
Beneficial Effects of Flaxseed and/or Mulberry Extracts Supplementation in Ovariectomized Wistar Rats

 , , , , and
, , , , and
Abstract
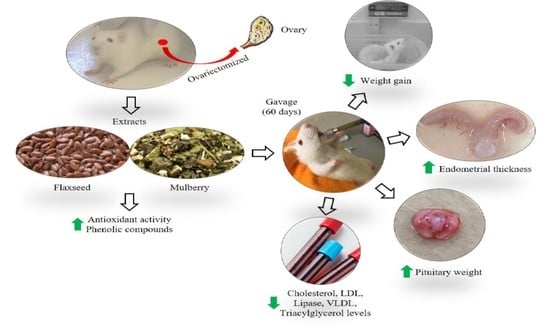
1. Introduction
2. Materials and Methods
2.1. Obtaining and Preparing the Administered Solutions
2.1.1. Flaxseed and Mulberry
2.1.2. Estrogen
2.2. In vitro Antioxidant Assays
2.2.1. Determination of Total Antioxidant Activity by Capturing the Free Radical DPPH (2,2′-Diphenyl-1-Picryl-Hydrazil)
2.2.2. Determination of Antioxidant Activity by the β-Carotene/Linoleic Acid System
2.2.3. Quantification of the Total Phenolic Content by the Folin-Ciocalteu Method
2.3. Animals
2.3.1. Bilateral Ovariectomy (OVX)
2.3.2. Experimental Groups
2.4. Experimental Protocol
2.4.1. Solution Administration
2.4.2. Body Weight
2.4.3. Euthanasia
2.4.4. Vaginal Wash
2.4.5. Pituitary Weight
2.5. Biochemical Analyses
2.5.1. Quantification of Total Cholesterol (TC), Triacylglycerols (TAG), Very-Low-Density Lipoprotein Cholesterol (VLDL-c), Low-Density Lipoprotein Cholesterol (LDL-c), and High-Density Lipoprotein Cholesterol (HDL-c):
2.5.2. Quantification of Lipase, Uric Acid, Glutamic-Oxalacetic Transaminase (GOT) and Glutamic-Pyruvic Transaminase (GPT)
2.6. Histological Analysis
2.6.1. Collection and Processing of the Uterus and Liver
2.6.2. Histological Procedures
2.6.3. Histomorphometric Analysis of the Uterus
2.6.4. Histopathological Analysis of the Liver
2.6.5. Histopathological Analysis of the Kidney
2.7. Statistical Analysis
3. Results
3.1. Total Antioxidant and Total Phenolic Compounds Analysis
3.2. Weight Gain
3.3. Cells Present in the Vaginal Lavage
3.4. Pituitary Weight
3.5. Biochemical Analysis
3.6. Endometrial Thickness
3.7. Liver and Kidney Histopathology
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Almeida, M.; Laurent, M.R.; Dubois, V.; Claessens, F.; O’Brien, C.A.; Bouillon, R.; Vanderschueren, D.; Manolagas, S.C. Estrogens and androgens in skeletal physiology and pathophysiology. Physiol. Rev. 2017, 97, 135–187. [Google Scholar] [CrossRef]
- Hildreth, K.L.; Ozemek, C.; Kohrt, W.M.; Blatchford, P.J.; Moreau, K.L. Vascular dysfunction across the stages of the menopausal transition is associated with menopausal symptoms and quality of life. Menopause 2018, 25, 1011–1019. [Google Scholar] [CrossRef]
- Patel, S.; Homaei, A.; Raju, A.B.; Meher, B.R. Estrogen: The necessary evil for human health, and ways to tame it. Biomed. Pharmacother. 2018, 102, 403–411. [Google Scholar] [CrossRef]
- Palmisano, B.T.; Zhu, L.; Stafford, J.M. Role of Estrogens in the Regulation of Liver Lipid Metabolism. In Sex and Gender Factors Affecting Metabolic Homeostasis, Diabetes and Obesity; Advances in Experimental Medicine and Biology; Springer: Cham, Switzerland, 2017; pp. 227–256. [Google Scholar]
- Nappi, R.E.; Martini, E.; Cucinella, L.; Martella, S.; Tiranini, L.; Inzoli, A.; Brambilla, E.; Bosoni, D.; Cassani, C.; Gardella, B. Addressing Vulvovaginal Atrophy (VVA)/Genitourinary Syndrome of Menopause (GSM) for Healthy Aging in Women. Front. Endocrinol. 2019, 10, 561. [Google Scholar] [CrossRef]
- Kanis, J.A.; Cooper, C.; Rizzoli, R.; Reginster, J.Y. European guidance for the diagnosis and management of osteoporosis in postmenopausal women. Osteoporos. Int. 2019, 30, 3–44, Erratum in Osteoporos. Int. 2020, 30, 3–44. [Google Scholar] [CrossRef]
- Agarwal, S.; Alzahrani, F.A.; Ahmed, A. Hormone replacement therapy: Would it be possible to replicate a functional ovary? Int. J. Mol. Sci. 2018, 19, 3160. [Google Scholar] [CrossRef] [PubMed]
