Nutrient Timing: A Garage Door of Opportunity?

 , ,
, , {kind=link}
Abstract
1. Introduction
2. Pre-Exercise Nutrition
3. Intra-Exercise Nutrition
4. Post-Exercise Nutrition
5. Additional Timing Considerations
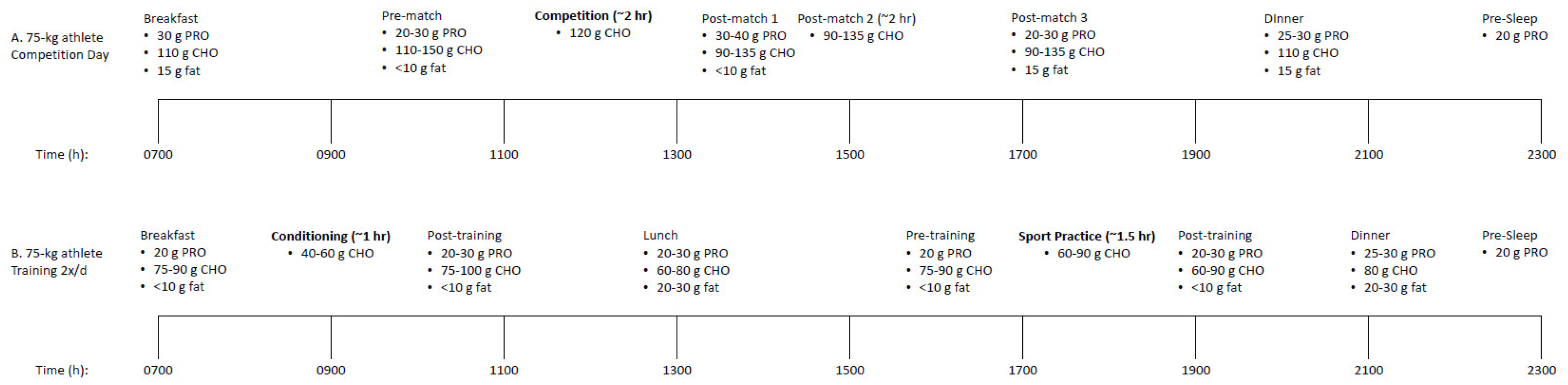
6. Practical Implications
Author Contributions
Funding
Conflicts of Interest
References
- Kerksick, C.M.; Arent, S.; Schoenfeld, B.J.; Stout, J.R.; Campbell, B.; Wilborn, C.D.; Taylor, L.; Kalman, D.; Smith-Ryan, A.E.; Kreider, R.B.; et al. International society of sports nutrition position stand: Nutrient timing. J. Int. Soc. Sport Nutr. 2017, 14, 33. [Google Scholar] [CrossRef]
- Karlsson, J.; Saltin, B. Diet, muscle glycogen, and endurance performance. J. Appl. Physiol. 1971, 31, 203–206. [Google Scholar] [CrossRef]
- Ivy, J.L.; Katz, A.L.; Cutler, C.L.; Sherman, W.M.; Coyle, E.F. Muscle glycogen synthesis after exercise: Effect of time of carbohydrate ingestion. J. Appl. Physiol. 1988, 64, 1480–1485. [Google Scholar] [CrossRef]
- Hawley, J.A.; Burke, L.M. Effect of meal frequency and timing on physical performance. Br. J. Nutr. 1997, 77 (Suppl. 1), S91–S103. [Google Scholar] [CrossRef]
- Gollnick, P.D.; Armstrong, R.B.; Sembrowich, W.L.; Shepherd, R.E.; Saltin, B. Glycogen depletion pattern in human skeletal muscle fibers after heavy exercise. J. Appl. Physiol. 1973, 34, 615–618. [Google Scholar] [CrossRef]
- Gollnick, P.D.; Piehl, K.; Saltin, B. Selective glycogen depletion pattern in human muscle fibres after exercise of varying intensity and at varying pedalling rates. J. Physiol. 1974, 241, 45–57. [Google Scholar] [CrossRef]
- Ivy, J.L.; Portman, R.M. Nutrient Timing: The Future of Sports Nutrition; Basic Health Publications: Laguna Beach, CA, USA, 2004; p. 211. [Google Scholar]
- Borer, K.T. Advanced Exercise Endocrinology; Human Kinetics: Champaign, IL, USA, 2012. [Google Scholar]
- Richter, E.A.; Hargreaves, M. Exercise, GLUT4, and skeletal muscle glucose uptake. Physiol. Rev. 2013, 93, 993–1017. [Google Scholar] [CrossRef] [PubMed]
- Ivy, J.L.; Kuo, C.H. Regulation of GLUT4 protein and glycogen synthase during muscle glycogen synthesis after exercise. Acta Physiol. Scand. 1998, 162, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Brooks, G.A.; Fahey, T.D.; Baldwin, K.M. Exercise Physiology: Human Bioenergetics and Its Applications, 4th ed.; McGraw Hill: New York, NY, USA, 2005. [Google Scholar]
- Atherton, P.J.; Smith, K. Muscle protein synthesis in response to nutrition and exercise. J. Physiol. 2012, 590, 1049–1057. [Google Scholar] [CrossRef]
- Cribb, P.J.; Hayes, A. Effects of supplement timing and resistance exercise on skeletal muscle hypertrophy. Med. Sci. Sports Exerc. 2006, 38, 1918–1925. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, J.R.; Ratamess, N.A.; Tranchina, C.P.; Rashti, S.L.; Kang, J.; Faigenbaum, A.D. Effect of protein-supplement timing on strength, power, and body-composition changes in resistance-trained men. Int. J. Sport Nutr. Exerc. Metab. 2009, 19, 172–185. [Google Scholar] [CrossRef] [PubMed]
- Koopman, R.; Beelen, M.; Stellingwerff, T.; Pennings, B.; Saris, W.H.; Kies, A.K.; Kuipers, H.; van Loon, L.J. Coingestion of carbohydrate with protein does not further augment postexercise muscle protein synthesis. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E833–E842. [Google Scholar] [CrossRef]
- Everman, S.; Meyer, C.; Tran, L.; Hoffman, N.; Carroll, C.C.; Dedmon, W.L.; Katsanos, C.S. Insulin does not stimulate muscle protein synthesis during increased plasma branched-chain amino acids alone but still decreases whole body proteolysis in humans. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E671–E677. [Google Scholar] [CrossRef]
- Aragon, A.A.; Schoenfeld, B.J. Nutrient timing revisited: Is there a post-exercise anabolic window? J. Int. Soc. Sport Nutr. 2013, 10, 5. [Google Scholar] [CrossRef] [PubMed]
- Schoenfeld, B.J.; Aragon, A.A. Is there a postworkout anabolic window of opportunity for nutrient consumption? Clearing up controversies. J. Orthop. Sports Phys. Ther. 2018, 48, 911–914. [Google Scholar] [CrossRef] [PubMed]
