
Gallation and B-Ring Dihydroxylation Increase Green Tea Catechin Residence Time in Plasma by Differentially Affecting Tissue-Specific Trafficking: Compartmental Model of Catechin Kinetics in Healthy Adults


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
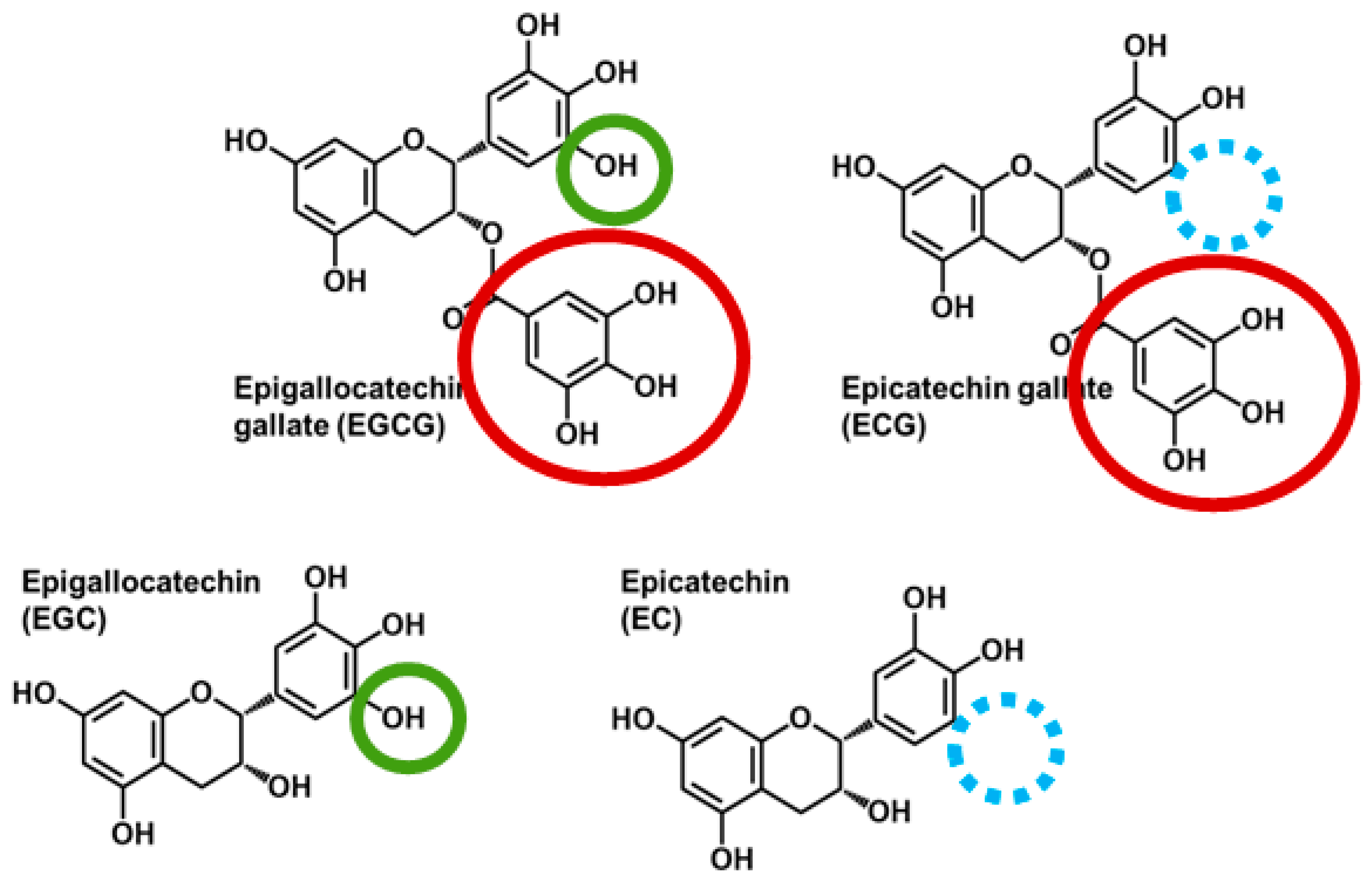
:1. Introduction
2. Materials and Methods
2.1. Participants
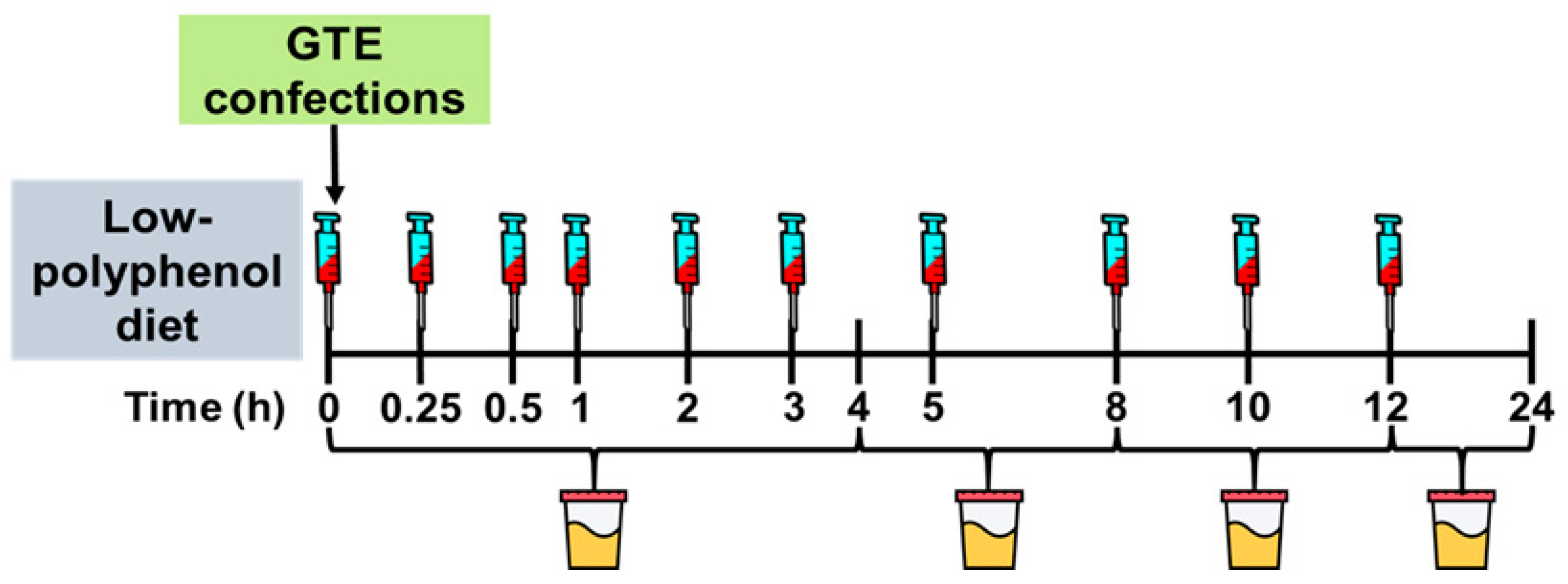
2.2. Study Design
2.3. GTE Confections
2.4. Biospecimen Preservation and Catechin Analysis in Plasma and Urine
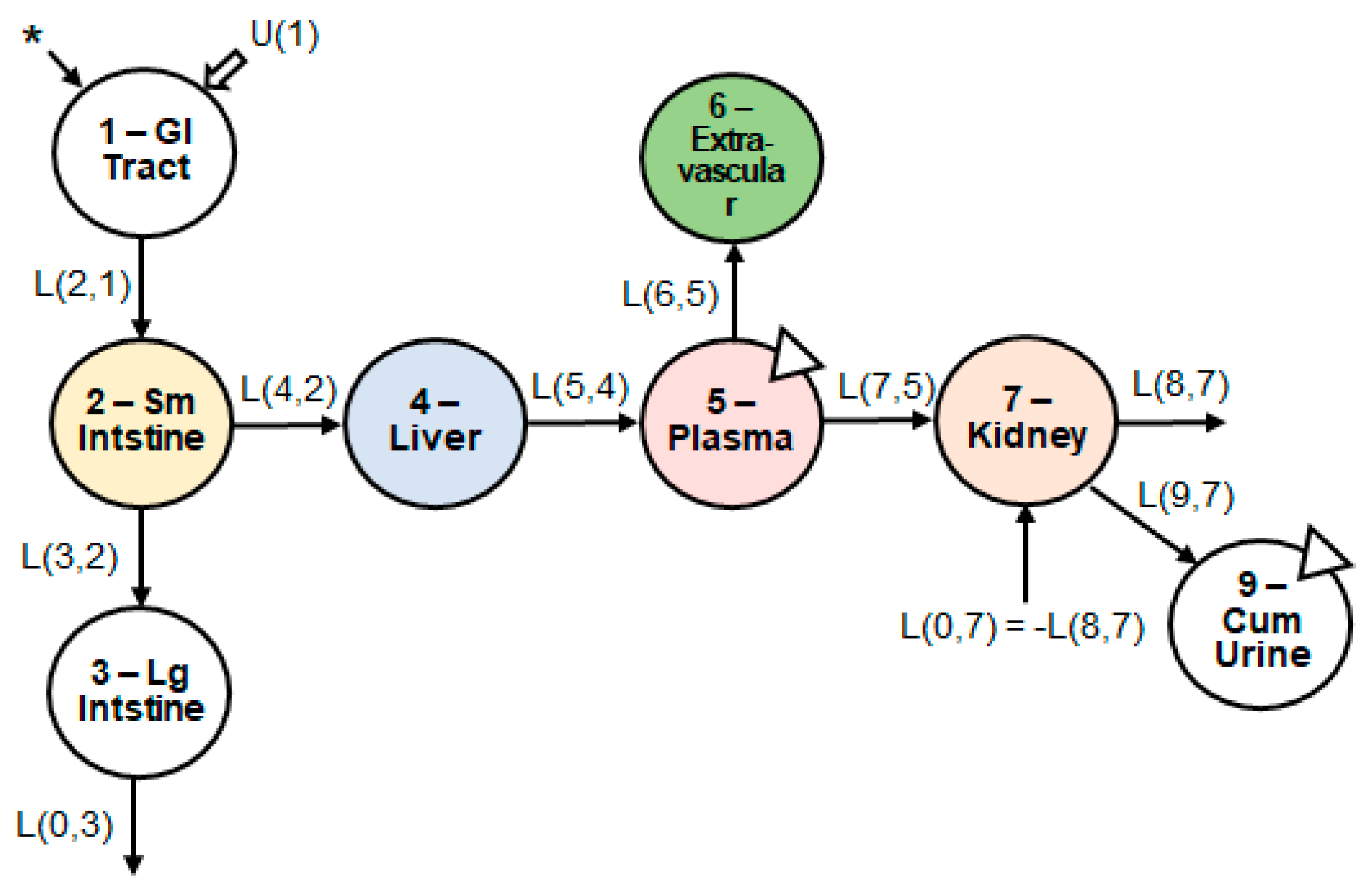
2.5. Compartmental Model Development and Kinetic Analysis
2.6. Statistical Analysis
3. Results
3.1. Model Structure
3.2. Response Profiles of Catechins in Plasma and Urine
3.3. Fractional Catabolic Rate and Residence Time
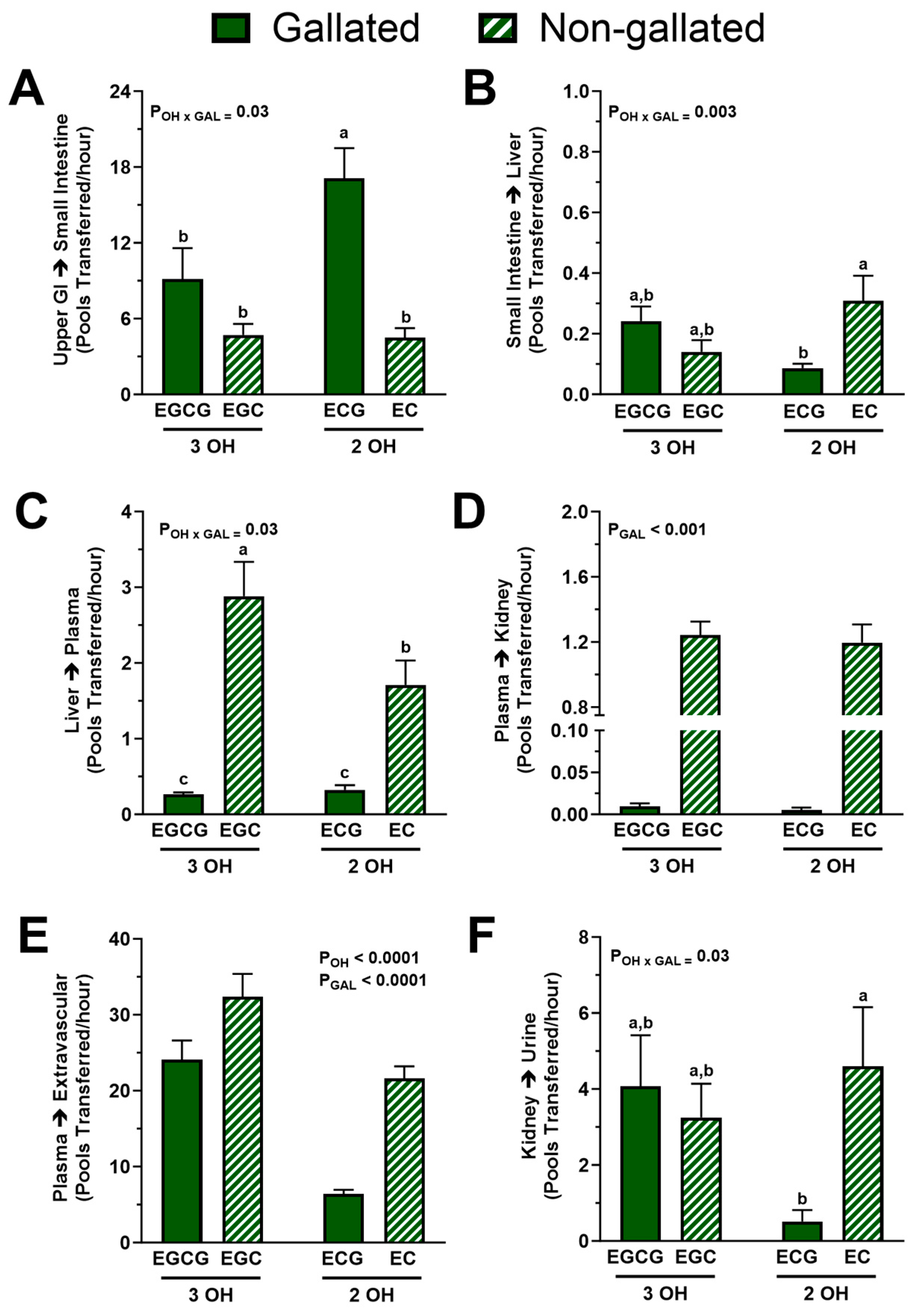
3.4. Tissue-Level Kinetics
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pang, J.; Zhang, Z.; Zheng, T.Z.; Bassig, B.A.; Mao, C.; Liu, X.; Zhu, Y.; Shi, K.; Ge, J.; Yang, Y.J.; et al. Green Tea Consumption and Risk of Cardiovascular and Ischemic Related Diseases: A Meta-Analysis. Int. J. Cardiol. 2016, 202, 967–974. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, F.; Li, J.; Yang, X.; Chen, J.; Cao, J.; Wu, X.; Lu, X.; Huang, J.; Li, Y.; et al. Tea Consumption and the Risk of Atherosclerotic Cardiovascular Disease and All-Cause Mortality: The China-Par Project. Eur. J. Prev. Cardiol. 2020, 27, 1956–1963. [Google Scholar] [CrossRef]
- Filippini, T.; Malavolti, M.; Borrelli, F.; Izzo, A.A.; Fairweather-Tait, S.J.; Horneber, M.; Vinceti, M. Green Tea (Camellia Sinensis) for the Prevention of Cancer. Cochrane Database Syst. Rev. 2020, 3, CD005004. [Google Scholar] [CrossRef] [PubMed]
