Genetic Risk Score Predictive of the Plasma Triglyceride Response to an Omega-3 Fatty Acid Supplementation in a Mexican Population

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Population
2.2. Intervention
2.3. Single-Nucleotide Polymorphisms Selection for Genotyping
2.4. Genotyping
2.5. SNP and Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kraja, A.T.; Borecki, I.B.; Tsai, M.Y.; Ordovas, J.M.; Hopkins, P.N.; Lai, C.Q.; Frazier-Wood, A.C.; Straka, R.J.; Hixson, J.E.; Province, M.A.; et al. Genetic analysis of 16 nmr-lipoprotein fractions in humans, the goldn study. Lipids 2013, 48, 155–165. [Google Scholar] [CrossRef]
- Healey, G.R.; Murphy, R.; Brough, L.; Butts, C.A.; Coad, J. Interindividual variability in gut microbiota and host response to dietary interventions. Nutr. Rev. 2017, 75, 1059–1080. [Google Scholar] [CrossRef] [PubMed]
- Zeevi, D.; Korem, T.; Zmora, N.; Israeli, D.; Rothschild, D.; Weinberger, A.; Ben-Yacov, O.; Lador, D.; Avnit-Sagi, T.; Lotan-Pompan, M.; et al. Personalized nutrition by prediction of glycemic responses. Cell 2015, 163, 1079–1094. [Google Scholar] [CrossRef]
- Jones, P.J. Inter-individual variability in response to plant sterol and stanol consumption. J. AOAC Int. 2015, 98, 724–728. [Google Scholar] [CrossRef]
- De Roos, B.; Brennan, L. Personalised interventions-a precision approach for the next generation of dietary intervention studies. Nutrients 2017, 9, 847. [Google Scholar] [CrossRef]
- Caslake, M.J.; Miles, E.A.; Kofler, B.M.; Lietz, G.; Curtis, P.; Armah, C.K.; Kimber, A.C.; Grew, J.P.; Farrell, L.; Stannard, J.; et al. Effect of sex and genotype on cardiovascular biomarker response to fish oils: The fingen study. Am. J. Clin. Nutr. 2008, 88, 618–629. [Google Scholar] [CrossRef] [PubMed]
- Rudkowska, I.; Paradis, A.M.; Thifault, E.; Julien, P.; Barbier, O.; Couture, P.; Lemieux, S.; Vohl, M.C. Differences in metabolomic and transcriptomic profiles between responders and non-responders to an n-3 polyunsaturated fatty acids (pufas) supplementation. Genes. Nutr. 2013, 8, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Caron-Dorval, D.; Paquet, P.; Paradis, A.M.; Rudkowska, I.; Lemieux, S.; Couture, P.; Vohl, M.C. Effect of the ppar-alpha l162v polymorphism on the cardiovascular disease risk factor in response to n-3 polyunsaturated fatty acids. J. Nutrigenet. Nutrigenomics 2008, 1, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Lindi, V.; Schwab, U.; Louheranta, A.; Laakso, M.; Vessby, B.; Hermansen, K.; Storlien, L.; Riccardi, G.; Rivellese, A. Impact of the pro12ala polymorphism of the ppar-gamma2 gene on serum triacylglycerol response to n-3 fatty acid supplementation. Mol. Genet. Metab. 2003, 79, 52–60. [Google Scholar] [CrossRef]
