
The Role of the Microbiome-Brain-Gut Axis in the Pathogenesis of Depressive Disorder

 , , ,
, , , 
Abstract
:1. Introduction
1.1. What Is Metagenome?
1.2. How Are the Brain and the Gut Communicating?
2. What Is the Gut Microbiota?
3. What Is Major Depressive Disorder (MDD)?
4. Does Depressive Disorder Cause Dysbiosis?
4.1. Stress and Gastrointestinal Disorders
4.2. Depressive State and Dysbiosis in Animal Models—Communication Routes
4.3. Which Alterations Are Observed in the Depressed Brain
4.4. Hypothalamic-Pituitary-Adrenal (HPA) Axis
4.5. Glucocorticoids and Suppression of the Inflammatory Response
4.6. Chronic Stress Affects the Inflammatory Response
4.7. Alterations of Intestinal Microbiota in Depressive State
5. Do Alterations in Gut Microbiota Lead to Depression?
5.1. What Are the Gut Peptides?
5.2. How Does the Microbiota Interact with the Secretion of Gut Peptides?
5.3. What Changes in the Composition of Gut Microbiota Might Cause Depressive Disorder?
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ding, R.X.; Goh, W.R.; Wu, R.N.; Yue, X.Q.; Luo, X.; Khine, W.; Wu, J.R.; Lee, Y.K. Revisit gut microbiota and its impact on human health and disease. J. Food Drug Anal. 2019, 27, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Tripathi, P. Gut microbiome and type 2 diabetes: Where we are and where to go? J. Nutr. Biochem. 2019, 63, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Sonnenburg, J.L.; Bäckhed, F. Diet–microbiota interactions as moderators of human metabolism. Nature 2016, 535, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Forte, N.; Fernández-Rilo, A.C.; Palomba, L.; di Marzo, V.; Cristino, L. Obesity Affects the Microbiota–Gut–Brain Axis and the Regulation Thereof by Endocannabinoids and Related Mediators. Int. J. Mol. Sci. 2020, 21, 1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrarese, R.; Ceresola, E.R.; Preti, A.; Canducci, F. Probiotics, prebiotics and synbiotics for weight loss and metabolic syndrome in the microbiome era. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7588–7605. [Google Scholar]
- Wilkowska, A.; Szałach, Ł.P.; Cubała, W.J. Gut Microbiota in Depression: A Focus on Ketamine. Front. Behav. Neurosci. 2021, 15, 134. [Google Scholar] [CrossRef]
- Lach, G.; Schellekens, H.; Dinan, T.G.; Cryan, J.F. Anxiety, Depression, and the Microbiome: A Role for Gut Peptides. Neurotherapeutics 2018, 15, 36–59. [Google Scholar] [CrossRef] [Green Version]
- Limbana, T.; Khan, F.; Eskander, N. Gut Microbiome and Depression: How Microbes Affect the Way We Think. Cureus 2020, 12, e9966. [Google Scholar] [CrossRef]
- Nikolova, V.L.; Hall, M.R.B.; Hall, L.J.; Cleare, A.J.; Stone, J.M.; Young, A.H. Perturbations in Gut Microbiota Composition in Psychiatric Disorders: A Review and Meta-analysis. JAMA Psychiatry 2021, 78, 1343–1354. [Google Scholar] [CrossRef]
- Winter, G.; Hart, R.A.; Charlesworth, R.P.G.; Sharpley, C.F. Gut microbiome and depression: What we know and what we need to know. Rev. Neurosci. 2018, 29, 629–643. [Google Scholar] [CrossRef]
- Gomaa, E.Z. Human gut microbiota/microbiome in health and diseases: A review. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2020, 113, 2019–2040. [Google Scholar] [CrossRef] [PubMed]
- Passos, M.d.C.F.; Moraes-Filho, J.P. Microbiota intestinal nas doenças digestivas. Arq. Gastroenterol. 2017, 54, 255–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adak, A.; Khan, M.R. An insight into gut microbiota and its functionalities. Cell. Mol. Life Sci. 2019, 76, 473–493. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bäckhed, F.; Ley, R.E.; Sonnenburg, J.L.; Peterson, D.A.; Gordon, J.I. Host-bacterial mutualism in the human intestine. Science 2005, 307, 1915–1920. [Google Scholar] [CrossRef] [Green Version]
- Mangiola, F.; Ianiro, G.; Franceschi, F.; Fagiuoli, S.; Gasbarrini, G.; Gasbarrini, A. Gut microbiota in autism and mood disorders. World J. Gastroenterol. 2016, 22, 361–368. [Google Scholar] [CrossRef]
- Sartor, R.B. Microbial Influences in Inflammatory Bowel Diseases. Gastroenterology 2008, 134, 577–594. [Google Scholar] [CrossRef]
- Quigley, E.M.M.; Eamonn, D.; Quigley, M.M. Gut Bacteria in Health and Disease. Gastroenterol. Hepatol. 2013, 9, 560–569. [Google Scholar]
- Mändar, R.; Mikelsaar, M. Transmission of mother’s microflora to the newborn at birth. Biol. Neonat. 1996, 69, 30–35. [Google Scholar] [CrossRef]
- Bergman, E.N. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef] [Green Version]
