In Vitro Evaluation of Prebiotic Properties of a Commercial Artichoke Inflorescence Extract Revealed Bifidogenic Effects


 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Short-Term Colonic Fermentation
2.3. Microbial Metabolic Activity Analysis
2.4. Microbial Community Analysis
2.5. Statistics
3. Results
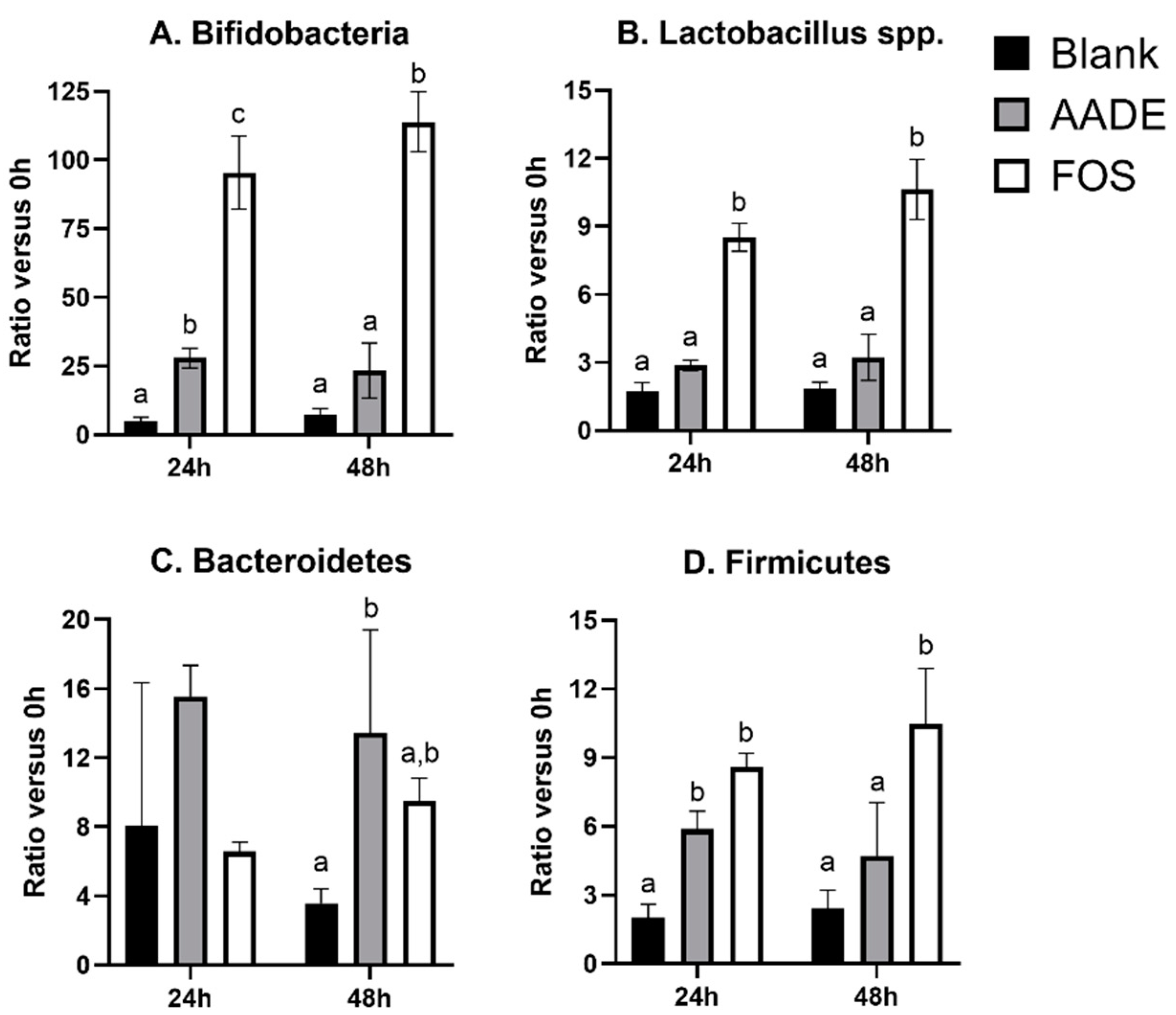
3.1. Microbial Composition
3.2. pH
3.3. Gas Pressure
3.4. Lactate and Carbohydrate (SCFAs, Acetate, Butyrate, and Propionate) and Protein Metabolites (BCFAs and Ammonium)
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Frank, D.N.; Amand, A.L.S.; Feldman, R.A.; Boedeker, E.C.; Harpaz, N.; Pace, N.R. Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc. Natl. Acad. Sci. 2007, 104, 13780–13785. [Google Scholar] [CrossRef] [PubMed]
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; Vuyyuru, H.; Sasikala, M.; Reddy, D.N. Role of the normal gut microbiota. World J. Gastroenterol. 2015, 21, 8787. [Google Scholar] [CrossRef] [PubMed]
- Jandhyala, S.M.; Madhulika, A.; Deepika, G.; Rao, G.V.; Reddy, D.N.; Subramanyam, C.; Sasikala, M.; Talukdar, R. Altered intestinal microbiota in patients with chronic pancreatitis: Implications in diabetes and metabolic abnormalities. Sci. Rep. 2017, 7, 43640. [Google Scholar] [CrossRef]
- Blaser, M.J. The microbiome revolution. J. Clin. Investig. 2014, 124, 4162–4165. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef] [PubMed]
- A. Parnell, J.; A. Reimer, R. Prebiotic fiber modulation of the gut microbiota improves risk factors for obesity and the metabolic syndrome. Gut Microbes 2012, 3, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Oku, T.; Tokunaga, T.; Hosoya, N. Nondigestibility of a new sweetener, “Neosugar”, in the rat. J. Nutr. 1984, 114, 1574–1581. [Google Scholar] [CrossRef]
- Spiegel, J.E.; Rose, R.; Karabell, P.; Frankos, V.H.; Schmitt, D.F. Safety and benefits of fructooligosaccharides as food ingredients. Food Technol. (Chic.) 1994, 48, 85–89. [Google Scholar]
- Johnson, J. Are Fructooligosaccharides Safe? In Medical News Today. Available online: https://www.medicalnewstoday.com/articles/319299.php (accessed on 15 October 2018).
- Williams, C.; Witherly, S.; Buddington, R. Influence of dietary neosugar on selected bacterial groups of the human faecal microbiota. Microb. Ecol. Health Dis. 1994, 7, 91–97. [Google Scholar] [CrossRef]
- Gibson, G.R.; Beatty, E.R.; Wang, X.; Cummings, J.H. Selective stimulation of bifidobacteria in the human colon by oligofructose and inulin. Gastroenterology 1995, 108, 975–982. [Google Scholar] [CrossRef]