- Casanova, E.; Salvado, J.; Crescenti, A.; Gibert-Ramos, A. Epigallocatechin Gallate Modulates Muscle Homeostasis in Type 2 Diabetes and Obesity by Targeting Energetic and Redox Pathways: A Narrative Review. Int. J. Mol. Sci. 2019, 20, 532. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Wang, Y.; Xie, Z.; Zhou, Y.; Zhang, Y.; Wan, X. The Anti-Obesity Effects of Green Tea in Human Intervention and Basic Molecular Studies. Eur. J. Clin. Nutr. 2014, 68, 1075–1087. [Google Scholar] [CrossRef]
- Yang, W.S.; Wang, W.Y.; Fan, W.Y.; Deng, Q.; Wang, X. Tea Consumption and Risk of Type 2 Diabetes: A Dose-Response Meta-Analysis of Cohort Studies. Br. J. Nutr. 2014, 111, 1329–1339. [Google Scholar] [CrossRef]
- Alves Ferreira, M.; Oliveira Gomes, A.P.; Guimaraes de Moraes, A.P.; Ferreira Stringhini, M.L.; Mota, J.F.; Siqueira Guedes Coelho, A.; Borges Botelho, P. Green Tea Extract Outperforms Metformin in Lipid Profile and Glycaemic Control in Overweight Women: A Double-Blind, Placebo-Controlled, Randomized Trial. Clin. Nutr. ESPEN 2017, 22, 1–6. [Google Scholar] [CrossRef]
- Asbaghi, O.; Fouladvand, F.; Gonzalez, M.J.; Aghamohammadi, V.; Choghakhori, R.; Abbasnezhad, A. Effect of Green Tea on Anthropometric Indices and Body Composition in Patients with Type 2 Diabetes Mellitus: A Systematic Review and Meta-Analysis. Complement. Med. Res. 2021, 28, 244–251. [Google Scholar] [CrossRef]
- Asbaghi, O.; Fouladvand, F.; Gonzalez, M.J.; Ashtary-Larky, D.; Choghakhori, R.; Abbasnezhad, A. Effect of Green Tea on Glycemic Control in Patients with Type 2 Diabetes Mellitus: A Systematic Review and Meta-Analysis. Diabetes Metab. Syndr. 2021, 15, 23–31. [Google Scholar] [CrossRef]
- Dey, P.; Olmstead, B.D.; Sasaki, G.Y.; Vodovotz, Y.; Yu, Z.; Bruno, R.S. Epigallocatechin Gallate but Not Catechin Prevents Nonalcoholic Steatohepatitis in Mice Similar to Green Tea Extract While Differentially Affecting the Gut Microbiota. J. Nutr. Biochem. 2020, 84, 108455. [Google Scholar] [CrossRef]
