
Role of Chrononutrition in the Antihypertensive Effects of Natural Bioactive Compounds

 ,
,  , , ,
, , ,  and
and 
Abstract
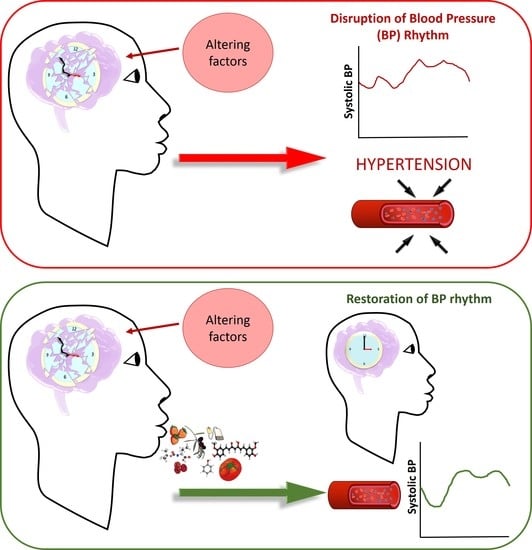
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Blood Pressure and Its Main Regulation Mechanisms
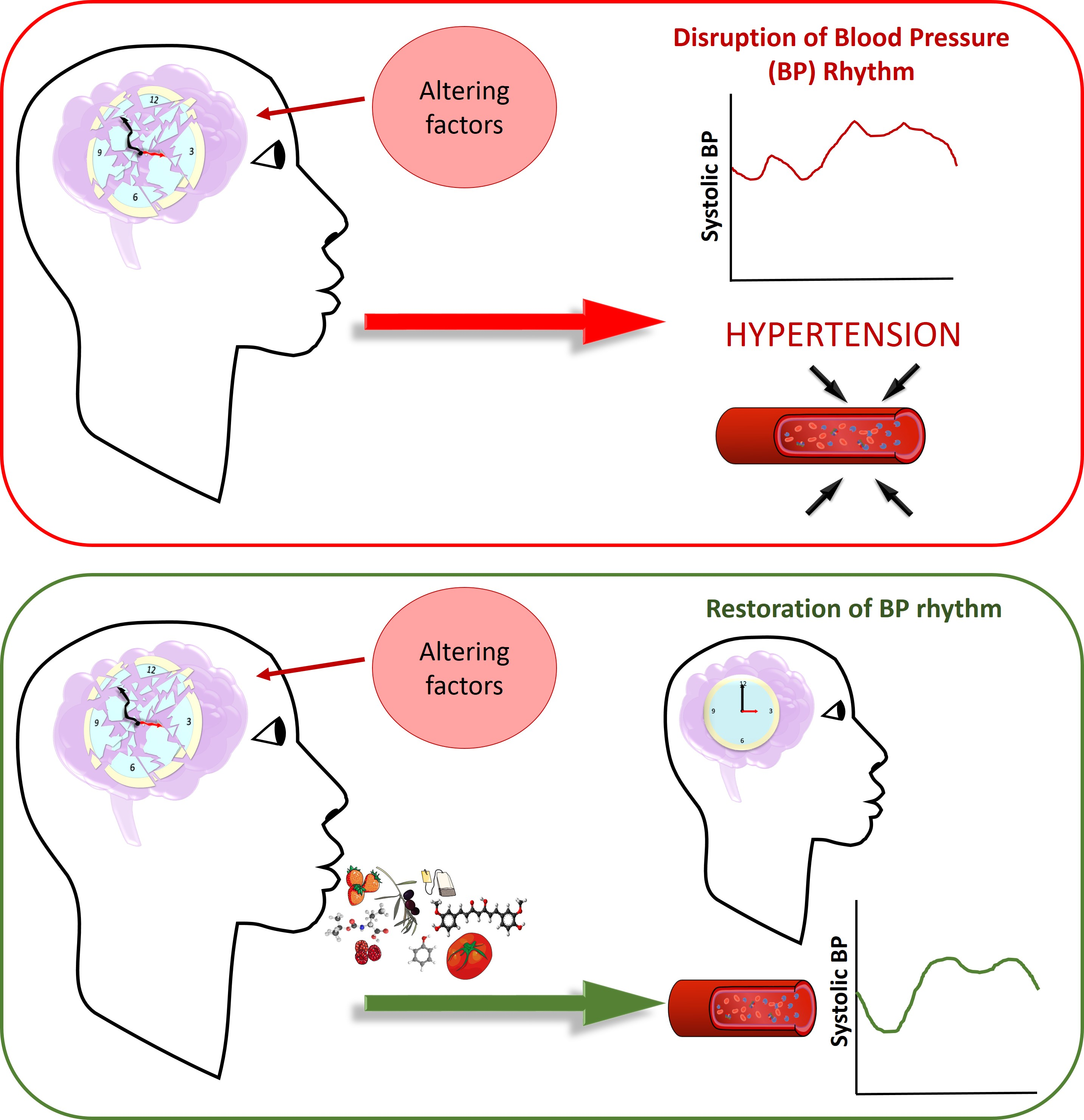
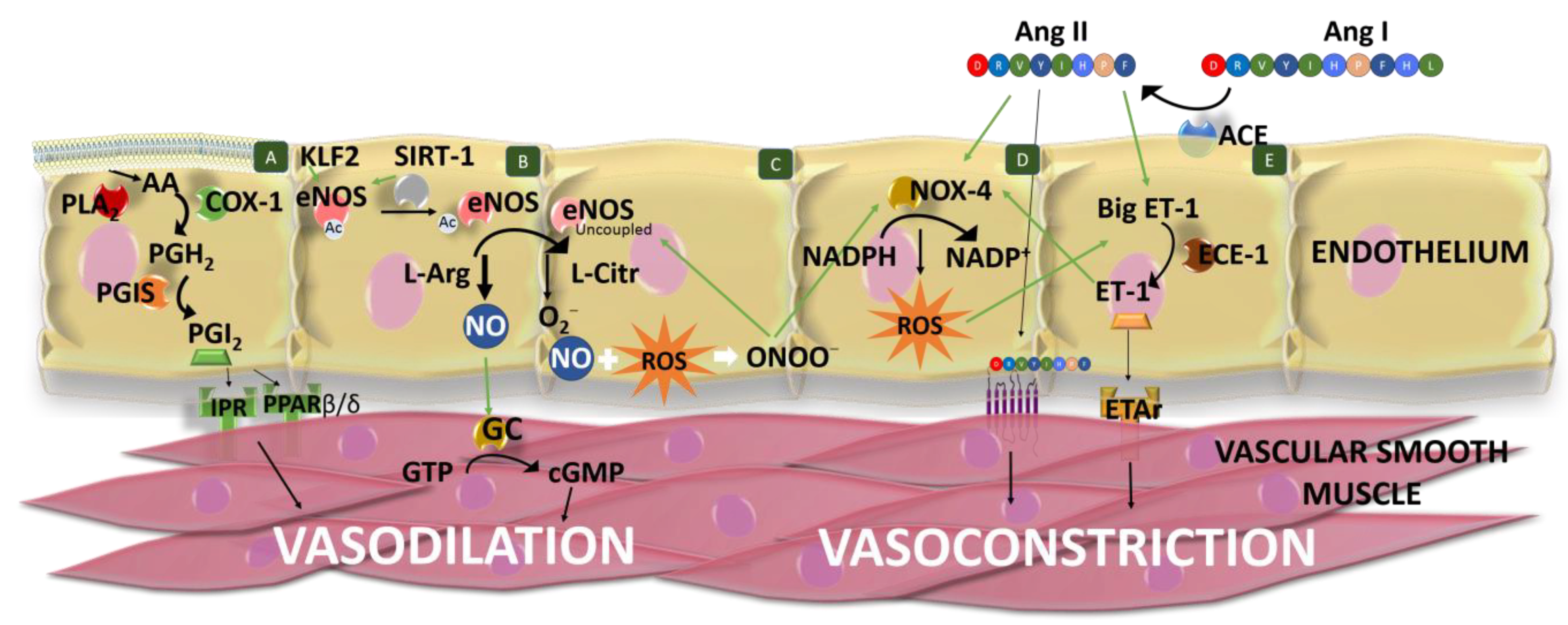
2.1. Renin–angiotensin–aldosterone System
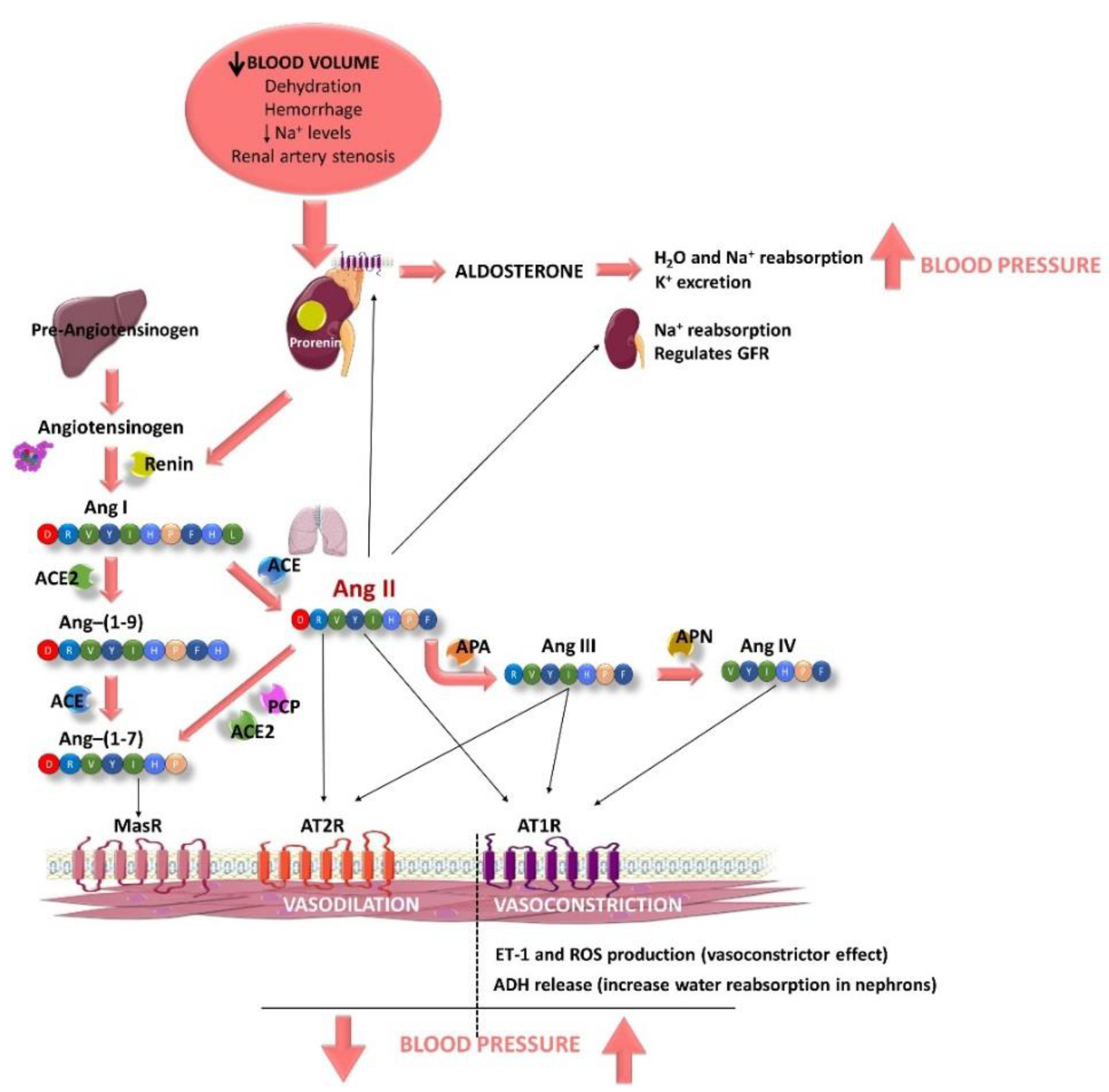
2.2. Endothelial Function
3. Biological Rhythms and Blood Pressure
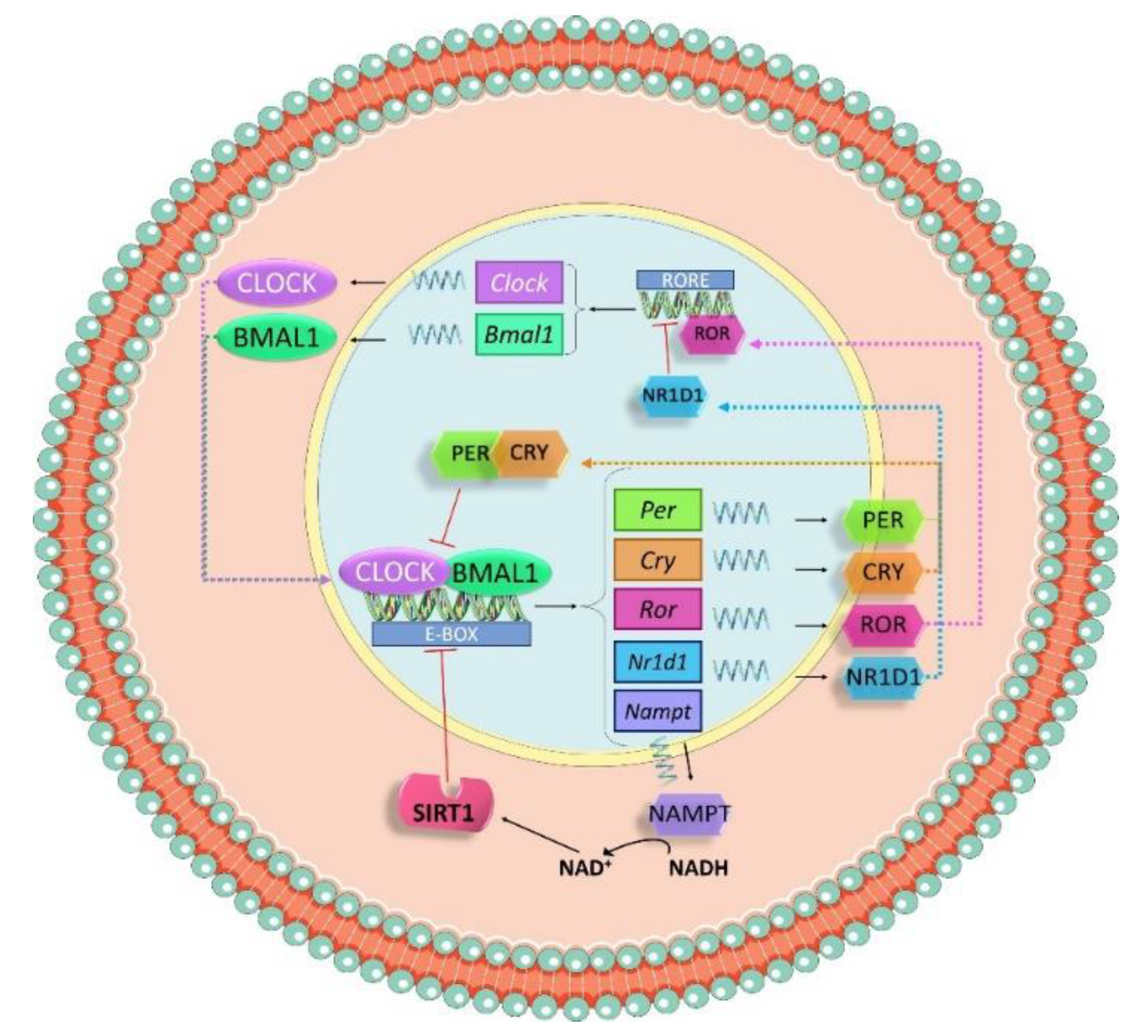
3.1. Molecular Machinery behind Circadian Rhtyhms
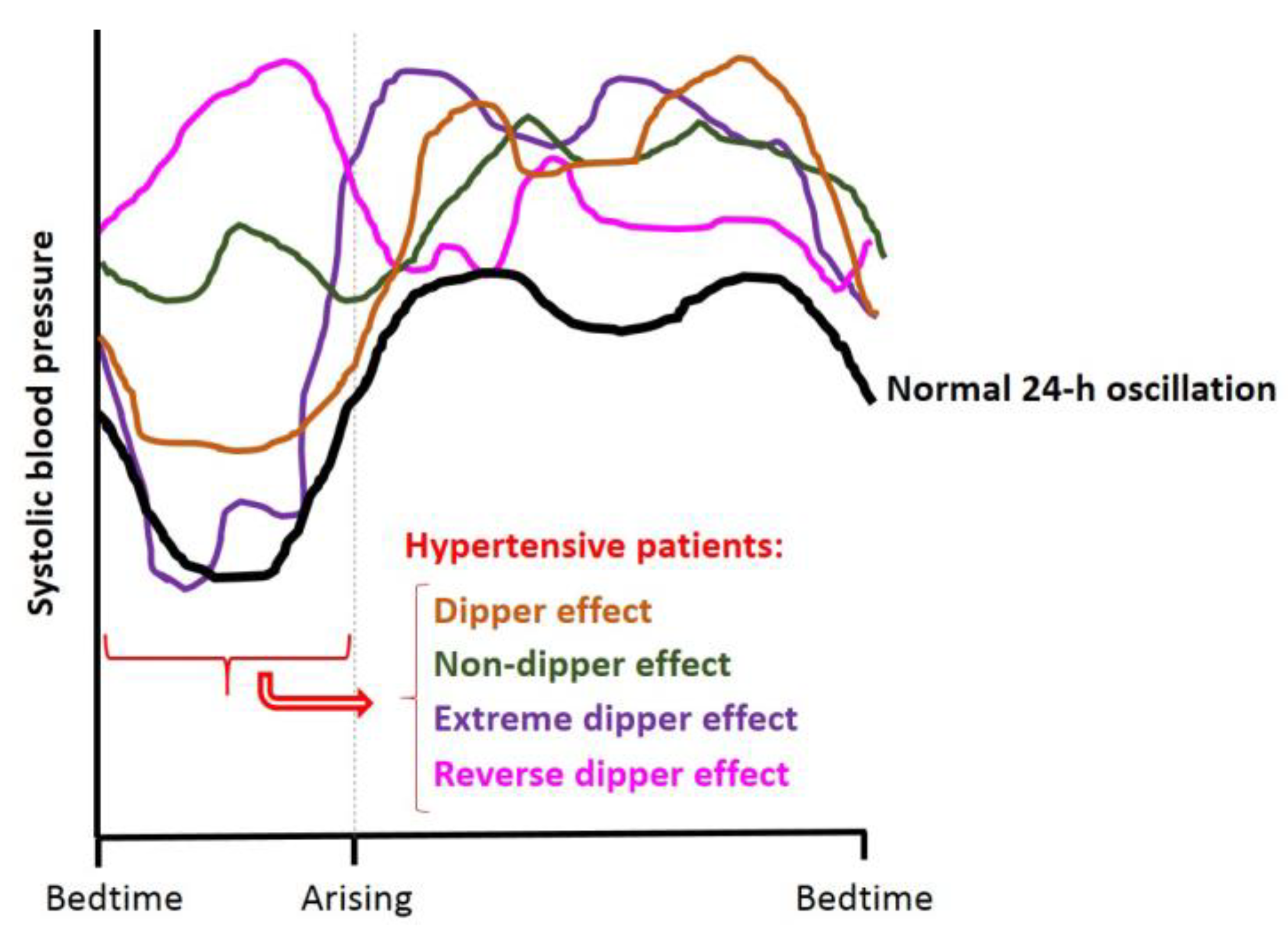
3.2. Circadian Blood Pressure Patterns
4. Hypertension
4.1. Treatments for Hypertension
4.1.1. Pharmacological Treatments
4.1.2. Natural Bioactive Compounds
5. Antihypertensive Compounds and their Role in Biological Rhythms
5.1. Pharmacological Treatments and Circadian Rhythms
5.2. Bioactive Compounds and Circadian Rhythms
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Unger, T.; Borghi, C.; Charchar, F.; Khan, N.A.; Poulter, N.R.; Prabhakaran, D.; Ramirez, A.; Schlaich, M.; Stergiou, G.S.; Tomaszewski, M.; et al. 2020 International Society of Hypertension Global Hypertension Practice Guidelines. Hypertension 2020, 75, 1334–1357. [Google Scholar] [CrossRef] [PubMed]
- Thomopoulos, C.; Parati, G.; Zanchetti, A. Effects of blood pressure lowering on outcome incidence in hypertension. 1. Overview, meta-analyses, and meta-regression analyses of randomized trials. J. Hypertens. 2014, 32, 2285–2295. [Google Scholar] [CrossRef] [PubMed]
- López-Fernández-Sobrino, R.L.; Torres-Fuentes, C.; Bravo, F.I.; Muguerza, B. Winery by-products as a valuable source for natural antihypertensive agents. Crit. Rev. Food Sci. Nutr. 2022, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Maria, A.G.; Graziano, R.; Nicolantonio, D. Carotenoids: Potential allies of cardiovascular health? Food Nutr. Res. 2015, 59, 26762. [Google Scholar] [CrossRef]
- Majumder, K.; Wu, J. Molecular Targets of Antihypertensive Peptides: Understanding the Mechanisms of Action Based on the Pathophysiology of Hypertension. Int. J. Mol. Sci. 2014, 16, 256–283. [Google Scholar] [CrossRef]
- Asif, M.A.; Lisa, S.R.; Qais, N. Exploring the Anti-Hypertensive Properties of Medicinal Plants and Their Bioactive Metabolites: An Extensive Review. Am. J. Plant Sci. 2021, 12, 1705–1740. [Google Scholar] [CrossRef]
- Mähler, A.; Wilck, N.; Rauch, G.; Dechend, R.; Müller, D.N. Effect of a probiotic on blood pressure in grade 1 hypertension (HYPRO): Protocol of a randomized controlled study. Trials 2020, 21, 1032. [Google Scholar] [CrossRef]
- Pimenta, F.S.; Luaces-Regueira, M.; Ton, A.M.; Campagnaro, B.P.; Campos-Toimil, M.; MC Pereira, T.; Vasquez, E.C. Mechanisms of Action of Kefir in Chronic Cardiovascular and Metabolic Diseases. Cell. Physiol. Biochem. 2018, 48, 1901–1914. [Google Scholar] [CrossRef]
- Silveira-Nunes, G.; Durso, D.F.; De Oliviera, L.R.A., Jr.; Cunha, E.H.M.; Maioli, T.U.; Vieira, A.T.; Speziali, E.; Corrêa-Oliveira, R.; Martins-Filho, O.A.; Teixeira-Carvalho, A.; et al. Hypertension Is Associated with Intestinal Microbiota Dysbiosis and Inflammation in a Brazilian Population. Front. Pharmacol. 2020, 11, 258. [Google Scholar] [CrossRef]
- Barona, J.; Aristizabal, J.C.; Blesso, C.N.; Volek, J.S.; Fernandez, M.L. Grape Polyphenols Reduce Blood Pressure and Increase Flow-Mediated Vasodilation in Men with Metabolic Syndrome. J. Nutr. 2012, 142, 1626–1632. [Google Scholar] [CrossRef]
- Li, S.-H.; Zhao, P.; Tian, H.-B.; Chen, L.-H.; Cui, L.-Q. Effect of Grape Polyphenols on Blood Pressure: A Meta-Analysis of Randomized Controlled Trials. PLoS ONE 2015, 10, e0137665. [Google Scholar] [CrossRef] [PubMed]
- Queipo-Ortuño, M.I.; Boto-Ordóñez, M.; Murri, M.; Gomez-Zumaquero, J.M.; Clemente-Postigo, M.; Estruch, R.; Cardona Diaz, F.; Andrés-Lacueva, C.; Tinahones, F.J. Influence of red wine polyphenols and ethanol on the gut microbiota ecology and biochemical biomarkers. Am. J. Clin. Nutr. 2012, 95, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Sano, A.; Uchida, R.; Saito, M.; Shioya, N.; Komori, Y.; Tho, Y.; Hashizume, N. Beneficial Effects of Grape Seed Extract on Malondialdehyde-Modified LDL. J. Nutr. Sci. Vitaminol. 2007, 53, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Mellen, P.B.; Daniel, K.R.; Brosnihan, K.B.; Hansen, K.J.; Herrington, D.M. Effect of Muscadine Grape Seed Supplementation on Vascular Function in Subjects with or at Risk for Cardiovascular Disease: A Randomized Crossover Trial. J. Am. Coll. Nutr. 2010, 29, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Ward, N.C.; Hodgson, J.M.; Croft, K.; Burke, V.; Beilin, L.J.; Puddey, I.B. The combination of vitamin C and grape-seed polyphenols increases blood pressure: A randomized, double-blind, placebo-controlled trial. J. Hypertens. 2005, 23, 427–434. [Google Scholar] [CrossRef]
- Miralles, B.; Amigo, L.; Recio, I. Critical Review and Perspectives on Food-Derived Antihypertensive Peptides. J. Agric. Food Chem. 2018, 66, 9384–9390. [Google Scholar] [CrossRef]
- Gallego, M.; Mora, L.; Toldrá, F. Health relevance of antihypertensive peptides in foods. Curr. Opin. Food Sci. 2018, 19, 8–14. [Google Scholar] [CrossRef]
- Magder, S. The meaning of blood pressure. Crit. Care 2018, 22, 1–10. [Google Scholar] [CrossRef]
- Brzezinski, W.A. Blood pressure. In Clinical Methods: The History, Physical, and Laboratory Examinations; Walker, H., Hall, W., Hurst, J., Eds.; Butterworths: Boston, MA, USA, 1990; ISBN 040990077X. [Google Scholar]
- Bartels, K.; Esper, S.A.; Thiele, R. Blood Pressure Monitoring for the Anesthesiologist. Anesth. Analg. 2016, 122, 1866–1879. [Google Scholar] [CrossRef]
- Margalef, M.; Bravo, F.I.; Arola-Arnal, A.; Muguerza, B. Natural Angiotensin Converting Enzyme (ACE) inhibitors with antihypetensive properties. In Natural Products Targeting Clinically Relevant Enzymes; Andrade, P., Valentao, P., Pereira, D.M., Eds.; Wiley-VCH GmbH & Co.: Weinheim, Germany, 2017; pp. 45–67. [Google Scholar]
- Valensi, P. Autonomic nervous system activity changes in patients with hypertension and overweight: Role and therapeutic implications. Cardiovasc. Diabetol. 2021, 20, 1–12. [Google Scholar] [CrossRef]
- Hart, E.; Charkoudian, N. Sympathetic Neural Regulation of Blood Pressure: Influences of Sex and Aging. Physiology 2014, 29, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Tigerstedt, R.; Bergman, P.Q. Niere und Kreislauf1. Skand. Arch. Physiol. 1898, 8, 223–271. [Google Scholar] [CrossRef]