- Buddington, R.K.; Williams, C.H.; Chen, S.-C.; Witherly, S.A. Dietary supplement of neosugar alters the fecal flora and decreases activities of some reductive enzymes in human subjects. Am. J. Clin. Nutr. 1996, 63, 709–716. [Google Scholar] [CrossRef] [PubMed]
- Kleessen, B.; Sykura, B.; Zunft, H.-J.; Blaut, M. Effects of inulin and lactose on fecal microflora, microbial activity, and bowel habit in elderly constipated persons. Am. J. Clin. Nutr. 1997, 65, 1397–1402. [Google Scholar] [CrossRef] [PubMed]
- Bouhnik, Y.; Vahedi, K.; Achour, L.; Attar, A.; Salfati, J.; Pochart, P.; Marteau, P.; Flourie, B.; Bornet, F.; Rambaud, J.-C. Short-chain fructo-oligosaccharide administration dose-dependently increases fecal bifidobacteria in healthy humans. J. Nutr. 1999, 129, 113–116. [Google Scholar] [CrossRef]
- Kruse, H.-P.; Kleessen, B.; Blaut, M. Effects of inulin on faecal bifidobacteria in human subjects. Br. J. Nutr. 1999, 82, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Den Hond, E.; Geypens, B.; Ghoos, Y. Effect of high performance chicory inulin on constipation. Nutr. Res. 2000, 20, 731–736. [Google Scholar] [CrossRef]
- Van De Wiele, T.; Boon, N.; Possemiers, S.; Jacobs, H.; Verstraete, W. Inulin-type fructans of longer degree of polymerization exert more pronounced in vitro prebiotic effects. J. Appl. Microbiol. 2007, 102, 452–460. [Google Scholar] [CrossRef]
- Van den Abbeele, P.; Taminiau, B.; Pinheiro, I.; Duysburgh, C.; Jacobs, H.; Pijls, L.; Marzorati, M. Arabinoxylo-Oligosaccharides and Inulin Impact Inter-Individual Variation on Microbial Metabolism and Composition, Which Immunomodulates Human Cells. J. Agric. Food Chem. 2018, 66, 1121–1130. [Google Scholar] [CrossRef]
- Hardy, H.; Harris, J.; Lyon, E.; Beal, J.; Foey, A.D. Probiotics, prebiotics and immunomodulation of gut mucosal defences: Homeostasis and immunopathology. Nutrients 2013, 5, 1869–1912. [Google Scholar] [CrossRef]
- Kleessen, B.; Schwarz, S.; Boehm, A.; Fuhrmann, H.; Richter, A.; Henle, T.; Krueger, M. Jerusalem artichoke and chicory inulin in bakery products affect faecal microbiota of healthy volunteers. Br. J. Nutr. 2007, 98, 540–549. [Google Scholar] [CrossRef]
- Costabile, A.; Kolida, S.; Klinder, A.; Gietl, E.; Bäuerlein, M.; Frohberg, C.; Landschütze, V.; Gibson, G.R. A double-blind, placebo-controlled, cross-over study to establish the bifidogenic effect of a very-long-chain inulin extracted from globe artichoke (Cynara scolymus) in healthy human subjects. Br. J. Nutr. 2010, 104, 1007–1017. [Google Scholar] [CrossRef]