- Schoenfeld, B.J.; Aragon, A.A.; Krieger, J.W. The effect of protein timing on muscle strength and hypertrophy: A meta-analysis. J. Int. Soc. Sport Nutr. 2013, 10, 53. [Google Scholar] [CrossRef]
- Ivy, J.L.; Ferguson-Stegall, L.M. Nutrient Timing. Am. J. Lifestyle Med. 2014, 8, 246–259. [Google Scholar] [CrossRef]
- Burke, L.M.; Castell, L.M.; Casa, D.J.; Close, G.L.; Costa, R.J.S.; Desbrow, B.; Halson, S.L.; Lis, D.M.; Melin, A.K.; Peeling, P.; et al. International association of athletics federations consensus statement 2019: Nutrition for athletics. Int. J. Sport Nutr. Exerc. Metab. 2019, 29, 73–84. [Google Scholar] [CrossRef]
- Bytomski, J.R. Fueling for performance. Sports Health 2018, 10, 47–53. [Google Scholar] [CrossRef]
- Fritzen, A.M.; Lundsgaard, A.M.; Kiens, B. Dietary fuels in athletic performance. Annu. Rev. Nutr. 2019, 39, 45–73. [Google Scholar] [CrossRef]
- Hector, A.J.; Phillips, S.M. Protein recommendations for weight loss in elite athletes: A focus on body composition and performance. Int. J. Sport Nutr. Exerc. Metab. 2018, 28, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Stellingwerff, T.; Morton, J.P.; Burke, L.M. A framework for periodized nutrition for athletics. Int. J. Sport Nutr. Exerc. Metab. 2019, 29, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Cintineo, H.P.; Arent, M.A.; Antonio, J.; Arent, S.M. Effects of protein supplementation on performance and recovery in resistance and endurance training. Front. Nutr. 2018, 5, 83. [Google Scholar] [CrossRef]
- Ormsbee, M.J.; Bach, C.W.; Baur, D.A. Pre-exercise nutrition: The role of macronutrients, modified starches and supplements on metabolism and endurance performance. Nutrients 2014, 6, 1782–1808. [Google Scholar] [CrossRef]
- McArdle, W.D.; Katch, F.I.; Katch, V.L. Exercise Physiology: Nutrition, Energy, and Human Performance; Lippincott, Williams, and Wilkins: Philadelphia, PA, USA, 2014. [Google Scholar]
- Gropper, S.S.; Smith, J.L. Advanced Nutrition and Human Metabolism, 7th ed.; Cengage: Independence, KY, USA, 2018. [Google Scholar]
- Burke, L.M. Re-examining high-fat diets for sports performance: Did we call the ‘nail in the coffin’ too soon? Sports Med. 2015, 45 (Suppl. 1), S33–S49. [Google Scholar] [CrossRef] [PubMed]
- Jeukendrup, A.E.; Killer, S.C. The myths surrounding pre-exercise carbohydrate feeding. Ann. Nutr. Metab. 2010, 57 (Suppl. 2), 18–25. [Google Scholar] [CrossRef]
- Brouns, F.; Rehrer, N.J.; Saris, W.H.; Beckers, E.; Menheere, P.; ten Hoor, F. Effect of carbohydrate intake during warming-up on the regulation of blood glucose during exercise. Int. J. Sports Med. 1989, 10 (Suppl. 1), S68–S75. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, T.M.; Fox, E.L. The effect of muscular work upon the respiratory exchange of man after the ingestion of glucose and of fructose. Arbeitsphysiologie 1931, 4, 532–569. [Google Scholar]
- Haldi, J.; Wynn, W. Observations on efficiency of swimmers as related to some changes in pre-exercise nutriment. J. Nutr. 1946, 31, 525–532. [Google Scholar] [CrossRef]
- Hargreaves, M.; Costill, D.L.; Fink, W.J.; King, D.S.; Fielding, R.A. Effect of pre-exercise carbohydrate feedings on endurance cycling performance. Med. Sci. Sports Exerc. 1987, 19, 33–36. [Google Scholar] [CrossRef]
- Sherman, W.M.; Brodowicz, G.; Wright, D.A.; Allen, W.K.; Simonsen, J.; Dernbach, A. Effects of 4 h preexercise carbohydrate feedings on cycling performance. Med. Sci. Sports Exerc. 1989, 21, 598–604. [Google Scholar] [CrossRef] [PubMed]
- Sherman, W.M.; Peden, M.C.; Wright, D.A. Carbohydrate feedings 1 h before exercise improves cycling performance. Am. J. Clin. Nutr. 1991, 54, 866–870. [Google Scholar] [CrossRef] [PubMed]
- Achten, J.; Jeukendrup, A.E. The effect of pre-exercise carbohydrate feedings on the intensity that elicits maximal fat oxidation. J. Sports Sci. 2003, 21, 1017–1024. [Google Scholar] [CrossRef]
- Haff, G.G.; Lehmkuhl, M.J.; McCoy, L.B.; Stone, M.H. Carbohydrate supplementation and resistance training. J. Strength Cond. Res. 2003, 17, 187–196. [Google Scholar] [CrossRef]