- Goldblatt, H.; Lynch, J.; Hanzal, R.F.; Summerville, W.W. Studies on Experimental Hypertension. J. Exp. Med. 1934, 59, 347–379. [Google Scholar] [CrossRef] [PubMed]
- Milei, J. A cornerstone in the history of hypertension: The seventieth anniversary of the discovery of angiotensin. J. Cardiovasc. Med. 2010, 11, 260–264. [Google Scholar] [CrossRef]
- Basso, N.; Terragno, N.A. History About the Discovery of the Renin-Angiotensin System. Hypertension 2001, 38, 1246–1249. [Google Scholar] [CrossRef]
- Patel, S.; Rauf, A.; Khan, H.; Abu-Izneid, T. Renin-angiotensin-aldosterone (RAAS): The ubiquitous system for homeostasis and pathologies. Biomed. Pharmacother. 2017, 94, 317–325. [Google Scholar] [CrossRef]
- Soubrier, F.; Cambien, F. Renin-angiotensin system genes as candidate genes in cardiovascular diseases. Trends Cardiovasc. Med. 1993, 3, 250–258. [Google Scholar] [CrossRef]
- Riordan, J.F. Angiotensin-I-converting enzyme and its relatives. Genome Biol. 2003, 4, 225. [Google Scholar] [CrossRef][Green Version]
- Watanabe, T.; Barker, T.A.; Berk, B.C. Angiotensin II and the endothelium: Diverse signals and effects. Hypertension 2005, 45, 163–169. [Google Scholar] [CrossRef]
- Iwai, M.; Horiuchi, M. Devil and angel in the renin–angiotensin system: ACE–angiotensin II–AT1 receptor axis vs. ACE2–angiotensin-(1–7)–Mas receptor axis. Hypertens. Res. 2009, 32, 533–536. [Google Scholar] [CrossRef]
- Pueyo, M.E.; Michel, J.-B. Angiotensin II receptors in endothelial cells. Gen. Pharmacol. Vasc. Syst. 1997, 29, 691–696. [Google Scholar] [CrossRef]
- Fountain, J.H.; Lappin, S.L. Physiology, Renin Angiotensin System; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Touyz, R.M.; Chen, X.; Tabet, F.; Yao, G.; He, G.; Quinn, M.; Pagano, P.J.; Schiffrin, E.L. Expression of a Functionally Active gp91phox-Containing Neutrophil-Type NAD(P)H Oxidase in Smooth Muscle Cells from Human Resistance Arteries. Circ. Res. 2002, 90, 1205–1213. [Google Scholar] [CrossRef] [PubMed]
- Osborn, J.W.; Foss, J.D. Renal Nerves and Long-Term Control of Arterial Pressure. Compr. Physiol. 2017, 7, 263–320. [Google Scholar] [CrossRef]
- Gupta, M. Vasoactive substances as mediators of renal injury. In Comprehensive Toxicology; McQueen, C.A., Ed.; Elsevier: Amsterdam, The Netherlands, 2010; pp. 305–327. [Google Scholar]
- Blair-West, J.R.; Carey, K.D.; Denton, D.A.; Madden, L.J.; Weisinger, R.S.; Shade, R.E. Possible contribution of brain angiotensin III to ingestive behaviors in baboons. Am. J. Physiol. Integr. Comp. Physiol. 2001, 281, R1633–R1636. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.J.; Chou, L.L.; Ger, L.P.; Tung, C.S. Cardiovascular effects of angiotensin III in brainstem nuclei of normotensive and hypertensive rats. J. Pharmacol. Exp. Ther. 1994, 268, 558–564. [Google Scholar] [PubMed]
- Yugandhar, V.G.; Clark, M.A. Angiotensin III: A physiological relevant peptide of the renin angiotensin system. Peptides 2013, 46, 26–32. [Google Scholar] [CrossRef] [PubMed]
- De Bundel, D.; Smolders, I.; Vanderheyden, P.; Michotte, Y. Ang II and Ang IV: Unraveling the Mechanism of Action on Synaptic Plasticity, Memory, and Epilepsy. CNS Neurosci. Ther. 2008, 14, 315–339. [Google Scholar] [CrossRef]
- Touyz, R.M.; Montezano, A.C. Angiotensin-(1–7) and Vascular Function. Hypertension 2018, 71, 68–69. [Google Scholar] [CrossRef]
- Shang, J.; Ye, G.; Shi, K.; Wan, Y.; Luo, C.; Aihara, H.; Geng, Q.; Auerbach, A.; Li, F. Structural basis of receptor recognition by SARS-CoV-2. Nature 2020, 581, 221–224. [Google Scholar] [CrossRef]
- Loera-Valencia, R.; Eroli, F.; Garcia-Ptacek, S.; Maioli, S. Brain Renin–Angiotensin System as Novel and Potential Therapeutic Target for Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 10139. [Google Scholar] [CrossRef]
- Wright, J.W.; Harding, J.W. Contributions by the Brain Renin-Angiotensin System to Memory, Cognition, and Alzheimer’s Disease. J. Alzheimer’s Dis. 2019, 67, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Kauwe, J.S.K.; Bailey, M.H.; Ridge, P.G.; Perry, R.; Wadsworth, M.E.; Hoyt, K.L.; Staley, L.A.; Karch, C.; Harari, O.; Cruchaga, C.; et al. Genome-Wide Association Study of CSF Levels of 59 Alzheimer’s Disease Candidate Proteins: Significant Associations with Proteins Involved in Amyloid Processing and Inflammation. PLoS Genet. 2014, 10, e1004758. [Google Scholar] [CrossRef] [PubMed]
- Jochemsen, H.M.; Teunissen, C.E.; Ashby, E.L.; Van Der Flier, W.M.; Jones, R.E.; Geerlings, M.I.; Scheltens, P.; Kehoe, P.G.; Muller, M. The association of angiotensin-converting enzyme with biomarkers for Alzheimer’s disease. Alzheimer’s Res. Ther. 2014, 6, 27. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, H.A.; Ishrat, T. The Brain AT2R—A Potential Target for Therapy in Alzheimer’s Disease and Vascular Cognitive Impairment: A Comprehensive Review of Clinical and Experimental Therapeutics. Mol. Neurobiol. 2020, 57, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Flammer, A.J.; Lüscher, T.F. Three decades of endothelium research: From the detection of nitric oxide to the everyday implementation of endothelial function measurements in cardiovascular diseases. Swiss Med. Wkly. 2010, 140, 1–9. [Google Scholar] [CrossRef]
- Desideri, G.; Ferri, C. Cardiovascular risk and endothelial dysfunction: The preferential route for atherosclerosis. Curr. Pharm. Biotechnol. 2011, 12, 1343–1353. [Google Scholar] [CrossRef]
- Incalza, M.A.; D’Oria, R.; Natalicchio, A.; Perrini, S.; Laviola, L.; Giorgino, F. Oxidative stress and reactive oxygen species in endothelial dysfunction associated with cardiovascular and metabolic diseases. Vasc. Pharmacol. 2018, 100, 459–469. [Google Scholar] [CrossRef]
- Ghalayini, I.F. Nitric oxide–cyclic GMP pathway with some emphasis on cavernosal contractility. Int. J. Impot. Res. 2004, 16, 459–469. [Google Scholar] [CrossRef]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef]
- Chen, W.; Xiao, H.; Rizzo, A.; Zhang, W.; Mai, Y.; Ye, M. Endothelial Nitric Oxide Synthase Dimerization Is Regulated by Heat Shock Protein 90 Rather than by Phosphorylation. PLoS ONE 2014, 9, e105479. [Google Scholar] [CrossRef]
- Förstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2012, 33, 829–837. [Google Scholar] [CrossRef] [PubMed]
- Sowa, G.; Pypaert, M.; Sessa, W.C. Distinction between signaling mechanisms in lipid rafts vs. caveolae. Proc. Natl. Acad. Sci. USA 2001, 98, 14072–14077. [Google Scholar] [CrossRef] [PubMed]
