Beneficial Effects of a Low-dose of Conjugated Linoleic Acid on Body Weight Gain and other Cardiometabolic Risk Factors in Cafeteria Diet-fed Rats

 ,
, 
 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. The Conjugated Linoleic Acid
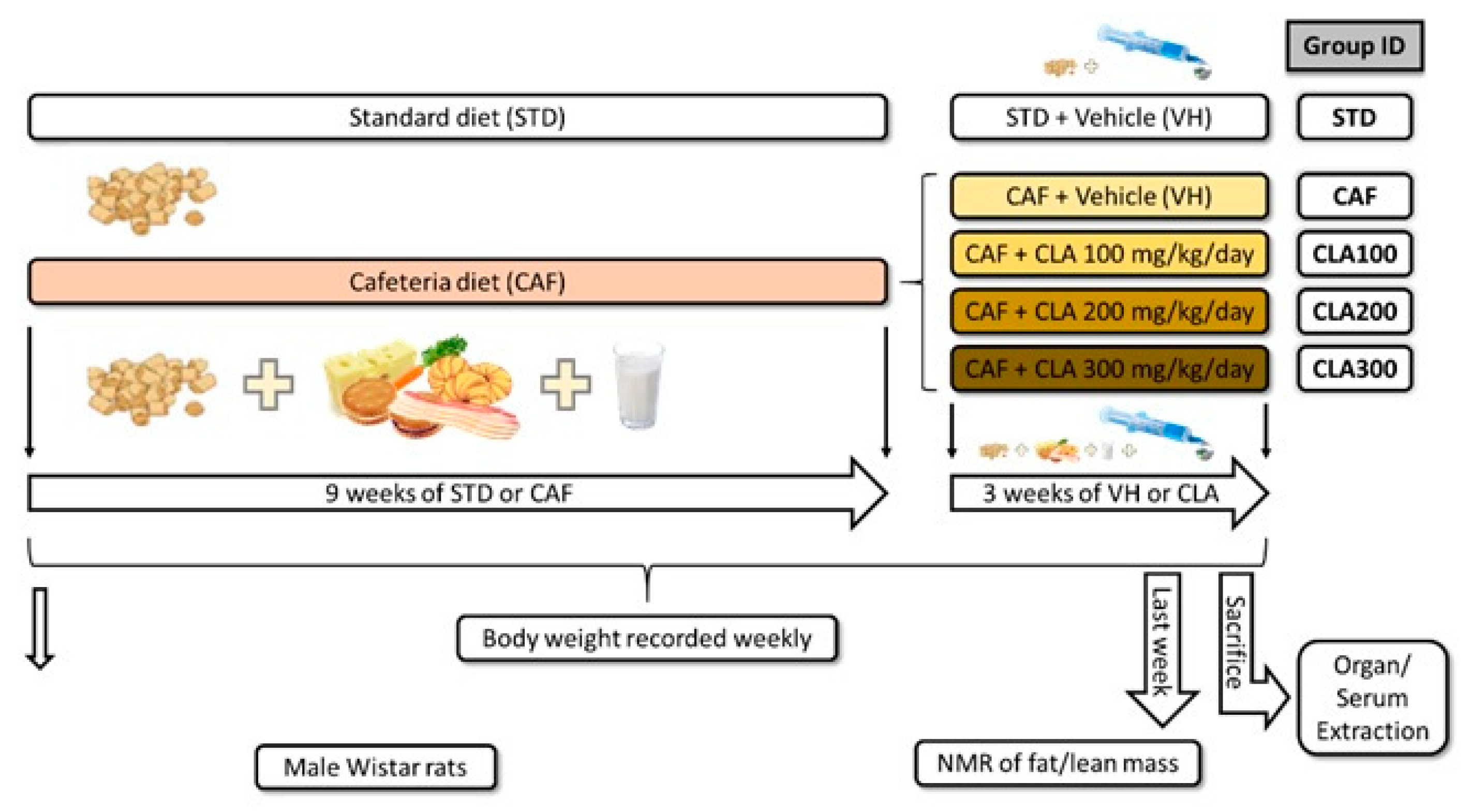
2.2. Experimental Procedure
2.3. Biochemical and Histopathological Analyses
2.4. Western Blot Analysis
2.5. Metabolite Extraction Procedure for 1H NMR Spectrometry
2.6. 1H NMR Spectrometry
2.7. Gene Expression Analysis
2.8. Statistical Analysis
3. Results
3.1. Body Weight Gain, Body Composition and Serum Leptin Levels
3.2. Glucose and Insulin Metabolism
3.3. Serum Lipid Profile
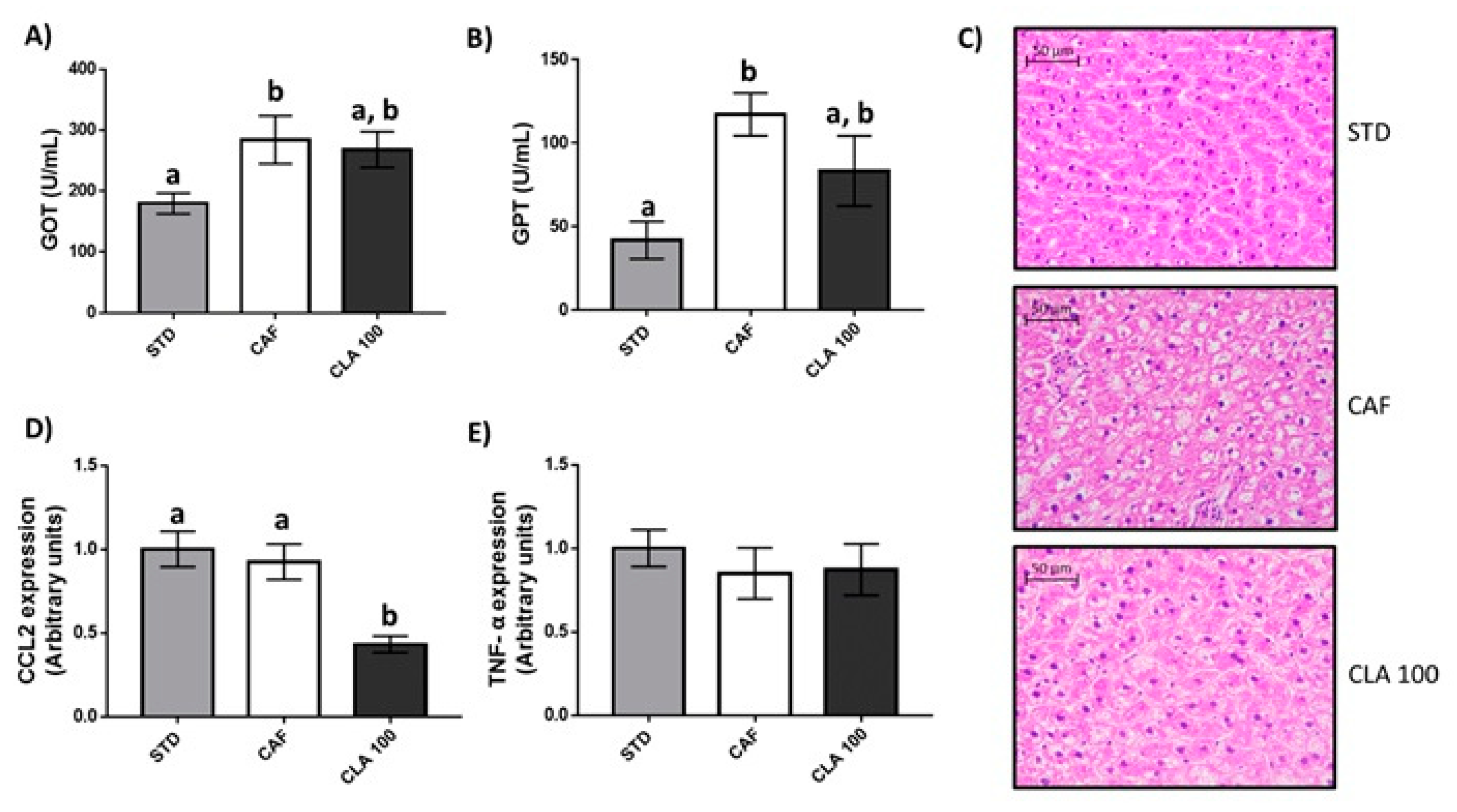
3.4. Hepatic Fat Accumulation and Liver Function
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bremer, A.A.; Mietus-Snyder, M.; Lustig, R.H. Toward a Unifying Hypothesis of Metabolic Syndrome. Pediatrics 2012, 129, 557–570. [Google Scholar] [CrossRef]
- Lent-Schochet, D.; McLaughlin, M.; Ramakrishnan, N.; Jialal, I. Exploratory metabolomics of metabolic syndrome: A status report. World J. Diabetes 2019, 10, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Rhee, E.P.; Larson, M.G.; Lewis, G.D.; McCabe, E.L.; Shen, D.; Palma, M.J.; Roberts, L.D.; Dejam, A.; Souza, A.L.; et al. Metabolite profiling identifies pathways associated with metabolic risk in humans. Circulation 2012, 125, 2222–2231. [Google Scholar] [CrossRef] [PubMed]
- Newgard, C.B. Interplay between lipids and branched-chain amino acids in development of insulin resistance. Cell Metab. 2012, 15, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Suliman, M.E.; Qureshi, A.R.; Garcia-Lopez, E.; Bárány, P.; Heimbürger, O.; Stenvinkel, P.; Lindholm, B. Consequences of low plasma histidine in chronic kidney disease patients: Associations with inflammation, oxidative stress, and mortality. Am. J. Clin. Nutr. 2008, 87, 1860–1866. [Google Scholar] [CrossRef]
- van der Veen, J.N.; Kennelly, J.P.; Wan, S.; Vance, J.E.; Vance, D.E.; Jacobs, R.L. The critical role of phosphatidylcholine and phosphatidylethanolamine metabolism in health and disease. Biochim. Biophys. Acta Biomembr. 2017, 1859, 1558–1572. [Google Scholar] [CrossRef] [PubMed]
- Sampey, B.P.; Vanhoose, A.M.; Winfield, H.M.; Freemerman, A.J.; Muehlbauer, M.J.; Fueger, P.T.; Newgard, C.B.; Makowski, L. Cafeteria diet is a robust model of human metabolic syndrome with liver and adipose inflammation: Comparison to high-fat diet. Obesity 2011, 19, 1109–1117. [Google Scholar] [CrossRef]
- Pons, Z.; Margalef, M.; Bravo, F.I.; Arola-Arnal, A.; Muguerza, B. Acute administration of single oral dose of grape seed polyphenols restores blood pressure in a rat model of metabolic syndrome: role of nitric oxide and prostacyclin. Eur. J. Nutr. 2016, 55, 749–758. [Google Scholar] [CrossRef]
- Evans, M.E.; Brown, J.M.; McIntosh, M.K. Isomer-specific effects of conjugated linoleic acid (CLA) on adiposity and lipid metabolism. J. Nutr. Biochem. 2002, 13, 508–516. [Google Scholar] [CrossRef]