- Duncan, S.H.; Belenguer, A.; Holtrop, G.; Johnstone, A.M.; Flint, H.J.; Lobley, G.E. Reduced Dietary Intake of Carbohydrates by Obese Subjects Results in Decreased Concentrations of Butyrate and Butyrate-Producing Bacteria in Feces. Appl. Environ. Microbiology 2007, 73, 1073–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis, P.; Flint, H.J. Diversity, metabolism and microbial ecology of butyrate-producing bacteria from the human large intestine. FEMS Microbiol. Lett. 2009, 294, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portune, K.J.; Beaumont, M.; Davila, A.M.; Tomé, D.; Blachier, F.; Sanz, Y. Gut microbiota role in dietary protein metabolism and health-related outcomes: The two sides of the coin. Trends Food Sci. Technol. 2016, 57, 213–232. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Huang, X.; Fang, S.; Yang, H.; He, M.; Zhao, Y.; Huang, L. Contribution of Host Genetics to the Variation of Microbial Composition of Cecum Lumen and Feces in Pigs. Front. Microbiol. 2018, 9, 2626. [Google Scholar] [CrossRef]
- Thompson, A.L.; Monteagudo-Mera, A.; Cadenas, M.B.; Lampl, M.L.; Azcarate-Peril, M.A. Milk- and solid-feeding practices and daycare attendance are associated with differences in bacterial diversity, predominant communities, and metabolic and immune function of the infant gut microbiome. Front. Cell. Infect. Microbiol. 2015, 5, 3. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.A.; Lim, J.Y.; Kim, B.S.; Cho, S.J.; Kim, N.Y.; Kim, O.B.; Kim, Y. Comparison of the gut microbiota profile in breast-fed and formula-fed Korean infants using pyrosequencing. Nutr. Res. Pract. 2015, 9, 242–248. [Google Scholar] [CrossRef] [Green Version]
- Mariat, D.; Firmesse, O.; Levenez, F.; Guimaraes, V.D.; Sokol, H.; Dore, J.; Corthier, G.; Furet, J.P. The Firmicutes/Bacteroidetes ratio of the human microbiota changes with age. BMC Microbiol. 2009, 9, 123. [Google Scholar] [CrossRef]
- Klingensmith, N.J.; Coopersmith, C.M. The Gut as the Motor of Multiple Organ Dysfunction in Critical Illness. Crit. Care Clin. 2016, 32, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Biedermann, L.; Brülisauer, K.; Zeitz, J.; Frei, P.; Scharl, M.; Vavricka, S.R.; Fried, M.; Loessner, M.J.; Rogler, G.; Schuppler, M. Smoking cessation alters intestinal microbiota: Insights from quantitative investigations on human fecal samples using FISH. Inflamm. Bowel. Dis. 2014, 20, 1496–1501. [Google Scholar] [CrossRef]
- Nishino, K.; Nishida, A.; Inoue, R.; Kawada, Y.; Ohno, M.; Sakai, S.; Inatomi, O.; Bamba, S.; Sugimoto, M.; Kawahara, M.; et al. Analysis of endoscopic brush samples identified mucosa-associated dysbiosis in inflammatory bowel disease. J. Gastroenterol. 2018, 53, 95–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokarek, J.; Gadzinowska, J.; Młynarska, E.; Franczyk, B.; Rysz, J. What Is the Role of Gut Microbiota in Obesity Prevalence? A Few Words about Gut Microbiota and Its Association with Obesity and Related Diseases. Microorganisms 2021, 10, 52. [Google Scholar] [CrossRef] [PubMed]
- Bunyavanich, S.; Shen, N.; Grishin, A.; Wood, R.; Burks, W.; Dawson, P.; Jones, S.M.; Leung, D.Y.; Sampson, H.; Sicherer, S.; et al. Early-life gut microbiome composition and milk allergy resolution. J. Allergy Clin. Immunol. 2016, 138, 1122–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, D.M.; Ma, J.; Prince, A.L.; Antony, K.M.; Seferovic, M.D.; Aagaard, K.M. Maturation of the infant microbiome community structure and function across multiple body sites and in relation to mode of delivery. Nat. Med. 2017, 23, 314–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jie, Z.; Xia, H.; Zhong, S.L.; Feng, Q.; Li, S.; Liang, S.; Zhong, H.; Liu, Z.; Gao, Y.; Zhao, H.; et al. The gut microbiome in atherosclerotic cardiovascular disease. Nat. Commun. 2017, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffery, I.B.; O’Toole, P.W.; Öhman, L.; Claesson, M.J.; Deane, J.; Quigley, E.M.; Simrén, M. An irritable bowel syndrome subtype defined by species-specific alterations in faecal microbiota. Gut 2012, 61, 997–1006. [Google Scholar] [CrossRef]
- Nishida, A.; Inoue, R.; Inatomi, O.; Bamba, S.; Naito, Y.; Andoh, A. Gut microbiota in the pathogenesis of inflammatory bowel disease. Clin. J. Gastroenterol. 2018, 11, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Le Chatelier, E.; Nielsen, T.; Qin, J.; Prifti, E.; Hildebrand, F.; Falony, G.; Almeida, M.; Arumugam, M.; Batto, J.M.; Kennedy, S.; et al. Richness of human gut microbiome correlates with metabolic markers. Nature 2013, 500, 541–546. [Google Scholar] [CrossRef]
- Muñoz-Garach, A.; Diaz-Perdigones, C.; Tinahones, F.J. Microbiota y diabetes mellitus tipo 2′. Endocrinol. Y Nutr. 2016, 63, 560–568. [Google Scholar] [CrossRef]