- Hecker, M.; Mülsch, A.; Bassenge, E.; Förstermann, U.; Busse, R. Subcellular localization and characterization of nitric oxide synthase(s) in endothelial cells: Physiological implications. Biochem. J. 1994, 299, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Andrew, P.J. Enzymatic function of nitric oxide synthases. Cardiovasc. Res. 1999, 43, 521–531. [Google Scholar] [CrossRef]
- Mattagajasingh, I.; Kim, C.-S.; Naqvi, A.; Yamamori, T.; Hoffman, T.A.; Jung, S.-B.; DeRicco, J.; Kasuno, K.; Irani, K. SIRT1 promotes endothelium-dependent vascular relaxation by activating endothelial nitric oxide synthase. Proc. Natl. Acad. Sci. USA 2007, 104, 14855–14860. [Google Scholar] [CrossRef]
- Zhang, Q.-J.; Wang, Z.; Chen, H.-Z.; Zhou, S.; Zheng, W.; Liu, G.; Wei, Y.-S.; Cai, H.; Liu, D.-P.; Liang, C.-C. Endothelium-specific overexpression of class III deacetylase SIRT1 decreases atherosclerosis in apolipoprotein E-deficient mice. Cardiovasc. Res. 2008, 80, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Fujiyoshi, M.; de Meijer, V.E.; Porte, R.J. Machine perfusion for donor organ repair: From vision to everyday clinical practice. In Organ Repair and Regeneration; Orlando, G., Shaf Keshavjee, Eds.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 43–73. [Google Scholar]
- Łuczak, A.; Madej, M.; Kasprzyk, A.; Doroszko, A. Role of the eNOS Uncoupling and the Nitric Oxide Metabolic Pathway in the Pathogenesis of Autoimmune Rheumatic Diseases. Oxidative Med. Cell. Longev. 2020, 2020, 1417981. [Google Scholar] [CrossRef]
- Rosenkranz-Weiss, P.; Sessa, W.C.; Milstien, S.; Kaufman, S.; A Watson, C.; Pober, J.S. Regulation of nitric oxide synthesis by proinflammatory cytokines in human umbilical vein endothelial cells. Elevations in tetrahydrobiopterin levels enhance endothelial nitric oxide synthase specific activity. J. Clin. Investig. 1994, 93, 2236–2243. [Google Scholar] [CrossRef]
- Li, H.; Förstermann, U. Uncoupling of ENOS in cardiovascular disease. In Nitric Oxide; Ignarro, L., Freeman, B., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 117–124. [Google Scholar]
- Rubanyi, G.M.; Vanhoutte, P.M. Superoxide anions and hyperoxia inactivate endothelium-derived relaxing factor. Am. J. Physiol. Circ. Physiol. 1986, 250, H822–H827. [Google Scholar] [CrossRef]
- Panth, N.; Paudel, K.R.; Parajuli, K. Reactive Oxygen Species: A Key Hallmark of Cardiovascular Disease. Adv. Med. 2016, 2016, 9152732. [Google Scholar] [CrossRef]
- Drummond, G.R.; Sobey, C.G. Endothelial NADPH Oxidases: Which NOX to Target in Vascular Disease? Trends Endocrinol. Metab. 2014, 25, 452–463. [Google Scholar] [CrossRef] [PubMed]
- Garrido, A.M.; Griendling, K.K. NADPH oxidases and angiotensin II receptor signaling. Mol. Cell. Endocrinol. 2009, 302, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Griendling, K.K.; Minieri, C.A.; Ollerenshaw, J.D.; Alexander, R.W. Angiotensin II stimulates NADH and NADPH oxidase activity in cultured vascular smooth muscle cells. Circ. Res. 1994, 74, 1141–1148. [Google Scholar] [CrossRef] [PubMed]
- Sandoo, A.; Veldhuijzen Van Zanten, J.J.C.S.; Metsios, G.S.; Carroll, D.; Kitas, G.D. The Endothelium and Its Role in Regulating Vascular Tone. Open Cardiovasc. Med. J. 2010, 4, 302–312. [Google Scholar] [CrossRef]
- Mitchell, J.A.; Ali, F.; Bailey, L.; Moreno, L.; Harrington, L.S. Role of Nitric Oxide and Prostacyclin as Vasoactive Hormones Released by the Endothelium. Exp. Physiol. 2008, 93, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Kudo, I. Phospholipase A2. J. Biochem. 2002, 131, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.A.; Warner, T. COX isoforms in the cardiovascular system: Understanding the activities of non-steroidal anti-inflammatory drugs. Nat. Rev. Drug Discov. 2006, 5, 75–86. [Google Scholar] [CrossRef]
- Frölich, J.C. Prostacyclin in Hypertension. Z. Kardiol. 1992, 81, 303–309. [Google Scholar]
- Spieker, L.E.; Flammer, A.J.; Lüscher, T.F. The vascular endothelium in hypertension. Handb. Exp. Pharmacol. 2006, 176, 249–283. [Google Scholar]
- Xu, D.; Emoto, N.; Giaid, A.; Slaughter, C.; Kaw, S.; Dewit, D.; Yanagisawa, M. ECE-1: A membrane-bound metalloprotease that catalyzes the proteolytic activation of big endothelin-1. Cell 1994, 78, 473–485. [Google Scholar] [CrossRef]
- Groenendijk, B.C.W.; Van Der Heiden, K.; Hierck, B.P.; Poelmann, R.E. The Role of Shear Stress on ET-1, KLF2, and NOS-3 Expression in the Developing Cardiovascular System of Chicken Embryos in a Venous Ligation Model. Physiology 2007, 22, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Corder, R.; Carrier, M.; Khan, N.; Klemm, P.; Vane, J.R. Cytokine Regulation of Endothelin-1 Release from Bovine Aortic Endothelial Cells. J. Cardiovasc. Pharmacol. 1995, 26 (Suppl. 3), S56-8. [Google Scholar] [CrossRef] [PubMed]
- Marasciulo, M.M.A.M.A.P.F.L.; Montagnani, M.; Potenza, M.A. Endothelin-1: The Yin and Yang on Vascular Function. Curr. Med. Chem. 2006, 13, 1655–1665. [Google Scholar] [CrossRef]
- Opgenorth, T.J. Endothelin receptor antagonism. In Advances in Pharmacology; August, J.T., Anders, M.W., Murad, F., Coyle, J.T., Eds.; Academic Press: Cambridge, MA, USA, 1995; pp. 1–65. [Google Scholar]
- Duerrschmidt, N.; Wippich, N.; Goettsch, W.; Broemme, H.-J.; Morawietz, H. Endothelin-1 Induces NAD(P)H Oxidase in Human Endothelial Cells. Biochem. Biophys. Res. Commun. 2000, 269, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.M.; Khokhlova, N.; Sutliff, R.L.; Adams, J.W.; Disher, K.M.; Zhao, H.; Capecchi, M.R.; Corvol, P.; Bernstein, K.E. Mice Lacking Endothelial ACE. Hypertension 2003, 41, 313–321. [Google Scholar] [CrossRef]
- Heyde, I.; Oster, H. Differentiating external zeitgeber impact on peripheral circadian clock resetting. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Gerhart-Hines, Z.; Lazar, M.A. Circadian Metabolism in the Light of Evolution. Endocr. Rev. 2015, 36, 289–304. [Google Scholar] [CrossRef]
- Raphael, T.E.; Au, K.H.; Goldman, S.R. Is Is. Change 2009, 198–229. [Google Scholar] [CrossRef]
- Lewis, P.; Korf, H.; Kuffer, L.; Groß, J.V.; Erren, T.C. Exercise time cues (zeitgebers) for human circadian systems can foster health and improve performance: A systematic review. BMJ Open Sport Exerc. Med. 2018, 4, e000443. [Google Scholar] [CrossRef]
- Hannibal, J. Neurotransmitters of the retino-hypothalamic tract. Cell Tissue Res. 2002, 309, 73–88. [Google Scholar] [CrossRef]