- Hodges, J.K.; Sasaki, G.Y.; Bruno, R.S. Anti-Inflammatory Activities of Green Tea Catechins Along the Gut-Liver Axis in Nonalcoholic Fatty Liver Disease: Lessons Learned from Preclinical and Human Studies. J. Nutr. Biochem. 2020, 85, 108478. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Lai, L.; You, Y.; Gao, R.; Xiang, J.; Wang, G.; Yu, W. Quantitative Analysis of Bioactive Compounds in Commercial Teas: Profiling Catechin Alkaloids, Phenolic Acids, and Flavonols Using Targeted Statistical Approaches. Foods 2023, 12, 3098. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Xu, L.; Yang, L.; Wang, X. Epigallocatechin Gallate Is the Most Effective Catechin against Antioxidant Stress via Hydrogen Peroxide and Radical Scavenging Activity. Med. Sci. Monit. 2018, 24, 8198–8206. [Google Scholar] [CrossRef]
- Lee, M.J.; Maliakal, P.; Chen, L.; Meng, X.; Bondoc, F.Y.; Prabhu, S.; Lambert, G.; Mohr, S.; Yang, C.S. Pharmacokinetics of Tea Catechins after Ingestion of Green Tea and (-)-Epigallocatechin-3-Gallate by Humans: Formation of Different Metabolites and Individual Variability. Cancer Epidemiol. Biomarkers Prev. 2002, 11, 1025–1032. [Google Scholar] [PubMed]
- Kikuchi, T.; Hayashi, A.; Ikeda, N.; Morita, O.; Tasaki, J. Multidrug Resistance-Associated Protein 2 (Mrp2) Is an Efflux Transporter of EGCG and its Metabolites in the Human Small Intestine. J. Nutr. Biochem. 2022, 107, 109071. [Google Scholar] [CrossRef]
- Chan, K.Y.; Zhang, L.; Zuo, Z. Intestinal Efflux Transport Kinetics of Green Tea Catechins in Caco-2 Monolayer Model. J. Pharm. Pharmacol. 2007, 59, 395–400. [Google Scholar] [CrossRef]
- Saeki, K.; Hayakawa, S.; Nakano, S.; Ito, S.; Oishi, Y.; Suzuki, Y.; Isemura, M. In Vitro and in Silico Studies of the Molecular Interactions of Epigallocatechin-3-O-Gallate (EGCG) with Proteins That Explain the Health Benefits of Green Tea. Molecules 2018, 23, 1295. [Google Scholar] [CrossRef]