- Minihane, A.M.; Khan, S.; Leigh-Firbank, E.C.; Talmud, P.; Wright, J.W.; Murphy, M.C.; Griffin, B.A.; Williams, C.M. Apoe polymorphism and fish oil supplementation in subjects with an atherogenic lipoprotein phenotype. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 1990–1997. [Google Scholar] [CrossRef] [PubMed]
- Thifault, E.; Cormier, H.; Bouchard-Mercier, A.; Rudkowska, I.; Paradis, A.M.; Garneau, V.; Ouellette, C.; Lemieux, S.; Couture, P.; Vohl, M.C. Effects of age, sex, body mass index and apoe genotype on cardiovascular biomarker response to an n-3 polyunsaturated fatty acid supplementation. J. Nutrigenet. Nutrigenomics 2013, 6, 73–82. [Google Scholar] [CrossRef]
- Ouellette, C.; Cormier, H.; Rudkowska, I.; Guenard, F.; Lemieux, S.; Couture, P.; Vohl, M.C. Polymorphisms in genes involved in the triglyceride synthesis pathway and marine omega-3 polyunsaturated fatty acid supplementation modulate plasma triglyceride levels. J. Nutrigenet. Nutrigenomics 2013, 6, 268–280. [Google Scholar] [CrossRef]
- Tremblay, B.L.; Cormier, H.; Rudkowska, I.; Lemieux, S.; Couture, P.; Vohl, M.C. Association between polymorphisms in phospholipase a2 genes and the plasma triglyceride response to an n-3 pufa supplementation: A clinical trial. Lipids Health Dis. 2015, 14, 12. [Google Scholar] [CrossRef] [PubMed]
- Cormier, H.; Rudkowska, I.; Paradis, A.M.; Thifault, E.; Garneau, V.; Lemieux, S.; Couture, P.; Vohl, M.C. Association between polymorphisms in the fatty acid desaturase gene cluster and the plasma triacylglycerol response to an n-3 pufa supplementation. Nutrients 2012, 4, 1026–1041. [Google Scholar] [CrossRef] [PubMed]
- Rudkowska, I.; Guenard, F.; Julien, P.; Couture, P.; Lemieux, S.; Barbier, O.; Calder, P.C.; Minihane, A.M.; Vohl, M.C. Genome-wide association study of the plasma triglyceride response to an n-3 polyunsaturated fatty acid supplementation. J. Lipid Res. 2014, 55, 1245–1253. [Google Scholar] [CrossRef]
- Cooke Bailey, J.N.; Igo, R.P., Jr. Genetic risk scores. Curr. Protoc. Hum. Genet. 2016. [Google Scholar] [CrossRef]