- Arola-Arnal, A.; Cruz-Carrión, Á.; Torres-Fuentes, C.; Ávila-Román, J.; Aragonès, G.; Mulero, M.; Bravo, F.I.; Muguerza, B.; Arola, L.; Suárez, M. Chrononutrition and Polyphenols: Roles and Diseases. Nutrients 2019, 11, 2602. [Google Scholar] [CrossRef] [PubMed]
- Golombek, D.A.; Bussi, I.L.; Agostino, P.V. Minutes, days and years: Molecular interactions among different scales of biological timing. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20120465. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.H.; McDearmon, E.L.; Panda, S.; Hayes, K.R.; Zhang, J.; Andrews, J.L.; Antoch, M.P.; Walker, J.R.; Esser, K.A.; Hogenesch, J.B.; et al. Circadian and CLOCK-controlled regulation of the mouse transcriptome and cell proliferation. Proc. Natl. Acad. Sci. USA 2007, 104, 3342–3347. [Google Scholar] [CrossRef]
- Latha, K.; Uhumwangho, M.; Sunil, S.; Srikanth, M.; Murthy, K.R. Chronobiology and Chronotherapy of Hypertension–A Review. Int. J. Health Res. 2010, 3, 121–131. [Google Scholar] [CrossRef][Green Version]
- Buurma, M.; Van Diemen, J.J.K.; Thijs, A.; Numans, M.E.; Bonten, T.N. Circadian Rhythm of Cardiovascular Disease: The Potential of Chronotherapy with Aspirin. Front. Cardiovasc. Med. 2019, 6, 84. [Google Scholar] [CrossRef]
- Hermida, R.C.; Ayala, D.E.; Fernández, J.R.; Portaluppi, F.; Fabbian, F.; Smolensky, M.H. Circadian Rhythms in Blood Pressure Regulation and Optimization of Hypertension Treatment with ACE Inhibitor and ARB Medications. Am. J. Hypertens. 2011, 24, 383–391. [Google Scholar] [CrossRef]
- Lemmer, B. The importance of circadian rhythms on drug response in hypertension and coronary heart disease—From mice and man. Pharmacol. Ther. 2006, 111, 629–651. [Google Scholar] [CrossRef]
- Hermida, R.C.; Ayala, D.E.; Portaluppi, F. Circadian variation of blood pressure: The basis for the chronotherapy of hypertension. Adv. Drug Deliv. Rev. 2007, 59, 904–922. [Google Scholar] [CrossRef]
- Elcik, D.; Kelesoglu, S.; Bing; Inan; Kalay, N.; Topsakal, R. Clinical effect of non-dipper and dipper hypertension with acute coronary syndrome patients. Erciyes Med. J. 2020, 43, 282. [Google Scholar] [CrossRef]
- Ohashi, N.; Isobe, S.; Ishigaki, S.; Yasuda, H. Circadian rhythm of blood pressure and the renin–angiotensin system in the kidney. Hypertens. Res. 2017, 40, 413–422. [Google Scholar] [CrossRef]
- Costello, H.M.; Gumz, M.L. Circadian Rhythm, Clock Genes, and Hypertension: Recent Advances in Hypertension. Hypertension 2021, 78, 1185–1196. [Google Scholar] [CrossRef] [PubMed]
- Douma, L.G.; Gumz, M.L. Circadian clock-mediated regulation of blood pressure. Free Radic. Biol. Med. 2018, 119, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Manfredini, R.; Fabbian, F.; Cappadona, R.; Modesti, P.A. Daylight saving time, circadian rhythms, and cardiovascular health. Intern. Emerg. Med. 2018, 13, 641–646. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Pollock, D.M. Diurnal Regulation of Renal Electrolyte Excretion: The Role of Paracrine Factors. Annu. Rev. Physiol. 2020, 82, 343–363. [Google Scholar] [CrossRef]
- Curtis, A.M.; Cheng, Y.; Kapoor, S.; Reilly, D.; Price, T.S.; FitzGerald, G.A. Circadian variation of blood pressure and the vascular response to asynchronous stress. Proc. Natl. Acad. Sci. USA 2007, 104, 3450–3455. [Google Scholar] [CrossRef]
- Crislip, G.R.; Douma, L.G.; Masten, S.H.; Cheng, K.-Y.; Lynch, I.J.; Johnston, J.G.; Barral, D.; Glasford, K.B.; Holzworth, M.R.; Verlander, J.W.; et al. Differences in renal BMAL1 contribution to Na+ homeostasis and blood pressure control in male and female mice. Am. J. Physiol. Physiol. 2020, 318, F1463–F1477. [Google Scholar] [CrossRef]
- Witte, K.; Schnecko, A.; Buijs, R.M.; van der Vliet, J.; Scalbert, E.; Delagrange, P.; Guardiola-Lemaitre, B.; Lemmer, B. Effects of Scn Lesions on Orcadian Blood Pressure Rhythm in Normotensive and Transgenic Hypertensive Rats. Chrono. Int. 1998, 15, 135–145. [Google Scholar] [CrossRef]
- Xie, Z.; Su, W.; Liu, S.; Zhao, G.; Esser, K.; Schroder, E.A.; Lefta, M.; Stauss, H.; Guo, Z.; Gong, M.C. Smooth-muscle BMAL1 participates in blood pressure circadian rhythm regulation. J. Clin. Investig. 2015, 125, 324–336. [Google Scholar] [CrossRef]
- Anea, C.B.; Cheng, B.; Sharma, S.; Kumar, S.; Caldwell, R.W.; Yao, L.; Ali, M.I.; Merloiu, A.M.; Stepp, D.W.; Black, S.M.; et al. Increased Superoxide and Endothelial NO Synthase Uncoupling in Blood Vessels of Bmal1-Knockout Mice. Circ. Res. 2012, 111, 1157–1165. [Google Scholar] [CrossRef]
- Zuber, A.M.; Centeno, G.; Pradervand, S.; Nikolaeva, S.; Maquelin, L.; Cardinaux, L.; Bonny, O.; Firsov, D. Molecular clock is involved in predictive circadian adjustment of renal function. Proc. Natl. Acad. Sci. USA 2009, 106, 16523–16528. [Google Scholar] [CrossRef]
- Stow, L.R.; Richards, J.; Cheng, K.-Y.; Lynch, I.J.; Jeffers, L.; Greenlee, M.M.; Cain, B.D.; Wingo, C.S.; Gumz, M.L. The Circadian Protein Period 1 Contributes to Blood Pressure Control and Coordinately Regulates Renal Sodium Transport Genes. Hypertension 2012, 59, 1151–1156. [Google Scholar] [CrossRef]
- Solocinski, K.; Holzworth, M.; Wen, X.; Cheng, K.-Y.; Lynch, I.J.; Cain, B.D.; Wingo, C.S.; Gumz, M.L. Desoxycorticosterone pivalate-salt treatment leads to non-dipping hypertension in Per1 knockout mice. Acta Physiol. 2017, 220, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Douma, L.G.; Solocinski, K.; Holzworth, M.R.; Crislip, G.R.; Masten, S.H.; Miller, A.H.; Cheng, K.-Y.; Lynch, I.J.; Cain, B.D.; Wingo, C.S.; et al. Female C57BL/6J mice lacking the circadian clock protein PER1 are protected from nondipping hypertension. Am. J. Physiol. Integr. Comp. Physiol. 2019, 316, R50–R58. [Google Scholar] [CrossRef]
- Viswambharan, H.; Carvas, J.M.; Antic, V.; Marecic, A.; Jud, C.; Zaugg, C.E.; Ming, X.-F.; Montani, J.-P.; Albrecht, U.; Yang, Z. Mutation of the Circadian Clock Gene Per2 Alters Vascular Endothelial Function. Circulation 2007, 115, 2188–2195. [Google Scholar] [CrossRef]
- Vukolic, A.; Antic, V.; Van Vliet, B.N.; Yang, Z.; Albrecht, U.; Montani, J.-P. Role of mutation of the circadian clock gene Per2 in cardiovascular circadian rhythms. Am. J. Physiol. Integr. Comp. Physiol. 2010, 298, R627–R634. [Google Scholar] [CrossRef]
- Doi, M.; Takahashi, Y.; Komatsu, R.; Yamazaki, F.; Yamada, H.; Haraguchi, S.; Emoto, N.; Okuno, Y.; Tsujimoto, G.; Kanematsu, A.; et al. Salt-sensitive hypertension in circadian clock–deficient Cry-null mice involves dysregulated adrenal Hsd3b6. Nat. Med. 2010, 16, 67–74. [Google Scholar] [CrossRef]
