
Probing the Double-Layered Cotyledon Cell Structure of Navy Beans: Barrier Effect of the Protein Matrix on In Vitro Starch Digestion



Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Navy Bean Materials
2.2.1. Isolation of Free Starch Granules
2.2.2. Isolation of Cotyledon Cells
2.2.3. Enzymatic Removal of Intracellular Protein Matrix
2.3. Determination of Physicochemical Properties
2.4. Determination of α-Amylase Activity
2.5. In Vitro Starch Digestion
2.5.1. Static In Vitro Starch Digestion Procedure
2.5.2. Effect of Proteolytic Enzymes on In Vitro Starch Digestion
2.6. Microscopy Analysis
2.7. Statistical Analysis
3. Results and Discussion
3.1. Microstructural Characteristics of Isolated Navy Bean Cotyledon Cells
3.2. Effect of the Intracellular Protein Matrix on Physicochemical Properties of Starch
3.3. Effect of the Intracellular Protein Matrix on In Vitro Starch Digestion
3.4. Effect of Proteolytic Enzymes on In Vitro Starch Digestion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Becerra-Tomás, N.; Díaz-López, A.; Rosique-Esteban, N.; Ros, E.; Buil-Cosiales, P.; Corella, D.; Estruch, R.; Fitó, M.; Serra-Majem, L.; Aros, F.; et al. Legume consumption is inversely associated with type 2 diabetes incidence in adults: A prospective assessment from the PREDIMED study. Clin. Nutr. 2018, 37, 906–913. [Google Scholar] [CrossRef] [PubMed]
- Sievenpiper, J.; Kendall, C.; Esfahani, A.; Wong, J.; Carleton, A.; Jiang, H.Y.; Jenkins, D.J.A. Effect of non-oil-seed pulses on glycaemic control: A systematic review and meta-analysis of randomised controlled experimental trials in people with and without diabetes. Diabetologia 2009, 52, 1479–1495. [Google Scholar] [CrossRef] [PubMed]
- Villegas, R.; Gao, Y.-T.; Yang, G.; Li, H.-L.; Elasy, T.A. Legume and soy food intake and the incidence of type 2 diabetes in the Shanghai Women’s Health Study. Am. J. Clin. Nutr. 2008, 87, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, D.; Ghafari, H.; Wolever, T.; Taylor, R.; Jenkins, A.L.; Barker, H.M.; Bowling, A.C. Relationship between rate of digestion of foods and post-prandial glycaemia. Diabetologia 1982, 22, 450–455. [Google Scholar] [CrossRef]