- Fusco, G.; Sanz-Hernandez, M.; Ruggeri, F.S.; Vendruscolo, M.; Dobson, C.M.; De Simone, A. Molecular Determinants of the Interaction of EGCG with Ordered and Disordered Proteins. Biopolymers 2018, 109, e23117. [Google Scholar] [CrossRef]
- Moeller, M.J.; Tenten, V. Renal Albumin Filtration: Alternative Models to the Standard Physical Barriers. Nat. Rev. Nephrol. 2013, 9, 266–277. [Google Scholar] [CrossRef]
- Li, M.; Hagerman, A.E. Role of the Flavan-3-Ol and Galloyl Moieties in the Interaction of (-)-Epigallocatechin Gallate with Serum Albumin. J. Agric. Food. Chem. 2014, 62, 3768–3775. [Google Scholar] [CrossRef]
- Tang, N.; Liu, J.; Cheng, Y. Effects of Zinc Binding on the Binding of Epigallocatechin Gallate (Green Tea) to Bovine Serum Albumin and Myoglobin. Food Chem. 2021, 357, 129750. [Google Scholar] [CrossRef] [PubMed]
- Bernatoniene, J.; Kopustinskiene, D.M. The Role of Catechins in Cellular Responses to Oxidative Stress. Molecules 2018, 23, 965. [Google Scholar] [CrossRef] [PubMed]
- Azman, N.A.M.; Peiró, S.; Fajarí, L.; Julià, L.; Almajano, M.P. Radical Scavenging of White Tea and Its Flavonoid Constituents by Electron Paramagnetic Resonance (Epr) Spectroscopy. J. Agric. Food Chem. 2014, 62, 5743–5748. [Google Scholar] [CrossRef]
- Sasaki, G.Y.; Vodovotz, Y.; Yu, Z.; Bruno, R.S. Catechin Bioavailability Following Consumption of a Green Tea Extract Confection Is Reduced in Obese Persons without Affecting Gut Microbial-Derived Valerolactones. Antioxidants 2022, 11, 2490. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.A.; Benedict, F.G. A Biometric Study of Human Basal Metabolism. Proc. Natl. Acad. Sci. USA 1918, 4, 370–373. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Vance, T.M.; Chun, O.K. Estimated Intake and Major Food Sources of Flavonoids among US Adults: Changes between 1999–2002 and 2007–2010 in NHANES. Eur. J. Nutr. 2016, 55, 833–843. [Google Scholar] [CrossRef] [PubMed]
