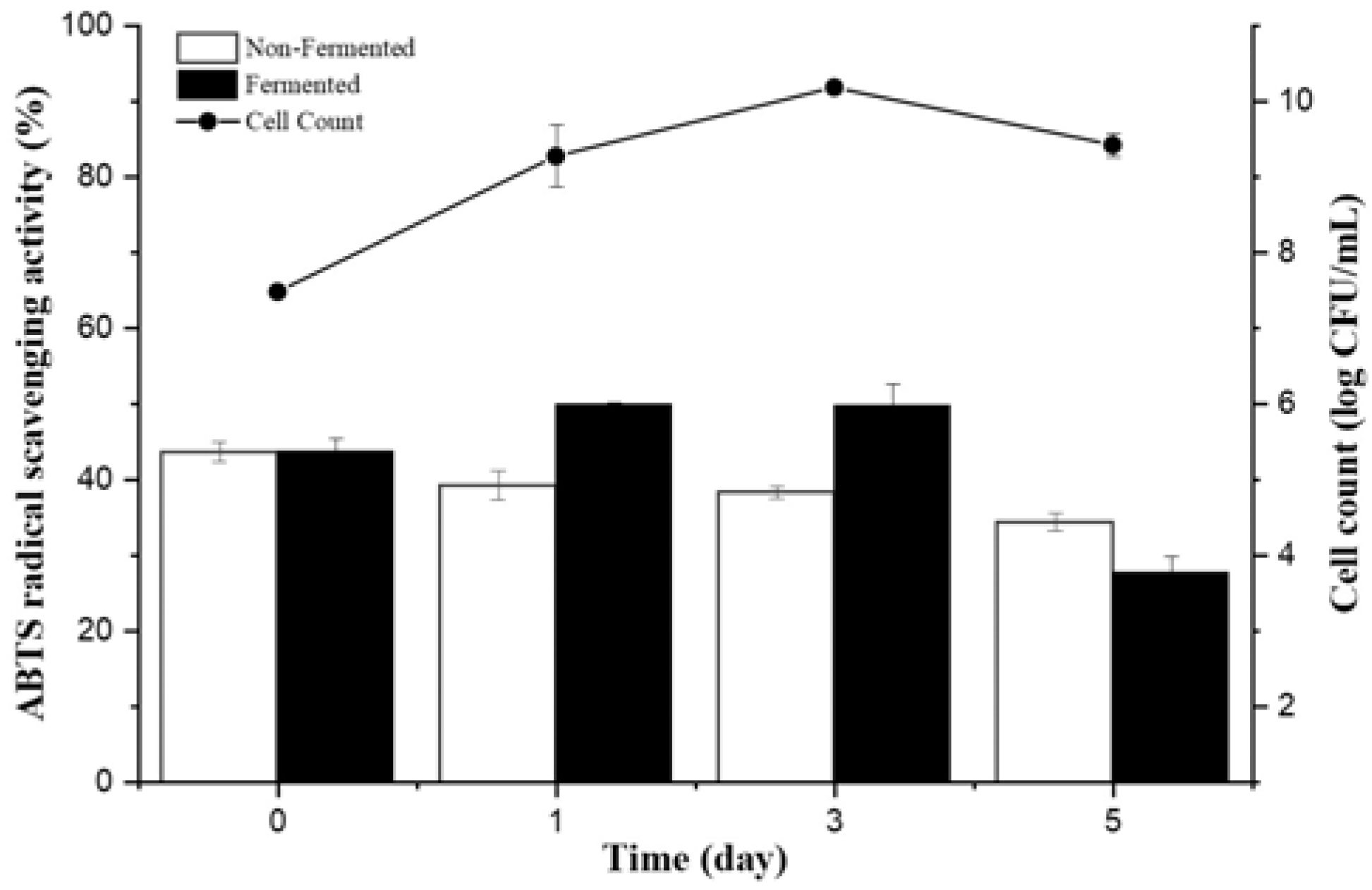
3.1. Screening Bacterial Strains for A. manihot Fermentation
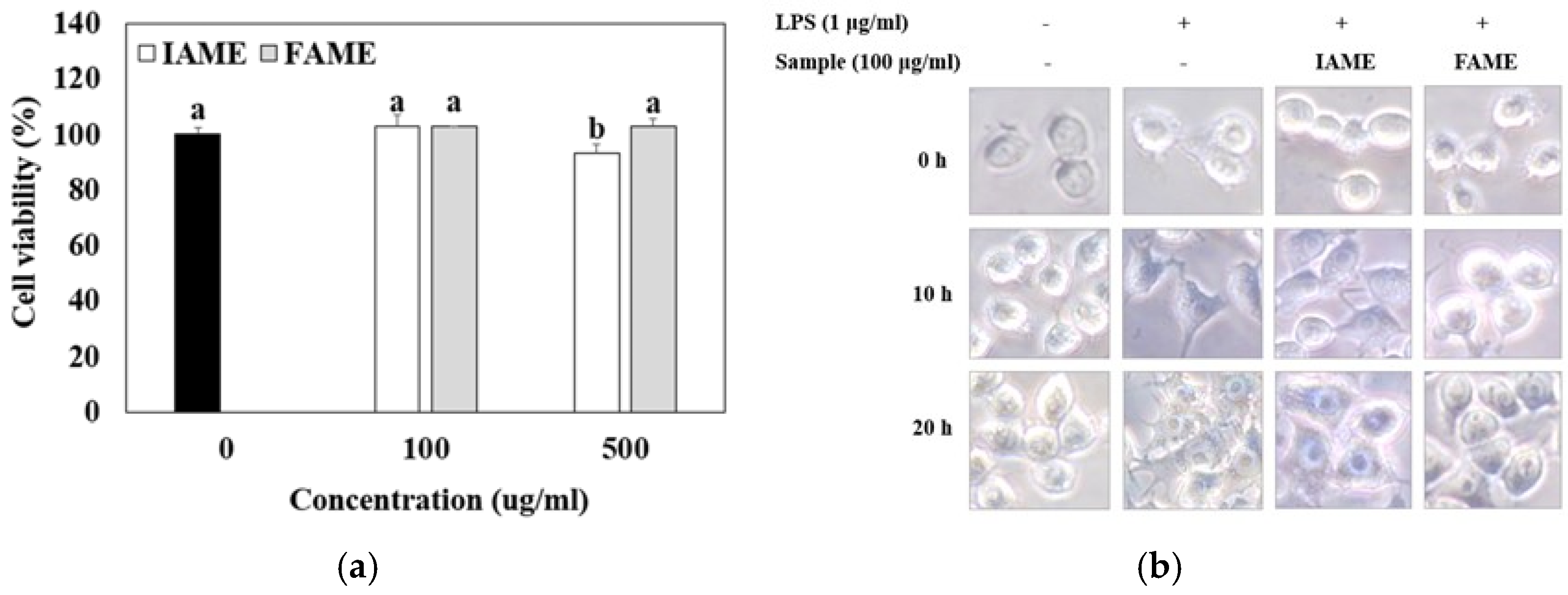
3.2. Effects of FAME on Cytotoxicity and Morphological Changes in Raw264.7 Cells
3.3. Effects of FAME on the Inhibition of NO Production in LPS-Stimulated Raw264.7 Cells
3.4. Effects of FAME on the Production of Inflammatory Cytokines in LPS-Stimulated Raw264.7 Cells
3.5. Effects of FAME on iNOS and COX-2 Expression in LPS-Stimulated Raw264.7 Cells
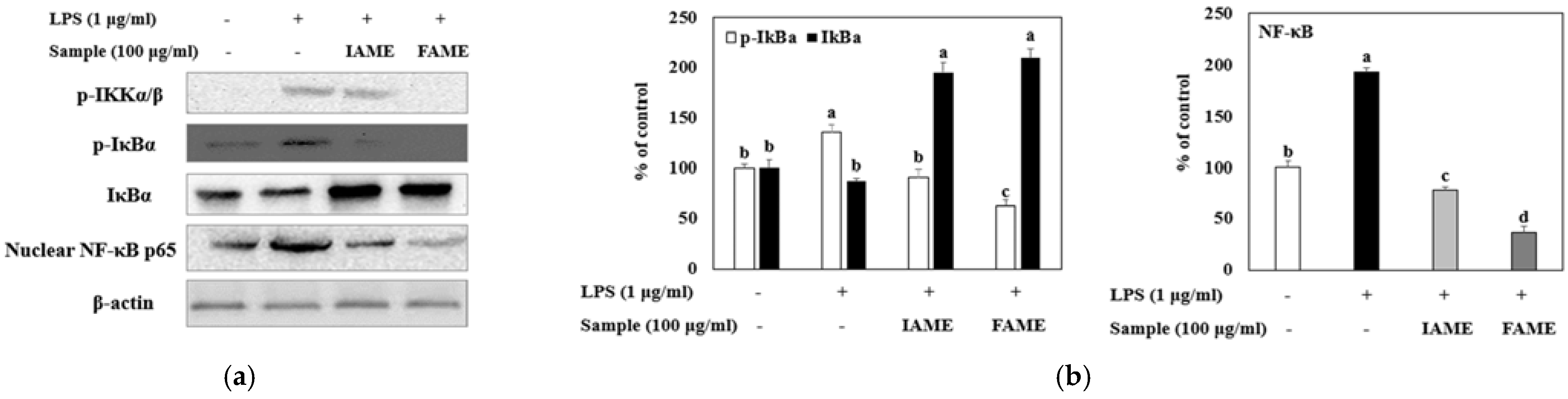
3.6. Effects of FAME on the NF-κB Signaling Pathway in LPS-Stimulated Raw264.7 Cells
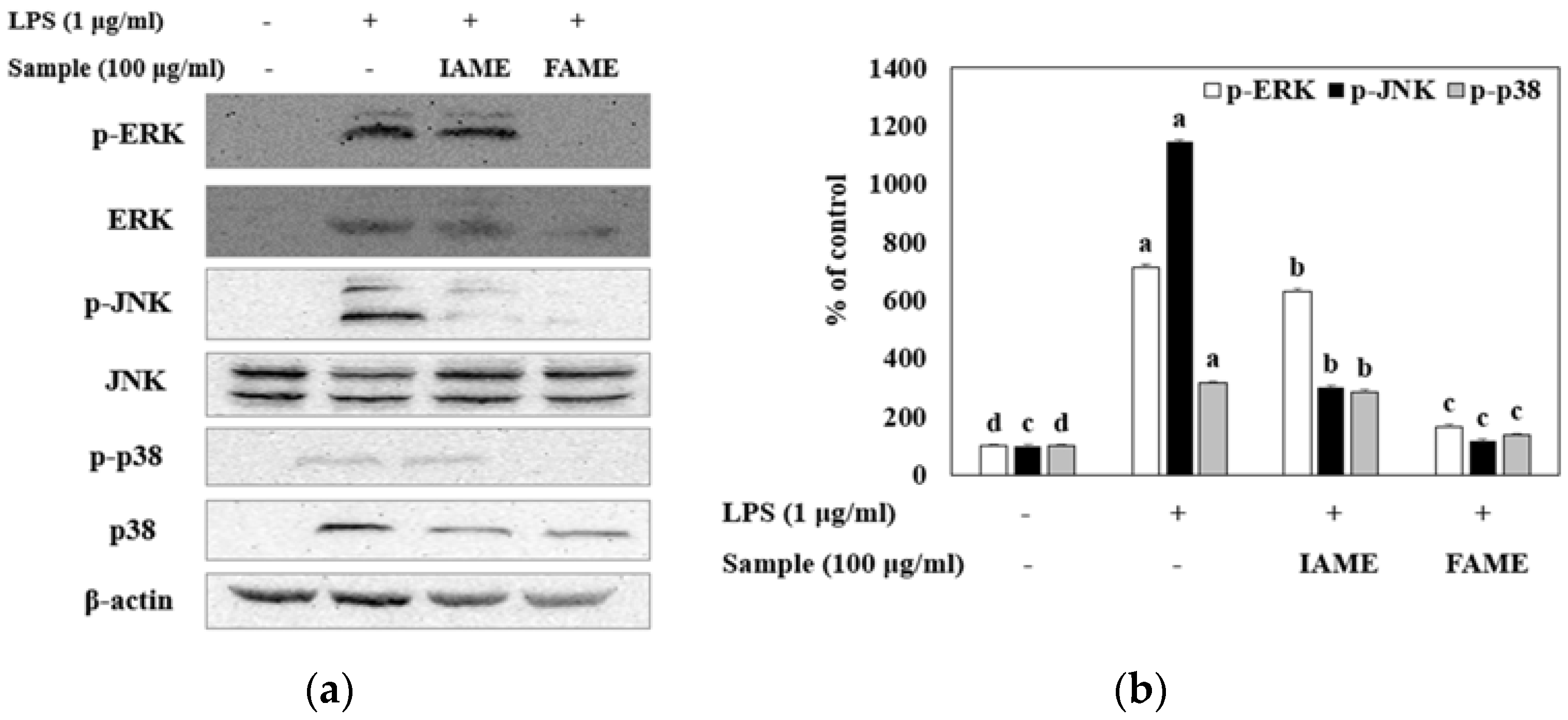
3.7. Effects of FAME on the MAPK Signaling Pathway in LPS-Stimulated Raw264.7 Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, D.Y.; Xue, M.Y.; Geng, Z.R.; Chen, P.Y. The suppressive effects of Bursopentine (BP5) on oxidative stress and NF-κB activation in lipopolysaccharide-activated murine peritoneal macrophages. Cell. Physiol. Biochem. 2012, 29, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, J.; Biswas, S.; Datta, A.G. Mode of action of endotoxin: Role of free radicals and antioxidants. Curr. Med. Chem. 2004, 11, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.O. Regulators of macrophage activation. Eur. J. Immunol. 2011, 41, 1531–1534. [Google Scholar] [CrossRef] [PubMed]
- Somensi, N.; Rabelo, T.K.; Guimarães, A.G.; Quintans-Junior, L.J.; Araújo, A.A.S.; Moreira, J.C.F.; Gelain, D.P. Carvacrol suppresses LPS-induced pro-inflammatory activation in RAW 264.7 macrophages through ERK1/2 and NF-κB pathway. Int. Immunopharmacol. 2019, 75, 105743. [Google Scholar] [CrossRef]
- Boscá, L.; Zeini, M.; Través, P.G.; Hortelano, S. Nitric oxide and cell viability in inflammatory cells: A role for NO in macrophage function and fate. Toxicology 2005, 208, 249–258. [Google Scholar] [CrossRef]