- Dan, X.; Mushi, Z.; Baili, W.; Han, L.; Enqi, W.; Huanhu, Z.; Shuchun, L. Differential Analysis of Hypertension-Associated Intestinal Microbiota. Int. J. Med. Sci. 2019, 16, 872–881. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, G.T.; Lynch, S.V.; Bloomberg, G.R.; Kattan, M.; Wood, R.A.; Gergen, P.J.; Jaffee, K.F.; Calatroni, A.; Bacharier, L.B.; Beigelman, A. Early-life home environment and risk of asthma among inner-city children. J. Allerg. Clin. Immunol. 2018, 141, 1468–1475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strati, F.; Cavalieri, D.; Albanese, D.; De Felice, C.; Donati, C.; Hayek, J.; Jousson, O.; Leoncini, S.; Renzi, D.; Calabrò, A.; et al. New evidences on the altered gut microbiota in autism spectrum disorders. Microbiome 2017, 5, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachmann, S. Epidemiology of Suicide and the Psychiatric Perspective. IJERPH 2018, 15, 1425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, K. Mental health: A world of depression. Nature 2014, 515, 180–181. [Google Scholar] [CrossRef]
- Ghannoum, M.A.; Ford, M.; Bonomo, R.A.; Gamal, A.; McCormick, T.S. A Microbiome-Driven Approach to Combating Depression During the COVID-19 Pandemic. Front. Nutr. 2021, 8, 576. [Google Scholar] [CrossRef]
- Malhi, G.S.; Mann, J.J. Depression. Lancet 2018, 392, 2299–2312. [Google Scholar] [CrossRef]
- Bains, N.; Abdijadid, S. Major Depressive Disorder. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021; Available online: http://www.ncbi.nlm.nih.gov/books/NBK559078/ (accessed on 5 December 2021).
- Becker, G.; Berg, D.; Lesch, K.P.; Becker, T. Basal limbic system alteration in major depression: A hypothesis supported by transcranial sonography and MRI findings. Int. J. Neuropsychopharm. 2001, 4, 2164. [Google Scholar] [CrossRef]
- Chevalier, G.; Siopi, E.; Guenin-Macé, L.; Pascal, M.; Laval, T.; Rifflet, A.; Boneca, I.G.; Demangel, C.; Colsch, B.; Pruvost, A.; et al. Effect of gut microbiota on depressive-like behaviors in mice is mediated by the endocannabinoid system. Nat. Commun. 2020, 11, 6363. [Google Scholar] [CrossRef]
- Kazemi, A.; Djafarian, K. Gut microbiota and depression. In The Neuroscience of Depression; Martin, C.R., Hunter, L.A., Patel, V.B., Preedy, V.R., Rajendram, R., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 463–472. [Google Scholar] [CrossRef]
- Selye, H. Stress and Disease. Science 1955, 122, 625–631. [Google Scholar] [CrossRef]
- Selye, H. The Stress of Life; McGraw-Hill: New York, NY, USA, 1978. [Google Scholar]
- Drossman, D.A. Gastrointestinal illness and the biopsychosocial model. Psychosom. Med. 1998, 60, 258–267. [Google Scholar] [CrossRef]
- Krych, L.; Hansen, C.H.F.; Hansen, A.K.; van den Berg, F.W.J.; Nielsen, D.S. Quantitatively Different, yet Qualitatively Alike: A Meta-Analysis of the Mouse Core Gut Microbiome with a View towards the Human Gut Microbiome. PLoS ONE 2013, 8, e62578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golden, S.A.; Covington, H.E.; Berton, O.; Russo, S.J. A standardized protocol for repeated social defeat stress in mice. Nat. Protoc. 2011, 6, 1183–1191. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Leonard, B.E. The olfactory bulbectomised rat as a model of depression. Neurosci. Biobehav. Rev. 2005, 29, 627–647. [Google Scholar] [CrossRef] [PubMed]
- Koenigs, M.; Grafman, J. The functional neuroanatomy of depression: Distinct roles for ventromedial and dorsolateral prefrontal cortex. Behav. Brain Res. 2009, 201, 239–243. [Google Scholar] [CrossRef] [Green Version]
- Harkin, A.; Kelly, J.P.; Leonard, B.E. A review of the relevance and validity of olfactory bulbectomy as a model of depression. Clin. Neurosci. Res. 2003, 3, 253–262. [Google Scholar] [CrossRef]
- Kelly, J.R.; Borre, Y.; O’ Brien, C.; Patterson, E.; El Aidy, S.; Deane, J.; Kennedy, P.J.; Beers, S.; Scott, K.; Moloney, G. Transferring the blues: Depression-associated gut microbiota induces neurobehavioural changes in the rat. J. Psychiatr. Res. 2016, 82, 109–118. [Google Scholar] [CrossRef]
- Marx, W.; McGuinness, A.J.; Rocks, T.; Ruusunen, A.; Cleminson, J.; Walker, A.J.; Gomes-da-Costa, S.; Lane, M.; Sanches, M.; Diaz, A.P. The kynurenine pathway in major depressive disorder, bipolar disorder, and schizophrenia: A meta-analysis of 101 studies. Mol. Psychiatry 2021, 26, 4158–4178. [Google Scholar] [CrossRef]
- Colle, R.; Masson, P.; Verstuyft, C.; Fève, B.; Werner, E.; Boursier-Neyret, C.; Walther, B.; David, D.J.; Boniface, B.; Falissard, B. Peripheral tryptophan, serotonin, kynurenine, and their metabolites in major depression: A case–control study. Psychiatry Clin. Neurosci. 2020, 74, 112–117. [Google Scholar] [CrossRef]
- Hoban, A.E.; Moloney, R.D.; Golubeva, A.V.; McVey Neufeld, K.A.; O’Sullivan, O.; Patterson, E.; Stanton, C.; Dinan, T.G.; Clarke, G.; Cryan, J.F. Behavioural and neurochemical consequences of chronic gut microbiota depletion during adulthood in the rat. Neuroscience 2016, 339, 463–477. [Google Scholar] [CrossRef]
