


Dietary Acid Load and Cancer Risk: A Review of the Uruguayan Experience


Abstract
1. Introduction
1.1. Dietary Acid Load
1.2. Dietary Acid Load and Cancer
2. Case-Control Studies in Uruguay
2.1. The Study Cohort
2.2. The Employed Questionnaires
2.3. Dietary Acid Load Estimation
2.4. Nutrient Intake Estimation
2.5. Statistical and Epidemiological Analyses Practices
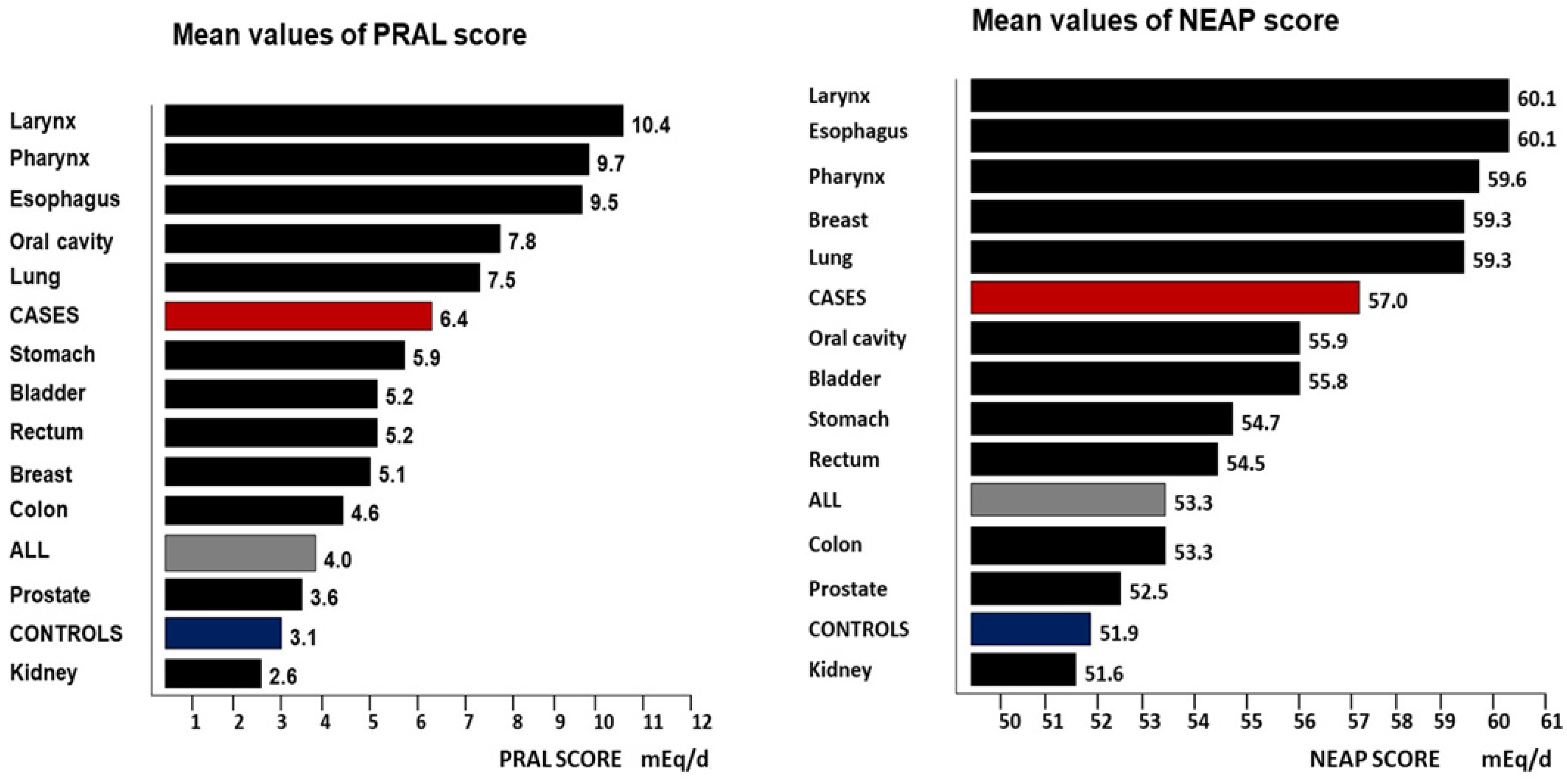
3. Results
4. Discussion
4.1. DAL Scores and Histopathological Features
4.2. DAL and Methionine Intake
4.3. DAL, Phosphorus, and Magnesium Intake
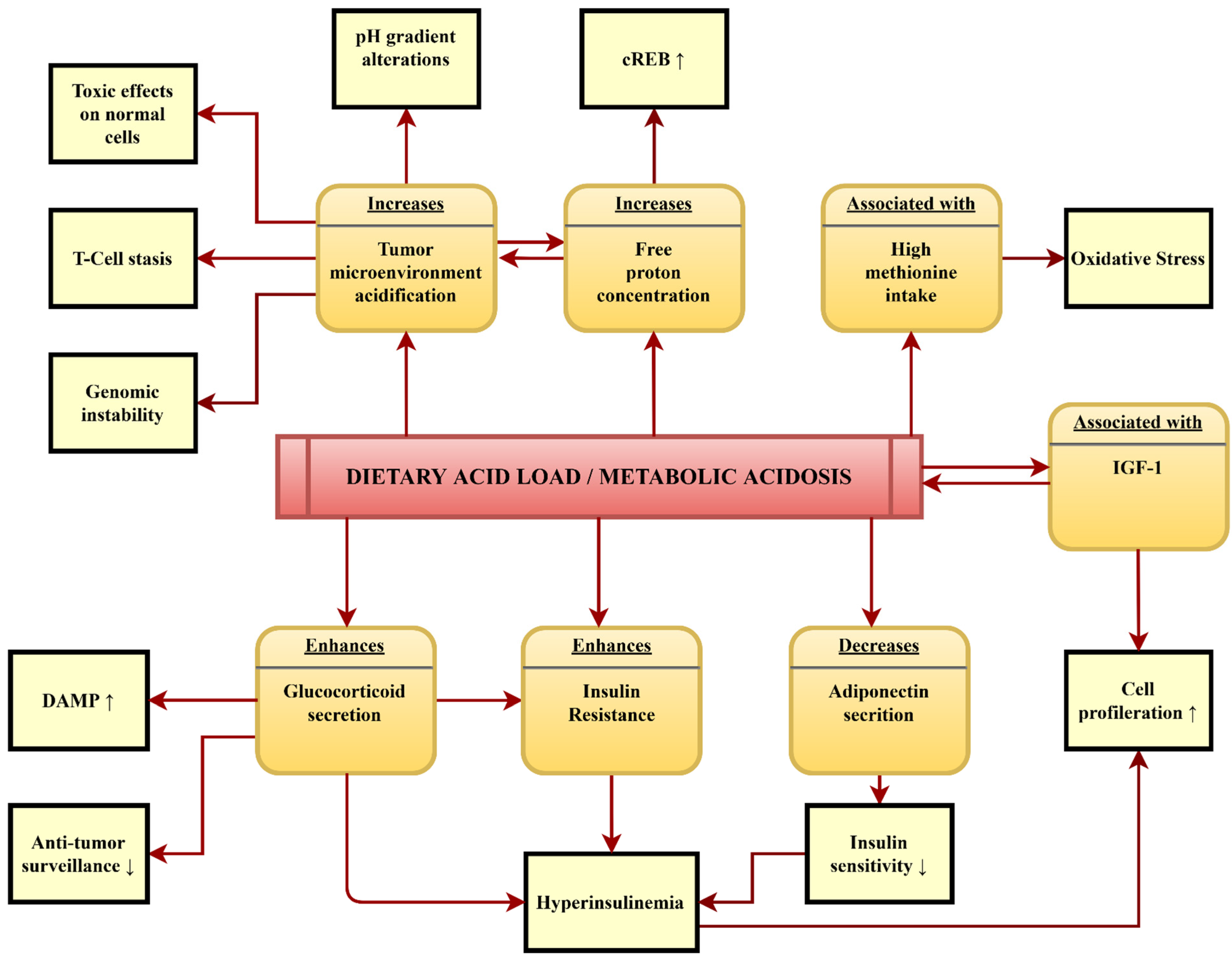
4.4. Potential Pathomechanisms Mediating the Association between DAL and Cancer
4.5. Limitations of Our Database and Future Fields of Study
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tran, K.B.; Lang, J.J.; Compton, K.; Xu, R.; Acheson, A.R.; Henrikson, H.J.; Kocarnik, J.M.; Penberthy, L.; Aali, A.; Abbas, Q.; et al. The global burden of cancer attributable to risk factors, 2010–2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2022, 400, 563–591. [Google Scholar] [CrossRef] [PubMed]
- Friedenreich, C.M.; Ryder-Burbidge, C.; McNeil, J. Physical activity, obesity and sedentary behavior in cancer etiology: Epidemiologic evidence and biologic mechanisms. Mol. Oncol. 2021, 15, 790–800. [Google Scholar] [CrossRef] [PubMed]
- Kamal, N.; Ilowefah, M.A.; Hilles, A.R.; Anua, N.A.; Awin, T.; Alshwyeh, H.A.; Aldosary, S.K.; Jambocus, N.G.S.; Alosaimi, A.A.; Rahman, A.; et al. Genesis and mechanism of some cancer types and an overview on the role of diet and nutrition in cancer prevention. Molecules 2022, 27, 1794. [Google Scholar] [CrossRef] [PubMed]
- Steck, S.E.; Murphy, E.A. Dietary patterns and cancer risk. Nat. Rev. Cancer 2020, 20, 125–138. [Google Scholar] [CrossRef]
- United Nations Development Program. Available online: http://hdr.undp.org/en/content/2019-human-development-index-ranking (accessed on 20 June 2023).
- Faostat. Food and Agriculture Organization. Available online: http://www.fao.org/faostat/en/#data/CL (accessed on 20 June 2023).
- De Stefani, E.; Oreggia, F.; Ronco, A.L.; Fierro, L.; Rivero, S. Salted meat consumption as a risk factor for cancer for the oral cavity and pharynx: A case-control study from Uruguay. Cancer Epidemiol. Biomark. Prev. 1994, 3, 381–385. [Google Scholar]
- De Stefani, E.; Correa, P.; Boffetta, P.; Deneo-Pellegrini, H.; Ronco, A.L.; Mendilaharsu, M. Dietary patterns and risk of gastric cancer: A case-control study in Uruguay. Gastric Cancer 2004, 7, 211–220. [Google Scholar] [CrossRef]
- De Stefani, E.; Boffetta, P.; Ronco, A.L.; Deneo-Pellegrini, H.; Acosta, G.; Mendilaharsu, M. Dietary patterns and risk of bladder cancer: A factor analysis in Uruguay. Cancer Causes Control. 2008, 19, 1243–1249. [Google Scholar] [CrossRef]
- De Stefani, E.; Deneo-Pellegrini, H.; Boffetta, P.; Ronco, A.L.; Aune, D.; Acosta, G.; Mendilaharsu, M.; Brennan, P.; Ferro, G. Dietary patterns and risk of cancer: A factor analysis in Uruguay. Int. J. Cancer 2009, 124, 1391–1397. [Google Scholar] [CrossRef]
- De Stefani, E.; Ronco, A.L.; Boffetta, P.; Deneo-Pellegrini, H.; Correa, P.; Acosta, G.; Mendilaharsu, M. Nutrient-derived dietary patterns and risk of colorectal cancer: A factor analysis in Uruguay. Asian Pac. J. Cancer Prev. 2012, 13, 231–235. [Google Scholar] [CrossRef]
- Ronco, A.L.; De Stefani, E.; Mendilaharsu, M.; Deneo-Pellegrini, H. Meat, fat and the risk of breast cancer: A case-control study from Uruguay. Int. J. Cancer 1996, 65, 328–331. [Google Scholar] [CrossRef]
- Ronco, A.L.; De Stefani, E.; Boffetta, P.; Deneo-Pellegrini, H.; Acosta, G.; Mendilaharsu, M. Food patterns and risk of breast cancer: A factor analysis study in Uruguay. Int. J. Cancer. 2006, 19, 1672–1678. [Google Scholar] [CrossRef] [PubMed]
- Ronco, A.L.; De Stefani, E.; Lasalvia-Galante, E.; Mendoza, B.; Vazquez, A.; Sanchez, G. Hot infusions and risk of colorectal cancer in Uruguay: A case-control study. Eur. J. Clin. Nutr. 2017, 71, 1429–1436. [Google Scholar] [CrossRef] [PubMed]
- Ronco, A.L.; Calderón, J.M.; Mendoza, B.A. Dietary iron, water intake and risk of urinary bladder cancer: A case-control study. World Cancer Res. J. 2020, 7, e1685. [Google Scholar]