- Karin, M.; Ben-Neriah, Y. Phosphorylation meets ubiquitination: The control of NF- κB activity. Annu. Rev. Immunol. 2000, 18, 621–663. [Google Scholar] [CrossRef] [PubMed]
- Lappas, M.; Permezel, M.; Georgiou, H.M.; Rice, G.E. Nuclear factor kappa B regulation of proinflammatory cytokines in human gestational tissues in vitro. Biol. Reprod. 2002, 67, 668–673. [Google Scholar] [CrossRef]
- Baeuerle, P.A.; Baltimore, D. NF-kappa B: Ten years after. Cell 1996, 87, 13–20. [Google Scholar] [CrossRef]
- Haddad, J.J. Mitogen-activated protein kinases and the evolution of Alzheimer’s: A revolutionary neurogenetic axis for therapeutic intervention? Prog. Neurobiol. 2004, 73, 359–377. [Google Scholar] [CrossRef]
- Lambert, W.S.; Carlson, B.J.; Formichella, C.R.; Sappington, R.M.; Ahlem, C.; Calkins, D.J. Oral delivery of a synthetic sterol reduces axonopathy and inflammation in a rodent model of glaucoma. Front. Neurosci. 2017, 11, 45. [Google Scholar] [CrossRef]
- Kim, Y.; Lee, S.; Ryu, J.H.; Yoon, K.D.; Shin, S.S. Effect of Aurea Helianthus stem extract on anti-melanogenesis. Biosci. Biothechnol. Biochem. 2018, 82, 1871–1879. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Park, C.G.; Varghese, R.; Lee, J.Y.; Kim, Y.O.; Sung, G.H. In-vitro antioxidative, antiinflammatory properties of Aurea helianthus leaf extract a Korean traditional medicinal plant. Saudi J. Biol. Sci. 2017, 24, 1943–1947. [Google Scholar] [CrossRef] [PubMed]
- Koyama, M.; Ogasawara, Y.; Endou, K.; Akano, H.; Nakajima, T.; Aoyama, T.; Nakamura, K. Fermentation-induced changes in the concentrations of organic acids, amino acids, sugars, and minerals and superoxide dismutase-like activity in tomato vinegar. Int. J. Food Prop. 2017, 20, 888–898. [Google Scholar] [CrossRef]
- Zhu, D.; Adebisi, W.A.; Ahmad, F.; Setupathy, S.; Danso, B.; Sun, J. Recent Development of extremophilic bacteria and their application in biorefinery. Front. Bioeng. Biotechnol. 2020, 8, 483. [Google Scholar] [CrossRef] [PubMed]