- Ciuculete, D.M.; Bandstein, M.; Benedict, C.; Waeber, G.; Vollenweider, P.; Lind, L.; Schioth, H.B.; Mwinyi, J. A genetic risk score is significantly associated with statin therapy response in the elderly population. Clin. Genet. 2017, 91, 379–385. [Google Scholar] [CrossRef]
- Svendstrup, M.; Allin, K.H.; Sorensen, T.I.A.; Hansen, T.H.; Grarup, N.; Hansen, T.; Vestergaard, H. Genetic risk scores for body fat distribution attenuate weight loss in women during dietary intervention. Int. J. Obes. 2018, 42, 370–375. [Google Scholar] [CrossRef] [PubMed]
- Rudkowska, I.; Paradis, A.M.; Thifault, E.; Julien, P.; Tchernof, A.; Couture, P.; Lemieux, S.; Barbier, O.; Vohl, M.C. Transcriptomic and metabolomic signatures of an n-3 polyunsaturated fatty acids supplementation in a normolipidemic/normocholesterolemic caucasian population. J. Nutr. Biochem. 2013, 24, 54–61. [Google Scholar] [CrossRef]
- Tai, E.S.; Demissie, S.; Cupples, L.A.; Corella, D.; Wilson, P.W.; Schaefer, E.J.; Ordovas, J.M. Association between the ppara l162v polymorphism and plasma lipid levels: The framingham offspring study. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 805–810. [Google Scholar] [CrossRef]
- Warodomwichit, D.; Arnett, D.K.; Kabagambe, E.K.; Tsai, M.Y.; Hixson, J.E.; Straka, R.J.; Province, M.; An, P.; Lai, C.Q.; Borecki, I.; et al. Polyunsaturated fatty acids modulate the effect of tcf7l2 gene variants on postprandial lipemia. J. Nutr. 2009, 139, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Binia, A.; Vargas-Martinez, C.; Ancira-Moreno, M.; Gosoniu, L.M.; Montoliu, I.; Gamez-Valdez, E.; Soria-Contreras, D.C.; Angeles-Quezada, A.; Gonzalez-Alberto, R.; Fernandez, S.; et al. Improvement of cardiometabolic markers after fish oil intervention in young mexican adults and the role of pparalpha l162v and ppargamma2 p12a. J. Nutr. Biochem. 2017, 43, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Maddison, R.; Ni Mhurchu, C.; Jiang, Y.; Vander Hoorn, S.; Rodgers, A.; Lawes, C.M.; Rush, E. International physical activity questionnaire (ipaq) and new zealand physical activity questionnaire (nzpaq): A doubly labelled water validation. Int. J. Behav. Nutr. Phys. Act. 2007, 4, 62. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Avila, M.; Romieu, I.; Parra, S.; Hernandez-Avila, J.; Madrigal, H.; Willett, W. Validity and reproducibility of a food frequency questionnaire to assess dietary intake of women living in mexico city. Salud. Publica. Mex. 1998, 40, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Lawson, L.D.; Hughes, B.G. Absorption of eicosapentaenoic acid and docosahexaenoic acid from fish oil triacylglycerols or fish oil ethyl esters co-ingested with a high-fat meal. Biochem. Biophys. Res. Commun. 1988, 156, 960–963. [Google Scholar] [CrossRef]