- Escobar, K.A.; VanDusseldorp, T.A.; Kerksick, C.M. Carbohydrate intake and resistance-based exercise: Are current recommendations reflective of actual need? Br. J. Nutr. 2016, 116, 2053–2065. [Google Scholar] [CrossRef]
- Camera, D.M.; West, D.W.; Burd, N.A.; Phillips, S.M.; Garnham, A.P.; Hawley, J.A.; Coffey, V.G. Low muscle glycogen concentration does not suppress the anabolic response to resistance exercise. J. Appl. Physiol. 2012, 113, 206–214. [Google Scholar] [CrossRef]
- Burke, L.M.; Hawley, J.A.; Ross, M.L.; Moore, D.R.; Phillips, S.M.; Slater, G.R.; Stellingwerff, T.; Tipton, K.D.; Garnham, A.P.; Coffey, V.G. Preexercise aminoacidemia and muscle protein synthesis after resistance exercise. Med. Sci. Sports Exerc. 2012, 44, 1968–1977. [Google Scholar] [CrossRef] [PubMed]
- Fujita, S.; Dreyer, H.C.; Drummond, M.J.; Glynn, E.L.; Volpi, E.; Rasmussen, B.B. Essential amino acid and carbohydrate ingestion before resistance exercise does not enhance postexercise muscle protein synthesis. J. Appl. Physiol. 2009, 106, 1730–1739. [Google Scholar] [CrossRef]
- Tipton, K.D.; Rasmussen, B.B.; Miller, S.L.; Wolf, S.E.; Owens-Stovall, S.K.; Petrini, B.E.; Wolfe, R.R. Timing of amino acid-carbohydrate ingestion alters anabolic response of muscle to resistance exercise. Am. J. Physiol. Endocrinol. Metab. 2001, 281, E197–E206. [Google Scholar] [CrossRef]
- Dalbo, V.J.; Roberts, M.D.; Hassell, S.; Kerksick, C.M. Effects of pre-exercise feeding on serum hormone concentrations and biomarkers of myostatin and ubiquitin proteasome pathway activity. Eur. J. Nutr. 2013, 52, 477–487. [Google Scholar] [CrossRef] [PubMed]
- Schoenfeld, B.J.; Aragon, A.; Wilborn, C.; Urbina, S.L.; Hayward, S.E.; Krieger, J. Pre-versus post-exercise protein intake has similar effects on muscular adaptations. PeerJ 2017, 5, e2825. [Google Scholar] [CrossRef]
- Chen, Y.J.; Wong, S.H.; Wong, C.K.; Lam, C.W.; Huang, Y.J.; Siu, P.M. The effect of a pre-exercise carbohydrate meal on immune responses to an endurance performance run. Br. J. Nutr. 2008, 100, 1260–1268. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.A.; Koch, A.J.; Benedict, S.H.; Potteiger, J.A. Influence of carbohydrate ingestion on cytokine responses following acute resistance exercise. Int. J. Sport Nutr. Exerc. Metab. 2003, 13, 454–465. [Google Scholar] [CrossRef]
- Kraemer, W.J.; Ratamess, N.A. Hormonal responses and adaptations to resistance exercise and training. Sports Med. 2005, 35, 339–361. [Google Scholar] [CrossRef] [PubMed]
- Mager, M.; Iampietro, P.F.; Goldman, R.F. The effect of supplementary feeding on plasma free fatty acids during work. Metabolism 1964, 13, 823–830. [Google Scholar] [CrossRef]
- Bergstrom, J.; Hermansen, L.; Hultman, E.; Saltin, B. Diet, muscle glycogen and physical performance. Acta Physiol. Scand. 1967, 71, 140–150. [Google Scholar] [CrossRef]
- Vollestad, N.K.; Tabata, I.; Medbo, J.I. Glycogen breakdown in different human muscle fibre types during exhaustive exercise of short duration. Acta Physiol. Scand. 1992, 144, 135–141. [Google Scholar] [CrossRef]
- Ortenblad, N.; Nielsen, J.; Saltin, B.; Holmberg, H.C. Role of glycogen availability in sarcoplasmic reticulum Ca2+ kinetics in human skeletal muscle. J. Physiol. 2011, 589, 711–725. [Google Scholar] [CrossRef] [PubMed]
- Jeukendrup, A.E.; Jentjens, R. Oxidation of carbohydrate feedings during prolonged exercise: Current thoughts, guidelines and directions for future research. Sports Med. 2000, 29, 407–424. [Google Scholar] [CrossRef] [PubMed]
- Jentjens, R.L.; Moseley, L.; Waring, R.H.; Harding, L.K.; Jeukendrup, A.E. Oxidation of combined ingestion of glucose and fructose during exercise. J. Appl. Physiol 2004, 96, 1277–1284. [Google Scholar] [CrossRef] [PubMed]
- Cermak, N.M.; van Loon, L.J. The use of carbohydrates during exercise as an ergogenic aid. Sports Med. 2013, 43, 1139–1155. [Google Scholar] [CrossRef]