- Capuco, A.; Urits, I.; Hasoon, J.; Chun, R.; Gerald, B.; Wang, J.K.; Ngo, A.L.; Simopoulos, T.; Kaye, A.D.; Colontonio, M.M.; et al. Gut Microbiome Dysbiosis and Depression: A Comprehensive Review. Curr. Pain Headache Rep. 2020, 24, 36. [Google Scholar] [CrossRef]
- Zhang, F.F.; Peng, W.; Sweeney, J.A.; Jia, Z.Y.; Gong, Q.Y. Brain structure alterations in depression: Psychoradiological evidence. CNS Neurosci. Ther. 2018, 24, 994–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stockmeier, C.A.; Rajkowska, G. Cellular abnormalities in depression: Evidence from postmortem brain tissue. Dialogues Clin. Neurosci. 2004, 6, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Miguel-Hidalgo, J.J. Brain structural and functional changes in adolescents with psychiatric disorders. Int. J. Adolesc. Med. Health 2013, 25, 245–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Videbech, P.; Ravnkilde, B. Hippocampal volume and depression: A meta-analysis of MRI studies. Amu. J. Psychiatry 2004, 161, 1957–1966. [Google Scholar] [CrossRef]
- Furczyk, K.; Schutová, B.; Michel, T.M.; Thome, J.; Büttner, A. The neurobiology of suicide—A Review of post-mortem studies. J. Mol. Psychiatry 2013, 1, 2. [Google Scholar] [CrossRef] [Green Version]
- Qiao, H.; Li, M.X.; Xu, C.; Chen, H.B.; An, S.C.; Ma, X.M. Dendritic Spines in Depression: What We Learned from Animal Models. Neural Plast. 2016, 2016, 8056370. [Google Scholar] [CrossRef] [Green Version]
- Kida, S.; Kato, T. Microendophenotypes of psychiatric disorders: Phenotypes of psychiatric disorders at the level of molecular dynamics, synapses, neurons, and neural circuits. Curr. Mol. Med. 2015, 15, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Schoenfeld, T.J.; Cameron, H.A. Adult neurogenesis and mental illness. Neuropsychopharmacology 2015, 40, 113–128. [Google Scholar] [CrossRef] [Green Version]
- Coumans, J.V.F.; Palanisamy, S.K.A.; McFarlane, J.; Moens, P.D.J. Proteomic and Microscopic Strategies towards the Analysis of the Cytoskeletal Networks in Major Neuropsychiatric Disorders. Int. J. Mol. Sci. 2016, 17, E581. [Google Scholar] [CrossRef] [Green Version]
- Pandey, G.N.; Rizavi, H.S.; Ren, X.; Fareed, J.; Hoppensteadt, D.A.; Roberts, R.C.; Conley, R.R.; Dwivedi, Y. Proinflammatory cytokines in the prefrontal cortex of teenage suicide victims. J. Psychiatr. Res. 2012, 46, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Dean, B.; Gibbons, A.S.; Tawadros, N.; Brooks, L.; Everall, I.P.; Scarr, E. Different changes in cortical tumor necrosis factor-α-related pathways in schizophrenia and mood disorders. Mol. Psychiatry 2013, 18, 767–773. [Google Scholar] [CrossRef] [PubMed]
- Gold, P.W.; Machado-Vieira, R.; Pavlatou, M.G. Clinical and Biochemical Manifestations of Depression: Relation to the Neurobiology of Stress. Neural Plast. 2015, 2015, 581976. [Google Scholar] [CrossRef] [PubMed]
- Min, W.; Liu, C.; Yang, Y.; Sun, X.; Zhang, B.; Xu, L.; Sun, X. Alterations in hypothalamic–pituitary–adrenal/thyroid (HPA/HPT) axes correlated with the clinical manifestations of depression. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2012, 39, 206–211. [Google Scholar] [CrossRef]
- Kapfhammer, H.P. Somatic symptoms in depression. Dialogues Clin. Neurosci. 2006, 8, 227–239. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S. Protective and Damaging Effects of Stress Mediators. N. Engl. J. Med. 1998, 338, 171–179. [Google Scholar] [CrossRef] [Green Version]
- Zdrojewicz, Z.; Sztuka-Pietkiewicz, A.; Pietkiewicz, W. Rola receptorów mineralokortykoidowych w patogenezie nadciśnienia tętniczego. Arter. Hypertens. 2000, 4, 209–216. [Google Scholar]
- Keller, J.; Gomez, R.; Williams, G.; Lembke, A.; Lazzeroni, L.; Murphy, G.M.; Schatzberg, A. HPA Axis in Major Depression: Cortisol, Clinical Symptomatology, and Genetic Variation Predict Cognition. Mol. Psychiatry 2017, 22, 527–536. [Google Scholar] [CrossRef]
- Otis, J.M.; Zhu, M.; Namboodiri, V.; Cook, C.A.; Kosyk, O.; Matan, A.M.; Ying, R.; Hashikawa, Y.; Hashikawa, K.; Trujillo-Pisanty, I. Paraventricular Thalamus Projection Neurons Integrate Cortical and Hypothalamic Signals for Cue-Reward Processing. Neuron 2019, 103, 423–431. [Google Scholar] [CrossRef]
- Mikulska, J.; Juszczyk, G.; Gawrońska-Grzywacz, M.; Herbet, M. HPA Axis in the Pathomechanism of Depression and Schizophrenia: New Therapeutic Strategies Based on Its Participation. Brain Sci. 2021, 11, 1298. [Google Scholar] [CrossRef]
- Bellavance, M.A.; Rivest, S. The HPA–Immune Axis and the Immunomodulatory Actions of Glucocorticoids in the Brain. Front. Immunol. 2014, 5, 136. [Google Scholar] [CrossRef] [Green Version]