- Whigham, L.D.; Watras, A.C.; Schoeller, D.A. Efficacy of conjugated linoleic acid for reducing fat mass: a meta-analysis in humans. Am. J. Clin. Nutr. 2007, 85, 1203–1211. [Google Scholar] [CrossRef]
- Pariza, M.W.; Park, Y.; Cook, M.E. The biologically active isomers of conjugated linoleic acid. Prog. Lipid Res. 2001, 40, 283–298. [Google Scholar] [CrossRef]
- Shen, W.; Baldwin, J.; Collins, B.; Hixson, L.; Lee, K.T.; Herberg, T.; Starnes, J.; Cooney, P.; Chuang, C.C.; Hopkins, R.; et al. Low level of trans-10, cis-12 conjugated linoleic acid decreases adiposity and increases browning independent of inflammatory signaling in overweight Sv129 mice. J. Nutr. Biochem. 2015, 26, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.R.; Park, Y.; Albright, K.J.; Pariza, M.W. Differential responses of hamsters and rats fed high-fat or low-fat diets supplemented with conjugated linoleic acid. Nutr. Res. 2002, 22, 715–722. [Google Scholar] [CrossRef]
- Koba, K.; Akahoshi, A.; Yamasaki, M.; Tanaka, K.; Yamada, K.; Iwata, T.; Kamegai, T.; Tsutsumi, K.; Sugano, M. Dietary conjugated linolenic acid in relation to CLA differently modifies body fat mass and serum and liver lipid levels in rats. Lipids 2002, 37, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Ostrowska, E.; Muralitharan, M.; Cross, R.F.; Bauman, D.E.; Dunshea, F.R. Dietary Conjugated Linoleic Acids Increase Lean Tissue and Decrease Fat Deposition in Growing Pigs. J. Nutr. 2018, 129, 2037–2042. [Google Scholar] [CrossRef]
- Szymczyk, B.; Pisulewski, P.M.; Szczurek, W.; Hanczakowski, P. Effects of conjugated linoleic acid on growth performance, feed conversion efficiency, and subsequent carcass quality in broiler chickens. Br. J. Nutr. 2001, 85, 465–473. [Google Scholar] [CrossRef]
- Benjamin, S.; Prakasan, P.; Sreedharan, S.; Wright, A.D.; Spener, F. Pros and cons of CLA consumption: An insight from clinical evidences. Nutr. Metab. (Lond.) 2015, 12, 4. [Google Scholar] [CrossRef]
- Lamarche, B.; Desroches, S. Metabolic syndrome and effects of conjugated linoleic acid in obesity and lipoprotein disorders: the Québec experience. Am. J. Clin. Nutr. 2004, 79, 1149S–1152S. [Google Scholar] [CrossRef]
- Tsuboyama-Kasaoka, N.; Takahashi, M.; Tanemura, K.; Kim, H.J.; Tange, T.; Okuyama, H.; Kasai, M.; Ikemoto, S.; Ezaki, O. Conjugated linoleic acid supplementation reduces adipose tissue by apoptosis and develops lipodystrophy in mice. Diabetes 2000, 49, 1534–1542. [Google Scholar] [CrossRef]
- Poirier, H.; Shapiro, J.S.; Kim, R.J.; Lazar, M.A. Nutritional supplementation with trans-10, cis-12-conjugated linoleic acid induces inflammation of white adipose tissue. Diabetes 2006, 55, 1634–1641. [Google Scholar] [CrossRef] [PubMed]