- Kawasaki, T.; Cugini, P.; Uezono, K.; Sasaki, H.; Itoh, K.; Nishiura, M.; Shinkawa, K. Circadian Variations of Total Renin, Active Renin, Plasma Renin Activity and Plasma Aldosterone in Clinically Healthy Young Subjects. Horm. Metab. Res. 1990, 22, 636–639. [Google Scholar] [CrossRef] [PubMed]
- Richards, A.M.; Nicholls, M.G.; Espiner, E.A.; Ikram, H.; Cullens, M.; Hinton, D. Diurnal Patterns of Blood Pressure, Heart Rate and Vasoactive Hormones in Normal Man. Clin. Exp. Hypertens. Part A Theory Pr. 1986, 8, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Denniff, M.; Turrell, H.E.; Vanezis, A.; Rodrigo, G.C. The time-of-day variation in vascular smooth muscle contractility depends on a nitric oxide signalling pathway. J. Mol. Cell. Cardiol. 2014, 66, 133–140. [Google Scholar] [CrossRef]
- Shang, X.; Pati, P.; Anea, C.B.; Fulton, D.J.; Rudic, R.D. Differential Regulation of BMAL1, CLOCK, and Endothelial Signaling in the Aortic Arch and Ligated Common Carotid Artery. J. Vasc. Res. 2016, 53, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Lemmer, B.; Arraj, M. Effect of NO Synthase Inhibition on Cardiovascular Circadian Rhythms in Wild-Type and eNOS-Knock-Out Mice. Chrono. Int. 2008, 25, 501–510. [Google Scholar] [CrossRef]
- Arraj, M.; Lemmer, B. Endothelial Nitric Oxide Is Not Involved in Circadian Rhythm Generation of Blood Pressure: Experiments in Wild-Type C57 and eNOS Knock-Out Mice under Light-Dark and Free-Run Conditions. Chrono. Int. 2007, 24, 1231–1240. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo, G.C.; Herbert, K.E. Regulation of vascular function and blood pressure by circadian variation in redox signalling. Free Radic. Biol. Med. 2018, 119, 115–120. [Google Scholar] [CrossRef]
- Anea, C.B.; Zhang, M.; Chen, F.; Ali, M.I.; Hart, C.M.M.; Stepp, D.W.; Kovalenkov, Y.; Merloiu, A.-M.; Pati, P.; Fulton, D.; et al. Circadian Clock Control of Nox4 and Reactive Oxygen Species in the Vasculature. PLoS ONE 2013, 8, e78626. [Google Scholar] [CrossRef]
- Hwang, Y.-S.; Hsieh, T.-J.; Lee, Y.-J.; Tsai, J.-H. Circadian rhythm of urinary endothelin-1 excretion in mild hypertensive patients. Am. J. Hypertens. 1998, 11, 1344–1351. [Google Scholar] [CrossRef][Green Version]
- Douma, L.G.; Barral, D.; Gumz, M.L. Interplay of the Circadian Clock and Endothelin System. Physiology 2021, 36, 35–43. [Google Scholar] [CrossRef]
- Mancia, G.; Zanchetti, A. Choice of antihypertensive drugs in the European Society of Hypertension–European Society of Cardiology guidelines: Specific indications rather than ranking for general usage. J. Hypertens. 2008, 26, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Mc Namara, K.; Alzubaidi, H.; Jackson, J.K. Cardiovascular disease as a leading cause of death: How are pharmacists getting involved? Integr. Pharm. Res. Pr. 2019, 8, 1–11. [Google Scholar] [CrossRef]
- Blacher, J.; Kretz, S.; Sorbets, E.; Lelong, H.; Vallée, A.; Lopez-Sublet, M. Épidémiologie de l’HTA: Différences femme/homme. Press. Med. 2019, 48, 1240–1243. [Google Scholar] [CrossRef]
- Liu, C.; Huang, Y. Chinese Herbal Medicine on Cardiovascular Diseases and the Mechanisms of Action. Front. Pharmacol. 2016, 7, 469. [Google Scholar] [CrossRef]
- Tabassum, N.; Ahmad, F. Role of natural herbs in the treatment of hypertension. Pharmacogn. Rev. 2011, 5, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Elliott, W.J. Systemic Hypertension. Curr. Probl. Cardiol. 2007, 32, 201–259. [Google Scholar] [CrossRef] [PubMed]
- Guzik, T.J.; Touyz, R.M. Oxidative Stress, Inflammation, and Vascular Aging in Hypertension. Hypertension 2017, 70, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Masi, S.; Uliana, M.; Virdis, A. Angiotensin II and vascular damage in hypertension: Role of oxidative stress and sympathetic activation. Vasc. Pharmacol. 2019, 115, 13–17. [Google Scholar] [CrossRef]
- Cannizzaro, E.; Cirrincione, L.; Mazzucco, W.; Scorciapino, A.; Catalano, C.; Ramaci, T.; Ledda, C.; Plescia, F. Night-Time Shift Work and Related Stress Responses: A Study on Security Guards. Int. J. Environ. Res. Public Health 2020, 17, 562. [Google Scholar] [CrossRef]
- Depner, C.M.; Stothard, E.R.; Wright, K.P., Jr. Metabolic Consequences of Sleep and Circadian Disorders. Curr. Diabetes Rep. 2014, 14, 507. [Google Scholar] [CrossRef]
- Torres-Fuentes, C.; Suárez, M.; Aragonès, G.; Mulero, M.; Ávila-Román, J.; Arola-Arnal, A.; Salvadó, M.J.; Arola, L.; Bravo, F.I.; Muguerza, B. Cardioprotective properties of phenolic compounds: A role for biological rhythms. Mol. Nutr. Food Res. 2022, 2100990. [Google Scholar] [CrossRef]
- Ghatage, T.; Goyal, S.G.; Dhar, A.; Bhat, A. Novel therapeutics for the treatment of hypertension and its associated complications: Peptide- and nonpeptide-based strategies. Hypertens. Res. 2021, 44, 740–755. [Google Scholar] [CrossRef]
- Sayarlioglu, H. Rational approaches to the treatment of hypertension: Modification of lifestyle measures. Kidney Int. Suppl. 2013, 3, 346–348. [Google Scholar] [CrossRef]
- Huang, W.-Y.; Davidge, S.T.; Wu, J. Bioactive Natural Constituents from Food Sources—Potential Use in Hypertension Prevention and Treatment. Crit. Rev. Food Sci. Nutr. 2013, 53, 615–630. [Google Scholar] [CrossRef]
- Bhatti, M.S.; Alseud, K.M.; Uddin, J.; El-Seedi, H.R.; Musharraf, S.G. Repurposing of Pharmaceutical Drugs by High-throughput Approach for Antihypertensive Activity as Inhibitors of Angiotensin-Converting Enzyme (ACE) Using HPLC-ESI-MS/MS Method. Arab. J. Chem. 2021, 14, 103279. [Google Scholar] [CrossRef]
- Hermida, R.C.; Ayala, D.E.; Mojón-Ojea, A.; Fernández, J.R. Influence of circadian time of hypertension treatment on cardiovascular risk: Results of the MAPEC study. Chrono. Int. 2010, 27, 1629–1651. [Google Scholar] [CrossRef] [PubMed]
- Frishman, W.H. Beta-Adrenergic Receptor Blockers in Hypertension: Alive and Well. Prog. Cardiovasc. Dis. 2016, 59, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Tocci, G.; Battistoni, A.; Passerini, J.; Musumeci, M.B.; Francia, P.; Ferrucci, A.; Volpe, M. Calcium Channel Blockers and Hypertension. J. Cardiovasc. Pharmacol. Ther. 2015, 20, 121–130. [Google Scholar] [CrossRef]
- Vazir, A.; Sundaram, V.; Harper, A.R. Diuretic therapy. In Encyclopedia of Cardiovascular Research and Medicine; Vasan, R.S., Sawyer, D.B., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 107–116. [Google Scholar]