- Hodges, J.K.; Zhu, J.; Yu, Z.; Vodovotz, Y.; Brock, G.; Sasaki, G.Y.; Dey, P.; Bruno, R.S. Intestinal-Level Anti-Inflammatory Bioactivities of Catechin-Rich Green Tea: Rationale, Design, and Methods of a Double-Blind, Randomized, Placebo-Controlled Crossover Trial in Metabolic Syndrome and Healthy Adults. Contemp. Clin Trials Commun. 2020, 17, 100495. [Google Scholar] [CrossRef] [PubMed]
- Neilson, A.P.; Green, R.J.; Wood, K.V.; Ferruzzi, M.G. High-Throughput Analysis of Catechins and Theaflavins by High Performance Liquid Chromatography with Diode Array Detection. J. Chromatogr. A 2006, 1132, 132–140. [Google Scholar] [CrossRef]
- Stefanovski, D.; Moate, P.J.; Boston, R.C. WinSAAM: A Windows-Based Compartmental Modeling System. Metabolism 2003, 52, 1153–1166. [Google Scholar] [CrossRef]
- Retzlaff, J.A.; Tauxe, W.N.; Kiely, J.M.; Stroebel, C.F. Erythrocyte Volume, Plasma Volume, and Lean Body Mass in Adult Men and Women. Blood 1969, 33, 649–661. [Google Scholar] [CrossRef]
- Scholl, C.; Lepper, A.; Lehr, T.; Hanke, N.; Schneider, K.L.; Brockmoller, J.; Seufferlein, T.; Stingl, J.C. Population Nutrikinetics of Green Tea Extract. PLoS ONE 2018, 13, e0193074. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.Y.; Li, X.M.; Liang, J.P.; Xiang, L.P.; Wang, K.R.; Shi, Y.L.; Yang, R.; Shi, M.; Ye, J.H.; Lu, J.L.; et al. Bioavailability of Tea Catechins and its Improvement. Molecules 2018, 23, 2346. [Google Scholar] [CrossRef] [PubMed]
- Law, F.C.P.; Yao, M.; Bi, H.C.; Lam, S. Physiologically Based Pharmacokinetic Modeling of Tea Catechin Mixture in Rats and Humans. Pharmacol. Res. Perspect. 2017, 5, e00305. [Google Scholar] [CrossRef] [PubMed]
- Stalmach, A.; Mullen, W.; Steiling, H.; Williamson, G.; Lean, M.E.; Crozier, A. Absorption, Metabolism, and Excretion of Green Tea Flavan-3-Ols in Humans with an Ileostomy. Mol. Nutr. Food Res. 2010, 54, 323–334. [Google Scholar] [CrossRef]