- Vyas, D.; Kadegowda, A.K.G.; Erdman, R.A. Dietary conjugated linoleic acid and hepatic steatosis: Species-specific effects on liver and adipose lipid metabolism and gene expression. J. Nutr. Metab. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA). Scientific Opinion on the safety of “conjugated linoleic acid (CLA)-rich oil” (Clarinol®) as a Novel Food ingredient. EFSA J. 2010, 8, 1601. [Google Scholar] [CrossRef]
- Ulmer, C.Z.; Jones, C.M.; Yost, R.A.; Garrett, T.J.; Bowden, J.A. Optimization of Folch, Bligh-Dyer, and Matyash Sample-to-Extraction Solvent Ratios for Human Plasma-Based Lipidomics Studies. Anal. Chim. Acta 2018, 1037, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Vinaixa, M.; Rodríguez, M.A.; Rull, A.; Beltrán, R.; Bladé, C.; Brezmes, J.; Cañellas, N.; Joven, J.; Correig, X. Metabolomic assessment of the effect of dietary cholesterol in the progressive development of fatty liver disease. J. Proteome Res. 2010, 9, 2527–2538. [Google Scholar] [CrossRef]
- Egras, A.M.; Hamilton, W.R.; Lenz, T.L.; Monaghan, M.S. An evidence-based review of fat modifying supplemental weight loss products. J. Obes. 2011, 2011, 297515. [Google Scholar] [CrossRef]
- Bezan, P.N.; Holland, H.; De Castro, G.S.; Cardoso, J.F.R.; Ovidio, P.P.; Calder, P.C.; Jordao, A.A. High dose of a conjugated linoleic acid mixture increases insulin resistance in rats fed either a low fat or a high fat diet. Exp. Clin. Endocrinol. Diabetes 2018, 126, 379–386. [Google Scholar] [CrossRef]
- DeLany, J.P.; Blohm, F.; Truett, A.A.; Scimeca, J.A.; West, D.B. Conjugated linoleic acid rapidly reduces body fat content in mice without affecting energy intake. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1999, 276, R1172–1179. [Google Scholar] [CrossRef]
- Taylor, C.G.; Zahradka, P. Dietary conjugated linoleic acid and insulin sensitivity and resistance in rodent models. Am. J. Clin. Nutr. 2004, 79, 1164–1168. [Google Scholar] [CrossRef]
- Park, Y.; Storkson, J.M.; Albright, K.J.; Liu, W.; Pariza, M.W. Evidence that the trans-10,cis-12 isomer of conjugated linoleic acid induces body composition changes in mice. Lipids. 1999, 34, 235–241. [Google Scholar] [CrossRef]
- Engl, J.; Sturm, W.; Sandhofer, A.; Kaser, S.; Tschoner, A.; Tatarczyk, T.; Weiss, H.; Tilg, H.; Patsch, J.R.; Ebenbichler, C.F. Effect of pronounced weight loss on visceral fat, liver steatosis and adiponectin isoforms. Eur. J. Clin. Investig. 2008, 38, 238–244. [Google Scholar] [CrossRef]
- Mollica, M.P.; Trinchese, G.; Cavaliere, G.; De Filippo, C.; Cocca, E.; Gaita, M.; Della-Gatta, A.; Marano, A.; Mazzarella, G.; Bergamo, P. c9;t11-Conjugated linoleic acid ameliorates steatosis by modulating mitochondrial uncoupling and Nrf2 pathway. J. Lipid Res. 2014, 55, 837–849. [Google Scholar] [CrossRef] [PubMed]
- Parra, P.; Serra, F.; Palou, A. Moderate doses of conjugated linoleic acid isomers mix contribute to lowering body fat content maintaining insulin sensitivity and a noninflammatory pattern in adipose tissue in mice. J. Nutr. Biochem. 2010, 21, 107–115. [Google Scholar] [CrossRef]