- Lorand, T.; Vigh, E.; Garai, J. Hormonal Action of Plant Derived and Anthropogenic Non-Steroidal Estrogenic Compounds: Phytoestrogens and Xenoestrogens. Curr. Med. Chem. 2010, 17, 3542–3574. [Google Scholar] [CrossRef]
- Narukawa, M. Physiological responses to taste signals of functional food components. Biosci. Biotechnol. Biochem. 2018, 82, 200–206. [Google Scholar] [CrossRef]
- Viggiani, M.T.; Polimeno, L.; Di Leo, A.; Barone, M. Phytoestrogens: Dietary intake, bioavailability, and protective mechanisms against colorectal neoproliferative lesions. Nutrients 2019, 11, 1709. [Google Scholar] [CrossRef] [PubMed]
- Poluzzi, E.; Piccinni, C.; Raschi, E.; Rampa, A.; Recanatini, M.; Ponti, F. Phytoestrogens in Postmenopause: The State of the Art from a Chemical, Pharmacological and Regulatory Perspective. Curr. Med. Chem. 2014, 21, 417–436. [Google Scholar] [CrossRef]
- Petrine, J.C.P.; Del Bianco-Borges, B. The influence of phytoestrogens on different physiological and pathological processes: An overview. Phyther. Res. 2020, 35, 180–197. [Google Scholar] [CrossRef]
- Xu, D.P.; Li, Y.; Meng, X.; Zhou, T.; Zhou, Y.; Zheng, J.; Zhang, J.J.; Li, H. Bin Natural antioxidants in foods and medicinal plants: Extraction, assessment and resources. Int. J. Mol. Sci. 2017, 18, 96. [Google Scholar] [CrossRef]
- Dizavandi, F.R.; Ghazanfarpour, M.; Roozbeh, N.; Kargarfard, L.; Khadivzadeh, T.; Dashti, S. An overview of the phytoestrogen effect on vaginal health and dyspareunia in peri- and post-menopausal women. Post Reprod. Health 2019, 25, 11–20. [Google Scholar] [CrossRef]
- Kasote, D.M. Flaxseed phenolics as natural antioxidants. Int. Food Res. J. 2013, 20, 27–34. [Google Scholar]
- Khalesi, S.; Jamaluddin, R.; Ismail, A. Effect of Raw and Heated Flaxseed (Linum Usitatissimum L.) on Blood Lipid Profiles in Rats. Int. J. Appl. Sci. Technol. 2011, 1, 84–89. [Google Scholar]
- Hu, C.; Yuan, Y.V.; Kitts, D.D. Antioxidant activities of the flaxseed lignan secoisolariciresinol diglucoside, its aglycone secoisolariciresinol and the mammalian lignans enterodiol and enterolactone in vitro. Food Chem. Toxicol. 2007, 45, 2219–2227. [Google Scholar] [CrossRef]
- Brito, A.F.; Zang, Y. A review of lignan metabolism, milk enterolactone concentration, and antioxidant status of dairy cows fed flaxseed. Molecules 2019, 24, 41. [Google Scholar] [CrossRef]
- Parikh, M.; Maddaford, T.G.; Austria, J.A.; Aliani, M.; Netticadan, T.; Pierce, G.N. Dietary flaxseed as a strategy for improving human health. Nutrients 2019, 11, 1171. [Google Scholar] [CrossRef] [PubMed]
- Landete, J.M.; Arqués, J.; Medina, M.; Gaya, P.; de Las Rivas, B.D.; Muñoz, R. Bioactivation of Phytoestrogens: Intestinal Bacteria and Health. Crit. Rev. Food Sci. Nutr. 2016, 56, 1826–1843. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, E.L.; Marcelino, G.; Silva, G.T.; Figueiredo, P.S.; Garcez, W.S.; Corsino, J.; Guimarães, R.d.C.A.; Freitas, K.d.C. Nutraceutical and medicinal potential of the Morus species in metabolic dysfunctions. Int. J. Mol. Sci. 2019, 20, 301. [Google Scholar] [CrossRef]
- Chan, E.W.C.; Lye, P.Y.; Wong, S.K. Phytochemistry, pharmacology, and clinical trials of Morus alba. Chin. J. Nat. Med. 2016, 14, 17–30. [Google Scholar]
- Gryn-Rynko, A.; Bazylak, G.; Olszewska-Slonina, D. New potential phytotherapeutics obtained from white mulberry (Morus alba L.) leaves. Biomed. Pharmacother. 2016, 84, 628–636. [Google Scholar] [CrossRef]
- Lim, S.H.; Choi, C.I. Pharmacological properties of Morus nigra L. (Black Mulberry) as a promising nutraceutical resource. Nutrients 2019, 11, 437. [Google Scholar] [CrossRef]