- Tharanathan, R.; Mahadevamma, S. Grain legumes—A boon to human nutrition. Trends Food Sci. Technol. 2003, 14, 507–518. [Google Scholar] [CrossRef]
- Tovar, J. Bioavailability of carbohydrates in legumes: Digestible and indigestible fractions. Arch. Latinoam. Nutr. 1996, 44, 36S–40S. [Google Scholar]
- Berg, T.; Singh, J.; Hardacre, A.; Boland, M.J. The role of cotyledon cell structure during in vitro digestion of starch in navy beans. Carbohyd. Poly. 2012, 87, 1678–1688. [Google Scholar] [CrossRef]
- Brummer, Y.; Kaviani, M.; Tosh, S.M. Structural and functional characteristics of dietary fibre in beans, lentils, peas and chickpeas. Food Res. Internat. 2015, 67, 117–125. [Google Scholar] [CrossRef]
- Dhital, S.; Bhattarai, R.R.; Gorham, J.; Gidley, M.J. Intactness of cell wall structure controls the in vitro digestion of starch in legumes. Food Funct. 2016, 7, 1367–1379. [Google Scholar] [CrossRef]
- Do, D.T.; Singh, J.; Oey, I.; Singh, H. Modulating effect of cotyledon cell microstructure on in vitro digestion of starch in legumes. Food Hydrocol. 2019, 96, 112–122. [Google Scholar] [CrossRef]
- Do, D.T.; Singh, J.; Oey, I.; Singh, H.; Yada, R.Y.; Frostad, J.M. A novel apparatus for time-lapse optical microscopy of gelatinisation and digestion of starch inside plant cells. Food Hydrocol. 2020, 104, 105551. [Google Scholar] [CrossRef]
- Noah, L.; Guillon, F.; Bouchet, B.; Buleon, A.; Molis, C.; Gratas, M.; Champ, M. Digestion of carbohydrate from white beans (Phaseolus vulgaris L.) in healthy humans. J. Nutr. 1998, 128, 977–985. [Google Scholar] [CrossRef] [PubMed]
- Do, D.T.; Singh, J. Legume microstructure. In Encyclopedia of Food Chemistry; Varelis, P., Melton, L.D., Shahidi, F., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; Volume 3, pp. 15–21. [Google Scholar] [CrossRef]
- Pallaresares, A.P.; Gwala, S.; Pälchen, K.; Duijsens, D.; Hendrickx, M.; Grauwet, T. Pulse seeds as promising and sustainable source of ingredients with naturally bioencapsulated nutrients: Literature review and outlook. Comp. Rev. Food Sci. Food Saf. 2021, 20, 1524–1553. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, R.R.; Dhital, S.; Wu, P.; Chen, X.D.; Gidley, M.J. Digestion of isolated legume cells in a stomach-duodenum model: Three mechanisms limit starch and protein hydrolysis. Food Func. 2017, 8, 2573–2582. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Gidley, M.J.; Dhital, S. Wall porosity in isolated cells from food plants: Implications for nutritional functionality. Food Chem. 2019, 279, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Pallares, A.P.; Miranda, B.A.; Truong, N.Q.A.; Kyomugasho, C.; Chigwedere, C.M.; Hendrickx, M.; Grauwet, T. Process-induced cell wall permeability modulates the in vitro starch digestion kinetics of common bean cotyledon cells. Food Func. 2018, 9, 6544–6554. [Google Scholar] [CrossRef]
- Rovalino-Córdova, A.M.; Fogliano, V.; Capuano, E. A closer look to cell structural barriers affecting starch digestibility in beans. Carbohy. Poly. 2018, 181, 994–1002. [Google Scholar] [CrossRef]
- Rovalino-Córdova, A.M.; Fogliano, V.; Capuano, E. The effect of cell wall encapsulation on macronutrients digestion: A case study in kidney beans. Food Chem. 2019, 286, 557–566. [Google Scholar] [CrossRef]
- AOAC. Method 968.06. Protein (crude) in animal feed. In Official Methods of Analysis of the Association of Official Analytical Chemists (AOAC); Latimer, G.W., Horwitz, W., Eds.; AOAC International: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Leach, H.; McCowen, D.; Schoch, T. Swelling and solubility patterns of various starches, structure of starch granule. Cereal Chem. 1959, 36, 534–544. [Google Scholar]
- Bernfeld, P. Amylases, α and β. Methods Enzymol. 1955, 1, 149–158. [Google Scholar]
- Dartois, A.; Singh, J.; Kaur, L.; Singh, H. Influence of guar gum on the in vitro starch digestibility—Rheological and microstructural characteristics. Food Biophys. 2010, 5, 149–160. [Google Scholar] [CrossRef]
- Pharmacopeia, U.S. Pharmacopeia, U.S. Pharmacopeia, simulated gastric fluid, TS, simulated intestinal fluid, TS, United States Pharmacopeial Convention. In The National Formulary 9; U.S. Pharmacopeia Board of Trustees: Rockville, MD, USA, 2000; Volume 24, p. 2235. [Google Scholar]
- Ezeogu, L.; Duodu, K.; Taylor, J. Effects of endosperm texture and cooking conditions on the in vitro starch digestibility of sorghum and maize flours. J. Cereal Sci. 2005, 42, 33–44. [Google Scholar] [CrossRef]