- Aune, D.; De Stefani, E.; Ronco, A.; Boffetta, P.; Deneo-Pellegrini, H.; Acosta, G.; Mendilaharsu, M. Meat Consumption and Cancer Risk: A Case-Control Study in Uruguay. Asian Pac. J. Cancer Prev. 2009, 10, 429–436. [Google Scholar] [PubMed]
- De Stefani, E.; Aune, D.; Boffetta, P.; Deneo-Pellegrini, H.; Ronco, A.L.; Acosta, G.; Brennan, P.; Ferro, G.; Mendilaharsu, M. Salted Meat Consumption and the Risk of Cancer: A Multisite Case-Control Study in Uruguay. Asian Pac. J. Cancer Prev. 2009, 10, 853–857. [Google Scholar] [PubMed]
- Pizzorno, J.; Frassetto, L.A.; Katzinger, J. Diet-induced acidosis: Is it real and clinically relevant? Br. J. Nutr. 2009, 103, 1185–1194. [Google Scholar] [CrossRef]
- Carnauba, R.; Baptistella, A.; Paschoal, V.; Hübscher, G. Diet-Induced Low-Grade Metabolic Acidosis and Clinical Outcomes: A Review. Nutrients 2017, 9, 538. [Google Scholar] [CrossRef]
- Remer, T.; Manz, F. Estimation of the renal net acid excretion by adults consuming diets containing variable amounts of protein. Am. J. Clin. Nutr. 1994, 59, 1356–1361. [Google Scholar] [CrossRef]
- Frassetto, L.A.; Todd, K.M.; Curtis Morris, R., Jr.; Sebastian, A. Estimation of net endogenous noncarbonic acid production in humans from diet potassium and protein. Am. J. Clin. Nutr. 1998, 68, 576–583. [Google Scholar] [CrossRef]
- Storz, M.A.; Ronco, A.L. How Well Do Low-PRAL Diets Fare in Comparison to the 2020–2025 Dietary Guidelines for Americans? Healthcare 2023, 11, 180. [Google Scholar] [CrossRef]
- Abbasalizad Farhangi, M.; Nikniaz, L.; Nikniaz, Z. Higher dietary acid load potentially increases serum triglyceride and obesity prevalence in adults: An updated systematic review and meta-analysis. PLoS ONE 2019, 14, e0216547. [Google Scholar] [CrossRef] [PubMed]
- Robey, I.F. Examining the Relationship between Diet-Induced Acidosis and Cancer. Nutr. Metab. 2012, 9, 72. [Google Scholar] [CrossRef]
- Shi, Q.; Maas, L.; Veith, C.; Van Schooten, F.J.; Godschalk, R.W. Acidic cellular microenvironment modifies carcinogen-induced DNA damage and repair. Arch. Toxicol. 2017, 91, 2425–2441. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.X.; Choi, S.Y.C.; Niu, X.; Kang, N.; Xue, H.; Killam, J.; Wang, Y. Lactic Acid and an Acidic Tumor Microenvironment Suppress Anticancer Immunity. Int. J. Mol. Sci. 2020, 21, 8363. [Google Scholar] [CrossRef]
- Wada, H.; Hamaguchi, R.; Narui, R.; Morikawa, H. Meaning and significance of “Alkalization Therapy for Cancer”. Front. Oncol. 2022, 12, 920843. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Wen, Z.-Y.; Liu, F.-H.; Wei, Y.-F.; Xu, H.-L.; Sun, M.-L.; Zhao, Y.-H.; Gong, T.-T.; Wang, H.-H.; Wu, Q.-J. Association between Dietary Acid Load and Cancer Risk and Prognosis: An Updated Systematic Review and Meta-Analysis of Observational Studies. Front. Nutr. 2022, 9, 891936. [Google Scholar] [CrossRef] [PubMed]
- Keramati, M.; Kheirouri, S.; Musazadeh, V.; Alizadeh, M. Association of high dietary acid load with the risk of cancer: A systematic review and meta-analysis of observational studies. Front. Nutr. 2022, 9, 816797. [Google Scholar] [CrossRef]
- Bahrami, A.; Khalesi, S.; Ghafouri-Taleghani, F.; Alibeyk, S.; Hajigholam-Saryazdi, M.; Haghighi, S.; Ehsan, H. Dietary acid load and the risk of cancer: A systematic review and dose-response meta-analysis of observational studies. Eur. J. Cancer Prev. 2022, 31, 577–584. [Google Scholar] [CrossRef]
- Park, Y.-M.M.; Steck, S.E.; Fung, T.T.; Merchant, A.T.; Elizabeth Hodgson, M.; Keller, J.A.; Sandler, D.P. Higher Diet-Dependent Acid Load Is Associated with Risk of Breast Cancer: Findings from the Sister Study. Int. J. Cancer 2019, 144, 1834–1843. [Google Scholar] [CrossRef]
- Jafari Nasab, S.; Rafiee, P.; Bahrami, A.; Rezaeimanesh, N.; Rashidkhani, B.; Sohrab, G.; Naja, F.; Hejazi, E.; Sadeghi, A. Diet-Dependent Acid Load and the Risk of Colorectal Cancer and Adenoma: A Case-Control Study. Public Health Nutr. 2021, 24, 4474–4481. [Google Scholar] [CrossRef]
- Shi, L.-W.; Wu, Y.-L.; Hu, J.-J.; Yang, P.-F.; Sun, W.-P.; Gao, J.; Wang, K.; Peng, Y.; Wu, J.-J.; Zhong, G.-C. Dietary Acid Load and the Risk of Pancreatic Cancer: A Prospective Cohort Study. Cancer Epidemiol. Biomark. Prev. 2021, 30, 1009–1019. [Google Scholar] [CrossRef] [PubMed]