- De Freitas, M.M.; Fontes, P.R.; Souza, P.M.; Fagg, C.W.; Guerra, E.N.S.; De Medeiros Nóbrega, Y.K.; Silveira, D.; Fonseca-Bazzo, Y.; Simeoni, L.A.; Homem-de-Mello, M.; et al. Extracts of Morus nigra L. Leaves Standardized in chlorogenic acid, rutin and isoquercitrin: Tyrosinase inhibition and cytotoxicity. PLoS ONE 2016, 11, e0163130. [Google Scholar] [CrossRef]
- Naderi, G.A.; Asgary, S.; Sarraf-Zadegan, N.; Oroojy, H.; Afshin-Nia, F. Antioxidant activity of three extracts of Morus nigra. Phyther. Res. 2004, 18, 365–369. [Google Scholar] [CrossRef]
- Prasad, K.; Dhar, A. Flaxseed and Diabetes. Curr. Pharm. Des. 2016, 22, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Zeni, A.L.B.; Moreira, T.D.; Dalmagro, A.P.; Camargo, A.; Bini, L.A.; Simionatto, E.L.; Scharf, D.R. Evaluation of phenolic compounds and lipid-lowering effect of Morus nigra leaves extract. An. Acad. Bras. Cienc. 2017, 89, 2805–2815. [Google Scholar] [CrossRef]
- Domínguez-López, I.; Yago-Aragón, M.; Salas-Huetos, A.; Tresserra-Rimbau, A.; Hurtado-Barroso, S. Effects of dietary phytoestrogens on hormones throughout a human lifespan: A review. Nutrients 2020, 12, 2456. [Google Scholar] [CrossRef]
- Moslehi, N.; Mirmiran, P.; Tehrani, F.R.; Azizi, F. Current evidence on associations of nutritional factors with ovarian reserve and timing of menopause: A systematic review. Adv. Nutr. 2017, 8, 597–612. [Google Scholar] [CrossRef] [PubMed]
- Galvão, E.L.; Da Silva, D.C.F.; Da Silva, J.O.; Moreira, A.V.B.; De Sousa, E.M.B.D. Evaluation of the antioxidant potential and sub-critical extraction of linseed oil. Cienc. Tecnol. Aliment. 2008, 28, 551–557. [Google Scholar] [CrossRef]
- Fu, M.; Feng, H.J.; Chen, Y.; Wang, D.B.; Yang, G.Z. Antioxidant activity of Garcinia xanthochymus leaf, root and fruit extracts in vitro. Chin. J. Nat. Med. 2012, 10, 129–134. [Google Scholar] [CrossRef]
- Sawant, S.H.; Bodhankar, S.L. Flax lignan concentrate reverses alterations in blood pressure, left ventricular functions, lipid profile and antioxidant status in DOCA-salt induced renal hypertension in rats. Ren. Fail. 2016, 38, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Do, S.H.; Lee, J.W.; Jeong, W.I.; Chung, J.Y.; Park, S.J.; Hong, I.H.; Jeon, S.K.; Lee, I.S.; Jeong, K.S. Bone-protecting effect of Rubus coreanus by dual regulation of osteoblasts and osteoclasts. Menopause 2008, 15, 676–683. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Wen, H.X.; Zheng, H.L.; Sun, S.X.; Sun, D.J.; Ni, J. Action mechanism of Zuo Gui Yin Decoction’s promotion on estradiol production in rats during the peri-menopausal period. J. Ethnopharmacol. 2011, 134, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Rufino, M.d.S.M.; Alves, R.E.; de Brito, E.S.; de Morais, S.M.; Sampaio, C.d.G.; Pérez-Jiménez, J.; Saura-Calixto, F.D. Metodologia científica: Determinação da atividade antioxidante total em frutas pela captura do radical livre DPPH. Embrapa Agroindústria Trop.-Comun. Técnico 2007, 1–4. Available online: https://www.infoteca.cnptia.embrapa.br/bitstream/doc/426953/1/Cot127.pdf (accessed on 3 June 2022).
- Rufino, M.d.S.M.; Alves, R.E.; de Brito, E.S.; Filho, J.M.; Moreira, A.V.B. Metodologia Científica: Determinação da Atividade Antioxidante Total em Frutas no Sistema β-caroteno/Ácido Linoléico. Embrapa Agroindústria Trop.-Comun. Técnico 2006, 1–4. Available online: https://ainfo.cnptia.embrapa.br/digital/bitstream/CNPAT-2010/11963/1/cot-126.pdf (accessed on 3 June 2022).