- Willoughby, D.; Hewlings, S.; Kalman, D. Body composition changes in weight loss: strategies and supplementation for maintaining lean body mass: A brief review. Nutrients 2018, 10, 1876. [Google Scholar] [CrossRef] [PubMed]
- Pinto Júnior, D.A.C.; Seraphim, P.M. Cafeteria diet intake for fourteen weeks can cause obesity and insulin resistance in wistar rats. Rev. Nutr. 2012, 25, 313–319. [Google Scholar] [CrossRef]
- Petersen, M.C.; Vatner, D.F.; Shulman, G.I. Regulation of hepatic glucose metabolism in health and disease. Nat. Rev. Endocrinol. 2017, 13, 572–587. [Google Scholar] [CrossRef]
- Rui, L. Energy metabolism in the liver. Compr. Physiol. 2014, 4, 177–197. [Google Scholar] [PubMed]
- Chen, X.; Yang, W. Branched-chain amino acids and the association with type 2 diabetes. J. Diabetes Investig. 2015, 6, 369–370. [Google Scholar] [CrossRef] [PubMed]
- Connelly, M.A.; Wolak-Dinsmore, J.; Dullaart, R.P.F. Branched chain amino acids are associated with insulin resistance independent of leptin and adiponectin in subjects with varying degrees of glucose tolerance. Metab. Syndr. Relat. Disord. 2017, 15, 183–186. [Google Scholar] [CrossRef]
- Yu, C.; Chen, Y.; Cline, G.W.; Zhang, D.; Zong, H.; Wang, Y.; Bergeron, R.; Kim, J.K.; Cushman, S.W.; Cooney, G.J.; et al. Mechanism by which fatty acids inhibit insulin activation of insulin receptor substrate-1 (IRS-1)-associated phosphatidylinositol 3-kinase activity in muscle. J. Biol. Chem. 2002, 277, 50230–50236. [Google Scholar] [CrossRef]
- Arnold, R.S.; Newton, A.C. Diacylglycerol directly stimulates the insulin receptor tyrosine kinase. FEBS Lett. 1996, 380, 58–62. [Google Scholar] [CrossRef]
- Erion, D.M.; Shulman, G.I. Diacylglycerol-mediated insulin resistance. Nat. Med. 2010, 16, 400–402. [Google Scholar] [CrossRef] [PubMed]
- Kloss, R.; Linscheid, J.; Johnson, A.; Lawson, B.; Edwards, K.; Linder, T.; Stocker, K.; Petitte, J.; Kern, M. Effects of conjugated linoleic acid supplementation on blood lipids and adiposity of rats fed diets rich in saturated versus unsaturated fat. Pharmacol. Res. 2005, 51, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Blankson, H.; Stakkestad, J.A.; Fagertun, H.; Thom, E.; Wadstein, J.; Gudmundsen, O. Conjugated Linoleic Acid Reduces Body Fat Mass in Overweight and Obese Humans. J. Nutr. 2018, 130, 2943–2948. [Google Scholar] [CrossRef] [PubMed]
- Belury, M.A. Dietary conjugated linoleic acid in health: Physiological Effects and Mechanisms of Action. Annu. Rev. Nutr. 2002, 22, 505–531. [Google Scholar] [CrossRef] [PubMed]
- Valeille, K.; Gripois, D.; Blouquit, M.-F.; Souidi, M.; Riottot, M.; Bouthegourd, J.-C.; Sérougne, C.; Martin, J.-C. Lipid atherogenic risk markers can be more favourably influenced by the cis -9, trans -11-octadecadienoate isomer than a conjugated linoleic acid mixture or fish oil in hamsters. Br. J. Nutr. 2004, 91, 191–199. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tricon, S.; Burdge, G.C.; Kew, S.; Banerjee, T.; Russell, J.J.; Jones, E.L.; Grimble, R.F.; Williams, C.M.; Yaqoob, P.; Calder, P.C. Opposing effects of cis-9,trans-11 and trans-10,cis-12 conjugated linoleic acid on blood lipids in healthy humans. Am. J. Clin. Nutr. 2004, 80, 614–620. [Google Scholar] [CrossRef]
