
A Novel Sprouted Oat Fermented Beverage: Evaluation of Safety and Health Benefits for Celiac Individuals


 , , , and
, , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals, Reagents and Standards
2.2. Preparation of Sprouted Oat Flour
2.3. Bacterial Strain Activation and Inoculum Preparation
2.4. Manufacture of SOFB
2.5. Manufacture of Placebo Beverage for the Intervention Study
2.6. Analysis of Gluten Content
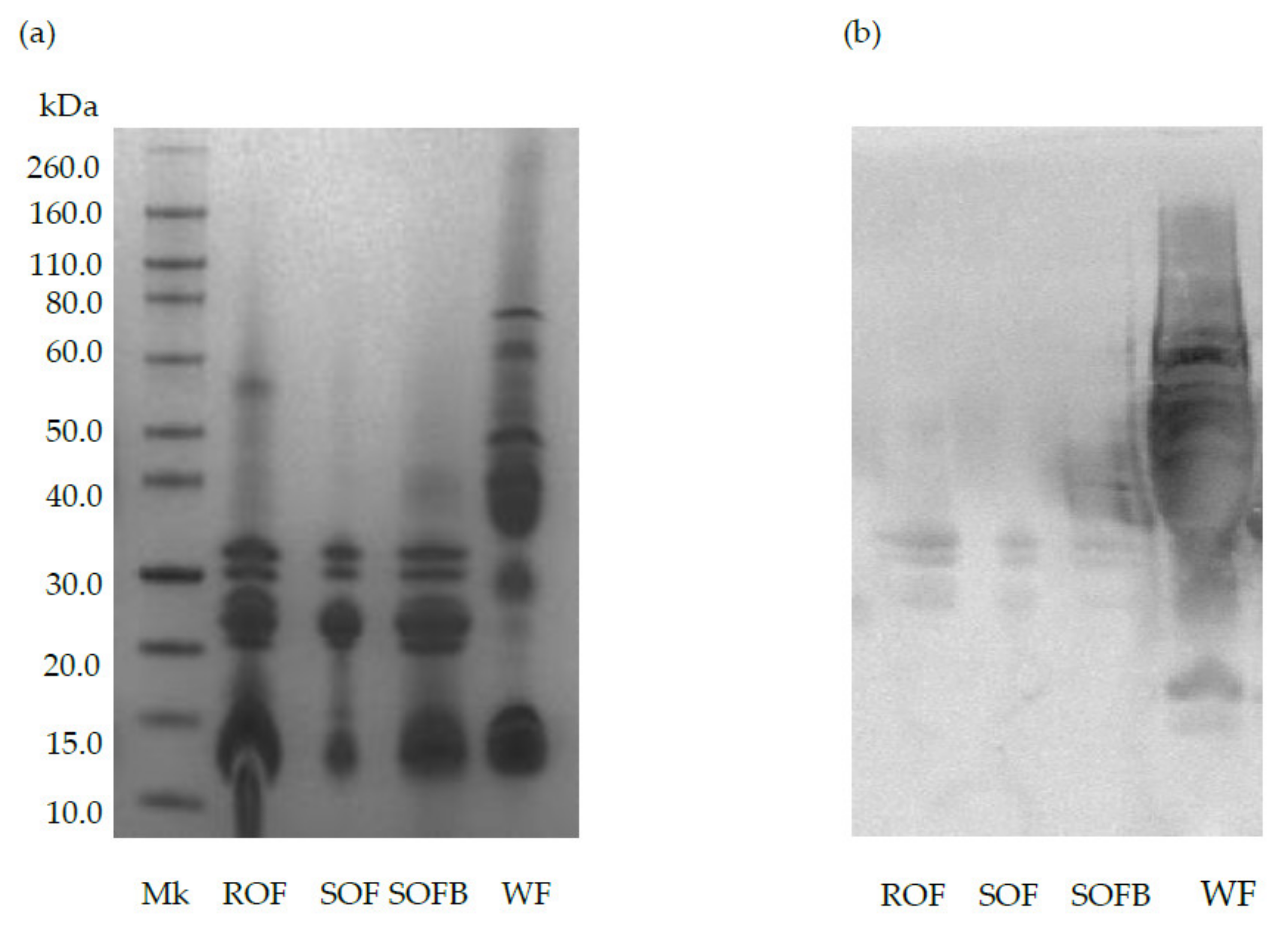
2.7. Evaluation of SOFB Protein Profile by Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.8. Determination of SOFB Immunotoxicity
2.9. Determination of SOFB Antioxidant and Antiinflammatory Activities
2.9.1. Cell Lines
2.9.2. In Vitro Gastrointestinal Digestion
2.9.3. Cell Treatments
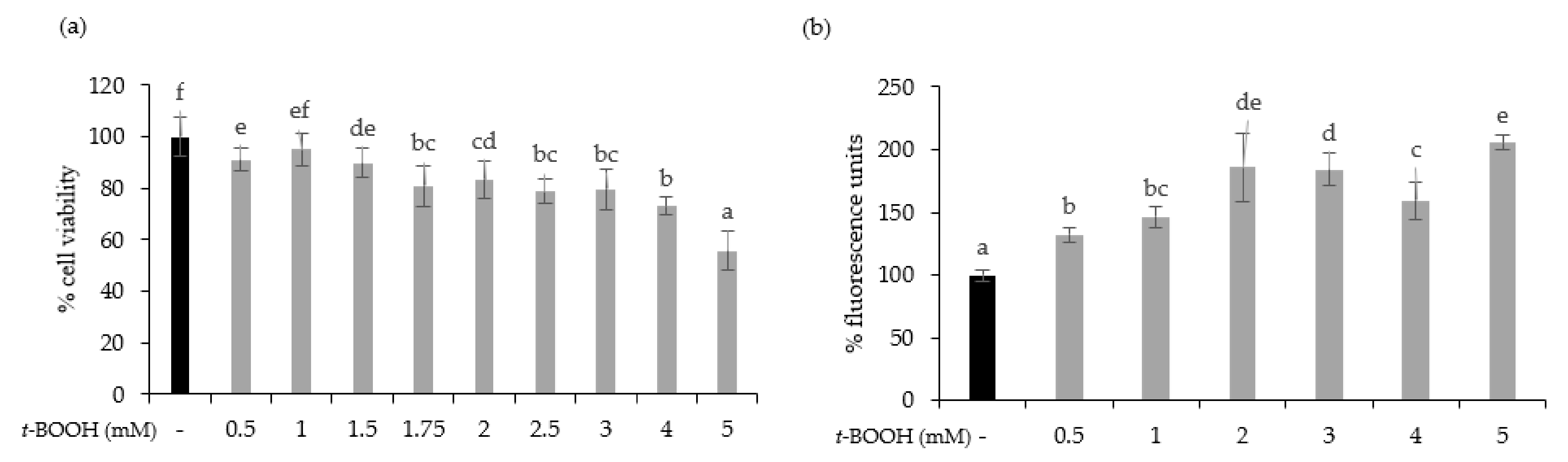
2.9.4. Cell Viability
2.9.5. In Vitro Determination of Reactive Oxygen Species (ROS) Concentration
2.9.6. Anti-Inflammatory In Vitro Assay
2.10. In Vivo Evaluation of Safety and Health Benefits of SOFB in CD Individuals
2.10.1. Study Population and Design
2.10.2. Serological and Histological Analysis
2.10.3. Fecal Samples Collection and Metataxonomic Analysis
2.11. Statistical Analysis
3. Results
3.1. Evaluation of SOFB Gluten Content and Immunotoxicity
3.2. SOFB Did Not Protect Caco-2 Cells against t-BOOH-Induced Oxidative Stress
3.3. SOFB Attenuates Inflammatory Response in LPS-Induced RAW 264.7 Macrophages
3.4. In Vivo Evaluation of SOFB Safety and Health Benefits in CD Subjects
3.4.1. Patient Baseline Characteristics
3.4.2. SOFB Safety Assessment in CD Subjects
3.4.3. SOFB Effects on Body Weight and Serological Markers of Nutritional Status in CD Subjects
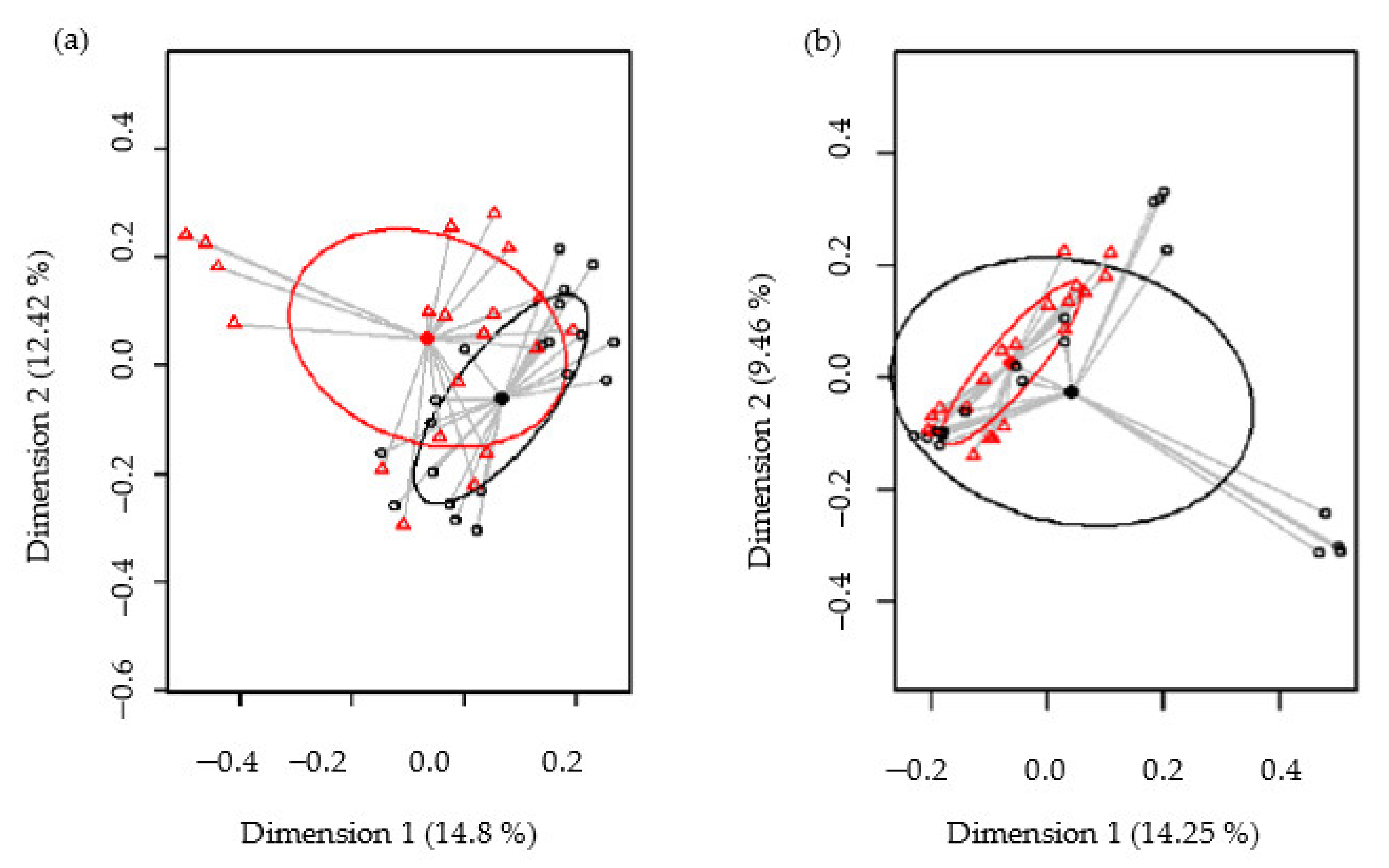
3.4.4. Metataxonomic Analysis of Intestinal Microbiota in CD Subjects
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Catassi, C.; Fasano, A. Clincal Practice. Celiac disease. N. Engl. J. Med. 2012, 237, 2419–2426. [Google Scholar]
- Dotsenko, V.; Oittinen, M.; Taavela, J.; Popp, A.; Peräaho, M.; Staff, S.; Sarin, J.; Leon, F.; Isola, J.; Mäki, M.; et al. Genome-wide transcriptomic analysis of intestinal mucosa in celiac disese patients on a gluten-free diet and postgluten challenge. Cell. Mol. Gastroenterol. Hepatol. 2021, 11, 13–32. [Google Scholar] [CrossRef] [PubMed]
- Kramer, K.; Yeboah-Awudzi, M.; Magazine, N.; King, J.M.; Xu, Z.; Losso, J.N. Procyanidin B2 rich cocoa extracts inhibit inflammation in Caco-2 cell model of in vitro celiac disease by down-regulating interferon-gamma- or gliadin peptide 31-43-induced transglutaminase-2 and interleukin-15. J. Func. Foods 2019, 57, 112–1120. [Google Scholar] [CrossRef]
- Caio, G.; Volta, U.; Sapone, A.; Leffler, D.; Giorgio, R.; Catassi, C.; Fasano, A. Celiac disease: A comprehensive current review. BMC Med. 2019, 17, 142. [Google Scholar] [CrossRef]