- Vallee Marcotte, B.; Cormier, H.; Guenard, F.; Rudkowska, I.; Lemieux, S.; Couture, P.; Vohl, M.C. Novel genetic loci associated with the plasma triglyceride response to an omega-3 fatty acid supplementation. J. Nutrigenet. Nutrigenomics 2016, 9, 1–11. [Google Scholar] [CrossRef]
- Ko, A.; Cantor, R.M.; Weissglas-Volkov, D.; Nikkola, E.; Reddy, P.M.; Sinsheimer, J.S.; Pasaniuc, B.; Brown, R.; Alvarez, M.; Rodriguez, A.; et al. Amerindian-specific regions under positive selection harbour new lipid variants in latinos. Nat. Commun. 2014, 5, 3983. [Google Scholar] [CrossRef] [PubMed]
- Diabetes Genetics Initiative of Broad Institute of Harvard and MIT, Lund University, and Novartis Institutes of BioMedical Research; Saxena, R.; Voight, B.F.; Lyssenko, V.; Burtt, N.P.; de Bakker, P.I.; Chen, H.; Roix, J.J.; Kathiresan, S.; Hirschhorn, J.N.; et al. Genome-wide association analysis identifies loci for type 2 diabetes and triglyceride levels. Science 2007, 316, 1331–1336. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Go, M.J.; Hu, C.; Hong, C.B.; Kim, Y.K.; Lee, J.Y.; Hwang, J.Y.; Oh, J.H.; Kim, D.J.; Kim, N.H.; et al. Large-scale genome-wide association studies in east asians identify new genetic loci influencing metabolic traits. Nat. Genet. 2011, 43, 990–995. [Google Scholar] [CrossRef]
- Rivas-Gomez, B.; Almeda-Valdes, P.; Tussie-Luna, M.T.; Aguilar-Salinas, C.A. Dyslipidemia in mexico, a call for action. Rev. Invest. Clin. 2018, 70, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Solis, A.L.; Datta Banik, S.; Mendez-Gonzalez, R.M. Prevalence of metabolic syndrome in mexico: A systematic review and meta-analysis. Metab. Syndr. Relat. Disord. 2018, 16, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Shamah-Levy, T.; Ruiz-Matus, C.; Rivera-Dommarco, J.; Kuri-Morales, P.; Cuevas-Nasu, L.; Jiménez-Corona, M.E.; Romero-Martínez, M.; Méndez Gómez-Humarán, I.; Gaona-Pineda, E.B.; Gómez-Acosta, L.M.; et al. Encuesta Nacional de Salud y Nutrición de Medio Camino 2016; Resultados Nacionales: Cuernavaca, México; Instituto Nacional de Salud Pública: Cuernavaca, México, 2017. [Google Scholar]