- Milajerdi, A.; Shayanfar, M.; Benisi-Kohansal, S.; Mohammad-Shirazi, M.; Sharifi, G.; Tabibi, H.; Esmaillzadeh, A. A Case-Control Study on Dietary Acid Load in Relation to Glioma. Nutr. Cancer 2022, 74, 1644–1651. [Google Scholar] [CrossRef] [PubMed]
- Passey, C. Reducing the Dietary Acid Load: How a More Alkaline Diet Benefits Patients with Chronic Kidney Disease. J. Ren. Nutr. 2017, 27, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Stroup, B.M.; Sawin, E.A.; Murali, S.G.; Binkley, N.; Hansen, K.E.; Ney, D.M. Amino Acid Medical Foods Provide a High Dietary Acid Load and Increase Urinary Excretion of Renal Net Acid, Calcium, and Magnesium Compared with Glycomacropeptide Medical Foods in Phenylketonuria. J. Nutr. Metab. 2017, 2017, e1909101. [Google Scholar] [CrossRef]
- Herter, J.; Lederer, A.-K.; Ronco, A.L.; Hannibal, L.; Huber, R.; Storz, M.A. Dietary Acid Load Correlates with Serum Amino Acid Concentrations after a Four-Week Intervention with Vegan vs. Meat-Rich Diets: A Secondary Data Analysis. Nutrients 2023, 15, 2942. [Google Scholar] [CrossRef]
- Osuna-Padilla, I.A.; Leal-Escobar, G.; Garza-García, C.A.; Rodríguez-Castellanos, F.E. Dietary Acid Load: Mechanisms and evidence of its health repercussions. Nefrologia 2019, 39, 343–354. [Google Scholar] [CrossRef]
- Ronco, A.L.; Martínez-López, W.; Calderón, J.M.; Mendoza, B.A. Dietary acid load and colorectal cancer risk: A case-control study. World Cancer Res. J. 2020, 7, e1750. [Google Scholar]
- Ronco, A.L.; Martínez-López, W.; Calderón, J.M.; Golomar, W. Dietary acid load and lung cancer risk: A case-control study in men. Cancer Treat. Res. Commun. 2021, 28, 100382. [Google Scholar] [CrossRef]
- Ronco, A.L.; Martínez-López, W.; Mendoza, B.A.; Calderón, J.M. Epidemiologic evidence for association between a high dietary acid load and the breast cancer risk. SciMedicine J. 2021, 3, 166–176. [Google Scholar] [CrossRef]
- Ronco, A.L.; Storz, M.A.; Martínez-López, W.; Calderón, J.M.; Golomar, W. High dietary acid load is associated with prostate cancer risk: An epidemiological study. World Cancer Res. J. 2021, 8, e2119. [Google Scholar]
- Ronco, A.L.; Storz, M.A.; Martínez-López, W.; Calderón, J.M.; Golomar, W. Dietary acid load and risk of kidney cancer: An epidemiologic case-control study. World Cancer Res. J. 2021, 8, e2096. [Google Scholar]
- Ronco, A.L.; Storz, M.A.; Martínez-López, W.; Calderón, J.M.; Golomar, W. Dietary acid load and bladder cancer risk: An epidemiologic case-control study. Multidiscip. Cancer Investig. 2022, 6, 1–12. [Google Scholar] [CrossRef]
- Ronco, A.L.; Martínez-López, W.; Calderón, J.M.; Storz, M.A. Dietary acid load and risk of head and neck and oral cavity cancers: An epidemiologic study. Oral Sci. Int. 2022, 20, 78–87. [Google Scholar] [CrossRef]
- Ronco, A.L.; Martínez-López, W.; Calderón, J.M.; Storz, M.A. Dietary acid load and esophageal cancer risk: A case-control study. Thorac. Cancer 2022, 13, 2759–2766. [Google Scholar] [CrossRef]
- Ronco, A.L.; Martínez-López, W.; Calderón, J.M.; Mendoza, B.; Storz, M.A. Dietary acid load and risk of gastric cancer: A case-control study. World Cancer Res. J. 2022, 9, e2403. [Google Scholar]
- Ronco, A.L.; Martínez-López, W.; Calderón, J.M.; Mintegui, M.; Storz, M.A. Dietary acid load and risk of cancer: New insights from a nationwide case-control study. World Cancer Res. J. 2023, 10, e2547. [Google Scholar]
- Mazzei, M.E.; Puchulu, M.R.; Rochaix, M.A. Table of Food Chemical Composition, 2nd ed.; Cenexa y Feiden Publishers: Buenos Aires, Argentina, 1995. (In Spanish) [Google Scholar]
- Food Data Central. United States Department of Agriculture. Available online: https://fdc.nal.usda.gov/fdc-app.html#/?component=1215 (accessed on 1 December 2022).
- Storz, M.A.; Ronco, A.L.; Hannibal, L. Observational and Clinical Evidence That Plant-Based Nutrition Reduces Dietary Acid Load. J. Nutr. Sci. 2022, 11, e93. [Google Scholar] [CrossRef]
- Elango, R. Methionine nutrition and metabolism: Insights from animal studies to inform human nutrition. J. Nutr. 2020, 150, 2518S–2523S. [Google Scholar] [CrossRef]
- McCarty, M.F.; Barroso-Aranda, J.; Contreras, F. The Low-Methionine Content of Vegan Diets May Make Methionine Restriction Feasible as a Life Extension Strategy. Med. Hypotheses 2009, 72, 125–128. [Google Scholar] [CrossRef]
- Nimni, M.E.; Han, B.; Cordoba, F. Are we getting enough sulfur in our diet? Nutr. Metab. 2007, 4, 24. [Google Scholar] [CrossRef]