- Elliot, C. The nutritional quality of gluten-free products for children. Pediatrics 2018, 142, e20180525. [Google Scholar] [CrossRef] [PubMed]
- Lionetti, E.; Antonucci, N.; Marinelli, M.; Bartolomei, B.; Franceschini, E.; Gatti, S.; Naspi Catassi, G.; Verma, A.K.; Monachesi, C.; Catassi, C. Nutritional status, dietary intake, and adherence to the Mediterranean diet on children with celiac disease on a gluten-free diet: A case-control prospective study. Nutrients 2019, 12, 143. [Google Scholar] [CrossRef]
- Larretxi, I.; Simon, E.; Benjumea, L.; Miranda, J.; Bustamante, M.A.; Lasa, A.; Euzaguirre, F.J.; Churruca, I. Gluten-free-rendered products contribute to imbalanced diets in children and adolescents with celiac disease. Eur. J. Nutr. 2019, 58, 775–783. [Google Scholar] [CrossRef]
- Arentz-Hansen, H.; Fleckenstein, B.; Molberg, O.; Scott, H.; Koning, F.; Jung, G.; Roepstorff, P.; Lundin, K.E.; Sollid, L.M. The molecular basis for oat intolerance in patients with celiac disease. PLoS Med. 2004, 1, e1. [Google Scholar] [CrossRef]
- Lundin, K.E.A.; Nilsen, E.M.; Scott, H.G.; Løberg, E.M.; Gjøen, A.; Bratlie, J.; Skar, V.; Mendez, E.; Løvik, A.; Kett, K. Oats induced villous atrophy in coeliac disease. Gut 2003, 52, 1649–1652. [Google Scholar] [CrossRef]
- Kaukinen, K.; Collin, P.; Huhtala, H.; Mäki, M. Long-term consumption of oats in adult celiac disease patients. Nutrients 2013, 5, 4380–4389. [Google Scholar] [CrossRef]
- Koskinen, O.; Villanen, M.; Korponay-Szabo, I.; Lindfords, K.; Mäki, M.; Kaukinen, K. Oats do no induce systemic or mucosa autoantibody response in children with coeliac disease. J. Pediatr. Gastroenterol. Nutr. 2009, 4, 559–565. [Google Scholar] [CrossRef]
- Lionetti, E.; Gatti, S.; Galeazzi, T.; Caporelli, N.; Francavilla, R.; Cucchiara, S.; Roggero, P.; Malamisura, B.; Iacono, G.; Tomarchiio, S.; et al. Safety of oats in children with celac disease: A double-blind, randomized, placebo-controlled trial. J. Pediatr. 2018, 194, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Sey, M.S.L.; Parfitt, J.; Gregor, J. Prospective study of clinical and histological safety of pure and uncontaminated Canadian oats in the management of celiac disease. J. Parenter. Enteral Nutr. 2011, 35, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Silano, M.; Peñas, E.; Uberti, F.; Manferdelli, S.; del Pinto, T.; Felli, C.; Budelli, A.; Vincentini, O.; Restani, P. Diversity of oat varieties in eliciting early inflammatory events in celiac disease. Eur. J. Nutr. 2014, 53, 1177–1186. [Google Scholar] [CrossRef] [PubMed]
- Aparicio-García, N.; Martínez-Villaluenga, C.; Frias, J.; Peñas, E. Sprouted oat as a potential gluten-free ingredient with enhanced nutritional and bioactive properties. Food Chem. 2021, 338, 127972. [Google Scholar] [CrossRef]