- EFSA BIOHAZ Panel. Uptated list of QPS-recommended biological agents for safety risk assessments carried out by EFSA. EFSA J. 2022, 20, 7408. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Srivastava, M.; Tiwari, R.; Sharma, N. Assessment of phenol and flavonoid content in the plant materials. J. New Biol. Rep. 2013, 2, 163–166. [Google Scholar]
- Hakiman, M.; Syed, M.A.; Syahidal, A.; Maziah, M. Total antioxidant, polyphenol, phenolic acid, and flavonoid content in Ficus deltoidea varieties. J. Med. Plants Res. 2012, 6, 4776–4784. [Google Scholar] [CrossRef]
- Hill, P.A.; Tumber, A.; Meikle, M.C. Multiple extracellular signals promote osteoblast survival and apoptosis. Endocrinology 1997, 138, 3849–3858. [Google Scholar] [CrossRef]
- Saxena, R.K.; Vallyathan, V.; Lewis, D.M. Evidence for lipopolysaccharide-induced differentiation of RAW264.7 murine macrophage cell line into dendritic like cells. J. Biosci. 2003, 28, 129–134. [Google Scholar] [CrossRef]
- Bogdan, C. Nitric oxide and the immune response. Nat. Immunol. 2001, 2, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Li, X.; Zhu, J.; Wu, J.; Geng, S.; Zhong, C. Magnesium isoglycyrrhizinate suppresses LPS-induced inflammation and oxidative stress through inhibiting NF-κB and MAPK pathways in RAW264.7 cells. Bioorg. Med. Chem. 2019, 27, 516–524. [Google Scholar] [CrossRef]
- Lee, A.K.; Sung, S.H.; Kim, Y.C.; Kim, S.G. Inhibition of lipopolysaccharide-inducible nitric oxide synthase, TNF-alpha and COX-2 expression by sauchinone effects on I-kappaBalpha phosphorylation, C/EBP and AP-1 activation. Br. J. Pharmacol. 2003, 139, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Bondeson, J. The mechanisms of action of disease-modifying antirheumatic drugs: A review with emphasis on macrophage signal transduction and the induction of proinflammatory cytokines. Gen. Pharmacol. 1997, 29, 127–150. [Google Scholar] [CrossRef] [PubMed]