- Currell, K.; Jeukendrup, A.E. Superior endurance performance with ingestion of multiple transportable carbohydrates. Med. Sci. Sports Exerc. 2008, 40, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Jentjens, R.L.; Jeukendrup, A.E. High rates of exogenous carbohydrate oxidation from a mixture of glucose and fructose ingested during prolonged cycling exercise. Br. J. Nutr. 2005, 93, 485–492. [Google Scholar] [CrossRef]
- Jeukendrup, A.E.; Moseley, L.; Mainwaring, G.I.; Samuels, S.; Perry, S.; Mann, C.H. Exogenous carbohydrate oxidation during ultraendurance exercise. J. Appl. Physiol. 2006, 100, 1134–1141. [Google Scholar] [CrossRef] [PubMed]
- Hulston, C.J.; Wallis, G.A.; Jeukendrup, A.E. Exogenous CHO oxidation with glucose plus fructose intake during exercise. Med. Sci. Sports Exerc. 2009, 41, 357–363. [Google Scholar] [CrossRef]
- Viribay, A.; Arribalzaga, S.; Mielgo-Ayuso, J.; Castaneda-Babarro, A.; Seco-Calvo, J.; Urdampilleta, A. Effects of 120 g/h of carbohydrates intake during a mountain marathon on exercise-induced muscle damage in elite runners. Nutrients 2020, 12. [Google Scholar] [CrossRef]
- Sutehall, S.; Muniz-Pardos, B.; Bosch, A.N.; Di Gianfrancesco, A.; Pitsiladis, Y.P. Sports drinks on the edge of a new era. Curr. Sports Med. Rep. 2018, 17, 112–116. [Google Scholar] [CrossRef]
- Pettersson, S.; Edin, F.; Bakkman, L.; McGawley, K. Effects of supplementing with an 18% carbohydrate-hydrogel drink versus a placebo during whole-body exercise in −5 °C with elite cross-country ski athletes: A crossover study. J. Int. Soc. Sport Nutr. 2019, 16, 46. [Google Scholar] [CrossRef]
- Nielsen, L.L.K.; Lambert, M.N.T.; Jeppesen, P.B. The effect of ingesting carbohydrate and proteins on athletic performance: A systematic review and meta-analysis of randomized controlled trials. Nutrients 2020, 12. [Google Scholar] [CrossRef]
- Betts, J.A.; Stevenson, E. Should protein be included in CHO-based sports supplements? Med. Sci. Sports Exerc. 2011, 43, 1244–1250. [Google Scholar] [CrossRef]
- Davis, J.M.; Bailey, S.P. Possible mechanisms of central nervous system fatigue during exercise. Med. Sci. Sports Exerc. 1997, 29, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Stearns, R.L.; Emmanuel, H.; Volek, J.S.; Casa, D.J. Effects of ingesting protein in combination with carbohydrate during exercise on endurance performance: A systematic review with meta-analysis. J. Strength Cond. Res. 2010, 24, 2192–2202. [Google Scholar] [CrossRef]
- Dardarian, N.; O’Donnell, K. Applying the science of nutrient timing and distribution on-and-off the soccer pitch: Sports nutritionist’s experience. Curr. Sports Med. Rep. 2018, 17, 242–243. [Google Scholar] [CrossRef]
- Saltin, B. Metabolic fundamentals in exercise. Med. Sci Sports Exerc 1973, 5, 137–146. [Google Scholar] [CrossRef]
- Baker, L.B.; Nuccio, R.P.; Jeukendrup, A.E. Acute effects of dietary constituents on motor skill and cognitive performance in athletes. Nutr. Rev. 2014, 72, 790–802. [Google Scholar] [CrossRef] [PubMed]
- Russell, M.; Benton, D.; Kingsley, M. Influence of carbohydrate supplementation on skill performance during a soccer match simulation. J. Sci. Med. Sport 2012, 15, 348–354. [Google Scholar] [CrossRef] [PubMed]
- Kingsley, M.; Penas-Ruiz, C.; Terry, C.; Russell, M. Effects of carbohydrate-hydration strategies on glucose metabolism, sprint performance and hydration during a soccer match simulation in recreational players. J. Sci. Med. Sport 2014, 17, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Grego, F.; Vallier, J.M.; Collardeau, M.; Bermon, S.; Ferrari, P.; Candito, M.; Bayer, P.; Magnie, M.N.; Brisswalter, J. Effects of long duration exercise on cognitive function, blood glucose, and counterregulatory hormones in male cyclists. Neurosci. Lett. 2004, 364, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, M. Immune function in sport and exercise. J. Appl. Physiol. 2007, 103, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Stellingwerff, T.; Spriet, L.L.; Watt, M.J.; Kimber, N.E.; Hargreaves, M.; Hawley, J.A.; Burke, L.M. Decreased PDH activation and glycogenolysis during exercise following fat adaptation with carbohydrate restoration. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E380–E388. [Google Scholar] [CrossRef]