- Ahmad, S.; Mora, S.; Franks, P.W.; Orho-Melander, M.; Ridker, P.M.; Hu, F.B.; Chasman, D.I. Adiposity and genetic factors in relation to triglycerides and triglyceride-rich lipoproteins in the women’s genome health study. Clin. Chem. 2018, 64, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Zubair, N.; Mayer-Davis, E.J.; Mendez, M.A.; Mohlke, K.L.; North, K.E.; Adair, L.S. Genetic risk score and adiposity interact to influence triglyceride levels in a cohort of filipino women. Nutr. Diabetes 2014, 4, e118. [Google Scholar] [CrossRef] [PubMed]
- Buscot, M.J.; Magnussen, C.G.; Juonala, M.; Pitkanen, N.; Lehtimaki, T.; Viikari, J.S.; Kahonen, M.; Hutri-Kahonen, N.; Schork, N.J.; Raitakari, O.T.; et al. The combined effect of common genetic risk variants on circulating lipoproteins is evident in childhood: A longitudinal analysis of the cardiovascular risk in young finns study. PLoS ONE 2016, 11, e0146081. [Google Scholar] [CrossRef] [PubMed]
- Leon-Mimila, P.; Vega-Badillo, J.; Gutierrez-Vidal, R.; Villamil-Ramirez, H.; Villareal-Molina, T.; Larrieta-Carrasco, E.; Lopez-Contreras, B.E.; Kauffer, L.R.; Maldonado-Pintado, D.G.; Mendez-Sanchez, N.; et al. A genetic risk score is associated with hepatic triglyceride content and non-alcoholic steatohepatitis in mexicans with morbid obesity. Exp. Mol. Pathol. 2015, 98, 178–183. [Google Scholar] [CrossRef] [PubMed]
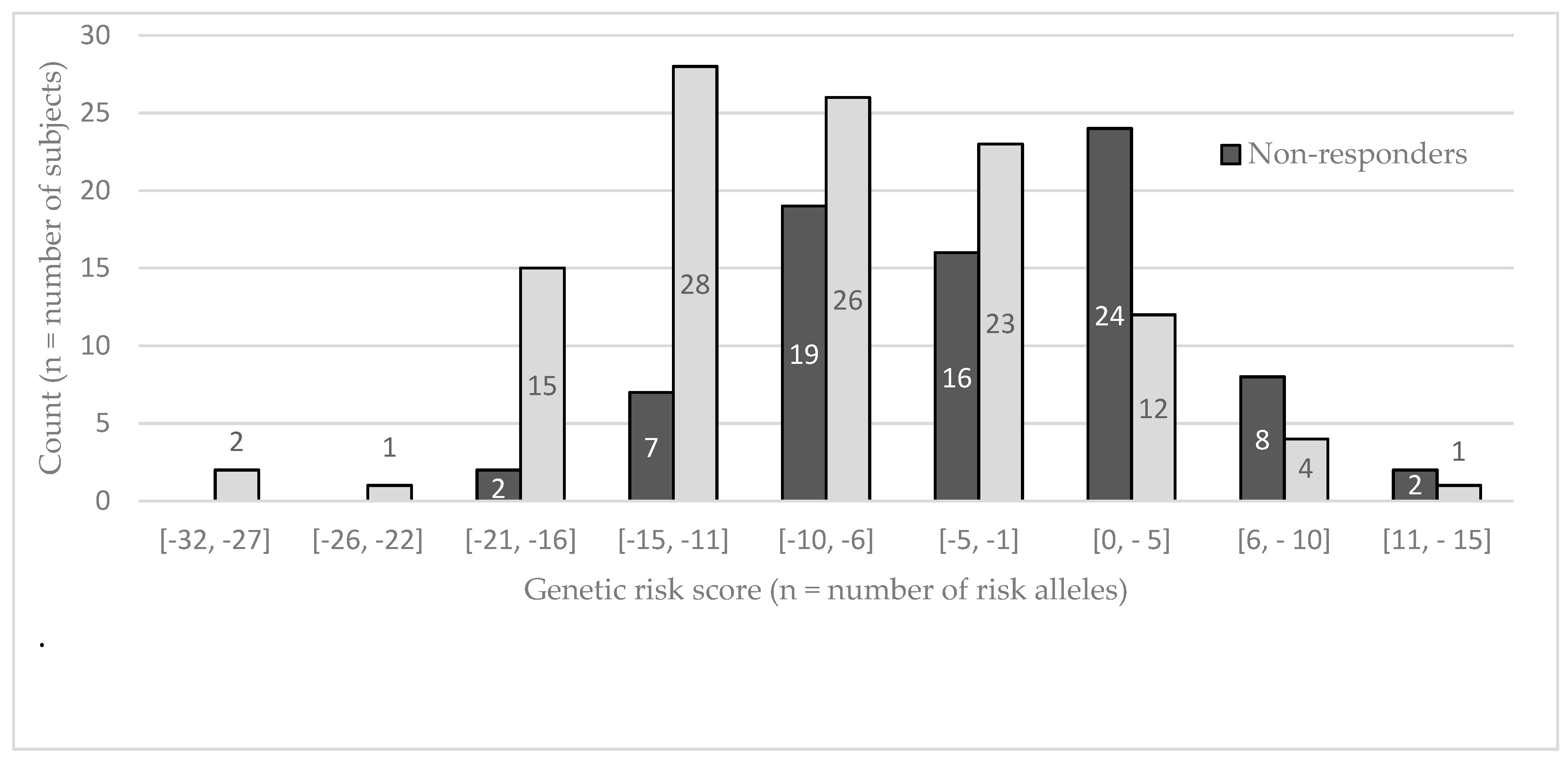

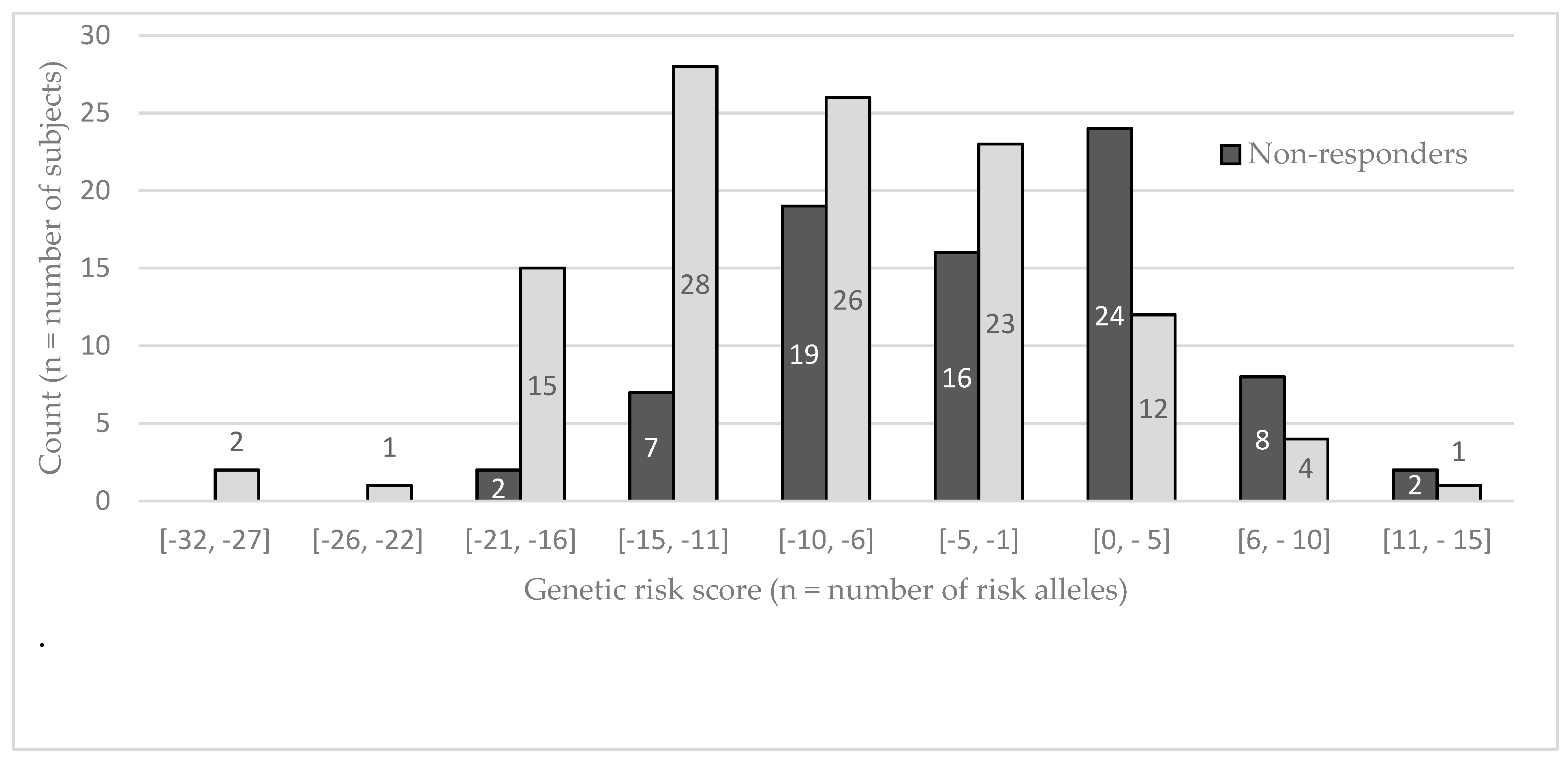{kind=link}
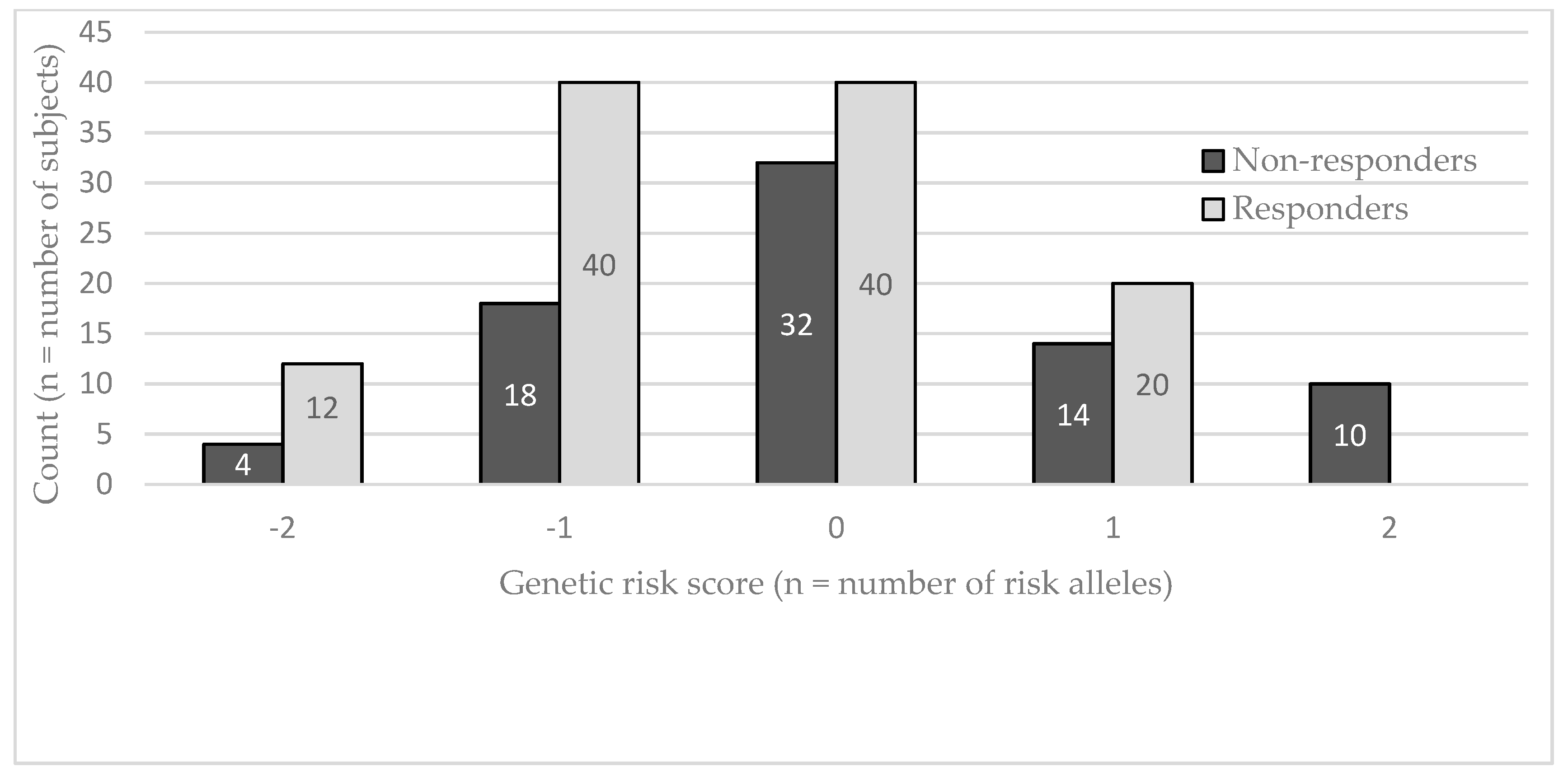
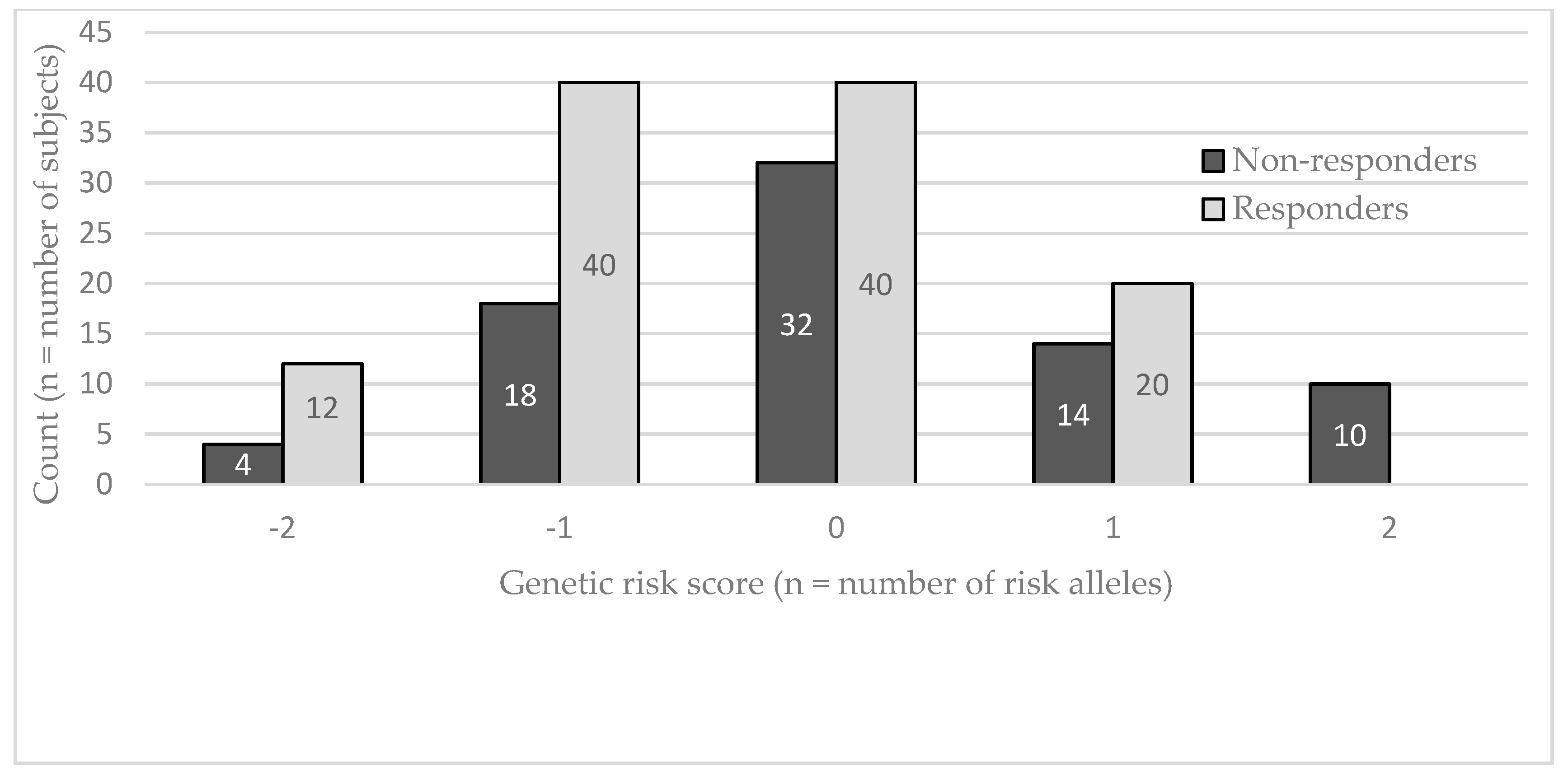{kind=link}
| Characteristics | Responders (n = 113) * | Non-Responders (n = 78) * | p between Responders and Non-Responders | |||||
|---|---|---|---|---|---|---|---|---|
| Baseline | Post-Intervention | pa | Baseline | Post-Intervention | pa | Baseline a | Post-Intervention a | |
| Sex (Male/Female) | 42/71 | - | - | 28/50 | - | - | - | - |
| Age (years) b | 26.1 ± 6.1 | - | - | 27.2 ± 6.4 | - | - | 0.24 | - |
| Weight (kg) b | 64.5 ± 9.7 | 64.6 ± 9.7 | 0.70 | 64.2 ± 10.9 | 64.3±10.6 | 0.27 | 0.81 | 0.85 |
| Height (m) b | 164.6 ± 7.9 | - | - | 164.2 ± 9.1 | - | - | 0.77 | 0.71 |
| Body mass index (kg/m2) b | 23.6 ± 2.7 | 23.8 ± 2.6 | 0.23 | 23.7 ± 2.6 | 23.8 ± 2.6 | 0.053 | 0.84 | 0.99 |
| Triglycerides (mg/dL) b | 110.1 ± 60.7 | 81.4 ± 41.9 | <0.0001 | 80.3 ± 34.4 | 111.9 ± 71.9 | <0.0001 | <0.0001 | 0.001 |