- Oparil, S.; Schmieder, R.E. New Approaches in the Treatment of Hypertension. Circ. Res. 2015, 116, 1074–1095. [Google Scholar] [CrossRef] [PubMed]
- Feyh, A.; Bracero, L.; Lakhani, H.V.; Santhanam, P.; I Shapiro, J.; Khitan, Z.; Sodhi, K. Role of Dietary Components in Modulating Hypertension. J. Clin. Exp. Cardiol. 2016, 7, 1–7. [Google Scholar] [CrossRef]
- Siervo, M.; Lara, J.; Chowdhury, S.; Ashor, A.; Oggioni, C.; Mathers, J.C. Effects of the Dietary Approach to Stop Hypertension (DASH) diet on cardiovascular risk factors: A systematic review and meta-analysis. Br. J. Nutr. 2015, 113, 1–15. [Google Scholar] [CrossRef]
- Kaur, S.; Das, M. Functional foods: An overview. Food Sci. Biotechnol. 2011, 20, 861–875. [Google Scholar] [CrossRef]
- López-Fernández-Sobrino, R.; Soliz-Rueda, J.; Margalef, M.; Arola-Arnal, A.; Suárez, M.; Bravo, F.; Muguerza, B. ACE Inhibitory and Antihypertensive Activities of Wine Lees and Relationship among Bioactivity and Phenolic Profile. Nutrients 2021, 13, 679. [Google Scholar] [CrossRef]
- Morales, D.; Miguel, M.; Garcés-Rimón, M. Pseudocereals: A novel source of biologically active peptides. Crit. Rev. Food Sci. Nutr. 2021, 61, 1537–1544. [Google Scholar] [CrossRef]
- Shobako, N. Hypotensive peptides derived from plant proteins. Peptides 2021, 142, 170573. [Google Scholar] [CrossRef] [PubMed]
- Sosalagere, C.; Kehinde, B.A.; Sharma, P. Isolation and functionalities of bioactive peptides from fruits and vege16s: A reviews. Food Chem. 2021, 366, 130494. [Google Scholar] [CrossRef] [PubMed]
- Bravo, F.I.; Mas-Capdevila, A.; López-Fernández-Sobrino, R.; Torres-Fuentes, C.; Mulero, M.; Alcaide-Hidalgo, J.M.; Muguerza, B. Identification of novel antihypertensive peptides from wine lees hydrolysate. Food Chem. 2021, 366, 130690. [Google Scholar] [CrossRef] [PubMed]
- Mora, L.; Gallego, M.; Toldrá, F. ACEI-Inhibitory Peptides Naturally Generated in Meat and Meat Products and Their Health Relevance. Nutrients 2018, 10, 1259. [Google Scholar] [CrossRef] [PubMed]
- Morales, D.; Piris, A.J.; Ruiz-Rodriguez, A.; Prodanov, M.; Soler-Rivas, C. Extraction of bioactive compounds against cardiovascular diseases from Lentinula edodes using a sequential extraction method. Biotechnol. Prog. 2018, 34, 746–755. [Google Scholar] [CrossRef]
- Sultan, S.; Huma, N.; Butt, M.S.; Aleem, M.; Abbas, M. Therapeutic potential of dairy bioactive peptides: A contemporary perspective. Crit. Rev. Food Sci. Nutr. 2018, 58, 105–115. [Google Scholar] [CrossRef]
- Rathnapala, E.C.N.; Ahn, D.U.; Abeyrathne, S. Functional properties of ovotransferrin from chicken egg white and its derived peptides: A review. Food Sci. Biotechnol. 2021, 30, 619–630. [Google Scholar] [CrossRef]
- Mas-Capdevila, A.; Pons, Z.; Aleixandre, A.; Bravo, F.I.; Muguerza, B. Dose-Related Antihypertensive Properties and the Corresponding Mechanisms of a Chicken Foot Hydrolysate in Hypertensive Rats. Nutrients 2018, 10, 1295. [Google Scholar] [CrossRef]
- Bravo, F.I.; Mas-Capdevila, A.; Margalef, M.; Arola-Arnal, A.; Muguerza, B. Novel Antihypertensive Peptides Derived from Chicken Foot Proteins. Mol. Nutr. Food Res. 2019, 63, e1801176. [Google Scholar] [CrossRef]
- Paiva, L.; Lima, E.; Neto, A.; Baptista, J. Angiotensin I-Converting Enzyme (ACE) Inhibitory Activity, Antioxidant Properties, Phenolic Content and Amino Acid Profiles of Fucus spiralis L. Protein Hydrolysate Fractions. Mar. Drugs 2017, 15, 311. [Google Scholar] [CrossRef]
- Alcaide-Hidalgo, J.M.; Margalef, M.; Bravo, F.I.; Muguerza, B.; López-Huertas, E. Virgin olive oil (unfiltered) extract contains peptides and possesses ACE inhibitory and antihypertensive activity. Clin. Nutr. 2019, 39, 1242–1249. [Google Scholar] [CrossRef] [PubMed]
- Durak, M.Z. Antihypertensive Peptides in Dairy Products. Am. J. Biomed. Sci. Res. 2020, 7, 191–195. [Google Scholar] [CrossRef]
- Alcaide-Hidalgo, J.M.; Romero, M.; Duarte, J.; López-Huertas, E. Antihypertensive Effects of Virgin Olive Oil (Unfiltered) Low Molecular Weight Peptides with ACE Inhibitory Activity in Spontaneously Hypertensive Rats. Nutrients 2020, 12, 271. [Google Scholar] [CrossRef] [PubMed]
- Al Shukor, N.; Van Camp, J.; Gonzales, G.B.; Staljanssens, D.; Struijs, K.; Zotti, M.J.; Raes, K.; Smagghe, G. Angiotensin-Converting Enzyme Inhibitory Effects by Plant Phenolic Compounds: A Study of Structure Activity Relationships. J. Agric. Food Chem. 2013, 61, 11832–11839. [Google Scholar] [CrossRef]
- Guerrero, L.; Castillo, J.; Quiñones, M.; Garcia-Vallvé, S.; Arola, L.; Pujadas, G.; Muguerza, B. Inhibition of Angiotensin-Converting Enzyme Activity by Flavonoids: Structure-Activity Relationship Studies. PLoS ONE 2012, 7, e49493. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Pechan, T.; Chang, S.K. Antioxidant and angiotensin-I converting enzyme inhibitory activities of phenolic extracts and fractions derived from three phenolic-rich legume varieties. J. Funct. Foods 2018, 42, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Ra, J.-E.; Woo, S.-Y.; Jin, H.; Lee, M.J.; Kim, H.Y.; Ham, H.; Chung, I.-M.; Seo, W.D. Evaluation of antihypertensive polyphenols of barley (Hordeum vulgare L.) seedlings via their effects on angiotensin-converting enzyme (ACE) inhibition. Appl. Biol. Chem. 2020, 63, 38. [Google Scholar] [CrossRef]
- Paiva, L.; Lima, E.; Neto, A.; Baptista, J. Angiotensin I-converting enzyme (ACE) inhibitory activity of Fucus spiralis macroalgae and influence of the extracts storage temperature—A short report. J. Pharm. Biomed. Anal. 2016, 131, 503–507. [Google Scholar] [CrossRef]
- Margalef, M.; Bravo, F.I.; Muguerza, B.; Arola-Arnal, A. Natural Angiotensin Converting Enzyme (ACE) inhibitors with antihypertensive properties. In Natural Products Targeting Clinically Relevant Enzymes; Wiley: Hoboken, NJ, USA, 2017; pp. 45–67. [Google Scholar]
- Fitzgerald, C.; Mora-Soler, L.; Gallagher, E.; O’Connor, P.; Prieto, J.; Soler-Vila, A.; Hayes, M. Isolation and Characterization of Bioactive Pro-Peptides with in Vitro Renin Inhibitory Activities from the Macroalga Palmaria palmata. J. Agric. Food Chem. 2012, 60, 7421–7427. [Google Scholar] [CrossRef] [PubMed]
- Ciau-Solís, N.A.; Acevedo-Fernández, J.J.; Betancur-Ancona, D. In Vitro renin–angiotensin system inhibition and In Vivo antihypertensive activity of peptide fractions from lima bean (Phaseolus lunatus L.). J. Sci. Food Agric. 2018, 98, 781–786. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Hori, K.; Shinbo, M.; Hiwatashi, K.; Gotoh, T.; Yamada, S. Isolation of Human Renin Inhibitor from Soybean: Soyasaponin I Is the Novel Human Renin Inhibitor in Soybean. Biosci. Biotechnol. Biochem. 2008, 72, 3232–3236. [Google Scholar] [CrossRef] [PubMed]