- Haff, G.G.; Koch, A.J.; Potteiger, J.A.; Kuphal, K.E.; Magee, L.M.; Green, S.B.; Jakicic, J.J. Carbohydrate supplementation attenuates muscle glycogen loss during acute bouts of resistance exercise. Int. J. Sport Nutr. Exerc. Metab. 2000, 10, 326–339. [Google Scholar] [CrossRef]
- Haff, G.G.; Schroeder, C.A.; Koch, A.J.; Kuphal, K.E.; Comeau, M.J.; Potteiger, J.A. The effects of supplemental carbohydrate ingestion on intermittent isokinetic leg exercise. J. Sports Med. Phys. Fit. 2001, 41, 216–222. [Google Scholar]
- van Loon, L.J. Is there a need for protein ingestion during exercise? Sports Med. 2014, 44 (Suppl. 1), S105–S111. [Google Scholar] [CrossRef] [PubMed]
- Bird, S.P.; Tarpenning, K.M.; Marino, F.E. Liquid carbohydrate/essential amino acid ingestion during a short-term bout of resistance exercise suppresses myofibrillar protein degradation. Metabolism 2006, 55, 570–577. [Google Scholar] [CrossRef] [PubMed]
- Bird, S.P.; Tarpenning, K.M.; Marino, F.E. Effects of liquid carbohydrate/essential amino acid ingestion on acute hormonal response during a single bout of resistance exercise in untrained men. Nutrition 2006, 22, 367–375. [Google Scholar] [CrossRef]
- McConell, G.; Kloot, K.; Hargreaves, M. Effect of timing of carbohydrate ingestion on endurance exercise performance. Med. Sci. Sports Exerc. 1996, 28, 1300–1304. [Google Scholar] [CrossRef]
- Schweitzer, G.G.; Smith, J.D.; Lecheminant, J.D. Timing carbohydrate beverage intake during prolonged moderate intensity exercise does not affect cycling performance. Int. J. Exerc. Sci. 2009, 2, 4–18. [Google Scholar]
- Beelen, M.; Burke, L.M.; Gibala, M.J.; van Loon, L.J. Nutritional strategies to promote postexercise recovery. Int. J. Sport Nutr. Exerc. Metab. 2010, 20, 515–532. [Google Scholar] [CrossRef]
- Jentjens, R.; Jeukendrup, A. Determinants of post-exercise glycogen synthesis during short-term recovery. Sports Med. 2003, 33, 117–144. [Google Scholar] [CrossRef]
- Hayashi, T.; Wojtaszewski, J.F.; Goodyear, L.J. Exercise regulation of glucose transport in skeletal muscle. Am. J. Physiol. 1997, 273, E1039–E1051. [Google Scholar] [CrossRef]
- Goodyear, L.J.; Hirshman, M.F.; King, P.A.; Horton, E.D.; Thompson, C.M.; Horton, E.S. Skeletal muscle plasma membrane glucose transport and glucose transporters after exercise. J. Appl. Physiol. 1990, 68, 193–198. [Google Scholar] [CrossRef]
- Burke, L.M. Fueling strategies to optimize performance: Training high or training low? Scand. J. Med. Sci. Sports 2010, 20 (Suppl. 2), 48–58. [Google Scholar] [CrossRef]
- Alghannam, A.F.; Gonzalez, J.T.; Betts, J.A. Restoration of muscle glycogen and functional capacity: Role of post-exercise carbohydrate and protein co-ingestion. Nutrients 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- van Loon, L.J.; Saris, W.H.; Kruijshoop, M.; Wagenmakers, A.J. Maximizing postexercise muscle glycogen synthesis: Carbohydrate supplementation and the application of amino acid or protein hydrolysate mixtures. Am. J. Clin. Nutr. 2000, 72, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Burke, L.M.; Collier, G.R.; Hargreaves, M. Muscle glycogen storage after prolonged exercise: Effect of the glycemic index of carbohydrate feedings. J. Appl. Physiol. 1993, 75, 1019–1023. [Google Scholar] [CrossRef] [PubMed]
- Erith, S.; Williams, C.; Stevenson, E.; Chamberlain, S.; Crews, P.; Rushbury, I. The effect of high carbohydrate meals with different glycemic indices on recovery of performance during prolonged intermittent high-intensity shuttle running. Int. J. Sport Nutr. Exerc. Metab. 2006, 16, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, E.; Williams, C.; McComb, G.; Oram, C. Improved recovery from prolonged exercise following the consumption of low glycemic index carbohydrate meals. Int. J. Sport Nutr. Exerc. Metab. 2005, 15, 333–349. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.J.; Midgley, A.W.; Vince, R.V.; Madden, L.A.; McNaughton, L.R. High versus low glycemic index 3-h recovery diets following glycogen-depleting exercise has no effect on subsequent 5-km cycling time trial performance. J. Sci. Med. Sport 2013, 16, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, N.R.; DiMarco, N.M.; Langley, S. Position of the American dietetic association, dietitians of Canada, and the American college of sports medicine: Nutrition and athletic performance. J. Am. Diet. Assoc. 2009, 109, 509–527. [Google Scholar] [CrossRef]