- Buret, A.G.; Allain, T.; Motta, J.P.; Wallace, J.L. Effects of Hydrogen Sulfide on the Microbiome: From Toxicity to Therapy. Antioxid. Redox Signal. 2022, 36, 4–6. [Google Scholar] [CrossRef] [PubMed]
- Wanders, D.; Hobson, K.; Ji, X. Methionine Restriction and Cancer Biology. Nutrients 2020, 12, 684. [Google Scholar] [CrossRef] [PubMed]
- Harper, A.E.; Benevenga, N.J.; Wohlhueter, R.M. Effects of Ingestion of Disproportionate Amounts of Amino Acids. Physiol. Rev. 1970, 60, 429–558. [Google Scholar] [CrossRef] [PubMed]
- Celin, I.; Turkmen, I.I.; Kara, C.; Udum, D.; Orman, A.; Gencoglu, H. Effects of supplementation with rumen-protected choline and methionine on metabolic profile and some reproductive parameters in dairy cattle during transition period. Eur. J. Lipid Sci. Technol. 2022, 35, 474–481. [Google Scholar] [CrossRef]
- Coleman, D.N.; Totakul, P.; Onjai-uea, N.; Aboragah, A.; Jiang, Q.; Vailati-Riboni, M.; Pate, R.T.; Luchini, D.; Paengkoum, P.; Wanapat, M.; et al. Rumen-protected methionine during heat stress alters mTOR, insulin signaling, and 1-carbon metabolism protein abundance in liver, and whole-blood transsulfuration pathway genes in Holstein cows. J. Dairy Sci. 2022, 105, 7787–7804. [Google Scholar] [CrossRef]
- Kachungwa Lugata, J.; Ortega, A.D.S.V.; Szabó, C. The Role of Methionine Supplementation on Oxidative Stress and Antioxidant Status of Poultry—A Review. Agriculture 2022, 12, 1701. [Google Scholar] [CrossRef]
- Cynober, L.; Bier, D.M.; Stover, P.; Kadowaki, M.; Morris, S.M., Jr.; Elango, R.; Smriga, M. Proposals for upper limits of safe intake for methionine, histidine, and lysine in healthy humans. J. Nutr. 2020, 150, 2606S–2608S. [Google Scholar] [CrossRef]
- Storz, M.A.; Ronco, A.L. The 1995 potential renal acid load (PRAL) values may no longer adequately reflect the actual acid-base impact of certain foods: A hypothesis. Nutr. Health 2023. [Google Scholar] [CrossRef]
- Zhang, Y.; Jelleschitz, J.; Grune, T.; Chen, W.; Zhao, Y.; Jia, M.; Wang, Y.; Liu, Z.; Höhn, A. Methionine Restriction—Association with Redox Homeostasis and Implications on Aging and Diseases. Redox Biol. 2022, 57, 102464. [Google Scholar] [CrossRef]
- Ritz, E.; Hahn, K.; Ketteler, M.; Kuhlmann, M.K.; Mann, J. Phosphate additives in food—A health risk. Dtsch. Ärzteblatt Int. 2012, 109, 49–55. [Google Scholar] [CrossRef]
- Brown, R.B.; Razzaque, M.S. Phosphate toxicity and tumorigenesis. Biochim. Et Biophys. Acta Rev. Cancer 2018, 1869, 303–309. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA) Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific opinion on dietary reference values for phosphorus. EFSA J. 2015, 13, 4185. [Google Scholar] [CrossRef]
- Hannah, J. Phosphorus in food: Limitations of food composition data. J. Kidney Care 2018, 3, 362–367. [Google Scholar] [CrossRef]
- Carrigan, A.; Klinger, A.; Choquette, S.S.; Luzuriaga-McPherson, B.A.; Bell, E.K.; Darnell, B.; Gutierrez, O.M. Contribution of food additives to sodium and phosphorus content of diets rich in processed foods. J. Ren. Nutr. 2014, 24, 13–19. [Google Scholar] [CrossRef]
- Frassetto, L.A.; Morris, R.C., Jr.; Sebastian, A. Dietary sodium chloride intake independently predicts the degree of hyperchloremic metabolic acidosis in healthy humans consuming a net acid-producing diet. Am. J. Physiol. Ren. Physiol. 2007, 293, F521–F525. [Google Scholar] [CrossRef] [PubMed]
- Hijmans, R.S.; Shrestha, P.; Sarpong, K.A.; Yazdani, S.; el Masri, R.; de Jong, W.H.A.; Navis, G.; Vives, R.R.; van den Born, J. High sodium diet converts renal proteoglycans into pro-inflammatory mediators in rats. PLoS ONE 2017, 12, e0178940. [Google Scholar] [CrossRef]
- Fiorentini, D.; Cappadone, C.; Farruggia, G.; Prata, C. Magnesium: Biochemistry, Nutrition, Detection, and Social Impact of Diseases Linked to Its Deficiency. Nutrients 2021, 13, 1136. [Google Scholar] [CrossRef]
- Da Silva Lima, F.; Fock, R.A. A review of the action of magnesium on several processes involved in the modulation of hematopoiesis. Int. J. Mol. Sci. 2020, 21, 7084. [Google Scholar] [CrossRef]
- Williams, R.S.; Heilbronn, L.K.; Chen, D.L.; Coster, A.C.F.; Greenfield, J.R.; Samocha-Bonet, D. Dietary Acid Load, Metabolic Acidosis and Insulin Resistance—Lessons from Cross-Sectional and Overfeeding Studies in Humans. Clin. Nutr. 2016, 35, 1084–1090. [Google Scholar] [CrossRef]