- Udenigwe, C.C.; Adebiyi, A.P.; Doyen, A.; Li, H.; Bazinet, L.; Aluko, R.E. Low molecular weight flaxseed protein-derived arginine-containing peptides reduced blood pressure of spontaneously hypertensive rats faster than amino acid form of arginine and native flaxseed protein. Food Chem. 2012, 132, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Li, X.; Liu, X.; Yang, W.; Li, M.; Li, J.; Li, F. Aqueous extracts of tree peony petals: Renin and angiotensin I-converting enzyme inhibitory activities in different colours and flowering stages. RSC Adv. 2022, 12, 7735–7741. [Google Scholar] [CrossRef] [PubMed]
- Ismail, W.M.; Ezzat, S.M.; E Michel, H.; El Deeb, K.S.; El-Fishawy, A.M. Angiotensin-converting enzyme and renin inhibition activities, antioxidant properties, phenolic and flavonoid contents of Cuphea ignea A. DC. J. Rep. Pharm. Sci. 2020, 9, 92. [Google Scholar] [CrossRef]
- Salem, M.A.; Ezzat, S.M.; Ahmed, K.A.; Alseekh, S.; Fernie, A.R.; Essam, R.M. A Comparative Study of the Antihypertensive and Cardioprotective Potentials of Hot and Cold Aqueous Extracts of Hibiscus sabdariffa L. in Relation to Their Metabolic Profiles. Front. Pharmacol. 2022, 13. [Google Scholar] [CrossRef]
- Mas-Capdevila, A.; Iglesias-Carres, L.; Arola-Arnal, A.; Suarez, M.; Muguerza, B.; Bravo, F.I. Long-term administration of protein hydrolysate from chicken feet induces antihypertensive effect and confers vasoprotective pattern in diet-induced hypertensive rats. J. Funct. Foods 2019, 55, 28–35. [Google Scholar] [CrossRef]
- Mas-Capdevila, A.; Iglesias-Carres, L.; Arola-Arnal, A.; Aragonès, G.; Aleixandre, A.; Bravo, F.I.; Muguerza, B. Evidence that Nitric Oxide is Involved in the Blood Pressure Lowering Effect of the Peptide AVFQHNCQE in Spontaneously Hypertensive Rats. Nutrients 2019, 11, 225. [Google Scholar] [CrossRef]
- Tonolo, F.; Folda, A.; Cesaro, L.; Scalcon, V.; Marin, O.; Ferro, S.; Bindoli, A.; Rigobello, M.P. Milk-derived bioactive peptides exhibit antioxidant activity through the Keap1-Nrf2 signaling pathway. J. Funct. Foods 2020, 64, 103696. [Google Scholar] [CrossRef]
- Tonolo, F.; Sandre, M.; Ferro, S.; Folda, A.; Scalcon, V.; Scutari, G.; Feller, E.; Marin, O.; Bindoli, A.; Rigobello, M.P. Milk-derived bioactive peptides protect against oxidative stress in a Caco-2 cell model. Food Funct. 2018, 9, 1245–1253. [Google Scholar] [CrossRef]
- Aires, R.; Amorim, F.G.; Côco, L.Z.; da Conceição, A.P.; Zanardo, T.C.; Taufner, G.H.; Nogueira, B.V.; Vasquez, E.C.; Pereira, T.M.C.; Campagnaro, B.P.P.; et al. Use of kefir peptide (Kef-1) as an emerging approach for the treatment of oxidative stress and inflammation in 2K1C mice. Food Funct. 2022, 13, 1965–1974. [Google Scholar] [CrossRef]
- López-Fernández-Sobrino, R.; Soliz-Rueda, J.; Suárez, M.; Mulero, M.; Arola, L.; Bravo, F.; Muguerza, B. Blood Pressure-Lowering Effect of Wine Lees: Dose-Response Study, Effect of Dealcoholization and Possible Mechanisms of Action. Nutrients 2021, 13, 1142. [Google Scholar] [CrossRef]
- López-Fernández-Sobrino, R.; Soliz-Rueda, J.; Ávila-Román, J.; Arola-Arnal, A.; Suárez, M.; Muguerza, B.; Bravo, F. Blood Pressure-Lowering Effect of Wine Lees Phenolic Compounds Is Mediated by Endothelial-Derived Factors: Role of Sirtuin 1. Antioxidants 2021, 10, 1073. [Google Scholar] [CrossRef] [PubMed]
- Mas-Capdevila, A.; Iglesias-Carres, L.; Arola-Arnal, A.; Suárez, M.; Bravo, F.I.; Muguerza, B. Changes in arterial blood pressure caused by long-term administration of grape seed proanthocyanidins in rats with established hypertension. Food Funct. 2020, 11, 8735–8742. [Google Scholar] [CrossRef] [PubMed]
- Pons, Z.; Margalef, M.; Bravo, F.; Arola-Arnal, A.; Muguerza, B. Grape seed flavanols decrease blood pressure via Sirt-1 and confer a vasoprotective pattern in rats. J. Funct. Foods 2016, 24, 164–172. [Google Scholar] [CrossRef]
- Yousefian, M.; Shakour, N.; Hosseinzadeh, H.; Hayes, A.W.; Hadizadeh, F.; Karimi, G. The natural phenolic compounds as modulators of NADPH oxidases in hypertension. Phytomedicine 2019, 55, 200–213. [Google Scholar] [CrossRef]
- Pap, N.; Fidelis, M.; Azevedo, L.; do Carmo, M.A.V.; Wang, D.; Mocan, A.; Pereira, E.P.R.; Xavier-Santos, D.; Sant’Ana, A.S.; Yang, B.; et al. Berry polyphenols and human health: Evidence of antioxidant, anti-inflammatory, microbiota modulation, and cell-protecting effects. Curr. Opin. Food Sci. 2021, 42, 167–186. [Google Scholar] [CrossRef]
- Figueira, J.; Porto-Figueira, P.; Berenguer, C.; Pereira, J.; Câmara, J. Evaluation of the Health-Promoting Properties of Selected Fruits. Molecules 2021, 26, 4202. [Google Scholar] [CrossRef]
- Yan, Z.; Zhong, Y.; Duan, Y.; Chen, Q.; Li, F. Antioxidant mechanism of tea polyphenols and its impact on health benefits. Anim. Nutr. 2020, 6, 115–123. [Google Scholar] [CrossRef]
- Klippel, B.F.; Duemke, L.B.; Leal, M.A.; Friques, A.G.F.; Dantas, E.M.; Dalvi, R.F.; Gava, A.L.; Pereira, T.M.C.; Andrade, T.U.; Meyrelles, S.S.; et al. Effects of Kefir on the Cardiac Autonomic Tones and Baroreflex Sensitivity in Spontaneously Hypertensive Rats. Front. Physiol. 2016, 7, 211. [Google Scholar] [CrossRef]
- Friques, A.G.F.; Arpini, C.; Kalil, I.C.; Gava, A.L.; Leal, M.A.; Porto, M.L.; Nogueira, B.V.; Dias, A.T.; Andrade, T.U.; Pereira, T.M.C.; et al. Chronic administration of the probiotic kefir improves the endothelial function in spontaneously hypertensive rats. J. Transl. Med. 2015, 13, 1–16. [Google Scholar] [CrossRef]
- Robles-Vera, I.; Toral, M.; De La Visitación, N.; Sánchez, M.; Gómez-Guzmán, M.; Romero, M.; Yang, T.; Izquierdo-Garcia, J.L.; Jiménez, R.; Ruiz-Cabello, J.; et al. Probiotics Prevent Dysbiosis and the Rise in Blood Pressure in Genetic Hypertension: Role of Short-Chain Fatty Acids. Mol. Nutr. Food Res. 2020, 64, e1900616. [Google Scholar] [CrossRef] [PubMed]
- Rashid, S.K.; Khodja, N.I.; Auger, C.; Alhosin, M.; Boehm, N.; Oswald-Mammosser, M.; Schini-Kerth, V.B. Probiotics (VSL#3) Prevent Endothelial Dysfunction in Rats with Portal Hypertension: Role of the Angiotensin System. PLoS ONE 2014, 9, e97458. [Google Scholar] [CrossRef]