- Clément, L.; Poirier, H.; Niot, I.; Bocher, V.; Guerre-Millo, M.; Krief, S.; Staels, B.; Besnard, P. Dietary trans-10,cis-12 conjugated linoleic acid induces hyperinsulinemia and fatty liver in the mouse. J. Lipid Res. 2002, 43, 1400–1409. [Google Scholar] [CrossRef]
- Ide, T. Interaction of fish oil and conjugated linoleic acid in affecting hepatic activity of lipogenic enzymes and gene expression in liver and adipose tissue. Diabetes 2005, 54, 412–423. [Google Scholar] [CrossRef]
- Takahashi, Y.; Kushiro, M.; Shinohara, K.; Ide, T. Activity and mRNA levels of enzymes involved in hepatic fatty acid synthesis and oxidation in mice fed conjugated linoleic acid. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2003, 1631, 265–273. [Google Scholar] [CrossRef]
- Javadi, M.; Beynen, A.C.; Hovenier, R.; Lankhorst, Æ.; Lemmens, A.G.; Terpstra, A.H.M.; Geelen, M.J.H. Prolonged feeding of mice with conjugated linoleic acid increases hepatic fatty acid synthesis relative to oxidation. J. Nutr. Biochem. 2004, 15, 680–687. [Google Scholar] [CrossRef]
- Park, Y.; Pariza, M.W. Mechanisms of body fat modulation by conjugated linoleic acid (CLA). Food Res. Int. 2007, 40, 311–323. [Google Scholar] [CrossRef]
- Belury, M.A.; Kempa-Steczko, A. Conjugated linoleic acid modulates hepatic lipid composition in mice. Lipids 1997, 32, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Kelley, D.S.; Bartolini, G.L.; Warren, J.M.; Simon, V.A.; Mackey, B.E.; Erickson, K.L. Contrasting effects of t 10, c 12- and c 9, t 11-conjugated linoleic acid isomers on the fatty acid profiles of mouse liver lipids. Lipids 2004, 39, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-Y.; Ma, K.Y.; Liang, Y.; Peng, C.; Zuo, Y. Role and classification of cholesterol-lowering functional foods. J. Funct. Foods. 2011, 3, 61–69. [Google Scholar] [CrossRef]
- Reynolds, C.M.; Roche, H.M. Conjugated linoleic acid and inflammatory cell signalling. Prostaglandins Leukot. Essent. Fat. Acids 2010, 82, 199–204. [Google Scholar] [CrossRef]
- Haghighatdoost, F.; Nobakht, M.; Gh, B.F. Effect of conjugated linoleic acid on blood inflammatory markers: a systematic review and meta-analysis on randomized controlled trials. Eur. J. Clin. Nutr. 2018, 72, 1071–1082. [Google Scholar] [CrossRef]
- Derakhshandeh-Rishehri, S.-M.; Rahbar, A.R.; Ostovar, A. Effects of Conjugated Linoleic Acid Intake in the Form of Dietary Supplement or Enriched Food on C-Reactive Protein and Lipoprotein (a) Levels in Humans: A Literature Review and Meta-Analysis. Iran. J. Med. Sci. 2019, 44, 359–373. [Google Scholar]