- Williams, R.S.; Kozan, P.; Samocha-Bonet, D. The Role of Dietary Acid Load and Mild Metabolic Acidosis in Insulin Resistance in Humans. Biochimie 2016, 124, 171–177. [Google Scholar] [CrossRef]
- Chiefari, E.; Mirabelli, M.; La Vignera, S.; Tanyolaç, S.; Foti, D.P.; Aversa, A.; Brunetti, A. Insulin Resistance and Cancer: In Search for a Causal Link. Int. J. Mol. Sci. 2021, 22, 11137. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.M.Y.; Wellberg, E.A.; Kopp, J.L.; Johnson, J.D. Hyperinsulinemia in Obesity, Inflammation, and Cancer. Diabetes Metab. J. 2021, 45, 285–311. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, E.J.; LeRoith, D. Hyperinsulinaemia in Cancer. Nat. Rev. Cancer 2020, 20, 629–644. [Google Scholar] [CrossRef] [PubMed]
- Arcidiacono, B.; Iiritano, S.; Nocera, A.; Possidente, K.; Nevolo, M.T.; Ventura, V.; Foti, D.; Chiefari, E.; Brunetti, A. Insulin Resistance and Cancer Risk: An Overview of the Pathogenetic Mechanisms. Exp. Diabetes Res. 2012, 2012, 789174. [Google Scholar] [CrossRef] [PubMed]
- Shanmugalingam, T.; Bosco, C.; Ridley, A.J.; Van Hemelrijck, M. Is There a Role for IGF-1 in the Development of Second Primary Cancers? Cancer Med. 2016, 5, 3353–3367. [Google Scholar] [CrossRef]
- Klement, R.J.; Fink, M.K. Dietary and Pharmacological Modification of the Insulin/IGF-1 System: Exploiting the Full Repertoire against Cancer. Oncogenesis 2016, 5, e193. [Google Scholar] [CrossRef]
- Müller, A.; Zimmermann-Klemd, A.M.; Lederer, A.-K.; Hannibal, L.; Kowarschik, S.; Huber, R.; Storz, M.A. A Vegan Diet Is Associated with a Significant Reduction in Dietary Acid Load: Post Hoc Analysis of a Randomized Controlled Trial in Healthy Individuals. Int. J. Environ. Res. Public Health 2021, 18, 9998. [Google Scholar] [CrossRef]
- Lim, S.Y.; Chan, Y.M.; Ramachandran, V.; Shariff, Z.M.; Chin, Y.S.; Arumugam, M. Dietary Acid Load and Its Interaction with IGF1 (Rs35767 and Rs7136446) and IL6 (Rs1800796) Polymorphisms on Metabolic Traits among Postmenopausal Women. Nutrients 2021, 13, 2161. [Google Scholar] [CrossRef]
- Levine, M.E.; Suarez, J.A.; Brandhorst, S.; Balasubramanian, P.; Cheng, C.-W.; Madia, F.; Fontana, L.; Mirisola, M.G.; Guevara-Aguirre, J.; Wan, J.; et al. Low Protein Intake Is Associated with a Major Reduction in IGF-1, Cancer, and Overall Mortality in the 65 and Younger but Not Older Population. Cell Metab. 2014, 19, 407–417. [Google Scholar] [CrossRef]
- Gulick, C.N.; Peddie, M.C.; Jowett, T.; Hackney, A.C.; Rehrer, N.J. Exercise, Dietary Protein, and Combined Effect on IGF-1. Int. J. Sci. Res. Methodol. 2020, 16, 61–77. [Google Scholar]
- Esche, J.; Shi, L.; Sánchez-Guijo, A.; Hartmann, M.F.; Wudy, S.A.; Remer, T. Higher Diet-Dependent Renal Acid Load Associates with Higher Glucocorticoid Secretion and Potentially Bioactive Free Glucocorticoids in Healthy Children. Kidney Int. 2016, 90, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Souto, G.; Donapetry, C.; Calviño, J.; Adeva, M.M. Metabolic Acidosis-Induced Insulin Resistance and Cardiovascular Risk. Metab. Syndr. Relat. Disord. 2011, 9, 247–253. [Google Scholar] [CrossRef]
- Janssen, J.A.M.J.L. New Insights into the Role of Insulin and Hypothalamic-Pituitary-Adrenal (HPA) Axis in the Metabolic Syndrome. Int. J. Mol. Sci. 2022, 23, 8178. [Google Scholar] [CrossRef] [PubMed]
- Khadka, S.; Druffner, S.R.; Duncan, B.C.; Busada, J.T. Glucocorticoid Regulation of Cancer Development and Progression. Front. Endocrinol. 2023, 14, 1161768. [Google Scholar] [CrossRef] [PubMed]
- Fallo, F.; Scarda, A.; Sonino, N.; Paoletta, A.; Boscaro, M.; Pagano, C.; Federspil, G.; Vettor, R. Effect of Glucocorticoids on Adiponectin: A Study in Healthy Subjects and in Cushing’s Syndrome. Eur. J. Endocrinol. 2004, 150, 339–344. [Google Scholar] [CrossRef]
- de Oliveira, C.; de Mattos, A.B.; Biz, C.; Oyama, L.M.; Ribeiro, E.B.; Oller do Nascimento, C.M. High-Fat Diet and Glucocorticoid Treatment Cause Hyperglycemia Associated with Adiponectin Receptor Alterations. Lipids Health Dis. 2011, 10, 11. [Google Scholar] [CrossRef] [PubMed]
- Ziemke, F.; Mantzoros, C.S. Adiponectin in Insulin Resistance: Lessons from Translational Research. Am. J. Clin. Nutr. 2010, 91, 258S–261S. [Google Scholar] [CrossRef]