- Nuutila, A.M.; Puupponen-Pimiä, R.; Aarni, M.; Oksman-Caldentey, K.M. Comparison of antioxidant activities of onion and garlic extracts by inhibition of lipid peroxidation and radical scavenging activity. Food Chem. 2003, 81, 485–493. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A.J. Colorimetry to total phenolics with phosphomolybdic acid reagents. Am. J. Enol. Vinic. 1985, 20, 144–158. [Google Scholar]
- Gorriti, A.; Arroyo, J.; Quispe, F.; Cisneros, B.; Condorhuamán, M.; Almora, Y.; Chumpitaz, V. Oral toxicity at 60-days of sacha inchi oil (Plukenetia volubilis L.) and linseed (Linum usitatissimum L.), and determination of lethal dose 50 in rodents. Rev. Peru. Med. Exp. Salud Publica 2010, 27, 352–360. [Google Scholar] [CrossRef]
- Vilela, M.G.; dos Santos Júnior, J.L.; Silva, J.G.d.C. E Determinação do ciclo estral em ratas por lavado vaginal. Femina 2007, 35, 667–670. [Google Scholar]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Dursun, A.; Sendag, F.; Terek, M.C.; Yilmaz, H.; Oztekin, K.; Baka, M.; Tanyalcin, T. Morphometric changes in the endometrium and serum leptin levels during the implantation period of the embryo in the rat in response to exogenous ovarian stimulation. Fertil. Steril. 2004, 82, 1121–1126. [Google Scholar] [CrossRef]
- Bedossa, P. Pathology of non-alcoholic fatty liver disease. Liver Int. 2017, 37, 85–89. [Google Scholar] [CrossRef]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- Yarijani, M.Z.; Najafi, H.; Shackebaei, D.; Madani, S.H.; Modarresi, M.; Jassemi, S.V. Amelioration of renal and hepatic function, oxidative stress, inflammation and histopathologic damages by Malva sylvestris extract in gentamicin induced renal toxicity. Biomed. Pharmacother. 2019, 112, 108635. [Google Scholar] [CrossRef] [PubMed]
- Van Hung, P. Phenolic Compounds of Cereals and Their Antioxidant Capacity. Crit. Rev. Food Sci. Nutr. 2016, 56, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Khan, H.; Shah, M.; Khan, R.; Khan, F. Chemical composition and antioxidant activity of certain Morus species. J. Zhejiang Univ. Sci. B 2010, 11, 973–980. [Google Scholar] [CrossRef]
- Herchi, W.; Al Hujaili, A.D.; Sakouhi, F.; Sebei, K.; Trabelsi, H.; Kallel, H.; Boukhchina, S. Flaxseed hull: Chemical composition and antioxidant activity during development. J. Oleo Sci. 2014, 63, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Siger, A.; Nogala-Kalucka, M.; Lampart-Szczapa, E. The content and antioxidant activity of phenolic compounds in cold-pressed plant oils. J. Food Lipids 2008, 15, 137–149. [Google Scholar] [CrossRef]
- Hassimotto, N.M.A.; Da Mota, R.V.; Cordenunsi, B.R.; Lajolo, F.M. Physico-chemical characterization and bioactive compounds of blackberry fruits (Rubus sp.) grown in Brazil. Cienc. Tecnol. Aliment. 2008, 28, 702–708. [Google Scholar] [CrossRef]
- Tu, J.; Shi, D.; Wen, L.; Jiang, Y.; Zhao, Y.; Yang, J.; Liu, H.; Liu, G.; Yang, B. Identification of moracin N in mulberry leaf and evaluation of antioxidant activity. Food Chem. Toxicol. 2019, 132, 110730. [Google Scholar] [CrossRef] [PubMed]
- Marco, G.J. A rapid method for evaluation of antioxidants. J. Am. Oil Chem. Soc. 1968, 45, 594. [Google Scholar] [CrossRef]
- Miller, H.E. A simplified method for the evaluation of antioxidants. J. Am. Oil Chem. Soc. 1971, 48, 91. [Google Scholar] [CrossRef]
- Vizzotto, M.; Pereira, M.C. Amora-preta (Rubus sp.): Otimização do processo de extração para determinação de compostos fenólicos antioxidantes. Rev. Bras. Frutic. 2011, 33, 1209–1214. [Google Scholar] [CrossRef]