- Nieman, D.C.; Mitmesser, S.H. Potential impact of nutrition on immune system recovery from heavy exertion: A metabolomics perspective. Nutrients 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Scharhag, J.; Meyer, T.; Auracher, M.; Gabriel, H.H.; Kindermann, W. Effects of graded carbohydrate supplementation on the immune response in cycling. Med. Sci. Sports Exerc. 2006, 38, 286–292. [Google Scholar] [CrossRef] [PubMed]
- van Loon, L.J.; Kruijshoop, M.; Verhagen, H.; Saris, W.H.; Wagenmakers, A.J. Ingestion of protein hydrolysate and amino acid-carbohydrate mixtures increases postexercise plasma insulin responses in men. J. Nutr. 2000, 130, 2508–2513. [Google Scholar] [CrossRef] [PubMed]
- Kaastra, B.; Manders, R.J.; Van Breda, E.; Kies, A.; Jeukendrup, A.E.; Keizer, H.A.; Kuipers, H.; Van Loon, L.J. Effects of increasing insulin secretion on acute postexercise blood glucose disposal. Med. Sci. Sports Exerc. 2006, 38, 268–275. [Google Scholar] [CrossRef]
- Rustad, P.I.; Sailer, M.; Cumming, K.T.; Jeppesen, P.B.; Kolnes, K.J.; Sollie, O.; Franch, J.; Ivy, J.L.; Daniel, H.; Jensen, J. Intake of protein plus carbohydrate during the first two hours after exhaustive cycling improves performance the following day. PLoS ONE 2016, 11, e0153229. [Google Scholar] [CrossRef]
- de Oliveira, E.P.; Burini, R.C.; Jeukendrup, A. Gastrointestinal complaints during exercise: Prevalence, etiology, and nutritional recommendations. Sports Med. 2014, 44 (Suppl. 1), S79–S85. [Google Scholar] [CrossRef]
- Pedersen, D.J.; Lessard, S.J.; Coffey, V.G.; Churchley, E.G.; Wootton, A.M.; Ng, T.; Watt, M.J.; Hawley, J.A. High rates of muscle glycogen resynthesis after exhaustive exercise when carbohydrate is coingested with caffeine. J. Appl. Physiol. 2008, 105, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.; Higham, D.; Close, G.L.; Morton, J.P. The effect of adding caffeine to postexercise carbohydrate feeding on subsequent high-intensity interval-running capacity compared with carbohydrate alone. Int. J. Sport Nutr. Exerc. Metab. 2011, 21, 410–416. [Google Scholar] [CrossRef]
- Hawley, J.A.; Tipton, K.D.; Millard-Stafford, M.L. Promoting training adaptations through nutritional interventions. J. Sports Sci. 2006, 24, 709–721. [Google Scholar] [CrossRef]
- Ivy, J.L. Regulation of muscle glycogen repletion, muscle protein synthesis and repair following exercise. J. Sports Sci. Med. 2004, 3, 131–138. [Google Scholar]
- Moore, D.R. Maximizing post-exercise anabolism: The case for relative protein intakes. Front. Nutr. 2019, 6, 147. [Google Scholar] [CrossRef]
- Witard, O.C.; Jackman, S.R.; Breen, L.; Smith, K.; Selby, A.; Tipton, K.D. Myofibrillar muscle protein synthesis rates subsequent to a meal in response to increasing doses of whey protein at rest and after resistance exercise. Am. J. Clin. Nutr. 2014, 99, 86–95. [Google Scholar] [CrossRef]
- Tang, J.E.; Moore, D.R.; Kujbida, G.W.; Tarnopolsky, M.A.; Phillips, S.M. Ingestion of whey hydrolysate, casein, or soy protein isolate: Effects on mixed muscle protein synthesis at rest and following resistance exercise in young men. J. Appl. Physiol. 2009, 107, 987–992. [Google Scholar] [CrossRef]
- Rowlands, D.S.; Nelson, A.R.; Phillips, S.M.; Faulkner, J.A.; Clarke, J.; Burd, N.A.; Moore, D.; Stellingwerff, T. Protein-leucine fed dose effects on muscle protein synthesis after endurance exercise. Med. Sci. Sports Exerc. 2015, 47, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Macnaughton, L.S.; Wardle, S.L.; Witard, O.C.; McGlory, C.; Hamilton, D.L.; Jeromson, S.; Lawrence, C.E.; Wallis, G.A.; Tipton, K.D. The response of muscle protein synthesis following whole-body resistance exercise is greater following 40 g than 20 g of ingested whey protein. Physiol. Rep. 2016, 4, e12893. [Google Scholar] [CrossRef] [PubMed]