- Aparicio-García, N.; Martínez-Villaluenga, C.; Frias, J.; Peñas, E. Production and characterization of a novel gluten-free fermented beverage based on sprouted oat flour. Foods 2021, 10, 139. [Google Scholar] [CrossRef] [PubMed]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef] [PubMed]
- De Andrés, A.; Camarero, C.; Roy, G. Distal duodendenum versus duodenal bulb: Intraepihtelial lymphocytes have something to say in celiac disease diagnosis. Dig. Dis. Sci. 2015, 60, 1004–1009. [Google Scholar] [CrossRef] [PubMed]
- Castro, I.; Alba, C.; Fernández, L.; García, A.J.; Rodríguez, J.M. Culture-dependent and metataxonomic analysis of milk rom red deer (Cervus elaphus). Int. Dairy J. 2020, 102, 104610. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Van den Nieuwboer, M.; van Hemert, S.; Claassen, E.; de Vos, W.M. Lactobacillus plantarum WCFS1 and its host interaction: A dozen years after the genome. Microbial. Biotechnol. 2016, 9, 452–465. [Google Scholar] [CrossRef]
- Aydar, E.F.; Tutuncu, S.; Ozcelik, B. Plant-based milk substitutes: Bioactive compounds, conventional and novel processes, bioavailability studies, and health effects. J. Func. Foods 2020, 70, 103975. [Google Scholar] [CrossRef]
- Ballabio, C.; Uberti, F.; Manferdelli, S.; Vacca, E.; Boggini, G.; Radaelli, R.; Catassi, C.; Lionetti, E.; Peñas, E.; Restani, P. Molecular characterization of 36 oat varieties and in vitro assessment of their suitability for celiac diet. J. Cereal Sci. 2011, 54, 110–115. [Google Scholar] [CrossRef]
- Real, A.; Comino, I.; Lorenzo, L.; Merchán, F.; Gil-Humanes, J.; Giménez, M.J.; López-Casado, M.A.; Torres, M.I.; Cebolla, A.; Sousa, C.; et al. Molecular and immunological characerization of gluten proteins isolated from oat cultivars that differ in toxicity for celiac disease. PLoS ONE 2012, 7, e48365. [Google Scholar] [CrossRef] [PubMed]
- Giménez, M.J.; Real, A.; García-Molina, M.D.; Sousa, C.; Barro, F. Characterization of celiac disease related oat proteins: Bases for the development of high quality oat varieties suitable for celiac patients. Sci. Rep. 2017, 7, 42588. [Google Scholar] [CrossRef]
- Luciani, A.; Villella, V.R.; Vasaturo, A.; Giardino, I.; Pettoello-Mantovani, M.; Guido, S.; Cexus, O.N.; Peake, N.; Londei, M.; Quaratino, S.; et al. Lysosomal accumulation of gliadin p31–43 peptide induces oxidative stress and tissue transglutaminase-mediated PPARgamma downregulation intestinal epithelial cells and coeliac mucosa. Gut 2010, 59, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Wang, L.; Wang, R.; Luo, X.; Li, Y.; Li, J.; Li, Y.; Chen, Z. Phenolic contents, cellular antioxidant activity and antiproliferative capacity of different varieties of oats. Food Chem. 2018, 239, 260–267. [Google Scholar] [CrossRef]