- Abe, S.K.; Inoue, M. Green Tea and Cancer and Cardiometabolic Diseases: A Review of the Current Epidemiological Evidence. Eur. J. Clin. Nutr. 2021, 75, 865–876. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Nagar, N.; Kumar, V.; Joshi, N.; Roy, P.; Poluri, K.M. Gallate Moiety of Catechin Is Essential for Inhibiting Ccl2 Chemokine-Mediated Monocyte Recruitment. J. Agric. Food Chem. 2023, 71, 4990–5005. [Google Scholar] [CrossRef]
- Sun, X.; Dey, P.; Bruno, R.S.; Zhu, J. EGCG and Catechin Relative to Green Tea Extract Differentially Modulate the Gut Microbial Metabolome and Liver Metabolome to Prevent Obesity in Mice Fed a High-Fat Diet. J. Nutr. Biochem. 2022, 109, 109094. [Google Scholar] [CrossRef]
- Ozduran, G.; Becer, E.; Vatansever, H.S.; Yucecan, S. Neuroprotective Effects of Catechins in an Experimental Parkinson’s Disease Model and Sk-N-as Cells: Evaluation of Cell Viability, Anti-Inflammatory and Anti-Apoptotic Effects. Neurol. Res. 2022, 44, 511–523. [Google Scholar] [CrossRef]
- Djerir, D.; Iddir, M.; Bourgault, S.; Lamy, S.; Annabi, B. Biophysical Evidence for Differential Gallated Green Tea Catechins Binding to Membrane Type-1 Matrix Metalloproteinase and its Interactors. Biophys. Chem. 2018, 234, 34–41. [Google Scholar] [CrossRef]
- Lnenickova, K.; Prochazkova, E.; Skalova, L.; Matouskova, P.; Bartikova, H.; Soucek, P.; Szotakova, B. Catechins Variously Affect Activities of Conjugation Enzymes in Proliferating and Differentiated Caco-2 Cells. Molecules 2016, 21, 1186. [Google Scholar] [CrossRef]
- Apak, R.; Guclu, K.; Ozyurek, M.; Karademir, S.E. Novel Total Antioxidant Capacity Index for Dietary Polyphenols and Vitamins C and E, Using Their Cupric Ion Reducing Capability in the Presence of Neocuproine: Cuprac Method. J. Agric. Food Chem. 2004, 52, 7970–7981. [Google Scholar] [CrossRef] [PubMed]
