

A Scoping Review on the Effects of Carotenoids and Flavonoids on Skin Damage Due to Ultraviolet Radiation


 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Literature Review
2.2. Selection of Research Article
2.3. Data Extraction and Management
3. Results
3.1. Search Results
3.2. Study Characteristic
3.3. Findings from In Vitro Studies
3.4. Findings from In Vivo Studies
3.5. Findings from Human Studies
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Panich, U.; Sittithumcharee, G.; Rathviboon, N.; Jirawatnotai, S. Ultraviolet Radiation-Induced Skin Aging: The Role of DNA Damage and Oxidative Stress in Epidermal Stem Cell Damage Mediated Skin Aging. Stem Cells Int. 2016, 2016, 7370642. [Google Scholar] [CrossRef] [PubMed]
- Guan, L.L.; Lim, H.W.; Mohammad, T.F. Sunscreens and Photoaging: A Review of Current Literature. Am. J. Clin. Dermatol. 2021, 22, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxidative Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef] [PubMed]
- Kotsafti, A.; Scarpa, M.; Castagliuolo, I.; Scarpa, M. Reactive Oxygen Species and Antitumor Immunity—From Surveillance to Evasion. Cancers 2020, 12, 1748. [Google Scholar] [CrossRef] [PubMed]
- Nita, M.; Grzybowski, A. The Role of the Reactive Oxygen Species and Oxidative Stress in the Pathomechanism of the Age-Related Ocular Diseases and Other Pathologies of the Anterior and Posterior Eye Segments in Adults. Oxidative Med. Cell. Longev. 2016, 2016, 3164734. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.V.; Mistry, B.M.; Shinde, S.K.; Syed, R.; Singh, V.; Shin, H.S. Therapeutic Potential of Quercetin as a Cardiovascular Agent. Eur. J. Med. Chem. 2018, 155, 889–904. [Google Scholar] [CrossRef]
- Dayem, A.A.; Choi, H.Y.; Yang, G.; Kim, K.; Saha, S.K.; Cho, S. The Anti-Cancer Effect of Polyphenols against Breast Cancer and Cancer Stem Cells: Molecular Mechanisms. Nutrients 2016, 8, 581. [Google Scholar] [CrossRef]
- Nahar, L.; Xiao, J.; Sarker, S.D. Introduction of Phytonutrients. In Handbook of Dietary Phytochemicals; Xiao, J., Sarker, S.D., Asakawa, Y., Eds.; Springer: Singapore, 2020; pp. 1–17. [Google Scholar] [CrossRef]
- Mintie, C.A.; Singh, C.K.; Ahmad, N. Whole Fruit Phytochemicals Combating Skin Damage and Carcinogenesis. Transl. Oncol. 2020, 13, 146–156. [Google Scholar] [CrossRef]
- Ibrahim, N.I.; Wong, S.K.; Mohamed, I.N.; Mohamed, N.; Chin, K.; Ima-Nirwana, S.; Shuid, A.N. Wound Healing Properties of Selected Natural Products. Int. J. Environ. Res. Public Health 2018, 15, 2360. [Google Scholar] [CrossRef]
- de Andrade Monteiro, C.; Santos, J.R.A.d. Phytochemicals and Their Antifungal Potential against Pathogenic Yeasts. In Phytochemicals in Human Health; Rao, V., Mans, D.R., Rao, L., Eds.; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef]
- Habib, S.H.; Makpol, S.; Hamid, N.A.; Das, S.; Ngah, W.Z.; Yusof, Y.A. Ginger Extract (Zingiber Officinale) Has Anti-Cancer and Anti-Inflammatory Effects on Ethionine-Induced Hepatoma Rats. Clinics 2008, 63, 807–813. [Google Scholar] [CrossRef]
- Mlcek, J.; Jurikova, T.; Skrovankova, S.; Sochor, J. Quercetin and Its Anti-Allergic Immune Response. Molecules 2016, 21, 623. [Google Scholar] [CrossRef] [PubMed]
- Lanuzza, F.; Occhiuto, F.; Monforte, M.T.; Tripodo, M.M.; D’Angelo, V.; Galati, E.M. Antioxidant Phytochemicals of Opuntia Ficus-Indica (L.) Mill. Cladodes with Potential Anti-Spasmodic Activity. Pharmacogn. Mag. 2017, 13, 424. [Google Scholar] [CrossRef]