- D’Antuono, I.; Garbetta, A.; Linsalata, V.; Minervini, F.; Cardinali, A. Polyphenols from artichoke heads (Cynara cardunculus (L.) subsp. scolymus Hayek): In vitro bio-accessibility, intestinal uptake and bioavailability. Food Funct. 2015, 6, 1268–1277. [Google Scholar] [PubMed]
- Rocchetti, G.; Giuberti, G.; Lucchini, F.; Lucini, L. Polyphenols and sesquiterpene lactones from artichoke heads: Modulation of starch digestion, gut bioaccessibility, and bioavailability following in vitro digestion and large intestine fermentation. Antioxidants 2020, 9, 306. [Google Scholar] [CrossRef] [PubMed]
- Council of Europe. European Pharmacopoeia; Council of Europe: Strasbourg, France, Ph. Eur. 10.1 04/2019:0765.
- De Weirdt, R.; Possemiers, S.; Vermeulen, G.; Moerdijk-Poortvliet, T.C.; Boschker, H.T.; Verstraete, W.; Van de Wiele, T. Human faecal microbiota display variable patterns of glycerol metabolism. FEMS Microbiol. Ecol. 2010, 74, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Vilchez-Vargas, R.; Geffers, R.; Suarez-Diez, M.; Conte, I.; Waliczek, A.; Kaser, V.S.; Kralova, M.; Junca, H.; Pieper, D.H. Analysis of the microbial gene landscape and transcriptome for aromatic pollutants and alkane degradation using a novel internally calibrated microarray system. Environ. Microbiol. 2013, 15, 1016–1039. [Google Scholar] [CrossRef] [PubMed]
- Freire, F.C.; Adorno, M.A.T.; Sakamoto, I.K.; Antoniassi, R.; Chaves, A.C.S.D.; Dos Santos, K.M.O.; Sivieri, K. Impact of multi-functional fermented goat milk beverage on gut microbiota in a dynamic colon model. Food Res. Int. 2017, 99, 315–327. [Google Scholar] [CrossRef]
- Barszcz, M.; Taciak, M.; Skomiał, J. The effects of inulin, dried Jerusalem artichoke tuber and a multispecies probiotic preparation on microbiota ecology and immune status of the large intestine in young pigs. Arch. Anim. Nutr. 2016, 70, 278–292. [Google Scholar] [CrossRef]
- Van Hoek, M.J.; Merks, R.M. Emergence of microbial diversity due to cross-feeding interactions in a spatial model of gut microbial metabolism. BMC Syst. Biol. 2017, 11, 56. [Google Scholar]
- Cummings, J.H.; Macfarlane, G.T.; Englyst, H.N. Prebiotic digestion and fermentation. Am. J. Clin. Nutr. 2001, 73, 415s–420s. [Google Scholar] [CrossRef]
- Blaut, M. Relationship of prebiotics and food to intestinal microflora. Eur. J. Nutr. 2002, 41, i11–i16. [Google Scholar] [CrossRef]
- Rios-Covian, D.; Salazar, N.; Gueimonde, M.; de Los Reyes-Gavilan, C.G. Shaping the Metabolism of Intestinal Bacteroides Population through Diet to Improve Human Health. Front. Microbiol. 2017, 8, 376. [Google Scholar] [CrossRef]