| Gene | SNP | Minor Allele Frequency | Chi-Squared | p Value | |
|---|---|---|---|---|---|
| Caucasian | Mexican | ||||
| IQCJ-SCHIP1 | rs12497650 | 0.32 | 0.27 | 2.73 | 0.10 |
| IQCJ-SCHIP1 | rs4501157 | 0.35 | 0.24 | 12.57 | 0.00039 |
| IQCJ-SCHIP1 | rs13091349 | 0.17 | 0.09 | 9.10 | 0.0026 |
| IQCJ-SCHIP1 | rs2044704 | 0.26 | 0.41 | 21.92 | 2.84 × 10−6 |
| IQCJ-SCHIP1 | rs1962071 | 0.27 | 0.38 | 9.92 | 0.0016 |
| IQCJ-SCHIP1 | rs7634829 | 0.44 | 0.28 | 20.77 | 5.18 × 10−6 |
| IQCJ-SCHIP1 | rs2621294 | 0.38 | 0.23 | 21.57 | 3.41 × 10−6 |
| IQCJ-SCHIP1 | rs6800211 | 0.29 | 0.14 | 24.59 | 7.08 × 10−7 |
| IQCJ-SCHIP1 | rs17782879 | 0.30 | 0.39 | 7.12 | 0.0076 |
| IQCJ-SCHIP1 | rs1868414 | 0.33 | 0.17 | 25.85 | 3.69 × 10−7 |
| IQCJ-SCHIP1 | rs2595260 | 0.25 | 0.52 | 61.21 | 5.12 × 10−15 |
| IQCJ-SCHIP1 | rs6763890 | 0.34 | 0.20 | 18.19 | 2.00 × 10−5 |
| NXPH1 | rs6956210 | 0.24 | 0.13 | 14.02 | 0.00018 |
| NXPH1 | rs2107779 | 0.55 | 0.41 | 16.20 | 5.70 × 10−5 |
| NXPH1 | rs10273195 | 0.20 | 0.24 | 2.32 | 0.13 |
| NXPH1 | rs12216689 | 0.28 | 0.32 | 1.17 | 0.28 |
| NXPH1 | rs6963644 | 0.08 | 0.04 | 3.43 | 0.064 |
| NXPH1 | rs17150341 | 0.30 | 0.16 | 24.51 | 7.39 × 10−7 |
| NXPH1 | rs1013868 | 0.33 | 0.45 | 11.91 | 0.00056 |
| NXPH1 | rs4318981 | 0.36 | 0.36 | 0.03 | 0.873 |
| NXPH1 | rs17153997 | 0.43 | 0.30 | 15.21 | 9.61 × 10−5 |
| NXPH1 | rs7801099 | 0.45 | 0.53 | 5.60 | 0.018 |
| NXPH1 | rs4725120 | 0.46 | 0.47 | 0.16 | 0.69 |
| NXPH1 | rs10238726 | 0.31 | 0.38 | 3.61 | 0.057 |
| NXPH1 | rs1012960 | 0.50 | 0.46 | 1.61 | 0.20 |
| NXPH1 | rs11767429 | 0.30 | 0.34 | 1.25 | 0.26 |
| NXPH1 | rs4333500 | 0.40 | 0.45 | 2.03 | 0.15 |
| NXPH1 | rs7793115 | 0.10 | 0.05 | 6.60 | 0.01 |
| NXPH1 | rs7799856 | 0.43 | 0.39 | 1.10 | 0.30 |
| NXPH1 | rs7806226 | 0.16 | 0.41 | 62.40 | 2.80 × 10−15 |
| NXPH1 | rs13221144 | 0.23 | 0.11 | 22.21 | 2.45 × 10−6 |
| NXPH1 | rs17406479 | 0.19 | 0.31 | 14.86 | 0.00012 |
| NXPH1 | rs10486228 | 0.18 | 0.44 | 61.91 | 3.59 × 10−15 |
| NXPH1 | rs17154569 | 0.18 | 0.08 | 16.90 | 3.95 × 10−5 |
| NXPH1 | rs4141002 | 0.12 | 0.17 | 3.69 | 0.055 |
| NXPH1 | rs7805772 | 0.19 | 0.40 | 41.06 | 1.48 × 10−10 |
| NXPH1 | rs2349780 | 0.38 | 0.49 | 10.00 | 0.0016 |
| NXPH1 | rs2107474 | 0.42 | 0.49 | 4.03 | 0.045 |
| NXPH1 | rs11769942 | 0.37 | 0.43 | 2.98 | 0.084 |
| NXPH1 | rs6952383 | 0.10 | 0.05 | 6.26 | 0.012 |
| NXPH1 | rs6974252 | 0.14 | 0.23 | 11.04 | 0.00090 |