- Dipasquale, D.; Basiricò, L.; Morera, P.; Primi, R.; Tröscher, A.; Bernabucci, U. Anti-inflammatory effects of conjugated linoleic acid isomers and essential fatty acids in bovine mammary epithelial cells. Animal 2018, 12, 2108–2114. [Google Scholar] [CrossRef]
- Song, H.J.; Grant, I.; Rotondo, D.; Mohede, I.; Sattar, N.; Heys, S.D.; Wahle, K.W.J. Effect of CLA supplementation on immune function in young healthy volunteers. Eur. J. Clin. Nutr. 2005, 59, 508–517. [Google Scholar] [CrossRef]
- Pingitore, P.; Sasidharan, K.; Ekstrand, M.; Prill, S.; Lindén, D.; Romeo, S. Human Multilineage 3D Spheroids as a Model of Liver Steatosis and Fibrosis. Int. J. Mol. Sci. 2019, 20, 1629. [Google Scholar] [CrossRef]
- Ipsen, D.H.; Lykkesfeldt, J.; Tveden-Nyborg, P. Molecular mechanisms of hepatic lipid accumulation in non-alcoholic fatty liver disease. Cell. Mol. Life Sci. 2018, 75, 3313–3327. [Google Scholar] [CrossRef] [PubMed]
- Tilak, J.C.; Adhikari, S.; Devasagayam, T.P.A. Antioxidant properties of Plumbago zeylanica, an Indian medicinal plant and its active ingredient, plumbagin. Redox Rep. 2004, 9, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| STD | CAF | CLA 100 | |
|---|---|---|---|
| O-Acetylcarnitine | 0.113 ± 0.01 | 0.119 ± 0.01 | 0.11 ± 0.01 |
| Formate | 0.42 ± 0.03 | 0.42 ± 0.01 | 0.46 ± 0.02 |
| Glycerol | 1.32 ± 0.1 a | 1.71 ± 0.1 b | 1.52 ± 0.2 a,b |
| Acetate | 3.52 ± 0.1 | 3.65 ± 0.1 | 3.77 ± 0.1 |
| 3-Hidroxybutyrate | 1.25 ± 0.2 | 0.87 ± 0.1 | 1.21 ± 0.3 |
| Glucose | 2.00 ± 0.11 a | 2.43 ± 0.05 b | 2.13 ± 0.03 a |
| Piruvate | 0.44 ± 0.1 | 0.64 ± 0.1 | 0.6 ± 0.1 |
| Succinate | 0.19 ± 0.01 | 0.17 ± 0.01 | 0.2 ± 0.02 |
| Lactate | 20.13 ± 2.7 | 24.56 ± 3.1 | 24.14 ± 3.2 |
| Citrate | 0.17 ± 0.01 a | 0.22 ± 0.01 b | 0.18 ± 0.01 a |
| Asparagine | 0.46 ± 0.02 | 0.51 ± 0.04 | 0.53 ± 0.02 |
| Leucine | 0.97 ± 0.02 | 0.94 ± 0.08 | 0.96 ± 0.04 |
| Threonine | 1.35 ± 0.1 a | 1.67 ± 0.1 b | 1.57 ± 0.1 a,b |
| Tryptophan | 0.74 ± 0.04 | 0.87 ± 0.06 | 0.8 ± 0.05 |
| Tyrosine | 0.61 ± 0.04 | 0.76 ± 0.07 | 0.65 ± 0.03 |
| Proline | 0.89 ± 0.1 | 1.14 ± 0.1 | 1.04 ± 0.1 |
| Isoleucine | 0.68 ± 0.02 | 0.62 ± 0.05 | 0.6 ± 0.03 |
| Glycine | 1.9 ± 0.1 | 1.57 ± 0.1 | 1.64 ± 0.13 |
| Glutamate | 1.01 ± 0.05 | 1.11 ± 0.07 | 1.11 ± 0.04 |
| Glutamine | 5.52 ± 0.1 | 5.51 ± 0.4 | 5.39 ± 0.1 |
| Methionine | 0.61 ± 0.01 | 0.69 ± 0.04 | 0.61 ± 0.01 |
| Lysine | 2.3 ± 0.1 | 2.45 ± 0.2 | 2.57 ± 0.2 |
| Valine | 1.11 ± 0.1 | 1.17 ± 0.1 | 1.11 ± 0.1 |
| Serine | 2.2 ± 0.1 a | 2.6 ± 0.2 a,b | 2.75 ± 0.1 b |
| Alanine | 3.06 ± 0.1 | 3.99 ± 0.5 | 3.54 ± 0.2 |
| Phenylalanine | 0.56 ± 0.01 | 0.61 ± 0.03 | 0.55 ± 0.01 |
| Taurine | 0.29 ± 0.02 | 0.29 ± 0.01 | 0.31 ± 0.01 |
| Carnosine | 0.26 ± 0.01 | 0.23 ± 0.03 | 0.22 ± 0.01 |
| Choline | 0.16 ± 0.005 a | 0.14 ± 0.008 b | 0.14 ± 0.004 b |
| Betaine | 0.82 ± 0.07 a | 0.54 ± 0.04 b | 0.58 ± 0.03 b |
| Creatinine | 0.09 ± 0.003 | 0.08 ± 0.007 | 0.09 ± 0.003 |
| Glutathione | 0.04 ± 0.01 | 0.05 ± 0.01 | 0.05 ± 0.01 |