- Lee, J.H.; Lee, B.-K.; Park, H.-H.; Lee, B.W.; Woo, K.S.; Kim, H.-J.; Han, S.-I.; Lee, Y.Y. Oat germination and ultrafiltration process improves the polyphenol and avenanthramide contents with protective effect in oxidative-damaged HepG2 cells. J. Food Biochem. 2019, 43, e12799. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Wen, L.; Sun, J.; Bai, W.; Jiao, R.; Hu, Y.; Peng, X.; He, Y.; Ou, S. Cytoprotective mechanism of ferulic acid against high glucose-induced oxidative stress in cardiomyocytes and hepatocytes. Food Nutr. Res. 2016, 60, 30323. [Google Scholar] [CrossRef]
- Wang, C.; Eskiw, C.H. Cytoprotective effects of Avenathramide C against oxidative and inflammatory stress in normal human dermal fibroblasts. Sci. Rep. 2019, 9, 2932. [Google Scholar] [CrossRef]
- Deswal, A.; Deora, N.S.; Mishra, H.N. Effect of concentration and temperature on the rheological properties of oat milk. Food Bioproc. Tech. 2014, 7, 2451–2459. [Google Scholar] [CrossRef]
- Fitrotin, U.; Utami, T.; Hastuti, P.; Santoso, U. Antioxidant properties of fermented sesame milk using Lactobacillus plantarum Dad 13. Int. Res. J. Biol. Sci. 2015, 4, 56–61. [Google Scholar]
- Alasalvar, C.; Bolling, B.W. Review of nut phytochemicals, fat-soluble bioactives, antioxidant components and health effects. Br. J. Nut. 2015, 113, S68–S78. [Google Scholar] [CrossRef] [PubMed]
- Suchecka, D.; Błaszczyk, K.; Harasym, J.; Gudej, S.; Wilczak, J.; Gromadzka-Ostrowska, J. Impact of purified oat 1-3,1-4-β-d-glucan of different molecular weight on alleviation of inflammation parameters during gastritis. J. Func. Foods. 2017, 28, 11–18. [Google Scholar] [CrossRef]
- Chu, Y.-F.; Wise, M.L.; Gulvady, A.A.; Chang, T.; Kendra, D.F.; Jan-Willem van Klinken, B.; Shi, Y.; O’Shea, M. In vitro antioxidant capacity and anti-inflammatory activity of seven common oats. Food Chem. 2013, 139, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Charlton, K.E.; Tapsell, L.C.; Batterham, M.J.; O’Shea, J.; Thorne, R.; Beck, E.; Tosh, S.M. Effect of 6 weeks’ consumption of β-glucan-rich oat products on cholesterol levels in mildly hypercholesterolaemic overweight adults. Brit. J. Nutr. 2012, 107, 1037–1047. [Google Scholar] [CrossRef]
- Schuster, J.; Benincá, G.; Vitorazzi, R.; Morelo Dal Bosco, S. Effects of oats on lipid profile, insulin resistance and weight loss. Nutr. Hosp. 2015, 32, 2111–2116. [Google Scholar] [PubMed]
- Geliebter, A.; Grillot, C.L.; Aviram-Friedman, R.; Haq, S.; Yahav, E.; Hashim, S.A. Effects of oatmeal and corn flakes cereal breakfasts on satiety, gastric emptying, glucose, and appetite-related hormones. Ann. Nutr. Metab. 2015, 66, 93–103. [Google Scholar] [CrossRef]
- Kemppainen, T.; Heikkinen, M.; Ristikankare, M.; Kosma, V.-M.; Julkunen, R. Effect of unkilned and large amounts of oats on nutritional state of celiac patients in remission. e-Spen 2009, 4, e30–e34. [Google Scholar] [CrossRef][Green Version]