- Barroso, A.K.M.; Torres, A.G.; Castelo-Branco, V.N.; Ferreira, A.; Freitas, S.P.; da Rocha-Leão, M.H.M. Linhaça marrom e dourada Propriedades químicas e funcionais das sementes e dos óleos prensados a frio. Cienc. Rural 2014, 44, 181–187. [Google Scholar] [CrossRef]
- Goettems-Fiorin, P.B.; Costa-Beber, L.C.; dos Santos, J.B.; Friske, P.T.; Sulzbacher, L.M.; Frizzo, M.N.; Ludwig, M.S.; Rhoden, C.R.; Heck, T.G. Ovariectomy predisposes female rats to fine particulate matter exposure’s effects by altering metabolic, oxidative, pro-inflammatory, and heat-shock protein levels. Environ. Sci. Pollut. Res. 2019, 26, 20581–20594. [Google Scholar] [CrossRef]
- Rueda, C.; Osorio, A.M.; Avellaneda, A.C.; Pinzón, C.E.; Restrepo, O.I. The efficacy and safety of estriol to treat vulvovaginal atrophy in postmenopausal women: A systematic literature review. Climacteric 2017, 20, 321–330. [Google Scholar] [CrossRef]
- Rebouças, E.C.C.; Leal, S.; Sá, S.I. Regulation of NPY and α-MSH expression by estradiol in the arcuate nucleus of Wistar female rats: A stereological study. Neurol. Res. 2016, 38, 740–747. [Google Scholar] [CrossRef]
- Molena-Fernandes, C.; Schimidt, G.; Neto-Oliveira, E.; Bersani-Amado, C.; Cuman, R.K. Avaliação dos efeitos da suplementação com farinha de linhaça (Linum usitatissimum L.) marrom e dourada sobre o perfil lipídico e a evolução ponderal em ratos Wistar. Rev. Bras. Plantas Med. 2010, 12, 201–207. [Google Scholar] [CrossRef]
- Gullo, G.I.; Carvalho, C.W.; Ferreira, G. Efeito da Administração Crônica de Extrato Aquoso de Rubus rosifolius sobre o Perfil Glicêmico e Ponderal de Ratos Induzidos ao Diabetes e Obesidade. Rev. Ciênc. Saúde 2015, 5, 24–25. [Google Scholar] [CrossRef][Green Version]
- Shen, J.; Song, N.; Williams, C.J.; Brown, C.J.; Yan, Z.; Xu, C.; Forney, L.J. Effects of low dose estrogen therapy on the vaginal microbiomes of women with atrophic vaginitis. Sci. Rep. 2016, 6, 24380. [Google Scholar] [CrossRef]
- Wierman, M.E. Sex steroid effects at target tissues: Mechanisms of action. Am. J. Physiol.-Adv. Physiol. Educ. 2007, 31, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Mueck, A.O.; Ruan, X.; Prasauskas, V.; Grob, P.; Ortmann, O.; Wierman, M.E. Treatment of vaginal atrophy with estriol and lactobacilli combination: A clinical review. Am. J. Physiol.-Adv. Physiol. Educ. 2018, 31, 140–147. [Google Scholar] [CrossRef]
- Occhiuto, F.; De Pasquale, R.; Guglielmo, G.; Palumbo, D.R.; Zangla, G.; Samperi, S.; Renzo, A.; Circosta, C. Effects of phytoestrogenic isoflavones from red clover (Trifolium pratense L.) on experimental osteoporosis. Phyther. Res. 2007, 21, 130–134. [Google Scholar] [CrossRef]
- Zanwar, A.A.; Aswar, U.M.; Hegde, M.V.; Bodhankar, S.L. Estrogenic and embryo-fetotoxic effects of ethanol extract of Linum usitatissimum in rats. J. Complement. Integr. Med. 2010, 7, 1–17. [Google Scholar] [CrossRef]
- González, S.; Bisogno, T.; Wenger, T.; Manzanares, J.; Milone, A.; Berrendero, F.; Di Marzo, V.; Ramos, J.A.; Fernández-Ruiz, J.J. Sex steroid influence on cannabinoid CB1 receptor mRNA and endocannabinoid levels in the anterior pituitary gland. Biochem. Biophys. Res. Commun. 2000, 270, 260–266. [Google Scholar] [CrossRef]
- Kurz, S.G.; Dennison, K.L.; Samanas, N.B.; Hickman, M.P.; Eckert, Q.A.; Walker, T.L.; Cupp, A.S.; Shull, J.D. Ept7 influences estrogen action in the pituitary gland and body weight of rats. Mamm. Genome 2014, 25, 244–252. [Google Scholar] [CrossRef]
- Nolan, L.A.; Levy, A. The trophic effects of oestrogen on male rat anterior pituitary lactotrophs. J. Neuroendocrinol. 2009, 21, 457–464. [Google Scholar] [CrossRef]