| Allantoin | 0.24 ± 0.01 a | 0.19 ± 0.001 b | 0.2 ± 0.01 b |
| Creatine | 1.95 ± 0.2 | 2.57 ± 0.2 | 2.35 ± 0.2 |
| Creatine phosphate | 0.3 ± 0.01 | 0.33 ± 0.03 | 0.27 ± 0.01 |
| Pantothenate | 0.06 ± 0.002 | 0.07 ± 0.01 | 0.05 ± 0.007 |
| STD | CAF | CLA 100 | |
|---|---|---|---|
| Valine | 0.48 ± 0.02 | 0.57 ± 0.04 | 0.49 ± 0.04 |
| Isoleucine | 0.3 ± 0.01 a | 0.27 ± 0.02 a,b | 0.22 ± 0.01 b |
| Leucine | 1.17 ± 0.1 | 1.18 ± 0.1 | 1.04 ± 0.1 |
| Glycine | 0.1 ± 0.016 | 0.1 ± 0.007 | 0.09 ± 0.015 |
| Alanine | 3.8 ± 0.2 a | 4.99 ± 0.2 b | 3.54 ± 0.3 a |
| Glutamine | 5.51 ± 0.4 | 4.47 ± 0.4 | 5.13 ± 0.3 |
| Tyrosine | 0.27 ± 0.009 a | 0.26 ± 0.015 a | 0.2 ± 0.014 b |
| Histidine | 0.61 ± 0.03 | 0.57 ± 0.03 | 0.53 ± 0.02 |
| Methionine | 0.33 ± 0.01 a | 0.25 ± 0.01 b | 0.23 ± 0.01 b |
| Glutamate | 2.26 ± 0.1 | 2.89 ± 0.2 | 2.41 ± 0.3 |
| Phenylalanine | 0.8 ± 0.05 | 0.84 ± 0.07 | 0.7 ± 0.04 |
| Glucose | 32.38 ± 2.6 a | 35.79 ± 1.0 b | 27.95 ± 2.6 a,b |
| Succinate | 1.5 ± 0.1 | 1.23 ± 0.5 | 1.54 ± 0.3 |
| Acetate | 0.37 ± 0.02 | 0.47 ± 0.05 | 0.5 ± 0.13 |
| Lactate | 11.26 ± 1.0 a | 14.54 ± 1.0 b | 11.76 ± 0.8 a,b |
| Fumarate | 0.09 ± 0.009 | 0.07 ± 0.008 | 0.1 ± 0.01 |
| NAD+ | 0.64 ± 0.04 | 0.54 ± 0.1 | 0.64 ± 0.03 |
| NADP+ | 0.37 ± 0.03 | 0.32 ± 0.03 | 0.34 ± 0.02 |
| 3-Hydroxybutyrate | 0.47 ± 0.05 | 0.35 ± 0.02 | 0.41 ± 0.05 |
| Uridine | 0.55 ± 0.02 | 0.56 ± 0.09 | 0.43 ± 0.02 |
| Choline | 0.12 ± 0.005 a | 0.09 ± 0.005 a,b | 0.08 ± 0.016 b |
| Phosphocholine | 1.48 ± 0.1 | 1.13 ± 0.2 | 1.05 ± 0.3 |
| Beatine | 2.32 ± 0.2 a | 1.08 ± 0.1b | 1.04 ± 0.2 b |
| Glutathione | 4.07 ± 0.8 | 2.69 ± 0.4 | 3.15 ± 0.6 |
| Niacinamide | 0.3 ± 0.02 a | 0.31 ± 0.01 a | 0.22 ± 0.01 b |
| Ascorbate | 1.46 ± 0.1 | 1.52 ± 0.22 | 1.6 ± 0.1 |
| Dimethylamine | 0.04 ± 0.003 a | 0.03 ± 0.001 a,b | 0.02 ± 0.002 b |
| Inosine | 1.94 ± 0.1 | 1.98 ± 0.2 | 1.59 ± 0.1 |
| Creatinine | 0.86 ± 0.2 | 0.59 ± 0.1 | 0.44 ± 0.1 |
| Creatine phosphate | 1.86 ± 0.1 | 2.00 ± 0.3 | 1.59 ± 0.1 |
| Creatine | 0.09 ± 0.005 | 0.09 ± 0.01 | 0.08 ± 0.01 |
| STD | CAF | CLA 100 | |
|---|---|---|---|
| Total Cholesterol | 3.12 ± 0.8 a | 4.98 ± 0.1 b | 4.12 ± 0.1 a,b |
| Free Cholesterol | 1.55 ± 0.2 a | 2.23 ± 0.1 b | 1.94 ± 0.1 a,b |
| Esterified Cholesterol | 2.05 ± 0.5 a | 2.92 ± 0.1 b | 2.53 ± 0.1 a,b |
| Triglycerides | 1.76 ± 0.6 a | 10.91 ± 0.6 b | 6.89 ± 1.4 c |
| Diglycerides | 0.07 ± 0.01 a | 0.18 ± 0.01 b | 0.12 ± 0.02 c |
| Total Phospholipids | 1.62 ± 0.4 a | 3.95 ± 0.1 b | 3.01 ± 0.2 c |
| Linoleic acid | 1.25 ± 0.4 a | 3.42 ± 0.1 b | 2.09 ± 0.1 c |
| Oleic acid | 1.71 ± 0.6 a | 14.33 ± 0.7 b | 8.88 ± 2 c |
| Sphingomyelin | 0.51 ± 0.03 | 0.48 ± 0.01 | 0.49 ± 0.02 |
| ARA + EPA | 1.97 ± 0.5 | 2.22 ± 0.2 | 1.79 ± 0.1 |
| DHA | 0.13 ± 0.03 a | 0.26 ± 0.02 b | 0.2 ± 0.01 a,b |
| Omega-3 | 0.6 ± 0.14 a | 0.98 ± 0.05 b | 0.75 ± 0.02 b |
| Phosphocholine | 2.84 ± 0.7 a | 5.83 ± 0.2 b | 4.63 ± 0.2 b |