- Xiong, W.; Zhang, B.; Huang, Q.; Li, C.; Pletsch, E.A.; Fu, X. Variation in the rate and extent of starch digestion is not determined by the starch structural features of cooked whole pulses. Food Hydrocol. 2018, 83, 340–347. [Google Scholar] [CrossRef]
- Fujimura, T.; Kugimiya, M. Gelatinization of starches inside cotyledon cells of kidney beans. Starch-Stärke 1994, 46, 374–378. [Google Scholar] [CrossRef]
- Fujimuru, T.; Kugimiya, M. Estimation of intracellular water content during gelatinization of starches inside cotyledon cells of legumes. J. App. Glycosci. 1995, 42, 7–13. [Google Scholar]
- Gwala, S.; Pallares, A.P.; Pälchen, K.; Hendrickx, M.; Grauwet, T. In vitro starch and protein digestion kinetics of cooked Bambara groundnuts depend on processing intensity and hardness sorting. Food Res. Internat. 2020, 137, 109512. [Google Scholar] [CrossRef]
- Ezeogu, L.I.; Duodu, K.G.; Emmambux, M.N.; Taylor, J.R. Influence of cooking conditions on the protein matrix of sorghum and maize endosperm flours. Cereal Chem. 2008, 85, 397–402. [Google Scholar] [CrossRef]
- Kim, E.H.J.; Petrie, J.R.; Motoi, L.; Morgenstern, M.P.; Sutton, K.H.; Mishra, S.; Simmons, L.D. Effect of structural and physicochemical characteristics of the protein matrix in pasta on in vitro starch digestibility. Food Biophys. 2008, 3, 229–234. [Google Scholar] [CrossRef]
- Li, H.T.; Li, Z.; Fox, G.P.; Gidley, M.J.; Dhital, S. Protein-starch matrix plays a key role in enzymic digestion of high-amylose wheat noodle. Food Chem. 2021, 336, 127719. [Google Scholar] [CrossRef]
- Pallares, A.P.; Rousseau, S.; Chigwedere, C.M.; Kyomugasho, C.; Hendrickx, M.; Grauwet, T. Temperature-pressure-time combinations for the generation of common bean microstructures with different starch susceptibilities to hydrolysis. Food Res. Internat. 2018, 106, 105–115. [Google Scholar] [CrossRef]
- Wang, S.; Li, P.; Zhang, T.; Wang, S.; Copeland, L. Trypsin and chymotrypsin are necessary for in vitro enzymatic digestion of rice starch. RSC Adv. 2017, 7, 3660–3666. [Google Scholar] [CrossRef]
- Do, D.T.; Singh, J.; Oey, I.; Singh, H. Biomimetic plant foods: Structural design and functionality. Trends Food Sci. Technol. 2018, 82, 46–59. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
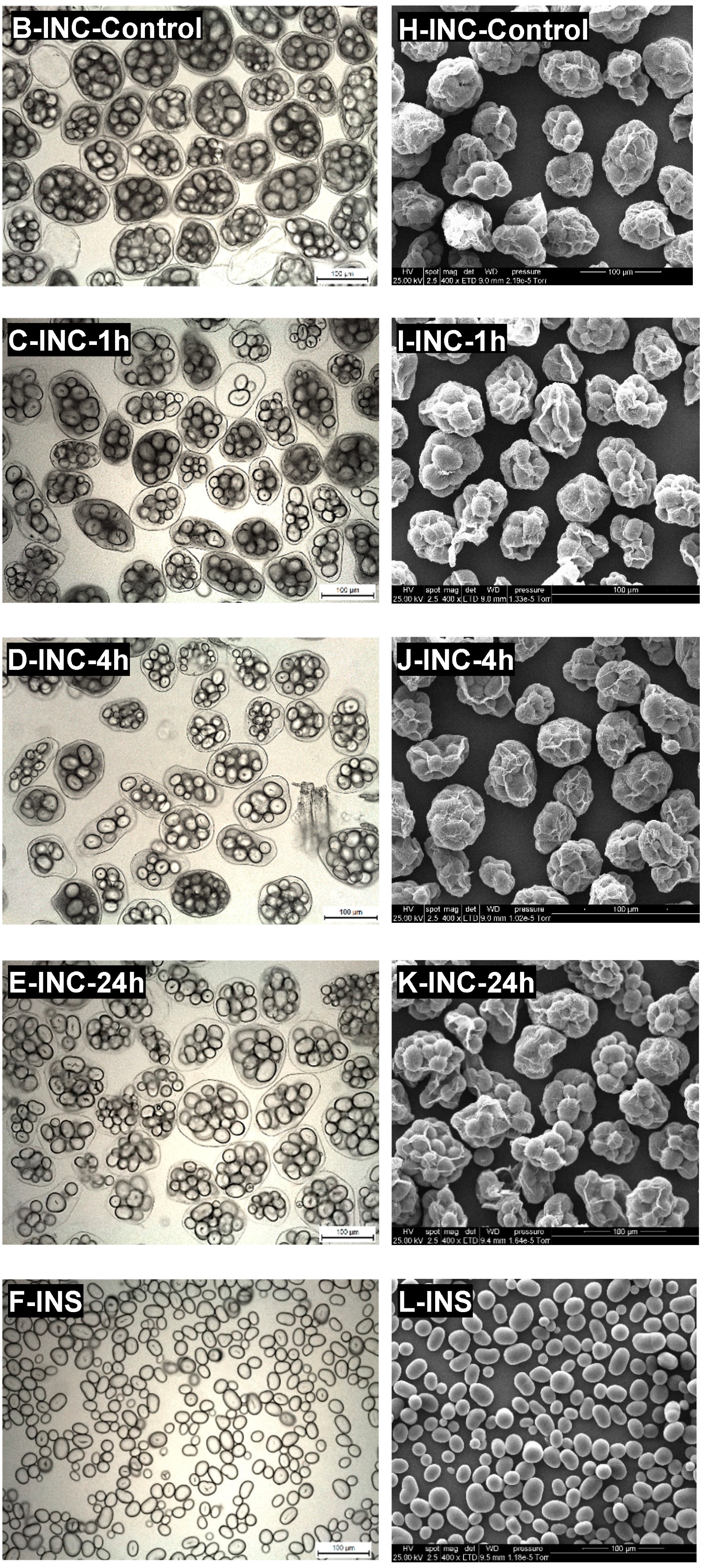{kind=link}