- Ronchetti, S.A.; Miler, E.A.; Duvilanski, B.H.; Cabilla, J.P. Cadmium mimics estrogen-driven cell proliferation and prolactin secretion from anterior pituitary cells. PLoS ONE 2013, 8, e81101. [Google Scholar] [CrossRef] [PubMed]
- Mendelsohn, M.E.; Karas, R.H. The protective effects of estrogen on the cardiovascular system. N. Engl. J. Med. 1999, 340, 1801–1811. [Google Scholar] [CrossRef]
- Hadi, A.; Askarpour, M.; Salamat, S.; Ghaedi, E.; Symonds, M.E.; Miraghajani, M. Effect of flaxseed supplementation on lipid profile: An updated systematic review and dose-response meta-analysis of sixty-two randomized controlled trials. Pharmacol. Res. 2020, 152, 104622. [Google Scholar] [CrossRef]
- Katare, C.; Saxena, S.; Agrawal, S.; Prasad, G. Flax Seed: A Potential Medicinal Food. J. Nutr. Food Sci. 2012, 02, 120–127. [Google Scholar] [CrossRef]
- Goyal, A.; Sharma, V.; Upadhyay, N.; Gill, S.; Sihag, M. Flax and flaxseed oil: An ancient medicine & modern functional food. J. Food Sci. Technol. 2014, 51, 1633–1653. [Google Scholar] [CrossRef]
- Dresseno, L.P.; Lehnen, A.M.; Teló, G.; Silveira, A.; Markoski, M.M.; Machado, U.F.; Schaan, B.D. Impact of flaxseed and soy nuts as dietary supplements on lipid profile, insulin sensitivity, and GLUT4 expression in ovariectomized rats. Appl. Physiol. Nutr. Metab. 2018, 43, 1282–1287. [Google Scholar] [CrossRef]
- Parikh, M.; Netticadan, T.; Pierce, G.N. Flaxseed: Its bioactive components and their cardiovascular benefits. Am. J. Physiol.-Heart Circ. Physiol. 2018, 314, H146–H159. [Google Scholar] [CrossRef] [PubMed]
- Morise, A.; Sérougne, C.; Gripois, D.; Blouquit, M.F.; Lutton, C.; Hermier, D. Effects of dietary alpha linolenic acid on cholesterol metabolism in male and female hamsters of the LPN strain. J. Nutr. Biochem. 2004, 15, 51–61. [Google Scholar] [CrossRef]
- Volpato, G.T.; Calderon, I.M.P.; Sinzato, S.; Campos, K.E.; Rudge, M.V.C.; Damasceno, D.C. Effect of Morus nigra aqueous extract treatment on the maternal-fetal outcome, oxidative stress status and lipid profile of streptozotocin-induced diabetic rats. J. Ethnopharmacol. 2011, 138, 691–696. [Google Scholar] [CrossRef] [PubMed]
- Pawlowska, A.M.; Oleszek, W.; Braca, A. Quali-quantitative analyses of flavonoids of Morus nigra L. and Morus alba L. (Moraceae) fruits. J. Agric. Food Chem. 2008, 56, 3377–3380. [Google Scholar] [CrossRef]
- Jung, U.J.; Lee, M.K.; Park, Y.B.; Kang, M.A.; Choi, M.S. Effect of citrus flavonoids on lipid metabolism and glucose-regulating enzyme mRNA levels in type-2 diabetic mice. Int. J. Biochem. Cell Biol. 2006, 38, 1134–1145. [Google Scholar] [CrossRef] [PubMed]
- Chae, H.J.; Yim, J.E.; Kim, K.A.; Chyun, J.H. Hepatoprotective effects of Rubus coreanus miquel concentrates on liver injuries induced by carbon tetrachloride in rats. Nutr. Res. Pract. 2014, 8, 40–45. [Google Scholar] [CrossRef]
- Mauvais-Jarvis, F.; Clegg, D.J.; Hevener, A.L. The role of estrogens in control of energy balance and glucose homeostasis. Endocr. Rev. 2013, 34, 309–338. [Google Scholar] [CrossRef] [PubMed]
- Hamden, K.; Jaouadi, B.; Zaraî, N.; Rebai, T.; Carreau, S.; Elfeki, A. Inhibitory effects of estrogens on digestive enzymes, insulin deficiency, and pancreas toxicity in diabetic rats. J. Physiol. Biochem. 2011, 67, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Tomaz Pacheco, J.; Beltrame Daleprame, J.; Teles Boaventura, G. Impact of dietary flaxseed (linum usitatissimum) supplementation on biochemical profile in healthy rats. Nutr. Hosp. 2011, 26, 798–802. [Google Scholar] [CrossRef]
- Oliveira, A.C.B.; Oliveira, A.P.; Guimarães, A.L.; Oliveira, R.A.; Silva, F.S.; Reis, S.A.G.B.; Ribeiro, L.A.A.; Almeida, J.R.G.S. Avaliação toxicológica pré-clínica do chá das folhas de Morus nigra l. (Moraceae). Rev. Bras. Plantas Med. 2013, 15, 244–249. [Google Scholar] [CrossRef][Green Version]