- Yuliani, S.; Mustofa; Partadiredja, G. Turmeric (Curcuma Longa L.) Extract May Prevent the Deterioration of Spatial Memory and the Deficit of Estimated Total Number of Hippocampal Pyramidal Cells of Trimethyltin-Exposed Rats. Drug Chem. Toxicol. 2018, 41, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Bunaim, M.K.; Kamisah, Y.; Mohd Mustazil, M.N.; Fadhlullah Zuhair, J.S.; Juliana, A.H.; Muhammad, N. Centella Asiatica (L.) Urb. Prevents Hypertension and Protects the Heart in Chronic Nitric Oxide Deficiency Rat Model. Front. Pharmacol. 2021, 12, 3311. [Google Scholar] [CrossRef] [PubMed]
- Niaz, K.; Shah, M.A.; Khan, F.; Saleem, U.; Vargas, C.; Panichayupakaranant, P. Bioavailability and Safety of Phytonutrients. In Phytonutrients in Food; Nabavi, S.M., Suntar, I., Barreca, D., Khan, H., Eds.; Woodhead Publishing: Cambridge, UK, 2020; pp. 117–136. [Google Scholar] [CrossRef]
- Kamaruddin, N.A.; Hakim Abdullah, M.N.; Tan, J.J.; Lim, V.; Fong, L.Y.; Abd Ghafar, S.A.; Yong, Y.K. Vascular Protective Effect and Its Possible Mechanism of Action on Selected Active Phytocompounds: A Review. Evid.-Based Complement. Altern. Med. 2022, 2022, 3311228. [Google Scholar] [CrossRef] [PubMed]
- Vayalil, P.K.; Mittal, A.; Hara, Y.; Elmets, C.A.; Katiyar, S.K. Green Tea Polyphenols Prevent Ultraviolet Light-Induced Oxidative Damage and Matrix Metalloproteinases Expression in Mouse Skin. J. Investig. Dermatol. 2004, 122, 1480–1487. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, U.; Moore, C.E.; De Spirt, S.; Tronnier, H.; Stahl, W. Green Tea Polyphenols Provide Photoprotection, Increase Microcirculation, and Modulate Skin Properties of Women. J. Nutr. 2011, 141, 1202–1208. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.Y.; Lim, S.S.; Kim, S.J.; Choi, J.S.; Park, J.; Ju, S.M.; Han, S.J.; Kang, I.J.; Kang, Y.H. Bog Blueberry Anthocyanins Alleviate Photoaging in Ultraviolet-B Irradiation-Induced Human Dermal Fibroblasts. Mol. Nutr. Food Res. 2009, 53, 726–738. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Sánchez, A.; Barrajón-Catalán, E.; Caturla, N.; Castillo, J.; Benavente-García, O.; Alcaraz, M.; Micol, V. Protective Effects of Citrus and Rosemary Extracts on UV-Induced Damage in Skin Cell Model and Human Volunteers. J. Photochem. Photobiol. B Biol. 2014, 136, 12–18. [Google Scholar] [CrossRef]
- Lim, S.H.; Jung, S.K.; Byun, S.; Lee, E.J.; Hwang, J.A.; Seo, S.G.; Kim, Y.A.; Yu, J.G.; Lee, K.W.; Lee, H.J. Luteolin Suppresses UVB-Induced Photoageing by Targeting JNK1 and P90RSK2. J. Cell. Mol. Med. 2013, 17, 672–680. [Google Scholar] [CrossRef]
- Groten, K.; Marini, A.; Grether-Beck, S.; Jaenicke, T.; Ibbotson, S.H.; Moseley, H.; Ferguson, J.; Krutmann, J. Tomato Phytonutrients Balance UV Response: Results from a Double-Blind, Randomized, Placebo-Controlled Study. Ski. Pharmacol. Physiol. 2019, 32, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Rizwan, M.; Rodriguez-Blanco, I.; Harbottle, A.; Birch-Machin, M.A.; Watson, R.E.B.; Rhodes, L.E. Tomato Paste Rich in Lycopene Protects against Cutaneous Photodamage in Humans in Vivo: A Randomized Controlled Trial. Br. J. Dermatol. 2011, 164, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Oh, J.; Averilla, J.N.; Kim, H.J.; Kim, J.S.; Kim, J.S. Grape Peel Extract and Resveratrol Inhibit Wrinkle Formation in Mice Model through Activation of Nrf2/HO-1 Signaling Pathway. J. Food Sci. 2019, 84, 1600–1608. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.Y.; Liu, W.; Hao, J.C.; Gu, W.J.; Zhao, Y.S. Topical Grape Seed Proanthocyandin Extract Reduces Sunburn Cells and Mutant P53 Positive Epidermal Cell Formation, and Prevents Depletion of Langerhans Cells in an Acute Sunburn Model. Photomed. Laser Surg. 2012, 30, 20–25. [Google Scholar] [CrossRef]