- Ahlstrom, P.; Rai, E.; Chakma, S.; Cho, H.H.; Rengasamy, P.; Sweeney, G. Adiponectin Improves Insulin Sensitivity via Activation of Autophagic Flux. J. Mol. Endocrinol. 2017, 59, 339–350. [Google Scholar] [CrossRef]
- Disthabanchong, S.; Niticharoenpong, K.; Radinahamed, P.; Stitchantrakul, W.; Ongphiphadhanakul, B.; Hongeng, S. Metabolic Acidosis Lowers Circulating Adiponectin through Inhibition of Adiponectin Gene Transcription. Nephrol. Dial. Transplant. 2011, 26, 592–598. [Google Scholar] [CrossRef]
- Williamson, M.; Moustaid-Moussa, N.; Gollahon, L. The Molecular Effects of Dietary Acid Load on Metabolic Disease (The Cellular PasaDoble: The Fast-Paced Dance of PH Regulation). Front. Mol. Med. 2021, 1, 777088. [Google Scholar] [CrossRef]
- Gillies, R.J.; Pilot, C.; Marunaka, Y.; Fais, S. Targeting Acidity in Cancer and Diabetes. Biochim. Et Biophys. Acta Rev. Cancer 2019, 1871, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Cassim, S.; Vučetić, M.; Ždralević, M.; Pouyssegur, J. Warburg and Beyond: The Power of Mitochondrial Metabolism to Collaborate or Replace Fermentative Glycolysis in Cancer. Cancers 2020, 12, 1119. [Google Scholar] [CrossRef] [PubMed]
- Moellering, R.E.; Black, K.C.; Krishnamurty, C.; Baggett, B.K.; Stafford, P.; Rain, M.; Gatenby, R.A.; Gillies, R.J. Acid Treatment of Melanoma Cells Selects for Invasive Phenotypes. Clin. Exp. Metastasis 2008, 25, 411–425. [Google Scholar] [CrossRef]
- Estrella, V.; Chen, T.; Lloyd, M.; Wojtkowiak, J.; Cornnell, H.H.; Ibrahim-Hashim, A.; Bailey, K.; Balagurunathan, Y.; Rothberg, J.M.; Sloane, B.F.; et al. Acidity Generated by the Tumor Microenvironment Drives Local Invasion. Cancer Res. 2013, 73, 1524–1535. [Google Scholar] [CrossRef] [PubMed]
- Gatenby, R.A.; Gillies, R.J. A Microenvironmental Model of Carcinogenesis. Nat. Rev. Cancer 2008, 8, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Lardner, A. The Effects of Extracellular PH on Immune Function. J. Leukoc. Biol. 2001, 69, 522–530. [Google Scholar] [CrossRef]
- Gillies, R.J.; Verduzco, D.; Gatenby, R.A. Evolutionary Dynamics of Carcinogenesis and Why Targeted Therapy Does Not Work. Nat. Rev. Cancer 2012, 12, 487–493. [Google Scholar] [CrossRef]
- Wilde, B.R.; Ye, Z.; Lim, T.-Y.; Ayer, D.E. Cellular Acidosis Triggers Human MondoA Transcriptional Activity by Driving Mitochondrial ATP Production. elife 2019, 8, e40199. [Google Scholar] [CrossRef]
- Chen, J.L.-Y.; Merl, D.; Peterson, C.W.; Wu, J.; Liu, P.Y.; Yin, H.; Muoio, D.M.; Ayer, D.E.; West, M.; Chi, J.-T. Lactic Acidosis Triggers Starvation Response with Paradoxical Induction of TXNIP through MondoA. PLOS Genet. 2010, 6, e1001093. [Google Scholar] [CrossRef]
- Riemann, A.; Ihling, A.; Schneider, B.; Gekle, M.; Thews, O. Impact of Extracellular Acidosis on Intracellular PH Control and Cell Signaling in Tumor Cells. Adv. Exp. Med. Biol. 2013, 789, 221–228. [Google Scholar] [CrossRef]
- Koul, H.K.; Pal, M.; Koul, S. Role of P38 MAP Kinase Signal Transduction in Solid Tumors. Genet. Cancer 2013, 4, 342–359. [Google Scholar] [CrossRef] [PubMed]
- Avnet, S.; Lemma, S.; Cortini, M.; Pellegrini, P.; Perut, F.; Zini, N.; Kusuzaki, K.; Chano, T.; Grisendi, G.; Dominici, M.; et al. Altered PH Gradient at the Plasma Membrane of Osteosarcoma Cells Is a Key Mechanism of Drug Resistance. Oncotarget 2016, 7, 63408–63423. [Google Scholar] [CrossRef] [PubMed]
- Moliterno, P.; Donangelo, C.M.; Borgarello, L.; Pécora, M.; Olascoaga, A.; Noboa, O.; Boggia, J. Association of dietary patterns with cardiovascular and kidney phenotypes in an Uruguayan population cohort. Nutrients 2021, 13, 2213. [Google Scholar] [CrossRef] [PubMed]
- Storz, M.A.; Ronco, A.L. Quantifying Dietary Acid Load in U.S. Cancer Survivors: An Exploratory Study Using NHANES Data. BMC Nutr. 2022, 8, 43. [Google Scholar] [CrossRef]
- Wu, T.; Hsu, F.-C.; Pierce, J.P. Increased Acid-Producing Diet and Past Smoking Intensity Are Associated with Worse Prognoses Among Breast Cancer Survivors: A Prospective Cohort Study. J. Clin. Med. 2020, 9, 1817. [Google Scholar] [CrossRef]
- Chronister, B.N.C.; Wu, T.; Santella, R.M.; Neugut, A.I.; Wolff, M.S.; Chen, J.; Teitelbaum, S.L.; Parada, H. Dietary Acid Load, Serum Polychlorinated Biphenyl Levels, and Mortality Following Breast Cancer in the Long Island Breast Cancer Study Project. Int. J. Environ. Res. Public Health 2021, 19, 374. [Google Scholar] [CrossRef]