- Diel, P.; Schulz, T.; Smolnikar, K.; Strunck, E.; Vollmer, G.; Michna, H. Ability of xeno- and phytoestrogens to modulate expression of estrogen-sensitive genes in rat uterus: Estrogenicity profiles and uterotropic activity. J. Steroid Biochem. Mol. Biol. 2000, 73, 1–10. [Google Scholar] [CrossRef]
- Jefferson, W.N.; Padilla-Banks, E.; Clark, G.; Newbold, R.R. Assessing estrogenic activity of phytochemicals using transcriptional activation and immature mouse uterotrophic responses. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2002, 777, 179–189. [Google Scholar] [CrossRef]
- Dilshad, S.M.R.; Najib-Ur-Rehman; Ahmad, N.; Iqbal, A.; Ali, M.A.; Ahmad, A. Effect of flax seeds (Linum usitatissimum) on uterine and ovarian protein contents, ovarian cholesterol, serum estradiol and onset of puberty in immature female mice. Int. J. Agric. Biol. 2012, 14, 781–786. [Google Scholar]
- Nandini, M.S.; Veena, T.; Swamy, M.N. Effect of extracts of murraya koenigii spreng. And morus alba linn. On the age of attainment of puberty and ovarian folliculogenesis in rats. J. Basic Clin. Pharm. 2010, 1, 203–207. [Google Scholar]
- Moshtaghin, F.N.; Moghadamnia, A.A.; Kazemi, S.; Arbabzadegan, N.; Moudi, E.; Haghanifar, S. Effect of hydroalcoholic extract of flaxseed on bone mineral density in Wistar rats using digital radiography. Casp. J. Intern. Med. 2020, 11, 92–99. [Google Scholar] [CrossRef]
- Toma, L.; Sanda, G.M.; Niculescu, L.S.; Deleanu, M.; Sima, A.V.; Stancu, C.S. Phenolic compounds exerting lipid-regulatory, anti-inflammatory and epigenetic effects as complementary treatments in cardiovascular diseases. Biomolecules 2020, 10, 641. [Google Scholar] [CrossRef] [PubMed]
- Giannini, E.G.; Testa, R.; Savarino, V. Liver enzyme alteration: A guide for clinicians. CMAJ 2005, 172, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.E.; Cain, J.; Small, C.; Hales, D.B. Therapeutic effect of flax-based diets on fatty liver in aged laying hens. Poult. Sci. 2016, 95, 2624–2632. [Google Scholar] [CrossRef] [PubMed]
- Palla, A.H.; Gilani, A.U.H.; Bashir, S.; Ur Rehman, N. Multiple Mechanisms of Flaxseed: Effectiveness in Inflammatory Bowel Disease. Evid.-Based Complement. Altern. Med. 2020, 2020, 7974835. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Deng, Q.; Tang, Y.; Xiao, L.; Liu, L.; Yao, P.; Tang, H.; Dong, X. Flaxseed Oil Attenuates Hepatic Steatosis and Insulin Resistance in Mice by Rescuing the Adaption to ER Stress. J. Agric. Food Chem. 2018, 66, 10729–10740. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, H.; Yin, P.; Fan, H.; Sun, L.; Liu, Y. Flaxseed oil ameliorates alcoholic liver disease via anti-inflammation and modulating gut microbiota in mice. Lipids Health Dis. 2017, 16, 44. [Google Scholar] [CrossRef] [PubMed]
- Vijaimohan, K.; Jainu, M.; Sabitha, K.E.; Subramaniyam, S.; Anandhan, C.; Shyamala Devi, C.S. Beneficial effects of alpha linolenic acid rich flaxseed oil on growth performance and hepatic cholesterol metabolism in high fat diet fed rats. Life Sci. 2006, 79, 448–454. [Google Scholar] [CrossRef]
- Rizwan, S.; Naqshbandi, A.; Farooqui, Z.; Khan, A.A.; Khan, F. Protective effect of dietary flaxseed oil on arsenic-induced nephrotoxicity and oxidative damage in rat kidney. Food Chem. Toxicol. 2014, 68, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Gupta, G.; Chellappan, D.K.; Kikuchi, I.S.; Pinto, T.d.J.A.; Pabreja, K.; Agrawal, M.; Singh, Y.; Tiwari, J.; Duaf, K. Nephrotoxicity in rats exposed to paracetamol: The protective role of moralbosteroid, a steroidal glycoside. J. Environ. Pathol. Toxicol. Oncol. 2017, 36, 113–119. [Google Scholar] [CrossRef] [PubMed]