- Taveira, I.C.; Nogueira, K.M.V.; Oliveira, D.L.G.; Silva, R.D.N. Fermentation: Humanity’s oldest biotechnological tool. Front. Young Minds 2021, 9, 568656. [Google Scholar] [CrossRef]
- Arora, N.K.; Ranosyan, H. Extremophiles: Applications and roles in environmental sustainability. Environ. Sustain. 2019, 2, 217–218. [Google Scholar] [CrossRef]
- Korhonen, R.; Lahti, A.; Kankaanranta, H.; Moilanen, E. Nitric oxide production and signaling in inflammation. Curr. Drug Targets Inflamm. Allergy 2005, 4, 471–479. [Google Scholar] [CrossRef]
- McAdam, E.; Haboubi, H.N.; Forrester, G.; Eltahir, Z.; Spencer-Harty, S.; Davies, C.; Griffiths, A.P.; Baxter, J.N.; Jenkins, G.J.S. Inducible nitric oxide synthase (iNOS) and nitric oxide (NO) are important mediators of reflux-induced cell signalling in esophageal cells. Carcinogenesis 2010, 33, 2035–2043. [Google Scholar] [CrossRef]
- Tsatsanis, C.; Androulidaki, A.; Venihaki, M.; Margioris, A.N. Signalling networks regulating cyclooxygenase-2. Int. J. Biochem. Cell Biol. 2006, 38, 1654–1661. [Google Scholar] [CrossRef]
- Kim, H.K.; Cheon, B.S.; Kim, Y.H.; Kim, S.Y.; Kim, H.P. Effects of naturally occurring flavonoids on nitric oxide production in the macrophage cell line RAW 264.7 and their structure–activity relationships. Biochem. Pharmacol. 1999, 58, 759–765. [Google Scholar] [CrossRef]
- Nam, N.H. Naturally occurring NF-κB inhibitors. Mini Rev. Med. Chem. 2006, 6, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Overview of the IL-1 family in innate inflammation and acquired immunity. Immunol. Rev. 2018, 281, 8–27. [Google Scholar] [CrossRef] [PubMed]
- Abbasifard, M.; Khorramdelazad, H. The bio-mission of interleukin-6 in the pathogenesis of COVID-19: A brief look at potential therapeutic tactics. Life Sci. 2020, 257, 118097. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.L.; Lamon-Fava, S.; Matthan, N.R.; Wu, D.; Lichtenstein, A.H. Docosahexaenoic acid differentially affects TNFα and IL-6 expression in LPS-stimulated RAW 264.7 murine macrophages. Prostaglandins Leukot. Essent. Fat. Acids 2015, 97, 27–34. [Google Scholar] [CrossRef]