- Borsheim, E.; Cree, M.G.; Tipton, K.D.; Elliott, T.A.; Aarsland, A.; Wolfe, R.R. Effect of carbohydrate intake on net muscle protein synthesis during recovery from resistance exercise. J. Appl. Physiol. 2004, 96, 674–678. [Google Scholar] [CrossRef] [PubMed]
- Roy, B.D.; Tarnopolsky, M.A.; MacDougall, J.D.; Fowles, J.; Yarasheski, K.E. Effect of glucose supplement timing on protein metabolism after resistance training. J. Appl. Physiol. 1997, 82, 1882–1888. [Google Scholar] [CrossRef] [PubMed]
- Nelson, A.R.; Phillips, S.M.; Stellingwerff, T.; Rezzi, S.; Bruce, S.J.; Breton, I.; Thorimbert, A.; Guy, P.A.; Clarke, J.; Broadbent, S.; et al. A protein-leucine supplement increases branched-chain amino acid and nitrogen turnover but not performance. Med. Sci. Sports Exerc. 2012, 44, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Di Donato, D.M.; West, D.W.; Churchward-Venne, T.A.; Breen, L.; Baker, S.K.; Phillips, S.M. Influence of aerobic exercise intensity on myofibrillar and mitochondrial protein synthesis in young men during early and late postexercise recovery. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E1025–E1032. [Google Scholar] [CrossRef]
- Phillips, S.M.; Tipton, K.D.; Aarsland, A.; Wolf, S.E.; Wolfe, R.R. Mixed muscle protein synthesis and breakdown after resistance exercise in humans. Am. J. Physiol. 1997, 273, 99–107. [Google Scholar] [CrossRef]
- Gabel, K.; Hoddy, K.K.; Haggerty, N.; Song, J.; Kroeger, C.M.; Trepanowski, J.F.; Panda, S.; Varady, K.A. Effects of 8-hour time restricted feeding on body weight and metabolic disease risk factors in obese adults: A pilot study. Nutr. Healthy Aging 2018, 4, 345–353. [Google Scholar] [CrossRef]
- Tinsley, G.M.; Moore, M.L.; Graybeal, A.J.; Paoli, A.; Kim, Y.; Gonzales, J.U.; Harry, J.R.; VanDusseldorp, T.A.; Kennedy, D.N.; Cruz, M.R. Time-restricted feeding plus resistance training in active females: A randomized trial. Am. J. Clin. Nutr. 2019, 110, 628–640. [Google Scholar] [CrossRef] [PubMed]
- Aird, T.P.; Davies, R.W.; Carson, B.P. Effects of fasted vs fed-state exercise on performance and post-exercise metabolism: A systematic review and meta-analysis. Scand. J. Med. Sci. Sports 2018, 28, 1476–1493. [Google Scholar] [CrossRef] [PubMed]
- Clayton, D.J.; James, L.J. The effect of breakfast on appetite regulation, energy balance and exercise performance. Proc. Nutr. Soc. 2016, 75, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Clayton, D.J.; Barutcu, A.; Machin, C.; Stensel, D.J.; James, L.J. Effect of breakfast omission on energy intake and evening exercise performance. Med. Sci. Sports Exerc. 2015, 47, 2645–2652. [Google Scholar] [CrossRef] [PubMed]
- Arciero, P.J.; Baur, D.; Connelly, S.; Ormsbee, M.J. Timed-daily ingestion of whey protein and exercise training reduces visceral adipose tissue mass and improves insulin resistance: The PRISE study. J. Appl. Physiol 2014, 117, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Arciero, P.J.; Edmonds, R.; He, F.; Ward, E.; Gumpricht, E.; Mohr, A.; Ormsbee, M.J.; Astrup, A. Protein-pacing caloric-restriction enhances body composition similarly in obese men and women during weight loss and sustains efficacy during long-term weight maintenance. Nutrients 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Arciero, P.J.; Edmonds, R.C.; Bunsawat, K.; Gentile, C.L.; Ketcham, C.; Darin, C.; Renna, M.; Zheng, Q.; Zhang, J.Z.; Ormsbee, M.J. Protein-pacing from food or supplementation improves physical performance in overweight men and women: The PRISE 2 study. Nutrients 2016, 8. [Google Scholar] [CrossRef]