- Parmar, D.; Shah, J. Role of Ultraviolet Radiation in Skin Cancer: A Review Article. J. Adv. Med. Dent. Sci. Res. 2015, 3, 83–85. [Google Scholar]
- Gilaberte, Y.; Prieto-Torres, L.; Pastushenko, I.; Juarranz, Á. Anatomy and Function of the Skin. In Nanoscience in Dermatology; Hamblin, M.R., Avci, P., Prow, T.W., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 1–14. [Google Scholar] [CrossRef]
- Solano, F. Photoprotection and Skin Pigmentation: Melanin-Related Molecules and Some Other New Agents Obtained from Natural Sources. Molecules 2020, 25, 1537. [Google Scholar] [CrossRef]
- Herndon, J.M.; Hoisington, R.D.; Whiteside, M. Deadly Ultraviolet UV-C and UV-B Penetration to Earth’s Surface: Human and Environmental Health Implications. J. Geogr. Environ. Earth Sci. Int. 2018, 14, 1–11. [Google Scholar] [CrossRef]
- D’Orazio, J.; Jarrett, S.; Amaro-Ortiz, A.; Scott, T. UV Radiation and the Skin. Int. J. Mol. Sci. 2013, 14, 12222–12248. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.W.; Hung, Y.C.; Lin, T.Y.; Fang, J.Y.; Yang, P.M.; Chen, M.H.; Pan, T.L. Comparison of the Biological Impact of UVA and UVB upon the Skin with Functional Proteomics and Immunohistochemistry. Antioxidants 2019, 8, 569. [Google Scholar] [CrossRef]
- Valejo Coelho, M.M.; Matos, T.R.; Apetato, M. The Dark Side of the Light: Mechanisms of Photocarcinogenesis. Clin. Dermatol. 2016, 34, 563–570. [Google Scholar] [CrossRef]
- Ghazali, N.I.; Mohd Rais, R.Z.; Makpol, S.; Chin, K.Y.; Yap, W.N.; Goon, J.A. Effects of Tocotrienol on Aging Skin: A Systematic Review. Front. Pharmacol. 2022, 13, 1006198. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.; Kim, S.; Shin, D.H.; Kim, M.S. Inhibition of SARS-CoV 3CL Protease by Flavonoids. J. Enzym. Inhib. Med. Chem. 2020, 35, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Kordmahalleh, M.M.; Sefidmazgi, M.G.; Harrison, S.H.; Homaifar, A. Identifying Time-Delayed Gene Regulatory Networks via an Evolvable Hierarchical Recurrent Neural Network. BioData Min. 2017, 10, 29. [Google Scholar] [CrossRef] [PubMed]
- Ahsanuddin, S.; Lam, M.; Baron, E.D. Skin Aging and Oxidative Stress. AIMS Mol. Sci. 2016, 3, 187–195. [Google Scholar] [CrossRef]
- Bosch, R.; Philips, N.; Suárez-Pérez, J.; Juarranz, A.J.; Devmurari, A.; Chalensouk-Khaosaat, J.; González, S. Mechanisms of Photoaging and Cutaneous Photocarcinogenesis, and Photoprotective Strategies with Phytochemicals. Antioxidants 2015, 4, 248–268. [Google Scholar] [CrossRef]
- Domínguez Díaz, L.; Fernández-Ruiz, V.; Cámara, M. The Frontier between Nutrition and Pharma: The International Regulatory Framework of Functional Foods, Food Supplements and Nutraceuticals. Crit. Rev. Food Sci. Nutr. 2020, 60, 1738–1746. [Google Scholar] [CrossRef]
- Martín, J.; Navas, M.J.; Jiménez-Moreno, A.M.; Asuero, A.G. Anthocyanin Pigments: Importance, Sample Preparation and Extraction. In Phenolic Compounds—Natural Sources, Importance and Applications; Soto-Hernandez, M., Palma-Tenango, M., Garcia-Mateos, R., Eds.; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef]