- Kurbitz, C.; Heise, D.; Redmer, T.; Goumas, F.; Arlt, A.; Lemke, J.; Rimbach, G.; Kalthoff, H.; Trauzold, A. Epicatechin Gallate and Catechin Gallate Are Superior to Epigallocatechin Gallate in Growth Suppression and Anti-Inflammatory Activities in Pancreatic Tumor Cells. Cancer Sci. 2011, 102, 728–734. [Google Scholar] [CrossRef]
- Matic, S.; Jadrijevic-Mladar Takac, M.; Barbaric, M.; Lucic, B.; Gall Troselj, K.; Stepanic, V. The Influence of in Vivo Metabolic Modifications on Admet Properties of Green Tea Catechins-in Silico Analysis. J. Pharm. Sci. 2018, 107, 2957–2964. [Google Scholar] [CrossRef] [PubMed]
- De Simone, G.; di Masi, A.; Ascenzi, P. Serum Albumin: A Multifaced Enzyme. Int. J. Mol. Sci. 2021, 22, 86. [Google Scholar] [CrossRef] [PubMed]
- Gburek, J.; Konopska, B.; Golab, K. Renal Handling of Albumin-from Early Findings to Current Concepts. Int. J. Mol. Sci. 2021, 22, 5809. [Google Scholar] [CrossRef]
- Henning, S.M.; Choo, J.J.; Heber, D. Nongallated Compared with Gallated Flavan-3-Ols in Green and Black Tea Are More Bioavailable. J. Nutr. 2008, 138, 1529S–1534S. [Google Scholar] [CrossRef]
- Fanali, G.; di Masi, A.; Trezza, V.; Marino, M.; Fasano, M.; Ascenzi, P. Human Serum Albumin: From Bench to Bedside. Mol. Aspects Med. 2012, 33, 209–290. [Google Scholar] [CrossRef]
- Zeng, L.; Ma, M.; Li, C.; Luo, L. Stability of Tea Polyphenols Solution with Different pH at Different Temperatures. Int J Food Prop 2017, 20, 1–18. [Google Scholar] [CrossRef]
- Qin, W.; Ketnawa, S.; Ogawa, Y. Effect of Digestive Enzymes and pH on Variation of Bioavailability of Green Tea During Simulated in Vitro Gastrointestinal Digestion. Food Sci. Hum. Wellness 2022, 11, 669–675. [Google Scholar] [CrossRef]
- Record, I.R.; Lane, J.M. Simulated Intestinal Digestion of Green and Black Teas. Food Chem. 2001, 73, 481–486. [Google Scholar] [CrossRef]