- Tessou, K.D.; Lemus, H.; Hsu, F.-C.; Pierce, J.; Hong, S.; Brown, L.; Wu, T. Independent and Joint Impacts of Acid-Producing Diets and Depression on Physical Health among Breast Cancer Survivors. Nutrients 2021, 13, 2422. [Google Scholar] [CrossRef]
- Wu, T.; Hsu, F.-C.; Pierce, J.P. Acid-Producing Diet and Depressive Symptoms among Breast Cancer Survivors: A Longitudinal Study. Cancers 2020, 12, 3183. [Google Scholar] [CrossRef]
- McAndrew, N.P.; Bottalico, L.; Mesaros, C.; Blair, I.A.; Tsao, P.Y.; Rosado, J.M.; Ganguly, T.; Song, S.J.; Gimotty, P.A.; Mao, J.J.; et al. Effects of Systemic Inflammation on Relapse in Early Breast Cancer. NPJ Breast Cancer 2021, 7, 7. [Google Scholar] [CrossRef]
- Ayoub, N.M.; Jaradat, S.K.; Alhusban, A.; Tahaineh, L. Glycosylated Hemoglobin A1c Is Associated with Anthropometric Measurements and Tumor Characteristics in Breast Cancer Patients. Int. J. Women’s Health 2020, 12, 139–149. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Ref. | Tumor Site | Cases/ Controls | Cat. 1 | DAL Score | Highest OR (95% CI) | Trend (p) |
|---|---|---|---|---|---|---|
| [45] | Oro-pharynx-larynx | 563/563 | 3 | PRAL | 2.10(1.46–3.03) | <0.001 |
| 3 | NEAP | 1.95 (1.37–2.78) | 0.002 | |||
| [45] | Oral cavity | 103/563 | 3 | PRAL | 1.29 (0.69–2.43) | 0.49 |
| 3 | NEAP | 1.06 (0.56–1.98) | 0.98 | |||
| [45] | Pharynx | 185/563 | 3 | PRAL | 2.40 (1.44–4.01) | 0.005 |
| 3 | NEAP | 2.28 (1.40–3.71) | 0.006 | |||
| [45] | Larynx | 275/563 | 3 | PRAL | 2.22 (1.42–3.47) | <0.001 |
| 3 | NEAP | 2.00 (1.29–3.09) | 0.006 | |||
| [40] | Lung | 843/1466 | 4 | PRAL | 0.99 (0.64–1.52) | 0.94 |
| 4 | NEAP | 2.22 (1.52–3.22) | <0.001 | |||
| [46] | Esophagus | 185/1110 | 3 | PRAL | 2.28 (1.44–3.61) | <0.001 |
| 3 | NEAP | 2.17 (1.38–3.41) | <0.001 | |||
| [47] | Stomach | 274/1096 | 3 | PRAL | 1.74 (1.13–2.66) | <0.001 |
| 3 | NEAP | 1.90 (1.26–2.84) | <0.001 | |||
| [39] | Colorectum | 611/2394 | 4 | PRAL | 1.53 (1.02–2.31) | 0.03 |
| 4 | NEAP | 1.29 (0.89–1.88) | 0.11 | |||
| [39] | Colon | 319/2394 | 4 | PRAL | 1.29 (0.76–2.19) | 0.052 |
| 4 | NEAP | 1.37 (0.85–2.24) | 0.20 | |||
| [39] | Rectum | 292/2394 | 4 | PRAL | 1.77 (1.00–3.12) | 0.048 |
| 4 | NEAP | 1.22 (0.73–2.04) | 0.24 | |||
| [41] | Breast | 572/889 | 4 | PRAL | 2.46 (1.76–3.44) | <0.001 |
| 4 | NEAP | 1.78 (1.30–2.42) | <0.001 | |||
| [43] | Kidney | 114/864 | 4 | PRAL | 0.98 (0.94–1.02) | 0.34 |
| 4 | NEAP | 1.00 (0.98–1.02) | 0.92 | |||
| [44] | Bladder | 255/510 | 3 | PRAL | 1.74 (1.08–2.82) | 0.002 |
| 3 | NEAP | 1.83 (1.15–2.89) | 0.02 | |||
| [42] | Prostate | 323/969 | 3 | PRAL | 1.52 (1.01–2.28) | 0.01 |
| 3 | NEAP | 1.72 (1.16–2.48) | 0.048 | |||
| [48] | Whole sample | 3736/9534 | 5 | PRAL | 2.20 (1.91–2.54) | <0.001 |
| 5 | NEAP | 2.35 (2.04–2.70) | <0.001 |
| Variables | Categories | PRAL Score: Controls (mEq/d ± SD) | PRAL Score: Cases (mEq/d ± SD) | p-Value for Difference |
|---|---|---|---|---|
| Age groups | <50 years | 5.0 ± 10.2 | 6.9 ± 10.9 | 0.001 |
| 50–59 years | 4.0 ± 9.0 | 7.4 ± 11.6 | <0.0001 | |
| 60–69 years | 3.3 ± 11.0 | 6.5 ± 11.2 | <0.0001 | |
| 70–79 years | 2.2 ± 10.5 | 5.4 ± 11.3 | <0.0001 | |
| 80–89 years | 0.4 ± 12.3 | 5.2 ± 11.2 | <0.0001 | |
| Urban/rural status | Urban | 2.9 ± 10.6 | 5.9 ± 11.1 | <0.0001 |
| Rural | 3.9 ± 10.3 | 8.3 ± 11.7 | <0.0001 | |
| Residence region | Montevideo | 3.2 ± 10.5 | 5.0 ± 10.9 | <0.0001 |
| Other counties | 3.0 ± 10.7 | 7.6 ± 11.5 | <0.0001 | |
| Sex | Men | 3.7 ± 10.7 | 7.3 ± 11.4 | <0.0001 |
| Women | 1.7 ± 9.9 | 3.7 ± 10.7 | <0.0001 | |
| Education years | <5 | 3.4 ± 10.6 | 6.8 ± 11.1 | <0.0001 |
| ≥5 | 2.7 ± 10.5 | 5.9 ± 11.6 | <0.0001 | |
| Body mass index (kg/m2) | <18.50 | 2.1 ± 11.7 | 8.7 ± 11.1 | 0.0002 |
| 18.50–24.99 | 3.6 ± 10.7 | 6.5 ± 11.1 | <0.0001 | |
| 25.00–29.99 | 2.7 ± 10.4 | 6.1 ± 11.5 | <0.0001 | |
| ≥30.00 | 2.9 ± 10.3 | 6.2 ± 11.7 | <0.0001 | |
| Family history of cancer for first- and second-degree relatives | No | 3.2 ± 10.4 | 6.6 ± 11.1 | <0.0001 |
| Yes | 2.9 ± 11.0 | 5.9 ± 11.7 | <0.0001 | |
| Smoking intensity (pack-years) | Non-smoker | 2.0 ± 10.3 | 3.3 ± 11.3 | 0.0004 |
| 0.01–27.0 | 2.8 ± 10.8 | 5.8 ± 11.3 | <0.0001 | |