- Zhu, X.; Sun, X.; Wang, M.; Zhang, C.; Cao, Y.; Mo, G.; Liang, J.; Zhu, S. Quantitative assessment of the effects of beta-glucan consumption on serum lipid profile and glucose level in hypercholesterolemic subjects. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 714–723. [Google Scholar] [CrossRef]
- Andersson, K.E.; Chawade, A.; Thuresson, N.; Rascon, A.; Öste, R.; Sterner, O.; Olsson, O.; Hellstrand, P. Wholegrain oat diet changes the expression of genes associated with intestinal bile acid transport. Mol. Nutr. Food Res. 2017, 61, 1600874. [Google Scholar] [CrossRef]
- Bodkhe, R.; Shetty, S.A.; Dhotre, D.P.; Verma, A.K.; Bhatia, K.; Mishra, A.; Kaur, G.; Pande, P.; Bangarusamy, D.K.; Santosh, B.P.; et al. Comparison of small gut and whole gut microbiota of first-degree relatives with adult celiac disease patients and controls. Front. Microbiol. 2019, 10, 164. [Google Scholar] [CrossRef]
- Pecora, F.; Persico, F.; Gismondi, P.; Fornaroli, F.; Luliano, K.S.; de’Angelis, G.L.; Esposito, S.D. Gut microbiota in celiac disease: Is there anay role for probiotics? Front. Immunol. 2020, 11, 957. [Google Scholar] [CrossRef] [PubMed]
- Nylund, L.; Hakkola, S.; Lahti, L.; Salminen, S.; Kalliomäki, M.; Yang, B.; Linderborg, K.M. Diet, perceived intestinal well-being and compositions of fecal microbiota and short chain fatty acids in oat-using subjects with celiac disease or gluten sensitivity. Nutrients 2020, 12, 2570. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, A.; Cerrone, R.; Capobianco, D.; Filardo, S.; Mancini, P.; Zanni, F.; Fanelli, S.; Mastromarino, P.; Mosca, L. A probiotic preparation hydrolyzes gliadin and protects intestinal cells from the toxicity of pro-inflammatory peptides. Nutrients 2020, 12, 495. [Google Scholar] [CrossRef] [PubMed]
- Ze, X.; Duncan, S.H.; Louis, P.; Flint, H.J. Ruminococcus bromii is a keystone species for the degradation of resistant starch in the human colon. ISME J. 2012, 6, 1535. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Baseline Characteristics | Control Group (n = 5) | SOFB Group (n = 5) | p |
|---|---|---|---|
| Gender (male/female) | 2/3 | 2/3 | |
| Age (year) | 45.20 ± 12.32 | 31.80 ± 12.52 | 0.126 |
| Duration GF diet (year) | 8.30 ± 9.36 | 8.80 ± 6.69 | 0.530 |
| Weight (Kg) | 72.20 ± 14.81 | 62.80 ± 5.80 | 0.168 |
| Height (m) | 1.68 ± 0.08 | 1.68 ± 0.08 | 0.727 |
| Body mass index (Kg/m2) | 25.54 ± 4.00 | 21.50 ± 1.99 | 0.078 |
| IgA Levels | Control Group | SOFB Group | p |
|---|---|---|---|
| Total serum IgA (mg/dL) | |||
| 0 months | 200.00 ± 67.99 (146.0–288.0) a | 197.20 ± 60.26 (104.0–271.0) a | 1.000 |
| 2 months | 188.40 ± 70.18 (126.0–286.0) a | 192.34 ± 61.95 (99.7–270.0) a | 1.000 |