- Mena, P.; Bresciani, L.; Brindani, N.; Ludwig, I.A.; Pereira-Caro, G.; Angelino, D.; Llorach, R.; Calani, L.; Brighenti, F.; Clifford, M.N.; et al. Phenyl-Gamma-Valerolactones and Phenylvaleric Acids, the Main Colonic Metabolites of Flavan-3-Ols: Synthesis, Analysis, Bioavailability, and Bioactivity. Nat. Prod. Rep. 2019, 36, 714–752. [Google Scholar] [CrossRef] [PubMed]
- Van Amelsvoort, J.M.; Van Hof, K.H.; Mathot, J.N.; Mulder, T.P.; Wiersma, A.; Tijburg, L.B. Plasma Concentrations of Individual Tea Catechins after a Single Oral Dose in Humans. Xenobiotica 2001, 31, 891–901. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hodges, J.K.; Sasaki, G.Y.; Vodovotz, Y.; Bruno, R.S. Gallation and B-Ring Dihydroxylation Increase Green Tea Catechin Residence Time in Plasma by Differentially Affecting Tissue-Specific Trafficking: Compartmental Model of Catechin Kinetics in Healthy Adults. Nutrients 2023, 15, 4021. https://doi.org/10.3390/nu15184021
Hodges JK, Sasaki GY, Vodovotz Y, Bruno RS. Gallation and B-Ring Dihydroxylation Increase Green Tea Catechin Residence Time in Plasma by Differentially Affecting Tissue-Specific Trafficking: Compartmental Model of Catechin Kinetics in Healthy Adults. Nutrients. 2023; 15(18):4021. https://doi.org/10.3390/nu15184021
Chicago/Turabian StyleHodges, Joanna K., Geoffrey Y. Sasaki, Yael Vodovotz, and Richard S. Bruno. 2023. "Gallation and B-Ring Dihydroxylation Increase Green Tea Catechin Residence Time in Plasma by Differentially Affecting Tissue-Specific Trafficking: Compartmental Model of Catechin Kinetics in Healthy Adults" Nutrients 15, no. 18: 4021. https://doi.org/10.3390/nu15184021
APA StyleHodges, J. K., Sasaki, G. Y., Vodovotz, Y., & Bruno, R. S. (2023). Gallation and B-Ring Dihydroxylation Increase Green Tea Catechin Residence Time in Plasma by Differentially Affecting Tissue-Specific Trafficking: Compartmental Model of Catechin Kinetics in Healthy Adults. Nutrients, 15(18), 4021. https://doi.org/10.3390/nu15184021