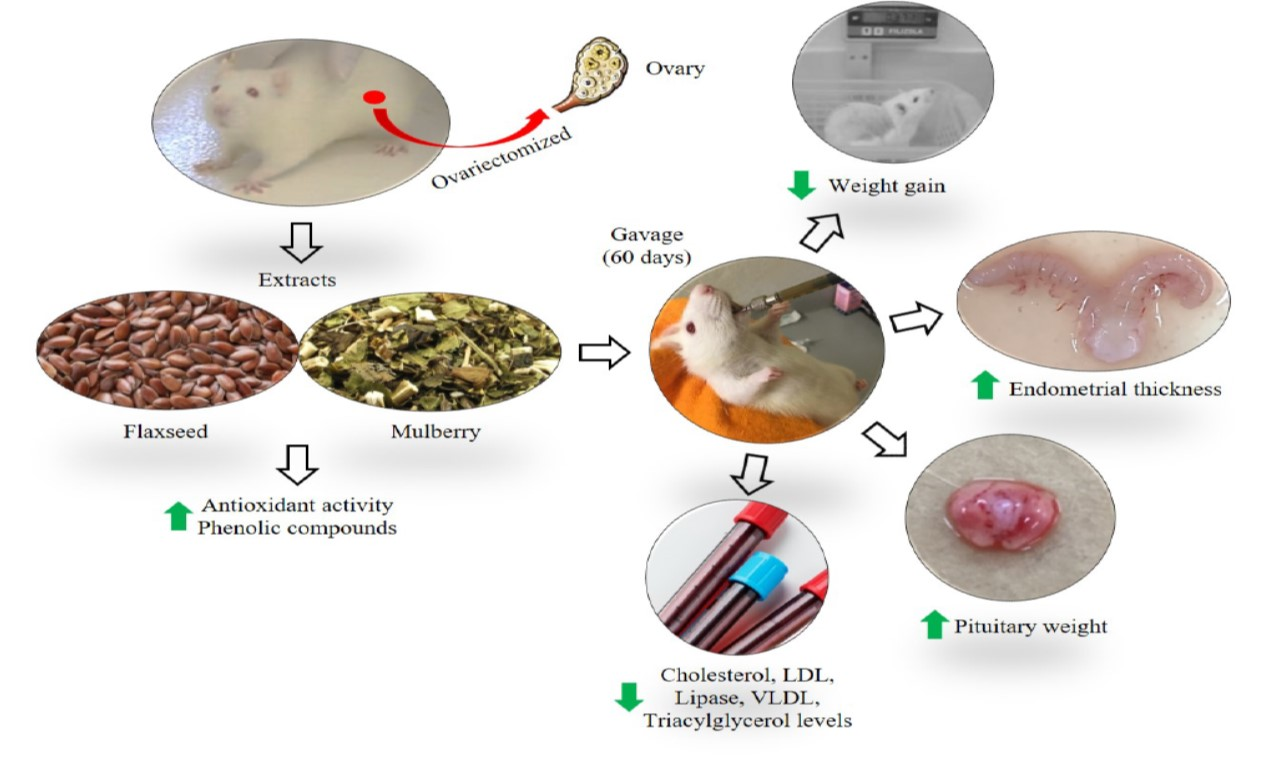{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50 | % Oxidation Inhibition | TPC | ||
|---|---|---|---|---|
| Extract | (mg/mL) | DPPH | β-Carotene/Linoleic Acid | (mg GAE/100g) |
| Flaxseed | 6.93 ± 0.431 | 74.55 ± 4.64 | - | 1395.4 ± 11.83 |
| Mulberry | 0.04 ± 0.007 | 73.44 ± 14.1 | 44.77 ± 28.90 | 1482.6 ± 37.08 |
| Experimental Groups | |||||||
|---|---|---|---|---|---|---|---|
| Histopathologic Damages | Definition | Score | Saline | Estrogen | Flaxseed | Mulberry | Flaxseed + Mulberry |
| Steatosis grade | <5% | 0 | 50% | 100% | 71.40% | 85.70% | 100% |
| 5–33% | 1 | 50% | 0 | 28.50% | 14.20% | 0 | |
| 33–66% | 2 | 0 | 0 | 0 | 0 | 0 | |
| >66% | 3 | 0 | 0 | 0 | 0 | 0 | |
| Inflammation | no focus | 0 | 66.60% | 80% | 71.40% | 85.70% | 57.10% |
| 2–4 foci per 10× field | 1 | 33.30% | 20% | 14.20% | 14.20% | 42.80% | |
| 4–8 foci per 10× field | 2 | 0 | 0 | 14.20% | 0 | 0 | |
| >8 foci per 10× field | 3 | 0 | 0 | 0 | 0 | 0 | |
| Fibrosis stage | no fibrosis | 0 | 100% | 100% | 100% | 100% | 100% |
| perisinusoidal or periportal | 1 | 0 | 0 | 0 | 0 | 0 | |
| perisinusoidal and periportal | 2 | 0 | 0 | 0 | 0 | 0 | |
| bridging | 3 | 0 | 0 | 0 | 0 | 0 | |
| cirrhosis | 4 | 0 | 0 | 0 | 0 | 0 | |
| Ballooning | ≤1:2 | 0 | 100% | 100% | 100% | 100% | 100% |
| 1:2 a 1:3 | 1 | 0 | 0 | 0 | 0 | 0 | |
| 1:3 a 1:4 | 2 | 0 | 0 | 0 | 0 | 0 | |
| ≥1:4 | 3 | 0 | 0 | 0 | 0 | 0 | |
| Total Score | 0 | 33.30% | 80% | 57.10% | 71.40% | 57.10% | |
| 1 | 50% | 20% | 28.50% | 28.50% | 42.80% | ||
| 2 | 16.60% | 0 | 0 | 0 | 0 | ||
| 3 | 0 | 0 | 14.20% | 0 | 0 |
| Experimental Groups | |||||
|---|---|---|---|---|---|
| Histopathologic Damages | Saline | Estrogen | Flaxseed | Mulberry | Flaxseed + Mulberry |
| Bowman’s space | 1.3 | 1.4 | 1.6 | 1.0 | 1.3 |
| Vascular congestion | 1.5 | 1.6 | 2.2 | 2.3 | 2.0 |
| Perivascular edema | 1.0 | 1.4 | 1.1 | 1.4 | 1.1 |
| Intra-tubular casts | 0.2 | 0 | 0 | 0 | 0 |
| Tubular vacuolization | 1.5 | 1.6 | 1.2 | 1.3 | 1.6 |
| Tubular dilatation | 1.0 | 1.2 | 1.4 | 2.0 | 1.4 |
| Exfoliated cells | 1.0 | 1.0 | 1.1 | 1.0 | 1.6 |
| Leucocyte infiltration | 0.2 | 0 | 0.3 | 0.1 | 0.4 |
| Brush border loss | 1.0 | 1.2 | 1.1 | 1.3 | 1.3 |
| Total histopathologic score | 8.7 | 9.4 | 10.3 | 10.4 | 10.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, J.P.C.; Oliveira, E.A.; Pereira, F.A.C.; Seixas, J.N.; Guimaraes, C.S.d.O.; Del Bianco Borges, B. Beneficial Effects of Flaxseed and/or Mulberry Extracts Supplementation in Ovariectomized Wistar Rats. Nutrients 2022, 14, 3238. https://doi.org/10.3390/nu14153238
Pereira JPC, Oliveira EA, Pereira FAC, Seixas JN, Guimaraes CSdO, Del Bianco Borges B. Beneficial Effects of Flaxseed and/or Mulberry Extracts Supplementation in Ovariectomized Wistar Rats. Nutrients. 2022; 14(15):3238. https://doi.org/10.3390/nu14153238
Chicago/Turabian StylePereira, Jéssica Petrine Castro, Erika Aparecida Oliveira, Fernanda Aparecida Castro Pereira, Josilene Nascimento Seixas, Camila Souza de Oliveira Guimaraes, and Bruno Del Bianco Borges. 2022. "Beneficial Effects of Flaxseed and/or Mulberry Extracts Supplementation in Ovariectomized Wistar Rats" Nutrients 14, no. 15: 3238. https://doi.org/10.3390/nu14153238
APA StylePereira, J. P. C., Oliveira, E. A., Pereira, F. A. C., Seixas, J. N., Guimaraes, C. S. d. O., & Del Bianco Borges, B. (2022). Beneficial Effects of Flaxseed and/or Mulberry Extracts Supplementation in Ovariectomized Wistar Rats. Nutrients, 14(15), 3238. https://doi.org/10.3390/nu14153238