| NXPH1 | rs10265408 | 0.28 | 0.31 | 1.04 | 0.31 |
| NXPH1 | rs2189904 | 0.33 | 0.18 | 24.95 | 5.88 × 10−7 |
| NXPH1 | rs2057862 | 0.41 | 0.53 | 10.99 | 0.00092 |
| PHF17 | rs2217023 | 0.19 | 0.74 | 237.40 | 1.45 × 10−53 |
| PHF17 | rs4975270 | 0.43 | 0.47 | 1.10 | 0.29 |
| PHF17 | rs11722830 | 0.21 | 0.18 | 1.16 | 0.28 |
| PHF17 | rs12505447 | 0.19 | 0.16 | 1.43 | 0.23 |
| PHF17 | rs6534704 | 0.08 | 0.03 | 8.76 | 0.0031 |
| PHF17 | rs13148510 | 0.04 | 0.01 | 8.69 | 0.0032 |
| PHF17 | rs13143771 | 0.28 | 0.32 | 1.54 | 0.21 |
| PHF17 | rs13142964 | 0.07 | 0.05 | 2.43 | 0.12 |
| MYB | rs9321493 | 0.45 | 0.45 | 0.02 | 0.89 |
| MYB | rs11154794 | 0.13 | 0.10 | 1.09 | 0.30 |
| MYB | rs210798 | 0.42 | 0.43 | 0.12 | 0.73 |
| MYB | rs210936 | 0.48 | 0.30 | 27.59 | 1.50 × 10−7 |
| MYB | rs7757388 | 0.16 | 0.05 | 26.67 | 2.42 × 10−7 |
| MYB | rs17639758 | 0.03 | 0.02 | 1.04 | 0.31 |
| MYB | rs1013891 | 0.35 | 0.23 | 12.98 | 0.00031 |
| MYB | rs2179308 | 0.51 | 0.43 | 4.57 | 0.033 |
| IQCJ-SCHIP1 | rs1449009 * | 0.29 | 0.60 | 77.70 | 1.20 × 10−18 |
| IQCJ-SCHIP1 | rs61332355 * | 0.18 | 0.33 | 23.30 | 1.38 × 10−6 |
| IQCJ-SCHIP1 | rs12485627 | 0.40 | 0.32 | 5.07 | 0.024 |
| IQCJ-SCHIP1 | rs2595242 | 0.52 | 0.26 | 53.59 | 2.47 × 10−13 |
| IQCJ-SCHIP1 | rs7639937 | 0.25 | 0.49 | 46.36 | 9.84 × 10−12 |
| IQCJ-SCHIP1 | rs9820807 | 0.16 | 0.07 | 17.52 | 2.84 × 10−5 |
| IQCJ-SCHIP1 | rs1375409 | 0.29 | 0.38 | 7.04 | 0.0080 |
| IQCJ-SCHIP1 | rs1967363 | 0.22 | 0.36 | 18.84 | 1.42 × 10−5 |
| IQCJ-SCHIP1 | rs9824310 | 0.40 | 0.46 | 3.45 | 0.063 |
| IQCJ-SCHIP1 | rs11915303 | 0.27 | 0.33 | 3.41 | 0.065 |
| IQCJ-SCHIP1 | rs9835214 | 0.46 | 0.39 | 3.79 | 0.051 |
| IQCJ-SCHIP1 | rs11921343 | 0.19 | 0.26 | 5.24 | 0.022 |
| IQCJ-SCHIP1 | rs13066560 | 0.16 | 0.08 | 13.52 | 0.00024 |
| IQCJ-SCHIP1 | rs1675497 | 0.29 | 0.32 | 0.61 | 0.44 |
| IQCJ-SCHIP1 | rs9839862 | 0.11 | 0.17 | 5.33 | 0.021 |
| IQCJ-SCHIP1 | rs16829875 | 0.22 | 0.37 | 22.29 | 2.34 × 10−6 |
| IQCJ-SCHIP1 | rs17795566 | 0.36 | 0.23 | 17.17 | 3.42 × 10−5 |
| IQCJ-SCHIP1 | rs9860588 | 0.23 | 0.10 | 21.25 | 4.02 × 10−6 |
| IQCJ-SCHIP1 | rs16830408 | 0.27 | 0.23 | 2.17 | 0.14 |
| IQCJ-SCHIP1 | rs17798579 | 0.17 | 0.16 | 0.19 | 0.66 |
| IQCJ-SCHIP1 | rs2364930 | 0.40 | 0.23 | 25.15 | 5.31 × 10−7 |
| IQCJ-SCHIP1 | rs9865997 | 0.14 | 0.29 | 25.63 | 4.14 × 10−7 |
| IQCJ-SCHIP1 | rs2595241 | 0.26 | 0.58 | 86.73 | 1.25 × 10−20 |