| 4 months | 196.00 ± 67.66 (143.0–288.0) a | 200.80 ± 66.38 (104.0–290.0) a | 0.841 |
| 6 months | 202.20 ± 79.90 (1.26–311.0) a | 201.80 ± 67.15 (102.0–290.0) a | 1.000 |
| IgA-tTG (U/mL) | |||
| 0 months | 1.76 ± 0.62 (0.7–2.4) a | 2.72 ± 3.29 (0.5–8.5) a | 0.841 |
| 2 months | 1.38 ± 0.62 (0.5–2.0) a | 2.64 ± 3.03 (0.9–8.0) a | 1.000 |
| 4 months | 1.83 ± 0.54 (1.0–2.4) a | 2.39 ± 2.41 (1.0–6.7) a | 0.548 |
| 6 months | 1.22 ± 0.56 (0.5–1.8) a | 2.55 ± 2.74 (0.2–7.2) a | 0.421 |
| IEL Levels | Months | Control Group | SOFB Group | p | |
|---|---|---|---|---|---|
| Second duodenal portion | Total IEL | 0 | 10.80 ± 2.83 (6.80–12.90) a | 19.90 ± 9.84 (5.20–25.80) a | 0.343 |
| 6 | 8.70 ± 4.04 (3.90–12.10) a | 8.24 ± 6.37 (2.50–0.40) a | 0.841 | ||
| TCR γδ+ IEL | 0 | 28.28 ± 12.41 (19.40–46.50) a | 24.32 ± 9.55 (17.00–40.90) a | 0.413 | |
| 6 | 22.98 ± 6.77 (16.30–31.60) a | 29.16 ± 12.20 (16.50–46.70) a | 0.548 | ||
| CD3−CD103+ IEL | 0 | 6.10 ± 5.74 (0.30–13.20) a | 3.12 ± 3.41 (0.40–8.40) a | 0.730 | |
| 6 | 7.44 ± 9.09 (1.20–23.50) a | 3.20 ± 4.15 (1.10–10.60) a | 0.222 | ||
| Duodenal bulb | Total IEL | 0 | 8.30 ± 4.51 (4.20–14.40) a | 11.57 ± 7.75 (5.9–20.40) a | 0.533 |
| 6 | 5.83 ± 1.28 (4.40–7.40) a | 3.97 ± 2.35 (1.40–6.00) a | 0.800 | ||
| TCR γδ+ IEL | 0 | 19.00 ± 15.67 (6.90–40.90) a | 24.46 ± 12.12 (14.90–38.10) a | 0.630 | |
| 6 | 20.80 ± 17.81 (8.20–47.20) a | 24.70 ± 10.07 (14.30–34.40) a | 0.533 | ||
| CD3−CD103+ IEL | 0 | 3.23 ± 1.61 (1.80–4.90) a | 3.30 ± 4.33 (0.70–8.30) a | 0.629 | |
| 6 | 1.98 ± 0.86 (1.20–3.20) a | 1.96 ± 0.83 (1.30–2.90) a | 0.857 |
| IgA Levels | Control Group | SOFB Group | p |
|---|---|---|---|
| Body weight (Kg) | |||
| 0 months | 72.2 ± 14.8 (54.0–95.0) a | 62.8 ± 5.8 (54.0–68.0) a | 0.151 |
| 2 months | 73.5 ± 16.8 (54.5–100.0) a | 62.7 ± 6.1 (52.5–68.0) a | 0.151 |
| 4 months | 73.5 ± 16.4 (54.0–99.0) a | 63.1 ± 6.4 (52.5–69.0) a | 0.095 |
| 6 months | 73.5 ± 16.7 (52.5–99.0) a | 62.8 ± 7.2 (51.0–70.0) a | 0.151 |
| Glucose (μg/dL) | |||
| 0 months | 89.4 ± 7.4 (79.0–97.0) a | 80.4 ± 6.9 (72.0–87.0) a | 0.151 |
| 2 months | 83.8 ± 6.5 (73.0–90.0) a | 76.6 ± 10.7 (64.0–92.0) a | 0.222 |
| 4 months | 88.2 ± 12.1 (75.0–106.0) a | 83.2 ± 1.7 (82.0–86.0) a | 0.690 |
| 6 months | 84.2 ± 12.0 (71.0–95.0) a | 80.8 ± 5.9 (71.0–85.0) a | 0.421 |
| Iron (mg/dL) | |||
| 0 months | 98.6 ± 22.1 (79.0–132.0) a | 100.0 ± 26.5 (73.0–127.0) a | 0.841 |
| 2 months | 86.2 ± 16.6 (64.0–108.0) a | 104.4 ± 33.8 (63.0–145.0) a | 0.420 |
| 4 months | 88.0 ± 27.2 (53.0–119.0) a | 69.6 ± 43.8 (38.0–145.0) a | 0.222 |
| 6 months | 101.6 ± 8.0 (95.0–114.0) a | 97.4 ± 61.7 (44.0–189.0) a | 0.690 |
| Ferritin (ng/mL) | |||
| 0 months | 62.9 ± 49.2 (20.9–143.5) a | 80.9 ± 66.5 (24.1–167.9) a | 0.917 |