- Domaszewska-Szostek, A.; Puzianowska-Kuźnicka, M.; Kuryłowicz, A. Flavonoids in Skin Senescence Prevention and Treatment. Int. J. Mol. Sci. 2021, 22, 6814. [Google Scholar] [CrossRef]
- Yusuf, N.; Irby, C.; Katiyar, S.K.; Elmets, C.A. Photoprotective Effects of Green Tea Polyphenols. Photodermatol. Photoimmunol. Photomed. 2007, 23, 48–56. [Google Scholar] [CrossRef]
- Kim, E.; Hwang, K.; Lee, J.; Han, S.Y.; Kim, E.; Park, J.; Cho, J.Y. Skin Protective Effect of Epigallocatechin Gallate. Int. J. Mol. Sci. 2018, 19, 173. [Google Scholar] [CrossRef]
- Franceschi, C.; Capri, M.; Monti, D.; Giunta, S.; Olivieri, F.; Sevini, F.; Panourgia, M.P.; Invidia, L.; Celani, L.; Scurti, M.; et al. Inflammaging and Anti-Inflammaging: A Systemic Perspective on Aging and Longevity Emerged from Studies in Humans. Mech. Ageing Dev. 2007, 128, 92–105. [Google Scholar] [CrossRef]
- Wang, J.; Bian, Y.; Cheng, Y.; Sun, R.; Li, G. Effect of Lemon Peel Flavonoids on UVB-Induced Skin Damage in Mice. RSC Adv. 2020, 10, 31470–31478. [Google Scholar] [CrossRef] [PubMed]
- Neha; Jaggi, A.S.; Singh, N. Silymarin and Its Role in Chronic Diseases. In Advances in Experimental Medicine and Biology; Gupta, S., Prasad, S., Aggarwal, B., Eds.; Springer: Cham, Switzerland, 2016; pp. 25–44. [Google Scholar] [CrossRef]
- Carrascosa, J.M.; Floriach, N.; Sala, E.; Aguilera, J. Increase in Minimal Erythemal Dose Following Oral Administration of an Antioxidant Complex Based on a Mix of Carotenoids: Double-blind, Placebo-controlled Trial. Photodermatol. Photoimmunol. Photomed. 2017, 33, 284–286. [Google Scholar] [CrossRef] [PubMed]
- Calniquer, G.; Khanin, M.; Ovadia, H.; Linnewiel-Hermoni, K.; Stepensky, D.; Trachtenberg, A.; Sedlov, T.; Braverman, O.; Levy, J.; Sharoni, Y. Combined Effects of Carotenoids and Polyphenols in Balancing the Response of Skin Cells to UV Irradiation. Molecules 2021, 26, 1931. [Google Scholar] [CrossRef]
- Zerres, S.; Stahl, W. Carotenoids in Human Skin. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2020, 1865, 158588. [Google Scholar] [CrossRef] [PubMed]
- Baswan, S.M.; Klosner, A.E.; Weir, C.; Salter-Venzon, D.; Gellenbeck, K.W.; Leverett, J.; Krutmann, J. Role of Ingestible Carotenoids in Skin Protection: A Review of Clinical Evidence. Photodermatol. Photoimmunol. Photomed. 2021, 37, 490–504. [Google Scholar] [CrossRef]
- Puah, B.P.; Jalil, J.; Attiq, A.; Kamisah, Y. New Insights into Molecular Mechanism behind Anti-Cancer Activities of Lycopene. Molecules 2021, 26, 3888. [Google Scholar] [CrossRef]
- Stahl, W.; Heinrich, U.; Wiseman, S.; Eichler, O.; Sies, H.; Tronnier, H. Dietary tomato paste protects against ultraviolet light-induced erythema in humans. J. Nutr. 2001, 131, 1449–1451. [Google Scholar] [CrossRef]
- Crascì, L.; Lauro, M.R.; Puglisi, G.; Panico, A. Natural Antioxidant Polyphenols on Inflammation Management: Anti-Glycation Activity vs. Metalloproteinases Inhibition. Crit. Rev. Food Sci. Nutr. 2017, 58, 893–904. [Google Scholar] [CrossRef]
- Sharma, P.; Montes de Oca, M.K.; Alkeswani, A.R.; McClees, S.F.; Das, T.; Elmets, C.A.; Afaq, F. Tea Polyphenols for the Prevention of UVB-Induced Skin Cancer. Photodermatol. Photoimmunol. Photomed. 2017, 34, 50–59. [Google Scholar] [CrossRef]