- Aguirre, M.; Eck, A.; Koenen, M.E.; Savelkoul, P.H.; Budding, A.E.; Venema, K. Diet drives quick changes in the metabolic activity and composition of human gut microbiota in a validated in vitro gut model. Res. Microbiol. 2016, 167, 114–125. [Google Scholar] [CrossRef]
- Salonen, A.; Lahti, L.; Salojärvi, J.; Holtrop, G.; Korpela, K.; Duncan, S.H.; Date, P.; Farquharson, F.; Johnstone, A.M.; Lobley, G.E.; et al. Impact of diet and individual variation on intestinal microbiota composition and fermentation products in obese men. ISME J. 2014, 8, 2218–2230. [Google Scholar] [CrossRef] [PubMed]
- Chambers, E.S.; Viardot, A.; Psichas, A.; Morrison, D.J.; Murphy, K.G.; Zac-Varghese, S.E.; MacDougall, K.; Preston, T.; Tedford, C.; Finlayson, G.S. Effects of targeted delivery of propionate to the human colon on appetite regulation, body weight maintenance and adiposity in overweight adults. Gut 2015, 64, 1744–1754. [Google Scholar] [CrossRef] [PubMed]
- Herreweghen, F.; Van den Abbeele, P.; De Mulder, T.; De Weirdt, R.; Geirnaert, A.; Hernandez-Sanabria, E.; Vilchez-Vargas, R.; Jauregui, R.; Pieper, D.H.; Belzer, C.; et al. In vitro colonisation of the distal colon by Akkermansia muciniphila is largely mucin and pH dependent. Benef. Microbes 2017, 8, 81–96. [Google Scholar] [CrossRef] [PubMed]
- Van den Abbeele, P.; Gérard, P.; Rabot, S.; Bruneau, A.; El Aidy, S.; Derrien, M.; Kleerebezem, M.; Zoetendal, E.G.; Smidt, H.; Verstraete, W.; et al. Arabinoxylans and inulin differentially modulate the mucosal and luminal gut microbiota and mucin-degradation in humanized rats. Environ. Microbiol. 2011, 13, 2667–2680. [Google Scholar] [CrossRef] [PubMed]
- Baxter, N.T.; Schmidt, A.W.; Venkataraman, A.; Kim, K.S.; Waldron, C.; Schmidt, T.M. Dynamics of Human Gut Microbiota and Short-Chain Fatty Acids in Response to Dietary Interventions with Three Fermentable Fibers. mBio 2019, 10, e02566-18. [Google Scholar] [CrossRef]
- Maathuis, A.J.; van den Heuvel, E.G.; Schoterman, M.H.; Venema, K. Galacto-oligosaccharides have prebiotic activity in a dynamic in vitro colon model using a 13C-labeling technique. J. Nutr. 2012, 142, 1205–1212. [Google Scholar] [CrossRef]
- Tolhurst, G.; Heffron, H.; Lam, Y.S.; Parker, H.E.; Habib, A.M.; Diakogiannaki, E.; Cameron, J.; Grosse, J.; Reimann, F.; Gribble, F.M. Short-chain fatty acids stimulate glucagon-like peptide-1 secretion via the G-protein–coupled receptor FFAR2. Diabetes 2012, 61, 364–371. [Google Scholar] [CrossRef]