{kind=link}
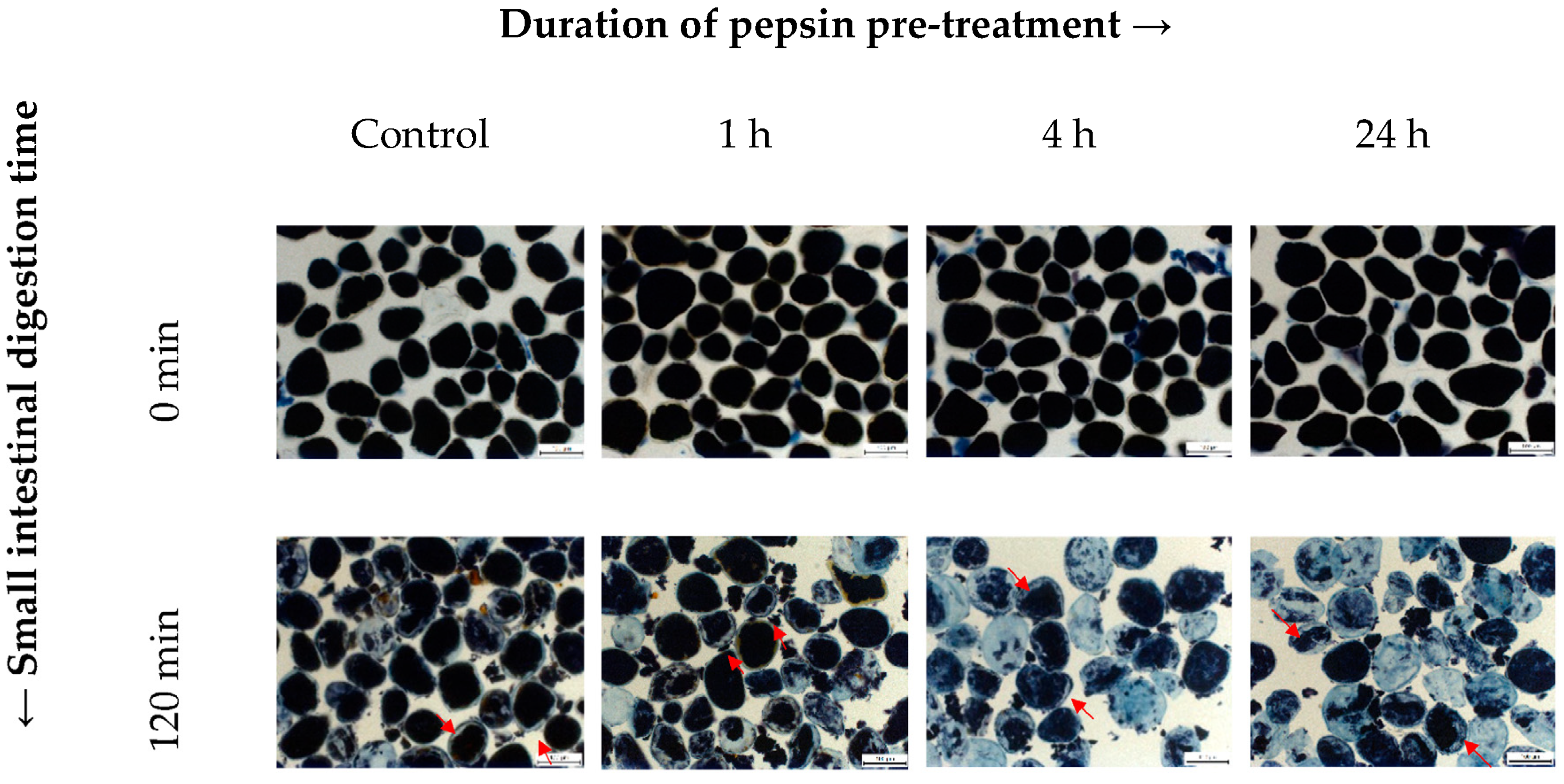{kind=link}
{kind=link}
| Navy Bean Material | AM (%) | TS (% dwb) | P (% dwb) | SP (g/g) | SS (%) |
|---|---|---|---|---|---|
| INC-Control | 28.8 ± 0.0 a | 64.3 ± 0.5 e | 17.6 ± 0.7 a | 5.2 ± 0.1 d | 5.5 ± 0.0 c |
| INC-1h | 28.6 ± 0.2 a | 70.5 ± 0.4 d | 11.4 ± 0.3 b | 5.4 ± 0.1 cd | 5.5 ± 0.0 c |
| INC-4h | 27.8 ± 0.3 a | 73.4 ± 0.2 c | 9.2 ± 0.3 c | 5.6 ± 0.0 c | 6.1 ± 0.1 c |
| INC-24h | 29.0 ± 0.0 a | 80.2 ± 0.5 b | 2.7 ± 0.2 d | 7.4 ± 0.2 b | 8.3 ± 0.3 b |
| INS | 28.0 ± 0.7 a | 91.9 ± 0.9 a | 0.3 ± 0.2 e | 12.6 ± 0.1 a | 16.9 ± 0.2 a |
| Navy Bean Materials | R10 (mg/mL/min) f | H120 (%) g |
|---|---|---|
| INC-Control | 1.40 ± 0.11 c | 70.6 ± 0.6 e |
| INC-1h | 1.50 ± 0.03 c | 73.2 ± 0.3 d |
| INC-4h | 2.07 ± 0.13 b | 77.6 ± 0.6 c |
| INC-24h | 2.11 ± 0.13 b | 80.6 ± 1.6 b |
| INS | 2.76 ± 0.13 a | 85.1 ± 0.4 a |
| Experiments | R10 (mg/mL/min) c | H120 (%) d |
|---|---|---|
| Exp.1 (GP + PP) | 1.40 ± 0.11 a | 70.6 ± 0.6 a |
| Exp.2 (PP only) | 1.41 ± 0.07 a | 69.9 ± 0.7 a |
| Exp.3 (GP only) | 0.97 ± 0.04 b | 70.8 ± 0.9 a |
| Exp.4 (without GP + PP) | 1.04 ± 0.04 b | 65.8 ± 1.6 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Do, D.T.; Singh, J.; Johnson, S.; Singh, H. Probing the Double-Layered Cotyledon Cell Structure of Navy Beans: Barrier Effect of the Protein Matrix on In Vitro Starch Digestion. Nutrients 2023, 15, 105. https://doi.org/10.3390/nu15010105
Do DT, Singh J, Johnson S, Singh H. Probing the Double-Layered Cotyledon Cell Structure of Navy Beans: Barrier Effect of the Protein Matrix on In Vitro Starch Digestion. Nutrients. 2023; 15(1):105. https://doi.org/10.3390/nu15010105
Chicago/Turabian StyleDo, Duc Toan, Jaspreet Singh, Stuart Johnson, and Harjinder Singh. 2023. "Probing the Double-Layered Cotyledon Cell Structure of Navy Beans: Barrier Effect of the Protein Matrix on In Vitro Starch Digestion" Nutrients 15, no. 1: 105. https://doi.org/10.3390/nu15010105
APA StyleDo, D. T., Singh, J., Johnson, S., & Singh, H. (2023). Probing the Double-Layered Cotyledon Cell Structure of Navy Beans: Barrier Effect of the Protein Matrix on In Vitro Starch Digestion. Nutrients, 15(1), 105. https://doi.org/10.3390/nu15010105