- Ramiro, E.; Franch, A.; Castellote, C.; Pérez-Cano, F.; Permanyer, J.; Izquierdo-Pulido, M.; Castell, M. Flavonoids from Theobroma cacao down-regulate inflammatory mediators. J. Agric. Food Chem. 2005, 53, 8506–8511. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, A. Signal transduction of inflammatory cytokines and tumor development. Cancer Sci. 2006, 97, 439–447. [Google Scholar] [CrossRef]
- Nepali, S.; Ki, H.H.; Lee, J.H.; Lee, H.Y.; Kim, D.K.; Lee, Y.M. Wheatgrass-derived polysaccharide has antiinflammatory, anti-oxidative and anti-apoptotic effects on LPS-induced hepatic injury in mice. Phytother. Res. 2017, 31, 1107–1116. [Google Scholar] [CrossRef]
- Gao, H.; Cui, Y.; Kang, N.; Liu, X.; Liu, Y.; Zou, Y.; Zhang, Z.; Li, X.; Yang, S.; Li, J.; et al. Isoacteoside, a dihydroxyphenylethyl glycoside, exhibits anti-inflammatory effects through blocking toll-like receptor 4 dimerization. Br. J. Pharmacol. 2017, 174, 2880–2896. [Google Scholar] [CrossRef]
- Li, L.; Zhang, X.H.; Liu, G.R.; Liu, C.; Dong, Y.M. Isoquercitrin suppresses the expression of histamine and pro-inflammatory cytokines by inhibiting the activation of MAP Kinases and NF-κB in human KU812 cells. Chin. J. Nat. Med. 2016, 14, 407–412. [Google Scholar] [CrossRef]
- Arthur, J.S.C.; Ley, S.C. Mitogen-activated protein kinases in innate immunity. Nat. Rev. Immunol. 2013, 13, 679–692. [Google Scholar] [CrossRef]
- Reibman, J.; Talbot, A.T.; Hsu, Y.; Ou, G.; Jover, J.; Nilsen, D.; Pillinger, M.H. Regulation of expression of granulocyte-macrophage colony-stimulating factor in human bronchial epithelial cells: Roles of protein kinase C and mitogen-activated protein kinases. J. Immunol. 2000, 165, 1618–1625. [Google Scholar] [CrossRef] [PubMed]