- Cohen, S.; Janicki-Deverts, D.; Doyle, W.J.; Miller, G.E.; Frank, E.; Rabin, B.S.; Turner, R.B. Chronic stress, glucocorticoid receptor resistance, inflammation, and disease risk. Proc. Natl. Acad. Sci. USA 2012, 109, 5995–5999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köhler, C.A.; Freitas, T.H.; Maes, M.; de Andrade, N.Q.; Liu, C.S.; Fernandes, B.S.; Stubbs, B.; Solmi, M.; Veronese, N.; Herrmann, N. Peripheral cytokine and chemokine alterations in depression: A meta-analysis of 82 studies. Acta Psychiatry Scand. 2017, 135, 373–387. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Liu, L.; Sheng, C.; Cheng, Z.; Cui, L.; Li, M.; Zhao, Y.; Shi, T.; Yau, T.O.; Li, F. Increased Serum Levels of Cortisol and Inflammatory Cytokines in People with Depression. J. Nerv. Ment. Dis. 2019, 207, 271–276. [Google Scholar] [CrossRef]
- Park, A.J.; Collins, J.; Blennerhassett, P.A.; Ghia, J.E.; Verdu, E.F.; Bercik, P.; Collins, S. Altered colonic function and microbiota profile in a mouse model of chronic depression. Neurogastroenterol. Motil. 2013, 25, 733-e575. [Google Scholar] [CrossRef] [Green Version]
- Knowles, S.R.; Nelson, E.A.; Palombo, E.A. Investigating the role of perceived stress on bacterial flora activity and salivary cortisol secretion: A possible mechanism underlying susceptibility to illness. Biol. Psychol. 2008, 77, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Goto, T.; Kubota, Y.; Tanaka, Y.; Iio, W.; Moriya, N.; Toyoda, A. Subchronic and mild social defeat stress accelerates food intake and body weight gain with polydipsia-like features in mice. Behav. Brain Res. 2014, 270, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Aoki-Yoshida, A.; Aoki, R.; Moriya, N.; Goto, T.; Kubota, Y.; Toyoda, A.; Takayama, Y.; Suzuki, C. Omics Studies of the Murine Intestinal Ecosystem Exposed to Subchronic and Mild Social Defeat Stress. J. Proteome Res. 2016, 15, 3126–3138. [Google Scholar] [CrossRef]
- Bangsgaard Bendtsen, K.M.; Krych, L.; Sørensen, D.B.; Pang, W.; Nielsen, D.S.; Josefsen, K.; Hansen, L.H.; Sørensen, S.J.; Hansen, A.K. Gut microbiota composition is correlated to grid floor induced stress and behavior in the BALB/c mouse. PLoS ONE 2012, 7, e46231. [Google Scholar] [CrossRef] [Green Version]
- Bharwani, A.; Mian, M.F.; Foster, J.A.; Surette, M.G.; Bienenstock, J.; Forsythe, P. Structural and functional consequences of chronic psychosocial stress on the microbiome and host. Psychoneuroendocrinology 2016, 63, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Galley, J.D.; Nelson, M.C.; Yu, Z.; Dowd, S.E.; Walter, J.; Kumar, P.S.; Lyte, M.; Bailey, M.T. Exposure to a social stressor disrupts the community structure of the colonic mucosa-associated microbiota. BMC Microbiol. 2014, 14, 189. [Google Scholar] [CrossRef] [Green Version]
- Bailey, M.T.; Dowd, S.E.; Galley, J.D.; Hufnagle, A.R.; Allen, R.G.; Lyte, M. Exposure to a Social Stressor Alters the Structure of the Intestinal Microbiota: Implications for Stressor-Induced Immunomodulation. Brain Behav. Immun. 2011, 25, 397–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farshim, P.; Walton, G.; Chakrabarti, B.; Givens, I.; Saddy, D.; Kitchen, I.; Swann, R.J.; Bailey, A. Maternal Weaning Modulates Emotional Behavior and Regulates the Gut-Brain Axis. Sci. Rep. 2016, 6, 21958. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Ling, Z.; Zhang, Y.; Mao, H.; Ma, Z.; Yin, Y.; Wang, W.; Tang, W.; Tan, Z.; Shi, J. Altered fecal microbiota composition in patients with major depressive disorder. Brain Behav. Immun. 2015, 48, 186–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, M.T.; Dowd, S.E.; Parry, N.M.A.; Galley, J.D.; Schauer, D.B.; Lyte, M. Stressor Exposure Disrupts Commensal Microbial Populations in the Intestines and Leads to Increased Colonization by Citrobacter rodentium. Infect. Immun. 2010, 78, 1509–1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naseribafrouei, A.; Hestad, K.; Avershina, E.; Sekelja, M.; Linløkken, A.; Wilson, R.; Rudi, K. Correlation between the human fecal microbiota and depression. Neurogastroenterol. Motil. 2014, 26, 1155–1162. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.A.; Tillisch, K.; Gupta, A. Gut/brain axis and the microbiota. J. Clin. Investig. 2015, 125, 926–938. [Google Scholar] [CrossRef] [PubMed]
- Sanada, K.; Nakajima, S.; Kurokawa, S.; Barceló-Soler, A.; Ikuse, D.; Hirata, A.; Yoshizawa, A.; Tomizawa, Y.; Salas-Valero, M.; Noda, Y.; et al. Gut microbiota and major depressive disorder: A systematic review and meta-analysis. J. Affect. Disord. 2020, 266, 1–13. [Google Scholar] [CrossRef]
- Nadeem, I.; Rahman, M.Z.; Ad-Dab’bagh, Y.; Akhtar, M. Effect of probiotic interventions on depressive symptoms: A narrative review evaluating systematic reviews. Psychiatry Clin. Neurosci. 2019, 73, 154–162. [Google Scholar] [CrossRef]