- Toral, M.; Gómez-Guzmán, M.; Jiménez, R.; Romero, M.; Sánchez, M.; Utrilla, M.P.; Garrido-Mesa, N.; Rodríguez-Cabezas, M.E.; Olivares, M.; Gálvez, J.; et al. The probiotic Lactobacillus coryniformis CECT5711 reduces the vascular pro-oxidant and pro-inflammatory status in obese mice. Clin. Sci. 2014, 127, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Lemmer, B. Chronopharmacology and its impact on antihypertensive treatment. Acta Physiol. Pharmacol. Bulg. 1999, 24, 71–80. [Google Scholar]
- de la Sierra, A.; Redon, J.; Banegas, J.R.; Segura, J.; Parati, G.; Gorostidi, M.; de la Cruz, J.J.; Sobrino, J.; Llisterri, J.L.; Alonso, J.; et al. Prevalence and Factors Associated with Circadian Blood Pressure Patterns in Hypertensive Patients. Hypertension 2009, 53, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Potucek, P.; Klimas, J. Chronopharmacology of high blood pressure—A critical review of clinical evidence. Eur. Pharm. J. 2019, 66, 32–36. [Google Scholar] [CrossRef]
- Hermida, R.C.; Ayala, D.E.; Smolensky, M.H.; Fernandez, J.R.; Mojón, A.; Portaluppi, F. Chronotherapy with conventional blood pressure medications improves management of hypertension and reduces cardiovascular and stroke risks. Hypertens. Res. 2016, 39, 277–292. [Google Scholar] [CrossRef]
- Kohno, I.; Ijiri, H.; Takusagawa, M.; Yin, D.; Sano, S.; Ishihara, T.; Sawanobori, T.; Komori, S.; Tamura, K. Effect of Imidapril in Dipper and Nondipper Hypertensive Patients: Comparison Between Morning and Evening Administration. Chrono. Int. 2000, 17, 209–219. [Google Scholar] [CrossRef]
- Hermida, R.C.; Ayala, D.E.; Chayán, L.; Mojón-Ojea, A.; Fernández, J.R. Administration-Time-Dependent Effects of Olmesartan on the Ambulatory Blood Pressure of Essential Hypertension Patients. Chrono. Int. 2009, 26, 61–79. [Google Scholar] [CrossRef]
- Smolensky, M.H.; Hermida, R.C.; Ayala, D.E.; Tiseo, R.; Portaluppi, F. Administration–time-dependent effects of blood pressure-lowering medications: Basis for the chronotherapy of hypertension. Blood Press. Monit. 2010, 15, 173–180. [Google Scholar] [CrossRef]
- Hermida, R.C.; Ayala, D.E.; Mojón, A.; Chayán, L.; Dominguez, M.J.; Fontao, M.J.; Soler, R.; Alonso, I.; Fernandez, J.R.; Mojón-Ojea, A. Comparison of the Effects on Ambulatory Blood Pressure of Awakening versus Bedtime Administration of Torasemide in Essential Hypertension. Chrono. Int. 2008, 25, 950–970. [Google Scholar] [CrossRef] [PubMed]
- Hermida, R.C.; Calvo, C.; Ayala, D.E.; Lopez, J.E.; Covelo, M. Administration time-dependent effects of nebivolol on ambulatory blood pressure in patients with essential hypertension. Am. J. Hypertens. 2006, 18, A63. [Google Scholar] [CrossRef][Green Version]
- Potter, G.D.M.; Cade, J.; Grant, P.J.; Hardie, L.J. Nutrition and the circadian system. Br. J. Nutr. 2016, 116, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Ávila-Román, J.; Soliz-Rueda, J.R.; Bravo, F.I.; Aragonès, G.; Suárez, M.; Arola-Arnal, A.; Mulero, M.; Salvadó, M.-J.; Arola, L.; Torres-Fuentes, C.; et al. Phenolic compounds and biological rhythms: Who takes the lead? Trends Food Sci. Technol. 2021, 113, 77–85. [Google Scholar] [CrossRef]
- Gadacha, W.; Ben-Attia, M.; Bonnefont-Rousselot, D.; Aouani, E.; Ghanem-Boughanmi, N.; Touitou, Y. Resveratrol opposite effects on rat tissue lipoperoxidation: Pro-oxidant during day-time and antioxidant at night. Redox Rep. 2009, 14, 154–158. [Google Scholar] [CrossRef] [PubMed]
- Ghiadoni, L.; Magagna, A.; Versari, D.; Kardasz, I.; Huang, Y.; Taddei, S.; Salvetti, A. Different Effect of Antihypertensive Drugs on Conduit Artery Endothelial Function. Hypertension 2003, 41, 1281–1286. [Google Scholar] [CrossRef]
- Ribas-Latre, A.; Baselga-Escudero, L.; Casanova, E.; Arola-Arnal, A.; Salvado, J.; Bladé, C.; Arola, L. Dietary proanthocyanidins modulate BMAL1 acetylation, Nampt expression and NAD levels in rat liver. Sci. Rep. 2015, 5, 10954. [Google Scholar] [CrossRef]
- Potenza, M.A.; Marasciulo, F.L.; Tarquinio, M.; Tiravanti, E.; Colantuono, G.; Federici, A.; Kim, J.-A.; Quon, M.J.; Montagnani, M. EGCG, a green tea polyphenol, improves endothelial function and insulin sensitivity, reduces blood pressure, and protects against myocardial I/R injury in SHR. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E1378–E1387. [Google Scholar] [CrossRef]
- Xu, R.; Yang, K.; Ding, J.; Chen, G. Effect of green tea supplementation on blood pressure. Medicine 2020, 99, e19047. [Google Scholar] [CrossRef]
- Mi, Y.; Qi, G.; Fan, R.; Ji, X.; Liu, Z.; Liu, X. EGCG ameliorates diet-induced metabolic syndrome associating with the circadian clock. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2017, 1863, 1575–1589. [Google Scholar] [CrossRef]
- Hironao, K.-Y.; Mitsuhashi, Y.; Huang, S.; Oike, H.; Ashida, H.; Yamashita, Y. Cacao polyphenols regulate the circadian clock gene expression and through glucagon-like peptide-1 secretion. J. Clin. Biochem. Nutr. 2020, 67, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Hironao, K.-Y.; Ashida, H.; Yamashita, Y. The cacao procyanidin extract-caused anti-hyperglycemic effect was changed by the administration timings. J. Clin. Biochem. Nutr. 2020, 67, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Arreaza-Gil, V.; Escobar-Martínez, I.; Suárez, M.; Bravo, F.I.; Muguerza, B.; Arola-Arnal, A.; Torres-Fuentes, C. Gut Seasons: Photoperiod Effects on Fecal Microbiota in Healthy and Cafeteria-Induced Obese Fisher 344 Rats. Nutrients 2022, 14, 722. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibarz-Blanch, N.; Morales, D.; Calvo, E.; Ros-Medina, L.; Muguerza, B.; Bravo, F.I.; Suárez, M. Role of Chrononutrition in the Antihypertensive Effects of Natural Bioactive Compounds. Nutrients 2022, 14, 1920. https://doi.org/10.3390/nu14091920
Ibarz-Blanch N, Morales D, Calvo E, Ros-Medina L, Muguerza B, Bravo FI, Suárez M. Role of Chrononutrition in the Antihypertensive Effects of Natural Bioactive Compounds. Nutrients. 2022; 14(9):1920. https://doi.org/10.3390/nu14091920
Chicago/Turabian StyleIbarz-Blanch, Néstor, Diego Morales, Enrique Calvo, Laura Ros-Medina, Begoña Muguerza, Francisca Isabel Bravo, and Manuel Suárez. 2022. "Role of Chrononutrition in the Antihypertensive Effects of Natural Bioactive Compounds" Nutrients 14, no. 9: 1920. https://doi.org/10.3390/nu14091920
APA StyleIbarz-Blanch, N., Morales, D., Calvo, E., Ros-Medina, L., Muguerza, B., Bravo, F. I., & Suárez, M. (2022). Role of Chrononutrition in the Antihypertensive Effects of Natural Bioactive Compounds. Nutrients, 14(9), 1920. https://doi.org/10.3390/nu14091920