| PUFA | 6.84 ± 1.8 | 9.56 ± 0.8 | 7.6 ± 0.2 |
| MUFA | 4.94 ± 1.5 a | 21.24 ± 1 b | 13.12 ± 2.4 c |
| STD | CAF | CLA 100 | |
|---|---|---|---|
| Steatosis degree (0 to 3 in severity) | 0.67 ± 0.3 a | 1.50 ± 0.2 b | 1.33 ± 0.2 a,b |
| Sinusoidal dilatation (0 to 2 in severity) | 0.17 ± 0.2 | 0.33 ± 0.2 | 0.67 ± 0.2 |
| Microgranulomes (number of samples) | 2/6 | 1/6 | 2/6 |
| Fibrosis degree (0 to 4 in severity) | 0 | 0 | 0 |
| Portal inflammation (0 to 2 in severity) | 0 | 0 | 0 |
| Lipogranulomes (number of samples) | 0/6 | 0/6 | 0/6 |
| STD | CAF | CLA 100 | |
|---|---|---|---|
| Total Cholesterol | 9.42 ± 0.7 a | 16.91 ± 2.6 a,b | 24.53 ± 4.6 b |
| Free Cholesterol | 1.26 ± 0.08 | 1.38 ± 0.16 | 1.41 ± 0.13 |
| Esterified Cholesterol | 1.05 ± 0.1 a | 7.83 ± 1.9 a,b | 15.17 ± 4.5 b |
| Triglycerides | 42.02 ± 3.5 a | 81.03 ± 11.3 b | 87.01 ± 11.1 b |
| Diglycerides | 7.14 ± 0.7 a | 11.51 ± 1.1 b | 9.37 ± 1 a,b |
| Sphingomyelin | 2.88 ± 0.1 | 2.76 ± 0.4 | 2.84 ± 0.3 |
| ARA + EPA | 28.66 ± 2.3 | 25.02 ± 3.5 | 25.61 ± 2 |
| Plasmalogen | 1.36 ± 0.1 a | 1.62 ± 0.2 a | 2.58 ± 0.3 b |
| Total Phospholipids | 48.1 ± 5.5 | 45.42 ± 7.4 | 45.34 ± 5.5 |
| Phosphoethanolamine | 17.37 ± 3.1 | 20.32 ± 2.6 | 20.68 ± 1.7 |
| Linoleic acid | 32.29 ± 2.8 | 36.05 ± 6.1 | 44.94 ± 5.7 |
| Oleic acid | 16.96 ± 2.4 a | 86.78 ± 14.6 b | 87 ± 16.3 b |
| Omega-3 | 0.05 ± 0.002 a | 0.03 ± 0.004 b | 0.03 ± 0.002 b |
| DHA | 4.19 ± 0.6 | 4.22 ± 0.8 | 4.64 ± 0.2 |
| PUFA | 120.63 ± 10.5 | 112.11 ± 15.6 | 118.57 ± 7.1 |
| MUFA | 38.89 ± 4.5 a | 109.45 ± 19.6 b | 120.16 ± 23.7 b |
| Total fatty acids | 150.44 ± 9.8 a | 277.54 ± 35.5 b | 290.55 ± 9.8 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martín-González, M.Z.; Palacios, H.; Rodríguez, M.A.; Arola, L.; Aragonès, G.; Muguerza, B. Beneficial Effects of a Low-dose of Conjugated Linoleic Acid on Body Weight Gain and other Cardiometabolic Risk Factors in Cafeteria Diet-fed Rats. Nutrients 2020, 12, 408. https://doi.org/10.3390/nu12020408
Martín-González MZ, Palacios H, Rodríguez MA, Arola L, Aragonès G, Muguerza B. Beneficial Effects of a Low-dose of Conjugated Linoleic Acid on Body Weight Gain and other Cardiometabolic Risk Factors in Cafeteria Diet-fed Rats. Nutrients. 2020; 12(2):408. https://doi.org/10.3390/nu12020408
Chicago/Turabian StyleMartín-González, Miguel Z, Héctor Palacios, Miguel A Rodríguez, Lluís Arola, Gerard Aragonès, and Begoña Muguerza. 2020. "Beneficial Effects of a Low-dose of Conjugated Linoleic Acid on Body Weight Gain and other Cardiometabolic Risk Factors in Cafeteria Diet-fed Rats" Nutrients 12, no. 2: 408. https://doi.org/10.3390/nu12020408
APA StyleMartín-González, M. Z., Palacios, H., Rodríguez, M. A., Arola, L., Aragonès, G., & Muguerza, B. (2020). Beneficial Effects of a Low-dose of Conjugated Linoleic Acid on Body Weight Gain and other Cardiometabolic Risk Factors in Cafeteria Diet-fed Rats. Nutrients, 12(2), 408. https://doi.org/10.3390/nu12020408