| 2 months | 73.6 ± 59.0 (17.3–164.9) a | 72.0 ± 60.1 (18.4–154.3) a | 0.917 |
| 4 months | 72.1 ± 81.2 (19.1–214.0) a | 76.5 ± 69.4 (15.6–177.7) a | 0.917 |
| 6 months | 80.5 ± 96.7 (13.1–247.3) a | 76.3 ± 69.2 (17.3–177.7) a | 0.754 |
| Vitamin B12 (pg/mL) | |||
| 0 months | 416.2 ± 143.8 (261.0–610.0) a | 407.8 ± 120.1 (261.0–589.0) a | 0.690 |
| 2 months | 428.2 ± 164.2 (246.0–644.0) a | 398.0 ± 136.8 (217.0–589.0) a | 1.000 |
| 4 months | 449.0 ± 202.7 (167.0–610.0) a | 393.8 ± 123.3 (218.0–544.0) a | 0.548 |
| 6 months | 387.6 ± 121.4 (261.0–527.0) a | 417.2 ± 186.7 (274.0–728.0) a | 0.841 |
| Folic acid (ng/mL) | |||
| 0 months | 7.5 ± 3.7 (3.7–13.2) a | 5.1 ± 2.6 (2.8–9.4) a | 0.310 |
| 2 months | 9.8 ± 4.2 (6.1–15.9) a | 5.7 ± 3.0 (2.1–9.0) a | 0.222 |
| 4 months | 6.6 ± 1.9 (4.5–8.8) a | 4.9 ± 2.4 (2.1–8.7) a | 0.220 |
| 6 months | 8.0 ± 2.2 (5.4–10.7) a | 5.1 ± 2.1 (3.1–8.7) a | 0.032 |
| Hemoglobin (g/dL) | |||
| 0 months | 14.4 ± 1.3 (12.5–16.2) a | 14.1 ± 0.8 (13.3–15.2) a | 0.690 |
| 2 months | 14.4 ± 1.5 (12.8–16.7) a | 12.0 ± 4.3 (4.4–14.3) a | 0.310 |
| 4 months | 14.3 ± 1.3 (12.8–16.0) a | 12.1 ± 4.1 (4.9–14.3) a | 0.548 |
| 6 months | 14.2 ± 1.9 (11.9–16.4) a | 13.9 ± 0.4 (13.3–14.3) a | 0.841 |
| Triglycerides (mg/mL) | |||
| 0 months | 133.8 ± 43.6 (64.0–174.0) a | 53.6 ± 19.3 (34.0–82.0) a | 0.016 |
| 2 months | 110.8 ± 47.8 (68.0–190.0) a | 69.6 ± 46.0 (33.0–150.0) a | 0.095 |
| 4 months | 94.8 ± 26.9 (64.0–130.0) a | 81.0 ± 48.0 (30.0–150.0) a | 0.069 |
| 6 months | 116.4 ± 59.7 (51.0–207.0) a | 51.6 ± 20.4 (41.0–88.0) a | 0.016 |
| Total cholesterol (mg/dL) | |||
| 0 months | 198.0 ± 41.2 (147.0–262.0) a | 147.2 ± 22.4 (119.0–170.0) a | 0.060 |
| 2 months | 188.2 ± 25.9 (158.0–215.0) a | 132.0 ± 28.4 (94.0–163.0) a | 0.016 |
| 4 months | 189.0 ± 42.3 (147.0–260.0) a | 137.4 ± 18.32 (115.0–158.0) a | 0.032 |
| 6 months | 199.0 ± 30.2 (163.0–235.0) a | 146.6 ± 21.4 (124.0–175.0) a | 0.016 |
| Phylum/Genus | Control Group | SOFB Group | p | ||
|---|---|---|---|---|---|
| n (%) | Median (IQR) | n (%) # | Median (IQR) | ||
| Firmicutes | 20 (100%) | 88.78 (80.85–92.2) | 20 (100%) | 91.46 (89.03–93.52) | 0.081 |
| Blautia | 20 (100%) | 9.79 (7–12.58) | 20 (100%) | 10.07 (6.32–14.58) | 0.95 |
| Subdoligranulum | 20 (100%) | 2.72 (1.42–6.05) | 20 (100%) | 6.99 (4.72–8.93) | 0.002 |
| Faecalibacterium | 16 (80%) | 3.83 (0.64–7.9) | 20 (100%) | 3.85 (1.61–8.07) | 0.6 |
| Agathobacter | 20 (100%) | 2.04 (1.58–5.25) | 20 (100%) | 1.96 (1.18–7.05) | 0.82 |
| Dorea | 20 (100%) | 3.19 (2.56–5.56) | 20 (100%) | 3.87 (1.6–5.11) | 0.56 |
| Anaerostipes | 20 (100%) | 3.41 (1.19–8.27) | 20 (100%) | 1.34 (0.6–3.13) | 0.02 |
| Streptococcus | 20 (100%) | 1.9 (1.23–4.22) | 20 (100%) | 1.11 (0.54–3.44) | 0.15 |
| Erysipelotrichaceae | 20 (100%) | 4.02 (1.75–4.83) | 19 (95%) | 2.61 (0.66–4.04) | 0.12 |