- Lee, S.A.; Lim, W.H.; Van Le, V.; Ko, S.R.; Kim, B.; Oh, H.M.; Ahn, C.Y. Lifespan Extension and Anti-Oxidant Effects of Carotenoid Pigments in Caenorhabditis Elegans. Bioresour. Technol. Rep. 2022, 17, 100962. [Google Scholar] [CrossRef]
- Trivedi, P.P.; Jena, G.B. Mechanistic Insight into Beta-Carotene-Mediated Protection against Ulcerative Colitis-Associated Local and Systemic Damage in Mice. Eur. J. Nutr. 2014, 54, 639–652. [Google Scholar] [CrossRef] [PubMed]
- Kanaze, F.I.; Bounartzi, M.I.; Georgarakis, M.; Niopas, I. Pharmacokinetics of the citrus flavanone aglycones hesperetin and naringenin after single oral administration in human subjects. Eur. J. Clin. Nutr. 2007, 61, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Gardana, C.; Guarnieri, S.; Riso, P.; Simonetti, P.; Porrini, M. Flavanone plasma pharmacokinetics from blood orange juice in human subjects. Br. J. Nutr. 2007, 98, 165–172. [Google Scholar] [CrossRef]
- Miyake, Y.; Mochizuki, M.; Okada, M.; Hiramitsu, M.; Morimitsu, Y.; Osawa, T. Isolation of antioxidative phenolic glucosides from lemon juice and their suppressive effect on the expression of blood adhesion molecules. Biosci. Biotechnol. Biochem. 2007, 71, 1911–1919. [Google Scholar] [CrossRef] [PubMed]
- Riva, A.; Ronchi, M.; Petrangolini, G.; Bosisio, S.; Allegrini, P. Improved oral absorption of quercetin from quercetin phytosome®, a new delivery system based on food grade lecithin. Eur. J. Drug Metab. Pharmacokinet. 2019, 44, 169–177. [Google Scholar] [CrossRef]
- Wang, S.T.; Fang, T.F.; Hsu, C.; Chen, C.H.; Lin, C.J.; Su, N.W. Biotransformed product, genistein 7-O-phosphate, enhances the oral bioavailability of genistein. J. Funct. Foods 2015, 13, 323–335. [Google Scholar] [CrossRef]
- Brett, G.M.; Hollands, W.; Needs, P.W.; Teucher, B.; Dainty, J.R.; Davis, B.D.; Brodbelt, J.S.; Kroon, P.A. Absorption, metabolism and excretion of flavanones from single portions of orange fruit and juice and effects of anthropometric variables and contraceptive pill use on flavanone excretion. Br. J. Nutr. 2009, 101, 664–675. [Google Scholar] [CrossRef]
- Lin, L.C.; Pai, Y.F.; Tsai, T.H. Isolation of luteolin and luteolin-7-O-glucoside from Dendranthema morifolium Ramat Tzvel and their pharmacokinetics in rats. J. Agric. Food Chem. 2015, 63, 7700–7706. [Google Scholar] [CrossRef]
- Auger, C.; Mullen, W.; Hara, Y.; Crozier, A. Bioavailability of polyphenon E flavan-3-ols in humans with an ileostomy. J. Nutr. 2008, 138, 1535S–1542S. [Google Scholar] [CrossRef]
- Chang, C.W.; Wong, C.Y.; Wu, Y.T.; Hsu, M.C. Development of a solid dispersion system for improving the oral bioavailability of resveratrol in rats. Eur. J. Drug Metab. Pharmacokinet. 2017, 42, 239–249. [Google Scholar] [CrossRef]
- Williamson, G.; Kay, C.D.; Crozier, A. The bioavailability, transport, and bioactivity of dietary flavonoids: A review from a historical perspective. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1054–1112. [Google Scholar] [CrossRef] [PubMed]
- Khan, J.; Deb, P.K.; Priya, S.; Medina, K.D.; Devi, R.; Walode, S.G.; Rudrapal, M. Dietary Flavonoids: Cardioprotective Potential with Antioxidant Effects and Their Pharmacokinetic, Toxicological and Therapeutic Concerns. Molecules 2021, 26, 4021. [Google Scholar] [CrossRef] [PubMed]