- Freeland, K.R.; Wolever, T.M. Acute effects of intravenous and rectal acetate on glucagon-like peptide-1, peptide YY, ghrelin, adiponectin and tumour necrosis factor-α. Br. J. Nutr. 2010, 103, 460–466. [Google Scholar] [CrossRef]
- Geboes, K.P.; De Hertogh, G.; De Preter, V.; Luypaerts, A.; Bammens, B.; Evenepoel, P.; Ghoos, Y.; Geboes, K.; Rutgeerts, P.; Verbeke, K. The influence of inulin on the absorption of nitrogen and the production of metabolites of protein fermentation in the colon. Br. J. Nutr. 2006, 96, 1078–1086. [Google Scholar] [CrossRef]
- Van den Abbeele, P.; Roos, S.; Eeckhaut, V.; MacKenzie, D.A.; Derde, M.; Verstraete, W.; Marzorati, M.; Possemiers, S.; Vanhoecke, B.; Van Immerseel, F. Incorporating a mucosal environment in a dynamic gut model results in a more representative colonization by lactobacilli. Microb. Biotechnol. 2012, 5, 106–115. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Target Species | Primer Sequences 5′-3′ and 3′-5′ |
|---|---|
| Bacteroidetes [27] | GGAACATGTGGTTTAATTCGATGAT |
| AGCTGACGACAACCATGCAG | |
| Firmicutes [27] | GGAGCATGTGGTTTAATTCGAAGCA |
| AGCTGACGACAACCATGCAC | |
| Lactobacillus spp. [20] | AGCAGTAGGGAATCTTCCA |
| CGCCACTGGTGTTCYTCCATATA | |
| Bifidobacterium spp. [28] | TCGCGTCYGGTGTGAAAG |
| CCACATCCAGCYTCCAC | |
| Akkermansia muciniphila [29] | CAGCACGTGAAGGTGGGGAC |
| CCTTGCGGTTGGCTTCAGAT |
| Levels of Microbial Groups (log (16S rRNA Copies/mL)) | |||||||
|---|---|---|---|---|---|---|---|
| Incubation Time (h) | 0 h | 24 h | 48 h | ||||
| Blank | AADE | FOS | Blank | AADE | FOS | ||
| Firmicutes | 9.96 ± 0.36 | 10.36 ± 0.13 a | 10.83 ± 0.06 b | 11.00 ± 0.03 b | 10.44 ± 0.14 a | 10.70 ± 0.25 a | 11.08 ± 0.10 b |
| Bacteroidetes | 9.73 ± 0.46 | 10.64 ± 0.47 | 11.09 ± 0.05 | 10.72 ± 0.04 | 10.44 ± 0.10 a | 10.99 ± 0.23 b | 10.87 ± 0.06 a,b |
| Bifidobacteria | 8.26 ± 0.25 | 8.94 ± 0.13 a | 9.71 ± 0.06 b | 10.24 ± 0.06 c | 9.12 ± 0.14 a | 9.60 ± 0.22 b | 10.32 ± 0.04 c |
| Lactobacillus spp. | 6.58 ± 0.19 | 6.84 ± 0.10 a | 7.06 ± 0.03 a | 7.54 ± 0.03 b | 6.87 ± 0.06 a | 7.01 ± 0.15 b | 7.63 ± 0.06 c |
| Akkermansia mucinphila | 7.02 ± 0.51 | 8.23 ± 0.39 | 8.30 ± 0.03 | 7.72 ± 0.08 | 8.08 ± 0.17 a,b | 8.19 ± 0.25 b | 7.85 ± 0.08 a |
| Incubation Time (h) | pH | ||
| Blank | AADE | FOS | |
| 0 | 6.49 ± 0.02 | 6.51 ± 0.00 | 6.51 ± 0.00 |
| 6 | 6.39 ± 0.01 a | 6.22 ± 0.01 b | 5.64 ± 0.13 c |
| 24 | 6.46 ± 0.02 a | 6.21 ± 0.01 b | 5.66 ± 0.02 c |
| 48 | 6.40 ± 0.04 a | 6.20 ± 0.02 b | 5.69 ± 0.03 c |
| Incubation Time (h) | Gas Pressure(kPa) | ||
| Blank | AADE | FOS | |
| 6 h | 13.2 ± 0.2 a | 22.8 ± 1.6 b | 29.9 ± 1.6 c |
| 24 h | 27.3 ± 0.6 a | 45.5 ± 0.5 b | 52.0 ± 2.8 c |