| Ruminococcus | 6 (30%) | <0.01 (<0.01–0.01) | 17 (85%) | 5.28 (0.16–7.17) | <0.001 |
| Lactobacillus | 15 (75%) | 0.04 (<0.01–0.13) | 17 (85%) | 0.25 (0.05–3.11) | 0.028 |
| Clostridium | 15 (75%) | 0.4 (0.01–1.19) | 20 (100%) | 0.38 (0.18–2.08) | 0.25 |
| Romboutsia | 17 (85%) | 0.78 (0.38–1.48) | 20 (100%) | 1.02 (0.43–2.62) | 0.39 |
| Holdemanella | 4 (20%) | <0.01 (<0.01–<0.01) | 8 (40%) | <0.01 (<0.01–1.56) | 0.43 |
| Monoglobus | 16 (80%) | 1.29 (0.1–1.89) | 19 (95%) | 0.95 (0.3–2.32) | 0.64 |
| Coprococcus | 17 (85%) | 1.35 (0.98–1.8) | 20 (100%) | 1.8 (1.17–2.1) | 0.13 |
| Catenibacterium | 5 (25%) | <0.01 (<0.01–<0.01) | 4 (20%) | <0.01 (<0.01–<0.01) | 0.81 |
| Actinobacteria | 20 (100%) | 5.69 (4.41–6.87) | 20 (100%) | 5.43 (3.58–6.51) | 0.24 |
| Bifidobacterium | 20 (100%) | 1.96 (0.79–4.09) | 19 (95%) | 1.72 (0.52–3.16) | 0.58 |
| Bacterioidota | 20 (100%) | 3.01 (0.86–11.09) | 19 (95%) | 2.17 (0.58–4.13 | 0.3 |
| Bacteroides | 18 (90%) | 2.5 (0.61–7.67) | 18 (90%) | 1.24 (0.25–2.58) | 0.16 |
| Proteobacteria | 18 (90%) | 0.17 (0.05–0.67) | 20 (100%) | 0.25 (0.14–0.64) | 36 |
| Fusobacteriota | 10 (50%) | 0.01 (<0.01–0.11) | 3 (15%) | <0.01 (<0.01–<0.01) | 0.011 |
| Minor phyla | 19 (95%) | 0.07 (0.02–0.21) | 16 (80%) | 0.06 (0.01–0.6) | 0.86 |
| Minor genera | 20 (100%) | 15.44 (14.57–21.34) | 20 (100%) | 17.55 (13.63–21.97) | 0.97 |
| Unclassified genera | 20 (100%) | 17.55 (13.63–21.97) | 20 (100%) | 13.3 (10.94–14.75) | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aparicio-García, N.; Martínez-Villaluenga, C.; Frias, J.; Crespo Perez, L.; Fernández, C.F.; Alba, C.; Rodríguez, J.M.; Peñas, E. A Novel Sprouted Oat Fermented Beverage: Evaluation of Safety and Health Benefits for Celiac Individuals. Nutrients 2021, 13, 2522. https://doi.org/10.3390/nu13082522
Aparicio-García N, Martínez-Villaluenga C, Frias J, Crespo Perez L, Fernández CF, Alba C, Rodríguez JM, Peñas E. A Novel Sprouted Oat Fermented Beverage: Evaluation of Safety and Health Benefits for Celiac Individuals. Nutrients. 2021; 13(8):2522. https://doi.org/10.3390/nu13082522
Chicago/Turabian StyleAparicio-García, Natalia, Cristina Martínez-Villaluenga, Juana Frias, Laura Crespo Perez, Cristina Fernández Fernández, Claudio Alba, Juan Miguel Rodríguez, and Elena Peñas. 2021. "A Novel Sprouted Oat Fermented Beverage: Evaluation of Safety and Health Benefits for Celiac Individuals" Nutrients 13, no. 8: 2522. https://doi.org/10.3390/nu13082522
APA StyleAparicio-García, N., Martínez-Villaluenga, C., Frias, J., Crespo Perez, L., Fernández, C. F., Alba, C., Rodríguez, J. M., & Peñas, E. (2021). A Novel Sprouted Oat Fermented Beverage: Evaluation of Safety and Health Benefits for Celiac Individuals. Nutrients, 13(8), 2522. https://doi.org/10.3390/nu13082522