- Rajapakse, N.; Kim, M.M.; Mendis, E.; Kim, S.K. Inhibition of inducible nitric oxide synthase and cyclooxygenase-2 in lipopolysaccharide-stimulated RAW264.7 cells by carboxybutyrylated glucosamine takes place via down-regulation of mitogen-activated protein kinase-mediated nuclear factor-kappaB signaling. Immunology 2008, 123, 348–357. [Google Scholar] [PubMed]
- Kaminska, B. MAPK signaling pathways as molecular targets for anti-inflammatory therapy from molecular mechanisms to therapeutic benefits. Biochim. Biophys. Acta 2005, 1754, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Yong, C.C.; Yoon, Y.; Yoo, H.S.; Oh, S. Effect of Lactobacillus fermentation on the anti-inflammatory potential of turmeric. J. Microbiol. Biotechnol. 2019, 29, 1561–1569. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef]
- Adebo, O.A.; Gabriela Medina-Meza, I. Impact of fermentation on the phenolic compounds and antioxidant activity of whole cereal grains: A mini review. Molecules 2020, 25, 927. [Google Scholar] [CrossRef]
- Yang, J.; Wu, X.; Chen, H.; Sun-waterhouse, D.; Zhong, H.; Cui, C. A value-added approach to improve the nutritional quality of soybean meal byproduct: Enhancing its antioxidant activity through fermentation by Bacillus amyloliquefaciens SWJS22. Food Chem. 2019, 272, 396–403. [Google Scholar] [CrossRef]
- Tuly, J.A.; Zabed, H.M.; Nizami, A.-S.; Mehedi Hassan, M.; Roknul Azam, S.M.; Kumar Awasthi, M.; Janet, Q.; Chen, G.; Dzidzorgbe Kwaku Akpabli-Tsigbe, N.; Ma, H. Bioconversion of agro-food industrial wastes into value-added peptides by a Bacillus sp. mutant through solid-state fermentation. Bioresour. Technol. 2022, 346, 126513. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Isolate | Closest Match | Growth of 1% AM Medium (log CFU/mL) after 5 Days | Antioxidant Activity (ABTS) % | Total Soluble Protein Concentration (BCA) µg/mL | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0D | 1D | 3D | 5D | 0D | 1D | 3D | 5D | ||||
| 1% AM Medium | 1% AM Medium | ||||||||||