| 48 h | 31.4 ± 0.6 a | 48.8 ± 0.6 b | 54.2 ± 1.1 c |
| Incubation Time (h) | 6 h | 24 h | 48 h | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Blank | AADE | FOS | Blank | AADE | FOS | Blank | AADE | FOS | |
| Carbohydrate Metabolite Levels (mean ± SD) (mM) | |||||||||
| Acetate | 8.1 ± 0.3 a | 17.2 ± 1.0 b | 31.4 ± 3.0 c | 19.2 ± 0.3 a | 34.9 ± 0.3 b | 40.1 ± 0.4 c | 20.8 ± 0.3 a | 37.1 ± 0.9 b | 43.7 ± 0.7 c |
| Butyrate | 0.41 ± 0.03 | 0.26 ± 0.05 | 0.48 ± 0.01 | 2.47 ± 0.03 a | 2.89 ± 0.53 a | 5.53 ± 0.64 b | 3.78 ± 0.02 a | 3.9 ± 0.55 a | 7.17 ± 0.22 b |
| Propionate | 3.5 ± 0.2 a | 7.6 ± 0.6 b | 8.7 ± 0.8 c | 7.1 ± 0.1 a | 15.6 ± 0.2 b | 23.6 ± 0.6 c | 7.8 ± 0.20 a | 16.3 ± 0.4 b | 24.1 ± 0.6 c |
| Total SCFAs | 12.1 ± 0.5 a | 25.0 ± 1.6 b | 40.6 ± 3.9 c | 31.6 ± 0.4 a | 56.7 ± 0.5 b | 69.5 ± 0.9 c | 38.3 ± 0.7 a | 65.6 ± 1.2 b | 75.7 ± 0.8 c |
| Lactate (mean ± SD) (mM) | 1.55 ± 0.03 a | 3.79 ± 0.04 b | 8.84 ± 1.16 c | 0.25 ± 0.01 | 0.33 ± 0.07 | 0.85 ± 0.78 | 0.49 ± 0.05 | 0.47 ± 0.21 | 0.10 ± 0.07 |
| Protein Metabolite Levels (mean ± SD) (mg/L) | |||||||||
| BCFAs | 0.10 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 1.34 ± 0.24 a | 1.70 ± 0.37 a | 0.00 ± 0.00 b | 4.01 ± 0.13 a | 3.42 ± 0.02 b | 0.34 ± 0.26 c |
| Ammonium | 0 ± 0 | 0 ± 0 | 0 ± 0 | 328 ± 1 a | 322 ± 7 a | 122 ± 5 b | 414 ± 3 a | 384 ± 13 b | 1874 ± 12 c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van den Abbeele, P.; Ghyselinck, J.; Marzorati, M.; Villar, A.; Zangara, A.; Smidt, C.R.; Risco, E. In Vitro Evaluation of Prebiotic Properties of a Commercial Artichoke Inflorescence Extract Revealed Bifidogenic Effects. Nutrients 2020, 12, 1552. https://doi.org/10.3390/nu12061552
Van den Abbeele P, Ghyselinck J, Marzorati M, Villar A, Zangara A, Smidt CR, Risco E. In Vitro Evaluation of Prebiotic Properties of a Commercial Artichoke Inflorescence Extract Revealed Bifidogenic Effects. Nutrients. 2020; 12(6):1552. https://doi.org/10.3390/nu12061552
Chicago/Turabian StyleVan den Abbeele, Pieter, Jonas Ghyselinck, Massimo Marzorati, Agusti Villar, Andrea Zangara, Carsten R. Smidt, and Ester Risco. 2020. "In Vitro Evaluation of Prebiotic Properties of a Commercial Artichoke Inflorescence Extract Revealed Bifidogenic Effects" Nutrients 12, no. 6: 1552. https://doi.org/10.3390/nu12061552
APA StyleVan den Abbeele, P., Ghyselinck, J., Marzorati, M., Villar, A., Zangara, A., Smidt, C. R., & Risco, E. (2020). In Vitro Evaluation of Prebiotic Properties of a Commercial Artichoke Inflorescence Extract Revealed Bifidogenic Effects. Nutrients, 12(6), 1552. https://doi.org/10.3390/nu12061552