- Liu, R.T.; Walsh, R.F.L.; Sheehan, A.E. Prebiotics and probiotics for depression and anxiety: A systematic review and meta-analysis of controlled clinical trials. Neurosci. Biobehav. Rev. 2019, 102, 13–23. [Google Scholar] [CrossRef]
- Dockray, G.J. Gastrointestinal hormones and the dialogue between gut and brain. J. Physiol. 2014, 592, 2927–2941. [Google Scholar] [CrossRef]
- Reichmann, F.; Holzer, P. Neuropeptide Y: A stressful review. Neuropeptides 2016, 55, 99–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farzi, A.; Reichmann, F.; Holzer, P. The homeostatic role of neuropeptide Y in immune function and its impact on mood and behaviour. Acta Physiol. 2015, 213, 603–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zietek, T.; Rath, E. Inflammation Meets Metabolic Disease: Gut Feeling Mediated by GLP-1′. Front. Immunol. 2016, 7, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosal, S.; Myers, B.; Herman, J.P. Role of central glucagon-like peptide-1 in stress regulation. Physiol. Behav. 2013, 122, 201–207. [Google Scholar] [CrossRef] [Green Version]
- del Boca, C.; Lutz, P.E.; le Merrer, J.; Koebel, P.; Kieffer, B.L. Cholecystokinin knock-down in the basolateral amygdala has anxiolytic and antidepressant-like effects in mice. Neuroscience 2012, 218, 185–195. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Wong, P.T.H.; Spiess, J.; Zhu, Y.Z. Cholecystokinin-2 (CCK2) receptor-mediated anxiety-like behaviors in rats. Neurosci. Biobehav. Rev. 2005, 29, 1361–1373. [Google Scholar] [CrossRef]
- Turnbull, A.V.; Rivier, C. Corticotropin-releasing factor (CRF) and endocrine responses to stress: CRF receptors, binding protein, and related peptides. Proc. Soc. Exp. Biol. Med. 1997, 215, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kawahito, Y.; Sano, H.; Kawata, M.; Yuri, K.; Mukai, S.; Yamamura, Y.; Kato, H.; Chrousos, G.P.; Wilder, R.L.; Kondo, M. Local secretion of corticotropin-releasing hormone by enterochromaffin cells in human colon. Gastroenterology 1994, 106, 859–865. [Google Scholar] [CrossRef]
- Rodiño-Janeiro, B.K.; Alonso-Cotoner, C.; Pigrau, M.; Lobo, B.; Vicario, M.; Santos, J. Role of Corticotropin-releasing Factor in Gastrointestinal Permeability. J. Neurogastroenterol. Motil. 2015, 21, 33–50. [Google Scholar] [CrossRef] [Green Version]
- Sanders, J.; Nemeroff, C. The CRF System as a Therapeutic Target for Neuropsychiatric Disorders. Trends Pharmacol. Sci. 2016, 37, 1045–1054. [Google Scholar] [CrossRef] [Green Version]
- Kojima, M.; Hosoda, H.; Date, Y.; Nakazato, M.; Matsuo, H.; Kangawa, K. Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature 1999, 402, 656–660. [Google Scholar] [CrossRef] [PubMed]
- Schaeffer, M.; Langlet, F.; Lafont, C.; Molino, F.; Hodson, D.J.; Roux, T.; Lamarque, L.; Verdié, P.; Bourrier, E.; Dehouck, B.; et al. Rapid sensing of circulating ghrelin by hypothalamic appetite-modifying neurons. Proc. Natl. Acad. Sci. USA 2013, 110, 1512–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, H.; Kaplan, Z.; Kozlovsky, N.; Gidron, Y.; Matar, M.A.; Zohar, J. Hippocampal microinfusion of oxytocin attenuates the behavioural response to stress by means of dynamic interplay with the glucocorticoid-catecholamine responses. J. Neuroendocrinol. 2010, 22, 889–904. [Google Scholar] [CrossRef] [PubMed]
- Obesity Programs of nutrition, Education, Research and Assessment (OPERA) Gut microbiota: A new path to treat obesity. Int. J. Obes. Supp. 2019, 9, 10–19. [CrossRef] [PubMed]
- Labus, J.S.; Hollister, E.B.; Jacobs, J.; Kirbach, K.; Oezguen, N.; Gupta, A.; Acosta, J.; Luna, R.A.; Aagaard, K.; Versalovic, J.; et al. Differences in gut microbial composition correlate with regional brain volumes in irritable bowel syndrome. Microbiome 2017, 5, 49. [Google Scholar] [CrossRef]
- Yu, M. Variations in gut microbiota and fecal metabolic phenotype associated with depression by 16S rRNA gene sequencing and LC/MS-based metabolomics. J. Pharm. Biomed. Anal. 2017, 138, 231–239. [Google Scholar] [CrossRef]
- Pirbaglou, M.; Katz, J.; de Souza, R.J.; Stearns, J.C.; Motamed, M.; Ritvo, P. Probiotic supplementation can positively affect anxiety and depressive symptoms: A systematic review of randomized controlled trials. Nutr. Res. 2016, 36, 889–898. [Google Scholar] [CrossRef]
- Wallace, C.J.K.; Milev, R. The effects of probiotics on depressive symptoms in humans: A systematic review. Annu. Gen. Psychiatry 2017, 16, 14. [Google Scholar] [CrossRef] [Green Version]
- Ng, Q.X.; Peters, C.; Ho, C.Y.X.; Lim, D.Y.; Yeo, W.S. A meta-analysis of the use of probiotics to alleviate depressive symptoms. J. Affect. Disord. 2018, 228, 13–19. [Google Scholar] [CrossRef]