| 27.1–53.0 | 4.5 ± 10.0 | 7.2 ± 11.0 | <0.0001 | |
| ≥53.1 | 3.8 ± 11.0 | 8.4 ± 10.9 | <0.0001 | |
| Alcohol drinking | Non-drinker | 1.8 ± 10.3 | 4.3 ± 11.5 | <0.0001 |
| Ex-drinker | 0.9 ± 10.8 | 7.1 ± 12.0 | <0.0001 | |
| Current drinker | 4.9 ± 10.5 | 7.9 ± 10.6 | <0.0001 | |
| “Mate’’ status | Non-drinker | 1.5 ± 10.1 | 3.9 ± 12.2 | 0.0001 |
| Ever drinker | 3.3 ± 10.6 | 6.7 ± 11.1 | <0.0001 | |
| “Mate’’ intensity (liter-years) | Non-drinker | 1.5 ± 10.1 | 3.9 ± 12.2 | 0.0001 |
| 0.1–39 | 2.5 ± 11.0 | 5.8 ± 11.0 | <0.0001 | |
| 39.1–62.9 | 3.8 ± 10.3 | 6.4 ± 10.9 | <0.0001 | |
| ≥63.0 | 3.7 ± 10.4 | 7.7 ± 11.5 | <0.0001 | |
| Whole sample | 3.1 ± 10.6 | 6.4 ± 11.3 | <0.0001 |
| Men | Women | Men | Women | |
|---|---|---|---|---|
| Tumor Site | PRAL ± SD | PRAL ± SD | NEAP ± SD | NEAP ± SD |
| Oral cavity | 7.8 ± 10.8 | ----- | 55.9 ± 19.0 | ----- |
| Pharynx | 9.7 ± 11.9 | ----- | 59.6 ± 16.8 | ----- |
| Larynx | 10.4 ± 11.1 | ----- | 60.1 ± 18.0 | ----- |
| Lung | 7.5 ± 10.7 | ----- | 59.3 ± 17.7 | ----- |
| Esophagus | 9.5 ± 10.7 | ----- | 60.1 ± 16.8 | ----- |
| Stomach | 7.6 ± 11.8 | 1.9 ± 10.6 | 57.3 ± 19.2 | 48.9 ± 14.5 |
| Colon | 6.2 ± 11.6 | 2.9 ± 12.4 | 54.9 ± 17.2 | 51.6 ± 16.6 |
| Rectum | 7.1 ± 12.5 | 1.1 ± 12.4 | 56.3 ± 16.8 | 50.6 ± 18.3 |
| Breast | ----- | 5.1 ± 10.1 | ----- | 59.3 ± 18.6 |
| Kidney | 4.9 ± 10.2 | −2.0 ± 8.5 | 55.5 ± 15.5 | 43.3 ± 16.2 |
| Bladder | 5.6 ± 10.6 | 2.7 ± 10.8 | 56.5 ± 17.5 | 50.8 ± 11.5 |
| Prostate | 3.6 ± 11.7 | ----- | 52.5 ± 16.5 | ----- |
| All cases | 7.3 ± 11.4 | 3.7 ± 10.7 | 57.6 ± 17.6 | 55.5 ± 18.3 |
| Controls | 3.5 ± 10.7 | 1.7 ± 9.9 | 52.4 ± 17.9 | 50.3 ± 16.0 |
| Whole sample | 4.6 ± 11.0 | 2.3 ± 10.2 | 53.8 ± 18.0 | 51.8 ± 16.9 |
| p-value * | <0.001 | <0.001 | <0.001 | <0.001 |
| Subgroup | Units | Controls | Cases | All | p-Value for Difference |
|---|---|---|---|---|---|
| Men | mg/1000 kcal/day | 1724 ± 395 | 1778 ± 388 | 1739 ± 394 | <0.0001 |
| Women | mg/1000 kcal/day | 1784 ± 465 | 1883 ± 470 | 1813 ± 469 | <0.0001 |
| All | mg/1000 kcal/day | 1739 ± 414 | 1805 ± 413 | 1758 ± 415 | <0.0001 |
| p-value for difference | <0.0001 | <0.0001 | <0.0001 | ||
| Men | mg/kg weight/day | 38.2 ± 15.4 | 42.7 ± 16.5 | 39.5 ± 15.8 | <0.0001 |
| Women | mg/kg weight/day | 37.6 ± 16.3 | 43.0 ± 19.3 | 39.2 ± 17.4 | <0.0001 |
| All | mg/kg weight/day | 38.1 ± 15.6 | 42.8 ± 17.3 | 39.4 ± 16.2 | <0.0001 |
| p-value for difference | 0.11 | 0.64 | 0.44 |
| Methionine Intake (g/Day) | All Women (%) | Female Controls (%) | All Female Cancer Cases (%) | Breast Cancer Cases (%) | Other Female Cancer Cases (%) |
|---|---|---|---|---|---|
| <2.05 | 1031 (31.1) | 795 (33.4) | 236 (24.2) | 124 (21.7) | 112 (27.9) |
| 2.05–2.66 | 836 (25.2) | 606 (25.9) | 230 (23.6) | 121 (21.1) | 109 (27.2) |
| 2.67–3.32 | 745 (22.5) | 520 (22.2) | 225 (23.1) | 127 (22.2) | 98 (24.4) |
| >3.32 | 700 (21.1) | 418 (17.9) | 282 (29.0) | 200 (35.0) | 82 (20.5) |
| Total numbers | 3312 (100.0) | 2339 (100.0) | 973 (100.0) | 572 (100.0) | 401 (100.0) |
| Food | % Methionine in Protein | mg Methionine/kcal |
|---|---|---|
| Almonds | 1.15 | 0.32 |
| Black beans | 1.51 | 1.01 |
| Brown rice | 2.26 | 0.52 |
| Chicken breast without skin | 2.77 | 4.94 |
| Corn | 2.11 | 0.64 |
| Eggs | 3.11 | 2.54 |
| Lean ground beef | 2.34 | 2.05 |
| Lentils | 0.85 | 0.66 |
| Low-fat milk | 2.51 | 1.97 |
| Oatmeal | 1.87 | 0.77 |
| Peanuts | 1.23 | 0.49 |
| Pinto beans | 1.51 | 0.90 |
| Potato | 1.61 | 0.42 |
| Soy (tofu) | 1.28 | 1.36 |
| Tuna | 2.96 | 6.48 |
| Wheat (bulgur) | 1.55 | 0.57 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ronco, A.L.; Storz, M.A. Dietary Acid Load and Cancer Risk: A Review of the Uruguayan Experience. Nutrients 2023, 15, 3098. https://doi.org/10.3390/nu15143098
Ronco AL, Storz MA. Dietary Acid Load and Cancer Risk: A Review of the Uruguayan Experience. Nutrients. 2023; 15(14):3098. https://doi.org/10.3390/nu15143098
Chicago/Turabian StyleRonco, Alvaro Luis, and Maximilian Andreas Storz. 2023. "Dietary Acid Load and Cancer Risk: A Review of the Uruguayan Experience" Nutrients 15, no. 14: 3098. https://doi.org/10.3390/nu15143098
APA StyleRonco, A. L., & Storz, M. A. (2023). Dietary Acid Load and Cancer Risk: A Review of the Uruguayan Experience. Nutrients, 15(14), 3098. https://doi.org/10.3390/nu15143098