| 1 | CP-6 | Bacillus licheniformis | 8.119 | 43.8 ± 1.6 | 50.0 ± 0.3 | 50.0 ± 2.7 | 27.8 ± 2.1 | 412.3 ± 15.7 | 243.58 ± 12.78 | 270.7 ± 6.3 | 149.3 ± 0.8 |
| 2 | CP-7 | Bacillus subtilis | 7.949 | 47.1 ± 0.5 | 37.5 ± 2.7 | 34.0 ± 1.5 | 299.1 ± 5.3 | 205.6 ± 3.2 | 215.2 ± 0.4 | ||
| 3 | CP-35 | Bacillus subtilis | 9.006 | 51.8 ± 3.1 | 44.9 ± 2.6 | 35.2 ± 2.4 | 344.0 ± 39.7 | 214.2 ± 4.4 | 195.0 ± 4.6 | ||
| 4 | CP-37 | Bacillus subtilis | 8.130 | 30.2 ± 5.4 | 43.4 ± 2.2 | 30.9 ± 0.6 | 230.5 ± 8.1 | 268.4 ± 19.2 | 215.2 ± 2.2 | ||
| 5 | CP-39 | Bacillus firmus | 7.845 | 43.7 ± 2.5 | 34.0 ± 0.5 | 36.0 ± 2.0 | 313.1 ± 11.5 | 216.8 ± 1.6 | 236.0 ± 3.8 | ||
| 6 | CP-42 | Bacillus velezensis | 6.667 | 50.1 ± 4.9 | 42.3 ± 1.2 | 39.7 ± 4.0 | 236.8 ± 22.9 | 270.1 ± 2.1 | 248.5 ± 20.6 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, M.H.; Yeom, Y.J.; Ganbat, D.; Kim, M.K.; Kim, S.-B.; Lee, Y.-J.; Lee, S.-J. Fermentation of Abelmoschus manihot Extract with Halophilic Bacillus licheniformis CP6 Results in Enhanced Anti-Inflammatory Activities. Nutrients 2023, 15, 309. https://doi.org/10.3390/nu15020309
Park MH, Yeom YJ, Ganbat D, Kim MK, Kim S-B, Lee Y-J, Lee S-J. Fermentation of Abelmoschus manihot Extract with Halophilic Bacillus licheniformis CP6 Results in Enhanced Anti-Inflammatory Activities. Nutrients. 2023; 15(2):309. https://doi.org/10.3390/nu15020309
Chicago/Turabian StylePark, Mi Hwa, Yu Jeong Yeom, Dariimaa Ganbat, Min Kyeong Kim, Seong-Bo Kim, Yong-Jik Lee, and Sang-Jae Lee. 2023. "Fermentation of Abelmoschus manihot Extract with Halophilic Bacillus licheniformis CP6 Results in Enhanced Anti-Inflammatory Activities" Nutrients 15, no. 2: 309. https://doi.org/10.3390/nu15020309
APA StylePark, M. H., Yeom, Y. J., Ganbat, D., Kim, M. K., Kim, S.-B., Lee, Y.-J., & Lee, S.-J. (2023). Fermentation of Abelmoschus manihot Extract with Halophilic Bacillus licheniformis CP6 Results in Enhanced Anti-Inflammatory Activities. Nutrients, 15(2), 309. https://doi.org/10.3390/nu15020309