- Kiokias, S.; Proestos, C.; Varzakas, T. A Review of the Structure, Biosynthesis, Absorption of Carotenoids-Analysis and Properties of their Common Natural Extracts. Curr. Res. Nutr. Food Sci. 2016, 4, 25–37. [Google Scholar] [CrossRef]
- Berni, P.; Chitchumroonchokchai, C.; Canniatti-Brazaca, G.S.; De Moura, F.F.; Failla, L.M. Comparison of Content and In vitro Bioaccessibility of Provitamin A Carotenoids in Home Cooked and Commercially Processed orange Fleshed Sweet Potato (Ipomea batatas Lam). Plant Foods Hum. Nutr. 2015, 70, 1–8. [Google Scholar] [CrossRef] [PubMed]
- van Het Hof, K.H.; West, C.E.; Weststrate, J.A.; Hautvast, J.G. Dietary factors that affect the bioavailability of carotenoids. J. Nutr. 2000, 130, 503–506. [Google Scholar] [CrossRef]
- Castenmiller, J.J.M.; West, C.E.; Linssen, J.P.H.; van het Hof, K.H.; Voragen, A.G.J. The Food Matrix of Spinach Is a Limiting Factor in Determining the Bioavailability of β-Carotene and to a Lesser Extent of Lutein in Humans. J. Nutr. 1999, 129, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Birt, D.F.; Jeffery, E. Flavonoids. Adv. Nutr. 2013, 4, 576–577. [Google Scholar] [CrossRef]
- Hammond, B.R., Jr.; Renzi, L.M. Carotenoids. Adv. Nutr. 2013, 4, 474–476. [Google Scholar] [CrossRef]
- Druesne Pecollo, N.; Latino-Martel, P.; Norat, T.; Barrandon, E.; Bertrais, S.; Galan, P.; Hercberg, S. Beta-carotene supplementation and cancer risk: A systematic review and meta-analysis of randomized controlled trials. Int. J. Cancer 2010, 127, 172–184. [Google Scholar] [CrossRef]



{kind=link}
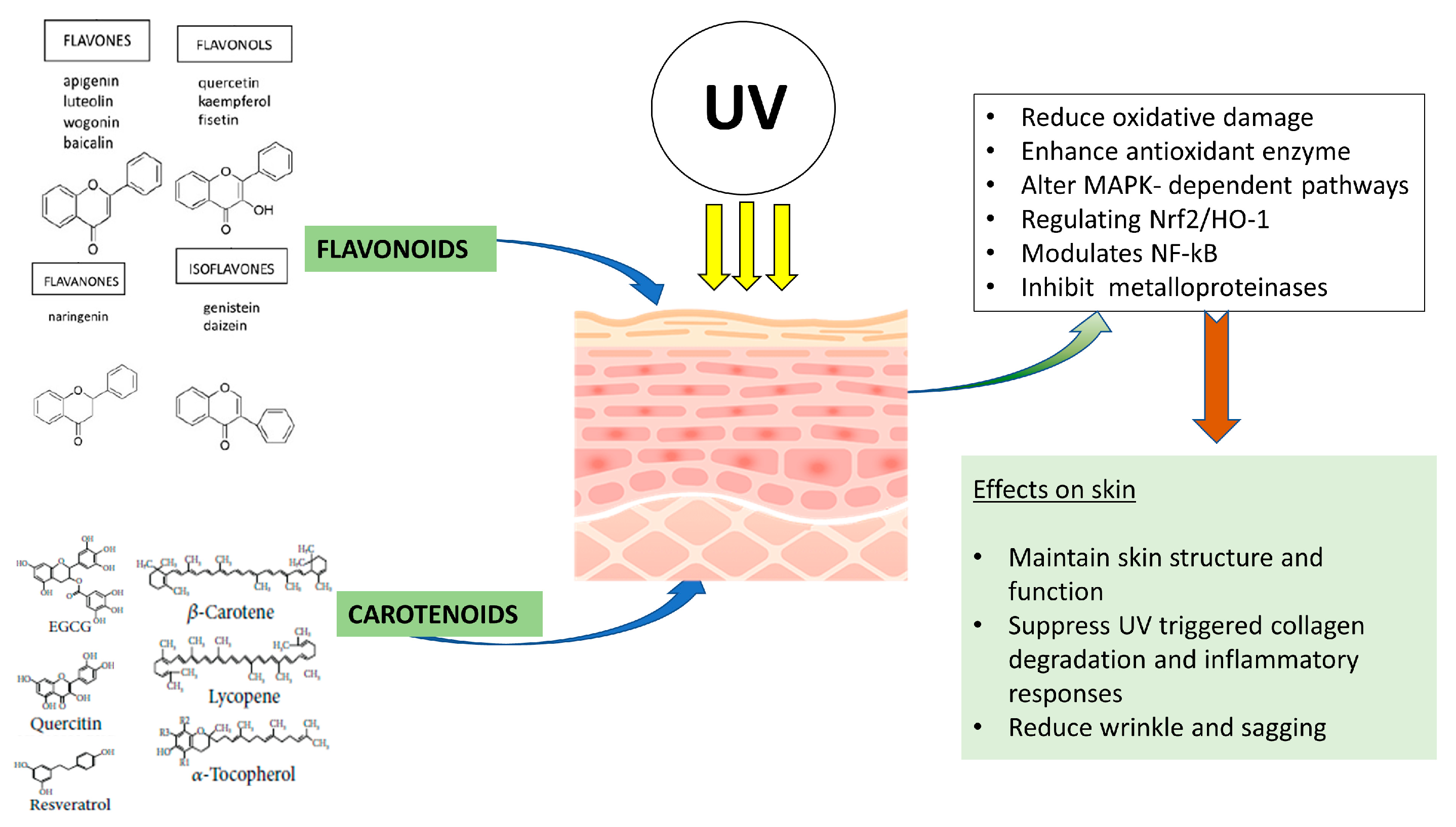{kind=link}
| Type of Plant Used | Sample/ Population | Methodology | Parameters Increased | Parameters Decreased | Parameters Unchanged | Conclusion |
|---|---|---|---|---|---|---|
| Green tea polyphenol (GTP) [19] | In vivo study Female SKH-1 hairless mice (6–7 weeks old). | GTP was given to SKH-1 hairless mice as drinking water (0.2%, wt/vol) before they were repeatedly exposed to UVB (90 mJ/cm2) for two months on alternate days. (a) H&E staining for microscopic histological evaluation (b) Skin biopsies were collected to determine protein carbonyl following DNPH analytical assay (c) Western blot analysis was done to assess the protein oxidation & MMP analysis |
| GTP exerts photoprotective effects by preventing oxidative damage and inhibition of MMP expression. | ||
| In vitro study Human fibroblast HS68. | Human fibroblast HS68 were cultured and treated with GTP before being exposed to UVB radiation (30 mJ per cm2) (a) Western blot analysis to access protein oxidation | |||||
| Green tea [20] | Double blind, placebo controlled clinicaltrial | (a) UV irradiation sensitivity was tested at weeks 0, 6, and 12 where dorsal skin at the back and scapular region was exposed to radiation. Skin tone was assessed before and after 24 h. (b) Skin elasticity was assessed using the Cutometer SEM 474 suction method on the inner forearm skin. (d) Skin density, thickness, structure and texture were examined using a high-frequency Ultrasound B-Scan. (e) Tewameter and Corneometer were used to measure transepidermal water loss (TEWL) and skin hydration in inner forearm. (f) The O2C-system (Lea Instruments) to assess peripheral blood flow and oxygen was used. |
|
| skin thinning | Regular intake of tea flavonoid in beverage was able to protect skin against UV radiation and improve skin quality and function |