- Akkasheh, G.; Kashani-Poor, Z.; Tajabadi-Ebrahimi, M.; Jafari, P.; Akbari, H.; Taghizadeh, M.; Memarzadeh, M.R.; Asemi, Z.; Esmaillzadeh, A. Clinical and metabolic response to probiotic administration in patients with major depressive disorder: A randomized, double-blind, placebo-controlled trial. Nutrition 2016, 32, 315–320. [Google Scholar] [CrossRef]
- Pinto-Sanchez, M.I.; Hall, G.B.C.; Ghajar, K.; Nardelli, A.; Bolino, C.; Lau, J.T.; Martin, F.; Cominetti, O.; Welsh, C.; Rieder, A.D. Probiotic Bifidobacterium longum NCC3001 Reduces Depression Scores and Alters Brain Activity: A Pilot Study in Patients with Irritable Bowel Syndrome. Gastroenterology 2017, 153, 448–459.e8. [Google Scholar] [CrossRef] [PubMed]
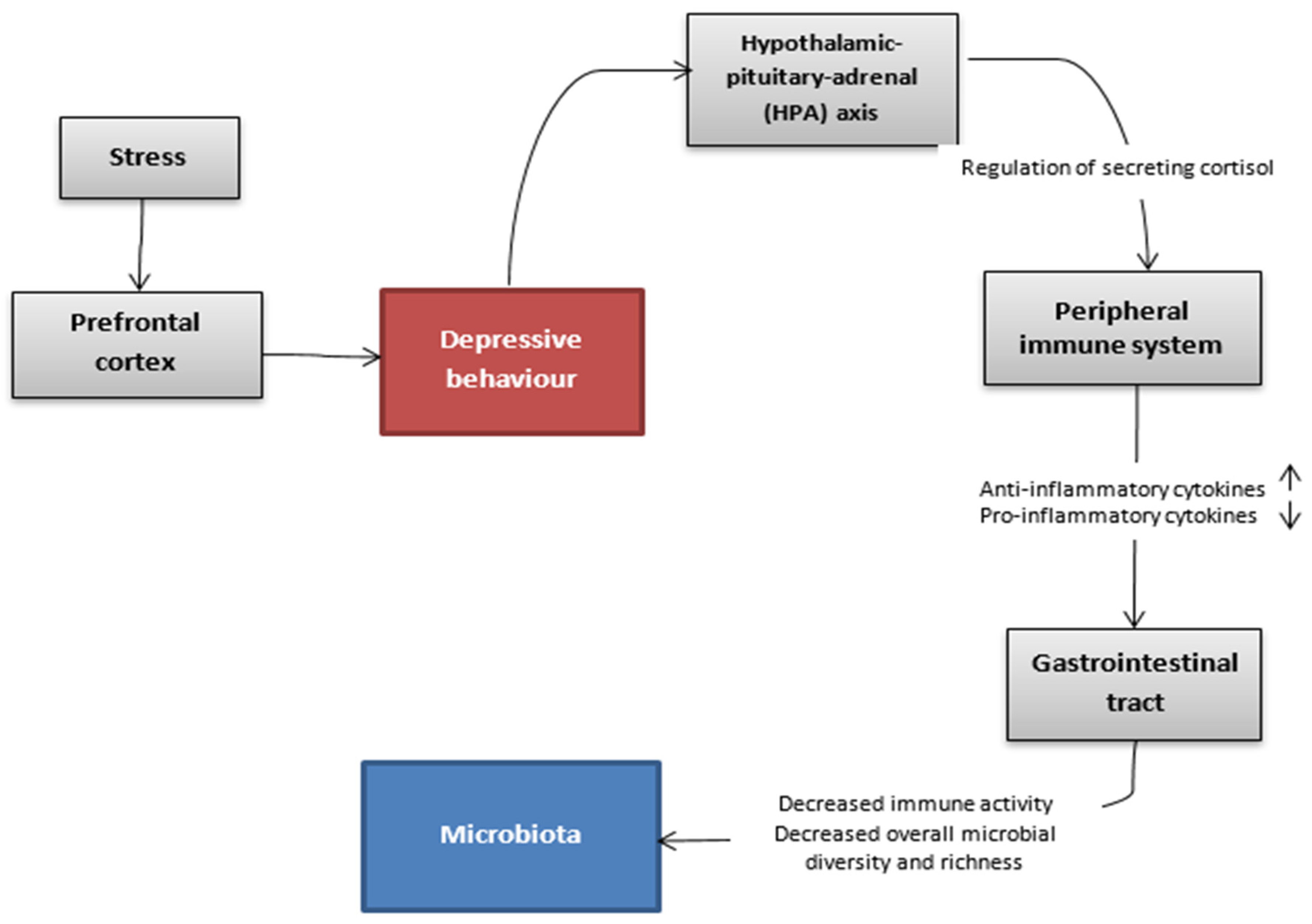

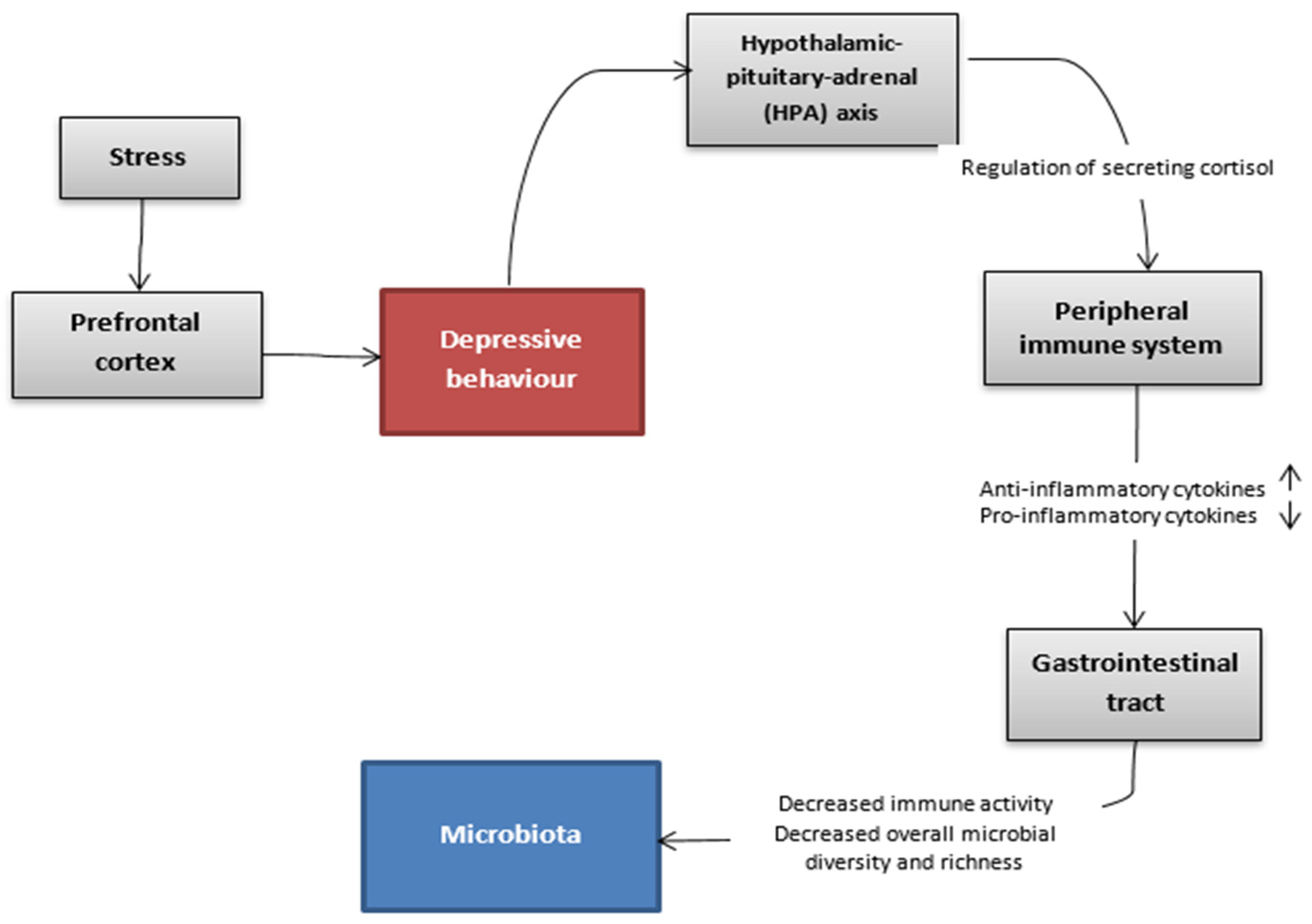{kind=link}
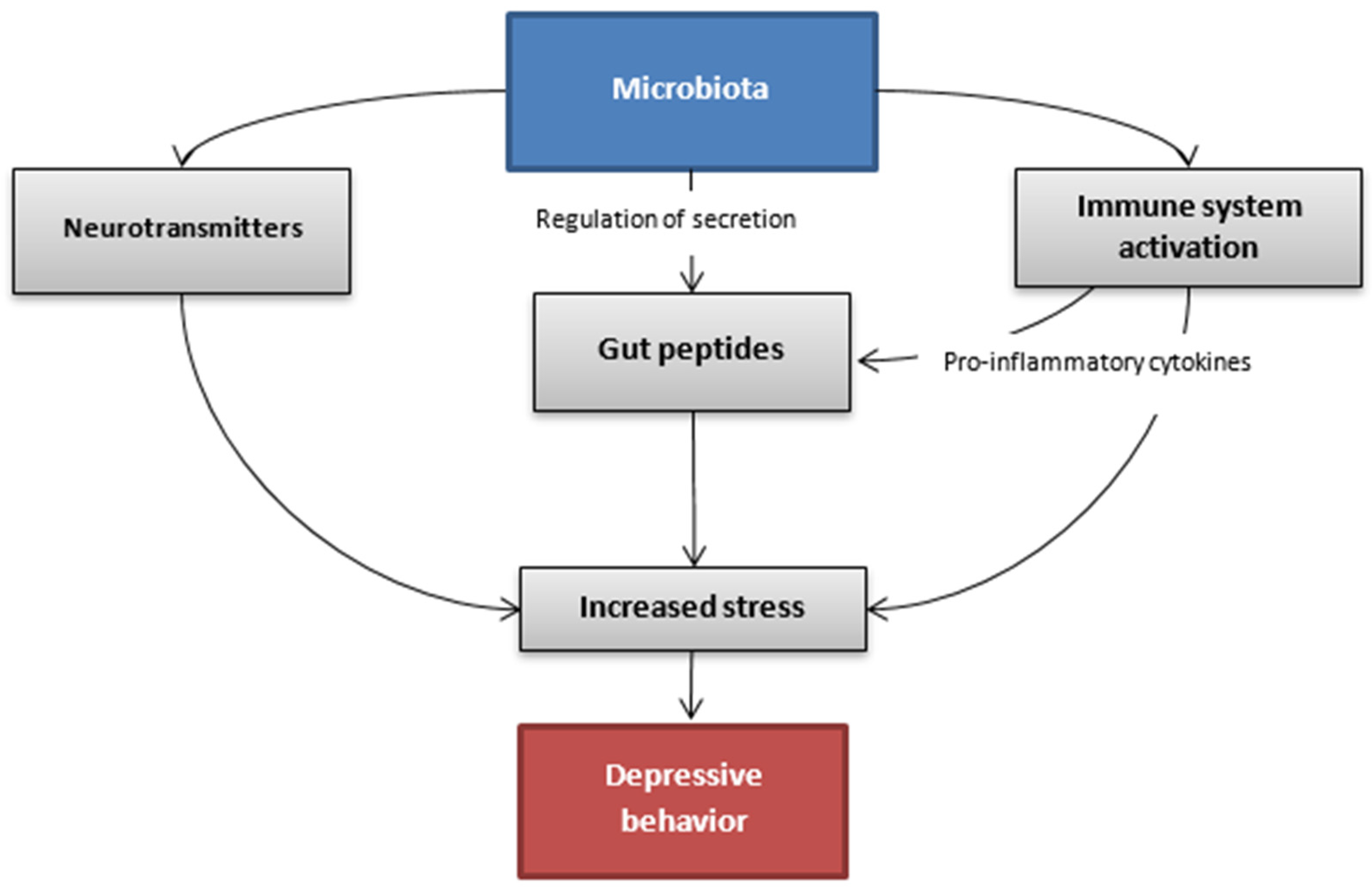
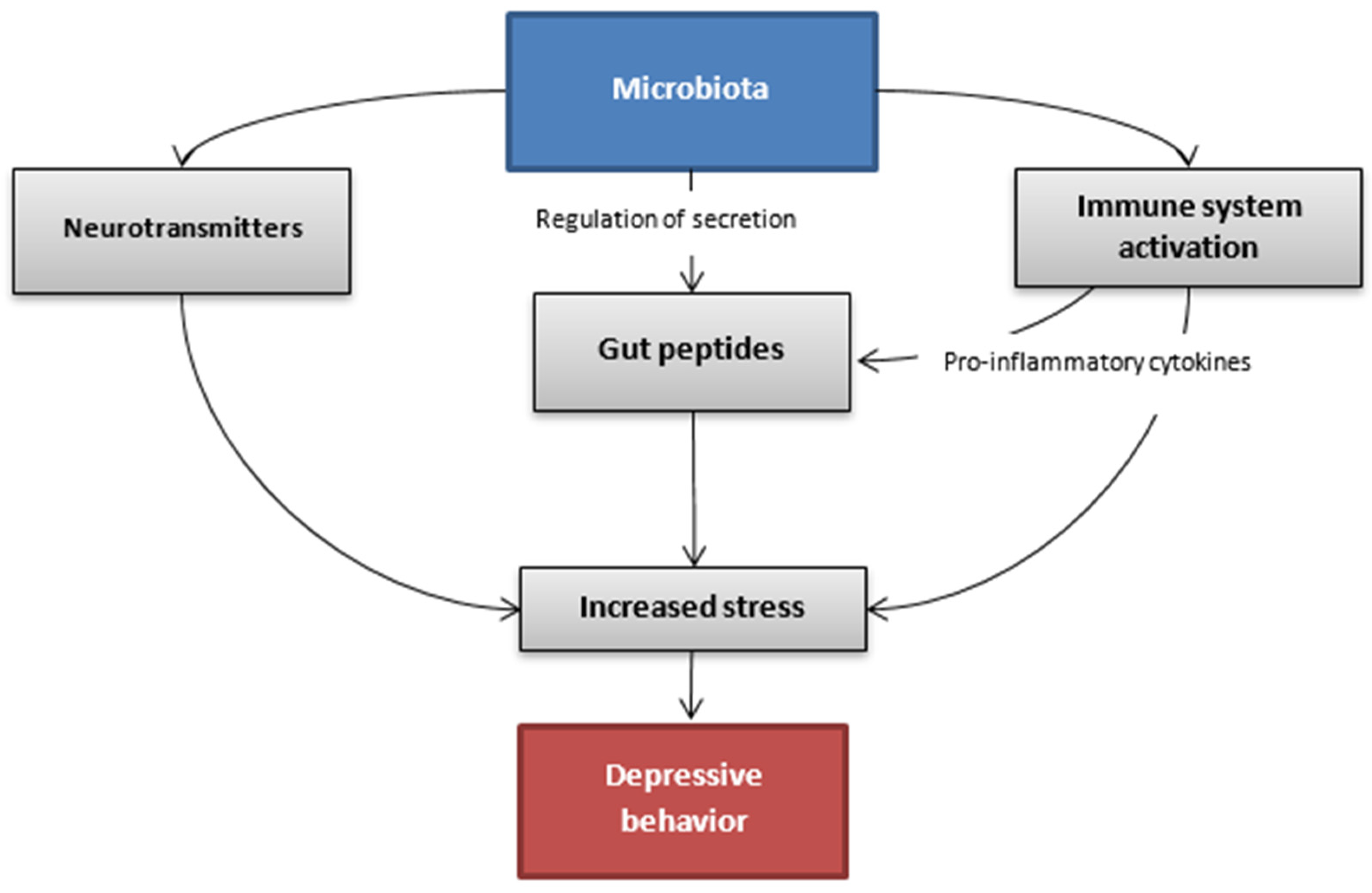{kind=link}
| Disease | Paper | Increase | Decrease |
|---|---|---|---|
| Irritable bowel syndrome | Jeffery et al. (2012) [36] | Firmicutes especially Clostridium, Ruminococcus and Dorea a | Ruminococcus albus, Bacteroides fragilis, Bacteroides vulgatus and Ruminococcus callidus a |
| Inflammatory bowel disease (IBD) | Nishida et al. (2018) [37] | Mucolytic bacteria (Ruminococcus gnavas, Ruminococcus torques), sulfate-reducing bacteria (Desulfovibrio), pathogenic bacteria (adhesion/invasive Escherichia coli) | Firmicutes, SCFA-producing bacteria (Clostridium cluster IV, XIVa, XVII and Faecalibacterium prausnitzzi) |
| Obesity | Le Chatelier et al. (2013) [38] | Porphyromonas, Campylobacter, Bacteroides, Staphylococcus, Parabacteroides, Dialister and Ruminococcus | Lactobacillus, Bifidobacterium, Faecalibacterium, Akkermansia, Methanobrevibacter and Coprococcus |
| Insulin resistance and Diabetes mellitus type 2 | Munoz-Garach et al. (2016) [39] | Firmicutes, Lactobacillus gasseri, Streptococcus mutans, Escherichia coli | Bacteroidetes, Roseburia, Eubacterium halli, Faecalibacterium prauznitzi |
| Hypertension | Dan et al. (2019) [40] | Acetobacteroides, Alistipes, Bacteroides, Christensenella, Clostridium sensu stricto, Desulfovibrio, Parabacteroides | Acetobacteroides, Clostridium, Coprobacter, Enterococcus, Enterorhabdus, Lachnospiracea, Lactobacillus, Paraprevotella, Prevotella Romboutsia, Ruminococcus, Veillonella |
| Asthma | O’Connor et al. (2018) [41] | Bifidobacterium adolescentis | Staphylococcus aureus, Faecalibacterium prausnitzii and Clostridium |
| Autistic spectrum disorder | Strati et al. (2017) [42] | Collinsella, Corynebacterium, Dorea and Lactobacillus | Alistipes, Bilophila, Dialister, Parabacteroides and Veillonella |
| Phylum | Class | Order | Family | Genus | Model Organism | Population Shift |
|---|---|---|---|---|---|---|
| Actinobacteria | Actinobacteria | Coriobacteriales | Coriobacteriaceae | Unidentified genera | Mice | Increase [91] |
| Bacteroidetes | Bacteroidia | Bacteroidales | Rikenellaceae | Unidentified genera | Mice | Increase [90], Decrease [92] |
| Bacteroidetes | Bacteroidia | Bacteroidales | Porphyromonadaceae | Odoribacter | Mice | Increase [91] |
| Proteobacteria | Deltaproteobacteria | Desulfovibrionales | Desulfovibrionaceae | Desulfovibrio | Mice | Increase [90] |
| Proteobacteria | Alphaproteobacteria | Rhodobacterales | Hyphomonadaceae | Ponticaulis | Mice | Increase [93] |
| Firmicutes | Clostridia | Clostridiales | Lachnospiraceae | Pseudobutyrivibrio | Mice | Decrease [94] |
| Firmicutes | Clostridia | Clostridiales | Lachnospiraceae | Coprococcus | Mice | Decrease [94] |
| Firmicutes | Clostridia | Clostridiales | Lachnospiraceae | Roseburia | Mice | Increase [94] |
| Firmicutes | Clostridia | Clostridiales | Lachnospiraceae | Dorea | Mice | Decrease [94] |
| Firmicutes | Clostridia | Clostridiales | Peptostreptococcaceae | Clostridium | Mice | Increase [94] |
| Firmicutes | Clostridia | Clostridiales | Ruminococcaceae | Oscillospira | Mice | Decrease [92] |
| Firmicutes | Bacilli | Lactobacillales | Enterococcaceae | Enterococcus | Mice | Increase [92], Decrease [95] |
| Firmicutes | Bacilli | Lactobacillales | Lactobacillaceae | Unidentified genera | Mice | Decrease [93] |
| Firmicutes | Bacilli | Lactobacillales | Lactobacillaceae | Lactobacillus | Mice | Increase [92], Decrease [93,94] |
| Firmicutes | Erysipelotrichia | Erysipelotrichales | Erysiopelotrichaceae | Allobaculum | Mice | Decrease [90] |
| Deferribacteres | Deferribacteres | Deferribacterales | Deferribacteraceae | Mucispirillum | Mice | Decrease [90] |
| Bacteroidetes | Bacteroidia | Bacteroidales | Rikenellaceae | Alistipes | Mice, Human | Increase [91,96] |
| Bacteroidetes | Bacteroidia | Bacteroidales | Porphyromonadaceae | Unidentified genera | Mice, Human | Increase [96], Decrease [93,97] |