| IQCJ-SCHIP1 | rs7632574 | 0.19 | 0.29 | 10.25 | 0.0014 |
| IQCJ-SCHIP1 | rs2621308 | 0.2589 | 0.5895 | 71.53 | 2.74 × 10−17 |
| SLIT2 | rs2952724 | 0.3511 | 0.4789 | 10.84 | 0.0010 |
| PHF17 | rs1216352 * | 0.3475 | 0.5921 | 38.76 | 4.80 × 10−10 |
| PHF17 | rs1216365 * | 0.6196 | 0.3421 | 49.57 | 1.92 × 10−12 |
| MYB | rs6920829 * | 0.1241 | 0.1032 | 0.7113 | 0.40 |
| NXPH1 | rs6463808 * | 0.1773 | 0.4 | 37.86 | 7.59 × 10−10 |
| NELL1 | rs752088 * | 0.3841 | 0.4579 | 3.563 | 0.059 |
| Number of Participants Excluded a | Number of Participants Included | % of TG Variance Explained by the GRS | p |
|---|---|---|---|
| None | 191 | 11.01 | <0.0001 |
| 35 responders b | 156 | 12.62 | <0.0001 |
| 45 responders; 10 non-responders | 136 | 13.20 | <0.0001 |
| 55 responders; 20 non-responders | 116 | 15.73 | <0.0001 |
| 65 responders; 30 non-responders | 96 | 17.74 | <0.0001 |
| 75 responders; 40 non-responders | 76 | 21.30 | <0.0001 |
| 85 responders; 50 non-responders | 56 | 29.10 | <0.0001 |
| 95 responders; 60 non-responders | 36 | 28.99 | 0.0005 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vallée Marcotte, B.; Guénard, F.; Marquis, J.; Charpagne, A.; Vadillo-Ortega, F.; Tejero, M.E.; Binia, A.; Vohl, M.-C. Genetic Risk Score Predictive of the Plasma Triglyceride Response to an Omega-3 Fatty Acid Supplementation in a Mexican Population. Nutrients 2019, 11, 737. https://doi.org/10.3390/nu11040737
Vallée Marcotte B, Guénard F, Marquis J, Charpagne A, Vadillo-Ortega F, Tejero ME, Binia A, Vohl M-C. Genetic Risk Score Predictive of the Plasma Triglyceride Response to an Omega-3 Fatty Acid Supplementation in a Mexican Population. Nutrients. 2019; 11(4):737. https://doi.org/10.3390/nu11040737
Chicago/Turabian StyleVallée Marcotte, Bastien, Frédéric Guénard, Julien Marquis, Aline Charpagne, Felipe Vadillo-Ortega, Maria Elizabeth Tejero, Aristea Binia, and Marie-Claude Vohl. 2019. "Genetic Risk Score Predictive of the Plasma Triglyceride Response to an Omega-3 Fatty Acid Supplementation in a Mexican Population" Nutrients 11, no. 4: 737. https://doi.org/10.3390/nu11040737
APA StyleVallée Marcotte, B., Guénard, F., Marquis, J., Charpagne, A., Vadillo-Ortega, F., Tejero, M. E., Binia, A., & Vohl, M.-C. (2019). Genetic Risk Score Predictive of the Plasma Triglyceride Response to an Omega-3 Fatty Acid Supplementation in a Mexican Population. Nutrients, 11(4), 737. https://doi.org/10.3390/nu11040737