| Bog blueberry extract (BBe) [21] | Human dermal fibroblast | Human dermal fibroblasts were pretreated with 1–10 mg/L ATH-BBe. Then, 100 mJ/cm2 UV-B was used to expose the pre-treated cell culture. (a) The MTT assay was used to assess cellular viability following UVB exposure. (b) Procollagen and collagenolytic MMP secretion expression levels were assessed using Western blot. (c) ELISA was used to measure the cytokine secretion from UV-B-irradiated fibroblasts. (d) Immunoblot analysis was used to assess NF-kB activation and MAPK signalling. |
|
| Anthocyanins were able to alleviate UV radiation injury via anti-inflammatory action and inhibition of NF-kB activation and MAPK signalling. | |
| Rosemary extract, citrus bioflavonoid extract [22] | In vitro study The human keratinocytes cell (HaCaT cells) | Human keratinocyte cells were cultured and treated with rosemary extract and citrus bioflavonoid extract before exposure to UVB. (a) Cell protection level was estimated as the percentage of cell viability recovered using the MTT assay, which was used to determine cellular viability. (b) The Folin-Ciocalteu technique was used to calculate the total phenolic compound (TPC). (c) The Comet assay was used to measure DNA damage. (d) A CBMN technique was used to conduct micronucleus assays on the irradiated lymphocytes. |
|
| Synergistic effect of both polyphenols in photoprotection by reducing oxidative damage. | |
| Human study Double blind, placebo controlled clinical trial | MED was measured pre and post-treatment. | |||||
| Luteolin from carrots [23] | In vitro study HaCaT human keratinocyte cell line | Before being exposed to UVB (0.01 J/cm2), HaCaT cells were treated for 1 h with luteolin (5 and 10 lM) and then incubated for 5 or 3 h, respectively. (a) The expression of MMP-1, JNK, c-Jun, ERK, and RSK phosphorylation were measured by Western blot analysis. (b) Using the luciferase assay, AP-1 transactivation and c-Fos promoter activation were quantified. (c) The expression of JNK1, JNK2, ERK2, and p90RSK was assessed using kinase assay. (d) JNK1, JNK2, and RSK2 expressions were analysed using pull-down assay. |
| Activation of JNK2 and ERK | Luteolin was able to provide protection by inhibiting UVB-induced signal transduction | |
| In vivo study SKH-1 hairless mice (5 weeks of age; mean body weight, 25 g) | For 15 weeks, luteolin (10 and 40 nmol) dissolved in 200L acetone was applied topically to the backs of SKH-1 hairless mice. This treatment began 1 h before UVB exposure (0.18 J/cm2). (a) Western blot analysis was done to assess MMP-13 expression (b) Grading of wrinkle formation. | |||||
| Tomato and rosemary extract [24] | Double blind, placebo controlled clinical trial | 149 participants were randomly assigned to carotenoid-rich tomato nutrient complex (TNC) or placebo for a 12-week period (a) A dose of 1.25 MED with Dermalight® 80 MED was administered to an additional field (12 × 12 mm) of the buttock to take biopsies and to determine the skin’s colour using chromametry. A change in skin tone quantified the development of erythema. (b) Blood samples were taken, and serum was evaluated for the presence of carotenoids at random intervals during a 5-week washout phase, after four weeks of therapy, and after the study. Two colourless carotenoids (phytofluene and phytoene), were also measured. (c) Biopsies were taken after 24 h following chromametry using 1.25 MED in order to evaluate UV-induced gene expression. |