- Arciero, P.J.; Ives, S.J.; Norton, C.; Escudero, D.; Minicucci, O.; O’Brien, G.; Paul, M.; Ormsbee, M.J.; Miller, V.; Sheridan, C.; et al. Protein-pacing and multi-component exercise training improves physical performance outcomes in exercise-trained women: The PRISE 3 study. Nutrients 2016, 8, 332. [Google Scholar] [CrossRef] [PubMed]
- Mamerow, M.M.; Mettler, J.A.; English, K.L.; Casperson, S.L.; Arentson-Lantz, E.; Sheffield-Moore, M.; Layman, D.K.; Paddon-Jones, D. Dietary protein distribution positively influences 24-h muscle protein synthesis in healthy adults. J. Nutr. 2014, 144, 876–880. [Google Scholar] [CrossRef]
- Arciero, P.J.; Ormsbee, M.J.; Gentile, C.L.; Nindl, B.C.; Brestoff, J.R.; Ruby, M. Increased protein intake and meal frequency reduces abdominal fat during energy balance and energy deficit. Obesity 2013, 21, 1357–1366. [Google Scholar] [CrossRef]
- Trommelen, J.; van Loon, L.J. Pre-sleep protein ingestion to improve the skeletal muscle adaptive response to exercise training. Nutrients 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Snijders, T.; Res, P.T.; Smeets, J.S.; van Vliet, S.; van Kranenburg, J.; Maase, K.; Kies, A.K.; Verdijk, L.B.; van Loon, L.J. Protein ingestion before sleep increases muscle mass and strength gains during prolonged resistance-type exercise training in healthy young men. J. Nutr. 2015, 145, 1178–1184. [Google Scholar] [CrossRef] [PubMed]
- Ormsbee, M.J.; Gorman, K.A.; Miller, E.A.; Baur, D.A.; Eckel, L.A.; Contreras, R.J.; Panton, L.B.; Spicer, M.T. Nighttime feeding likely alters morning metabolism but not exercise performance in female athletes. Appl. Physiol. Nutr. Me. 2016, 41, 719–727. [Google Scholar] [CrossRef] [PubMed]
- Antonio, J.; Ellerbroek, A.; Peacock, C.; Silver, T. Casein protein supplementation in trained men and women: Morning versus evening. Int. J. Exerc. Sci. 2017, 10, 479–486. [Google Scholar]
- Shukla, A.P.; Iliescu, R.G.; Thomas, C.E.; Aronne, L.J. Food order has a significant impact on postprandial glucose and insulin levels. Diabetes Care 2015, 38, e98–e99. [Google Scholar] [CrossRef] [PubMed]
- Knuiman, P.; Hopman, M.T.; Mensink, M. Glycogen availability and skeletal muscle adaptations with endurance and resistance exercise. Nutr. Metab. 2015, 12, 59. [Google Scholar] [CrossRef]
- Impey, S.G.; Hearris, M.A.; Hammond, K.M.; Bartlett, J.D.; Louis, J.; Close, G.L.; Morton, J.P. Fuel for the work required: A theoretical framework for carbohydrate periodization and the glycogen threshold hypothesis. Sports Med. 2018, 48, 1031–1048. [Google Scholar] [CrossRef]
- Marquet, L.A.; Brisswalter, J.; Louis, J.; Tiollier, E.; Burke, L.M.; Hawley, J.A.; Hausswirth, C. Enhanced endurance performance by periodization of carbohydrate intake: “Sleep Low” strategy. Med. Sci. Sports Exerc. 2016, 48, 663–672. [Google Scholar] [CrossRef]
- Marquet, L.A.; Hausswirth, C.; Molle, O.; Hawley, J.A.; Burke, L.M.; Tiollier, E.; Brisswalter, J. Periodization of carbohydrate intake: Short-term effect on performance. Nutrients 2016, 8. [Google Scholar] [CrossRef]
- Mitchell, C.J.; Gatta, P.A.D.; Petersen, A.C.; Cameron-Smith, D.; Markworth, J.F. Soy protein ingestion results in less prolonged p70S6 kinase phosphorylation compared to whey protein after resistance exercise in older men. J. Int. Soc. Sport Nutr. 2015, 12, 6. [Google Scholar] [CrossRef]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arent, S.M.; Cintineo, H.P.; McFadden, B.A.; Chandler, A.J.; Arent, M.A. Nutrient Timing: A Garage Door of Opportunity? Nutrients 2020, 12, 1948. https://doi.org/10.3390/nu12071948
Arent SM, Cintineo HP, McFadden BA, Chandler AJ, Arent MA. Nutrient Timing: A Garage Door of Opportunity? Nutrients. 2020; 12(7):1948. https://doi.org/10.3390/nu12071948
Chicago/Turabian StyleArent, Shawn M., Harry P. Cintineo, Bridget A. McFadden, Alexa J. Chandler, and Michelle A. Arent. 2020. "Nutrient Timing: A Garage Door of Opportunity?" Nutrients 12, no. 7: 1948. https://doi.org/10.3390/nu12071948
APA StyleArent, S. M., Cintineo, H. P., McFadden, B. A., Chandler, A. J., & Arent, M. A. (2020). Nutrient Timing: A Garage Door of Opportunity? Nutrients, 12(7), 1948. https://doi.org/10.3390/nu12071948