| Bacteroidetes | Bacteroidia | Bacteroidales | Porphyromonadaceae | Parabacteroides | Human, Mice | Increase [96], Decrease [93,94] |
| Firmicutes | Clostridia | Clostridiales | Lachnospiraceae | Unidentified genera | Human, Mice | Increase [90], Decrease [92,96,98] |
| Proteobacteria | Gammaproteobacteria | Enterobacteriales | Enterobacteriaceae | Unidentified genera | Human | Increase [96] |
| Actinobacteria | Actinobacteria | Coriobacteriales | Coriobacteriaceae | Eggerthella | Human | Increase [59] |
| Bacteroidetes | Bacteroidia | Bacteroidales | Bacteroidaceae | Unidentified genera | Human | Decrease [96] |
| Bacteroidetes | Bacteroidia | Bacteroidales | Bacteroidaceae | Bacteroides | Human | Decrease [96] |
| Bacteroidetes | Bacteroidia | Bacteroidales | Prevotellaceae | Unidentified genera | Human | Decrease [59,96] |
| Bacteroidetes | Bacteroidia | Bacteroidales | Prevotellaceae | Paraprevotella | Human | Increase [59] |
| Bacteroidetes | Bacteroidia | Bacteroidales | Prevotellaceae | Prevotella | Human | Decrease [59,96] |
| Firmicutes | Clostridia | Clostridiales | Lachnospiraceae | Anaerofilum | Human | Increase [59] |
| Firmicutes | Clostridia | Clostridiales | Lachnospiraceae | Blautia | Human | Increase [96] |
| Firmicutes | Clostridia | Clostridiales | Ruminococcaceae | Ruminococcus | Human | Decrease [96] |
| Firmicutes | Clostridia | Clostridiales | Clostridiaceae | Faecalibacterium | Human | Decrease [96] |
| Firmicutes | Clostridia | Thermoanaerobacterales | Thermoanaerobacteraceae | Gelria | Human | Increase [59] |
| Firmicutes | Erysipelotrichia | Erysipelotrichales | Erysiopelotrichaceae | Unidentified genera | Human | Decrease [96] |
| Firmicutes | Erysipelotrichia | Erysipelotrichales | Erysiopelotrichaceae | Turicibacter | Human | Increase [59] |
| Firmicutes | Erysipelotrichia | Erysipelotrichales | Erysipelotrichidae | Holdemania | Human | Increase [59] |
| Firmicutes | Negativicutes | Selenomonadales | Acidaminococcaceae | Unidentified genera | Human | Increase [96] |
| Firmicutes | Negativicutes | Veillonellales | Veillonellaceae | Unidentified genera | Human | Decrease [96] |
| Firmicutes | Negativicutes | Veillonellales | Veillonellaceae | Dialister | Human | Decrease [59,96] |
| Firmicutes | Negativicutes | Veillonellales | Veillonellaceae | Megamonas | Human | Increase [96] |
| Fusobacteria | Fusobacteriales | Fusobacteriaceae | Fusobacterium | Unidentified genera | Human | Increase [96] |
| Gut Peptide | Producing Cells | Releasing Factor | Peripheral Function | Central Function |
|---|---|---|---|---|
| PYY [104,105] | L-cells a | food intake | inhibition of gastric emptying and intestinal motor activity | modulation of anxiety and stress-related disorders |
| GLP-1 [106,107] | L-cells a | food intake | stimulation of insulin release and inhibition of glucagon secretion | modulation of the HPA axis and response to stress |
| CCK [108,109] | I-cells a | food intake | suppression of appetite, gastric emptying, gallbladder contraction, pancreatic enzymes release | increased anxiety-like behavior |
| CRF [110,111,112,113] | effector neurons of hypothalamus and enterochromaffin cells of the colon | stress | inhibition of gastric emptying, stimulation of colonic motility and impairment of the intestinal epithelial barrier | increased anxiety and depressive disorder |
| ghrelin [114,115] | A-cells a | starvation | increase of appetite and adipogenesis | modulation of stress response, anxiety and depressive disorder |
| oxytocin [116] | magnocellular neurons in hypothalamus | stress | facilitation of parturition and stimulation of lactation | reduced anxiety-like behavior and antidepressant effect |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Młynarska, E.; Gadzinowska, J.; Tokarek, J.; Forycka, J.; Szuman, A.; Franczyk, B.; Rysz, J. The Role of the Microbiome-Brain-Gut Axis in the Pathogenesis of Depressive Disorder. Nutrients 2022, 14, 1921. https://doi.org/10.3390/nu14091921
Młynarska E, Gadzinowska J, Tokarek J, Forycka J, Szuman A, Franczyk B, Rysz J. The Role of the Microbiome-Brain-Gut Axis in the Pathogenesis of Depressive Disorder. Nutrients. 2022; 14(9):1921. https://doi.org/10.3390/nu14091921
Chicago/Turabian StyleMłynarska, Ewelina, Joanna Gadzinowska, Julita Tokarek, Joanna Forycka, Aleksandra Szuman, Beata Franczyk, and Jacek Rysz. 2022. "The Role of the Microbiome-Brain-Gut Axis in the Pathogenesis of Depressive Disorder" Nutrients 14, no. 9: 1921. https://doi.org/10.3390/nu14091921
APA StyleMłynarska, E., Gadzinowska, J., Tokarek, J., Forycka, J., Szuman, A., Franczyk, B., & Rysz, J. (2022). The Role of the Microbiome-Brain-Gut Axis in the Pathogenesis of Depressive Disorder. Nutrients, 14(9), 1921. https://doi.org/10.3390/nu14091921