|
|
| The carotenoid-rich supplement significantly prevented UVB-induced erythema and cytokine upregulation. |
| Tomato [25] | Single blind, placebo controlled clinical trial. | Volunteers received either tomato paste and olive oil or just olive oil. Their skin was exposed to UVB and they were assessed for erythema at baseline and week 12. (a) Immunohistochemical analysis was done to identify a panel of ECM molecules or remodelling enzymes in frozen sections. (b) A five-point semiquantitative scale was used to evaluate the level of immunostaining for procollagen and fibrillin-1, (c) MMP-1-positive dermal cell counts were calculated with high-power field (×400) |
| MMP-1 expression |
| Lycopene-rich tomato paste offers defense against both immediate and possible long-term effects of photodamage. |
| Grape peel extract (GPE) [26] | In vitro study HaCaT cells In vivo study: ICR six-week-old male mice | HaCaT cells were treated for 24 h with GPE or resveratrol (RES). Then, HaCaT cells were subsequently exposed to UVB radiation at a dose of 25 mJ/cm2. (a) Assessment of fractionation of nuclear and cytoplasmic proteins using Western blot analysis. Before receiving UVB (280–315 nm) radiation treatment, the dorsal skin of mice was shaved and they were given GPE orally three times per week for seven weeks. (a) Histological analysis of the skin tissue from mice (b) Western blot analysis |
|
| Resveratrol exhibits photoprotection and photoageing effects via regulation of nuclear Nrf2. | |
| Grape seed proanthocyanidin extract (GSPE) [27] | Double blind, placebo controlled clinical trial. | Cutaneous areas on the backs of volunteers were untreated or treated with GSPE solutions, 30 min before exposure to two minimal erythema doses (MED) of solar simulated radiation. (a) Cutaneous regions at various sites were examined histologically for the number of sunburn cells. (b) Mutant p53-positive cells and CD1a+ Langerhans cells were identified using immunohistochemistry. | Langerhans cells exhibit more dendrites |
| Topical GSPE prevents formation of sunburn cells, mutant p53 epidermal cells. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anbualakan, K.; Tajul Urus, N.Q.; Makpol, S.; Jamil, A.; Mohd Ramli, E.S.; Md Pauzi, S.H.; Muhammad, N. A Scoping Review on the Effects of Carotenoids and Flavonoids on Skin Damage Due to Ultraviolet Radiation. Nutrients 2023, 15, 92. https://doi.org/10.3390/nu15010092
Anbualakan K, Tajul Urus NQ, Makpol S, Jamil A, Mohd Ramli ES, Md Pauzi SH, Muhammad N. A Scoping Review on the Effects of Carotenoids and Flavonoids on Skin Damage Due to Ultraviolet Radiation. Nutrients. 2023; 15(1):92. https://doi.org/10.3390/nu15010092
Chicago/Turabian StyleAnbualakan, Kirushmita, Nur Qisti Tajul Urus, Suzana Makpol, Adawiyah Jamil, Elvy Suhana Mohd Ramli, Suria Hayati Md Pauzi, and Norliza Muhammad. 2023. "A Scoping Review on the Effects of Carotenoids and Flavonoids on Skin Damage Due to Ultraviolet Radiation" Nutrients 15, no. 1: 92. https://doi.org/10.3390/nu15010092
APA StyleAnbualakan, K., Tajul Urus, N. Q., Makpol, S., Jamil, A., Mohd Ramli, E. S., Md Pauzi, S. H., & Muhammad, N. (2023). A Scoping Review on the Effects of Carotenoids and Flavonoids on Skin Damage Due to Ultraviolet Radiation. Nutrients, 15(1), 92. https://doi.org/10.3390/nu15010092